Insect pests of stored grains in hot climates
The insect species of most importance in the tropics as pests of stored foodgrains, including pulses (legume grains), are listed in Table 8.3. Recognition of these common major pests, at least to the genus level, is not difficult for a trained inspector and there are many useful recognition charts and some excellent keys to specific identification. One of the most recent is contained in a Training Manual "Insects and Arachnids of Tropical Stored Products: their Biology and Identification" produced by the Natural Resources Institute (Haines, C P [ed], 1991). This publication also contains summarised biodata for important species. Another, by Weidner and Rack (1984), may be more useful in francophone countries.
There are many other publications which illustrate the common insect pests of stored products and describe their biology. The familiar details are not all repeated here. Instead, the points which seem of particular importance with regard to pest status are discussed more broadly.
Table 8.3. Important insect pests of tropical stored grains or grain products
| COLEOPTERA: | ANOBIIDAE BOSTRICHIDAE BRUCHIDAE CUCUJIDAE CURCULIONIDAE DERMESTIDAE SILVANIDAE TENEBRIONIDAE |
Lasioderma
serricorne (F) Rhyzopertha dominica (F) Prostephanus truncatus (Horn). Acanthoscelides obtectus (Say) Callosobruchus spp. Zabrotes subfasciatus Boheman Cryptolestes spp. Sitophilus oryzae (L) S. zeamais Motschulsky Trogoderma granarium Everts Dermestes spp. Oryzuephilus surinamensis (L)* Tribolium castaneum (Herbs") |
| LEPIDOPTERA: | GELECHIIDAE PYRALIDAE |
Sitotroga
cerealella (Olivier) Ephestia cautella (Walker) Plodia interpunctella (Hubner) Corcyra cephalonica (Stainton) |
Notes:
*Now established in Africa as well as in the Americas. **Common only on very dry grain; especially in Sahelian North Africa. ***O. mercator (Fauvel) may also occur but is more commonly a pest of oilseeds.
The grain weevils (Curculionidae) are well-known as major primary pests of stored cereal grains. They are able to establish themselves on whole, undamaged grains of maize, sorghum, rice and wheat so long as the grains are not exceptionally dry. However, Sitophilus zeamais is the dominant species on maize while Sitophilus oryzae is dominant on wheat. Neither species is a significant pest of millets and other grains that are too small to permit the full development, within a single grain, of the weevil larva.
The bostrichid beetle Prostephanus truncatus (the Larger Grain Borer) is a highly destructive primary pest of maize, especially maize stored on the cob. This insect is now established in several East and West African countries following recent accidental introductions from its previously more limited indigenous range in meso-America (Dick, 1988; Golob, 1988; McFarlane, 1988a). The Lesser Grain Borer (Rhyzopertha dominica) is more cosmopolitan and is well-known as a destructive pest of most stored cereal grains including millet. It is not generally common on maize.
The bruchid beetles listed in Table 8.3 are the only significant pests of stored pulses. Acanthoscelides obtectus and the less cosmopolitan Zabrotes subfasciatus are generally restricted to dry beans (Phaseolus vulgaris) while Callosobruchus spp are generally restricted to the other legume grains, notably cowpeas and mungbeans (Vigna spp). The anobiid Lasioderma serricorne is sometimes reported as a significant pest of stored beans and other pulses but it is not a primary pest of foodgrains. It is, however, a considerable pest of cereal-based animal feeds, wholemeal flour and high-protein milling offals.
Apart from the one exception noted above (L. serricorne) all of these beetles are primary pests in the sense that they can initiate major damage to the grains and most of them are also able to commence their attack in the field before harvest. The other beetles listed in Table 8.3 are generally regarded as secondary pests which can be of major importance on grains previously damaged either mechanically or by other insects. The dermestid Trogoderma granarium is exceptional in that it can cause major primary damage but rarely occurs as a primary pest except in arid climates, or on very dry grain, where other primary pests are inhibited by the dryness.
Of the moths (Lepidoptera) listed in Table 8.3 only the gelechiid Sitotroga cerealella is able to cause substantial primary damage to the grain kernel. Like the grain weevils it can also infest the grains before harvest and like the Lesser Grain Borer it is a considerable pest of millets as well as all the larger cereal grains.
The other moths listed are warehouse moths, and the rice moth Corcyra cephalonica is included here although it is less commonly abundant on other cereal grains. The larvae of all three species can do substantial damage as secondary pests. They can also attack the whole grain at the site of the embryo, which is typically excised completely, and may thus be of special importance as pests in seed grain stores.
Many other insects may occur quite commonly and sometimes abundantly on stored cereal grains especially when they are underdried or have been heavily infested by the major primary and secondary pests. Some of these are listed in Table 8.4. This list, although quite extensive, is not exhaustive. Most of the species included are capable of doing some damage to the grain and while several are unable to thrive in the absence of mould growth, or are most commonly predators and scavengers rather than grain feeders, their presence would be unacceptable to many consumers. Evident signs of their previous activity, in the form of insect fragments and waste matter, will be equally unacceptable where high quality commands a premium price.
Table 8.4. Insect species (additional to those in Table 8.3) found on underdried stored grain or grain residues
| COLEOPTERA: | ANTHRIBIDAE BOSTRICHIDAE BRUCHIDAE CLERIDAE CRYPTOPHAGIDAE DERMESTIDAE LATHRIDIIDAE MYCETOPHAGIDAE NITIDULIDAE OSTOMIDAE PTINIDAE SILVANIDAE TENEBRIONIDAE |
Araecerus
fasciculatus Degeer Dinoderus spp. Bruchidius spp., Specularius spp. Necrobia rufipes Degeer Thaneroclerus buqueti Lefevre Henoticus californicus (Mann) Cryptophagus spp Attagenus spp., Dermestes spp. Corticaria spp., Lathridius spp. Typhaea stercorea (L) Carpophilus spp. Tenebroides mauritanicus (L) Ptinus spp.*, Trigonogenius spp., Gibbium spp. Cathartus quadricollis (Guerin) Alphitobius spp., Gnatocerus spp. Palorus spp. |
| LEPIDOPIERA: | OECOPHORIDAE | Endrosis sarcitrella (L) |
| PSOCOPTERA: | LIPOSCELIDAE | Liposcelis spp. |
Note: Common only in cool upland tropics.
The psocopteran (psocid) species listed in Table 8.4, which include the familiar 'dust lice', sometimes mistaken for mites (Acarina), have received more attention in recent years than previously; both in the tropics and in temperate countries. In the latter, they have attracted attention as occasional pests of various commodities including skimmed milk powder. In the humid tropics they sometimes occur as troublesome pests of cereal grains, notably milled rice, on which they are sometimes very abundant (Rees, 1990). It has been shown that a psocid population can feed and multiply on damaged or imperfect cereal grains (Shires, 1982). When milled rice is exposed to very dense populations the feeding damage, on some individual kernels, can be massive (McFarlane, 1982). Whether or not such damage occurs sufficiently extensively to constitute an economically significant weight loss in storage practice has yet to be demonstrated. However, it has been shown (V. Pike, personal communication) that measurable weight losses can occur, especially on under-milled rice, and that the protein and lipid content of the infested rice may be reduced. The nuisance-value of these small insects, when they occur as dense, swarming populations in warehouses, is also considerable. Furthermore, surface scarification due to psocid feeding may improve the apparent whiteness of infested milled rice and thus obscure the visible evidence of mould growth and possible contamination by mycotoxins.
Pest status
The status of any particular insect pest may vary between different commodities, different varieties of the same commodity, different climatic regions and agro-industrial systems and between different socio-economic groups. It is affected by the form in which the commodity is stored (Figure 8.2. Pest status as affected by handling and processing.), by the environmental conditions and by consumer attitudes. As with the psocid problem, referred to previously, it may also be affected by the sensitivities of store supervisors and their work-force.
| Key | ||
| S.zm | Sitophilus zeamais | - on maize, sorghum and rice |
| S.o | Sitophilus oryzae | - on wheat, sorghum and rice |
| S.c | Sitotroga cerealella | - on all cereals |
| P.t | Prostephanus truncatus | - on maize |
| R.d | Rhyzopertha dominica | - on all cereals |
| T.c etc | Tribolium castaneum & other secondary beetles | - ditto |
| E.c etc | Ephestia cautella & other warehouse moths | - ditto |
| L.s | Lasioderma serricorne | - on all grains |
| A.o | Acanthoscelides obtectus | - on beans |
| C.spp | Callosobruchus spp. | - on cowpeas, etc. |
| * | Very low status (possibly negligible) | |
| ** | Low status | |
| *** | Low - moderate status | |
| **** | Moderate - high status | |
| ***** | High status | |
Pest status may also vary between biotypes of the same insect species due to differences in the capacity to cause grain damage (McFarlane, 1990) or to adaptations to other foodstuffs. As an extreme example of this, although grain weevils are usually insignificant as pests of stored pulses there are biotypes of S. oryzae that multiply successfully on stored split peas (Holloway, 1986) and on mung beans (C P Haines, personal communication: including field records). Certain strains of S. oryzAe have been noted as having greater flight proclivity than others (Kiritani, 1959). This species, unlike the maize weevil S. zeamais, does not usually fly very readily, although it has wings.
The influence of maize varietal characteristics upon the preharvest infestation of maize cobs by S. zeamais has been much investigated (Floyd and Powell, 1958; Giles and Ashman, 1971; Schulten, 1976). It is clear that the cob sheath, in those cultivars which produce sheathing leaves completely enclosing the entire cob, provides considerable protection against the weevil. Storage of cobs in the sheath, which does not significantly impair the grain drying rate in ventilated cribs, therefore reduces the status of the grain weevil as a pest and will be beneficial where weevils are the main threat (Dick, 1988). Even without the sheath, grains on the cob are considerably less susceptible to weevil attack than the shelled grains. The reasons for this have been clarified recently; by Kossou, Bosque-Perez and Mareck (1992). Unfortunately, maize on the cob, especially without the sheath, is more heavily attacked by the grain moth S. cerealella. The Larger Grain Borer Prostephanus truncatus is also favoured by storage on the cob, with or without the sheath. The impact of this pest in those African countries where it has recently established itself has been dramatic for this reason as well as on account of the more rapid and destructive grain damage caused by the adult borers (Golob, 1988).
The status of this new pest in maize-based farming systems in Africa, traditionally dependent upon cob storage as a modest pest management stratagem, has been such that pragmatic thresholds for control action are commonly exceeded (McFarlane, 1988a). In consequence, demonstrably cost-effective treatments of shelled maize grain with admixed powder formulations of suitable synthetic insecticides have been readily adopted in several regions where they were, formerly, less often used. As a further consequence, it may be that the overall insect control level has been considerably enhanced, since the recommended formulations are generally 'cocktails' of two active ingredients that can give effective control of a broad spectrum of storage insect pests (Golob, 1988).
The storage of sorghum and rough rice (paddy) in the panicle, millet on the head and cowpeas in the pod also serves as a modest form of insect pest management. However, the storage of unthreshed grains, although it may retard the build-up of infestation by some pests, does not prevent it entirely and different insects are affected in different ways. Thus, on sorghum, infestation by grain weevils is usually reduced but the grain moth S. cerealella is likely to be more successful (Giles, 1965; Wongo and Pedersen, 1990). On unthreshed rice and millet, for similar reasons, the grain moth will have increased pest status.
The low status of grain weevils as pests of millets and other small grains, previously mentioned, is due primarily to the limited grain size since the complete larval development, in these species, has to take place within one kernel. The lesser grain borer (R. dominica) and the grain moth (S. cerealella) are not so handicapped because their larvae are able to migrate, if necessary, from one kernel to another. However, the grain moth is a low status pest on large grain bulks and tightly-built large bag-stacks, where the beetles are serious pests, because the moth is unable to move freely amongst close-packed grains to lay its eggs and the first instar larvae are unable to travel more than a few centimetres in search of food.
Most storage insects, especially the important pests, are able to survive and multiply rapidly on well-dried grain. However, grain dried to below 12% mc inhibits the development of most species to some extent and on exceptionally dry grain (<8% mc) the grain weevils, for example, are insignificant pests. The grain borers remain of considerable importance at these low moisture levels and the "khapra" beetle (T. granarium) becomes increasingly important.
This insect assumes major pest status and dominance over almost all other storage insects at the very low moisture contents (down to about 4%) which equate to the extremely low humidities ( < 20% rh) that characterise the most arid climates and, also, the insect ecosystem created by stored malting barley which is usually dried to this very low level and is not uncommonly imported into the tropics.
Factors affecting development and control
Grain moisture content considerably affects pest status but it is not a factor which can be cost-effectively manipulated, in most situations, to achieve sufficient control of insect pests. Cost-effective drying, in common practice, can achieve control of moulds and will lessen the problems of insect infestation; in particular it will greatly reduce the spectrum of pest species. However, it will not prevent significant damage by one or more of the major insect pests.
Insect development and population growth rates are more dramatically affected by temperature and here the developmental limits are more clearly defined and generally applicable. Upper limits for development and survival vary to some extent between species, with the grain borers again more resistant than the grain weevils, but temperatures above 45°C are eventually fatal to all storage insects. At 50°C most species will die quite quickly, within a matter of hours, and complete disinfestation of wheat grain can be achieved rapidly, economically and without damage to the grain by very short exposures to air heated to 60°C (Evans, 1987b).
Rapid insect development occurs within a fairly narrow range of 5-10 degrees around the optimal temperature which, for most storage insects, is in the region of 30°C. At temperatures nearer to 20°C development proceeds more slowly and population growth may be considerably reduced. At 17°C or less it is relatively negligible and pest status is consequently greatly reduced. However, even at 15°C some species are able to continue feeding, to some extent, so that grain damage may very slowly increase. Insect populations will certainly not be eliminated at these temperatures and, while grain may often be held safely in cool storage, any eventual transfer to warmer conditions will bring about a resurgence of the suppressed infestation. Even in cold storage (at 6-9 °C) some, at least, of the important insect pests of stored grain can survive longer than one year (Wohlgemuth, 1989).
Insects require oxygen for respiration. Living grains, when sufficiently dry (12-13% me), are dormant and respire very little. Grain properties, including viability, are virtually unaffected in cool conditions by protracted hermetic storage. Insects, however, will use up the oxygen and eventually die. The traditional concept of sealed (hermetic) storage as a means of controlling insect infestation depends upon this. Most storage insects will die when the oxygen in the storage atmosphere is reduced, by the insects' respiration, to 2% (Hyde et al., 1973). With light infestation the process may take 6-8 weeks but, if airtight conditions are maintained, the infestation will be controlled and probably eliminated before serious damage is done. There is new evidence (Donahaye, 1990) that insects may adapt to low oxygen tensions and evolve strains with considerable resistance to sub-optimal levels even down to about 1%. However, it seems very improbable that any storage insect would multiply rapidly in such conditions.
Physical disturbance of grain, by turning it from one elevator bin to another, can reduce live grain weevil infestation to a considerable extent and thus retard its further development (Joffe, 1963). A more complete kill of all insect life stages can be achieved, by mechanical high-speed impact, in the entoleters included in the processing line of many grain mills. In small-scale storage it may be less easy to achieve the degree of disturbance necessary for effective control of grain weevil and other cereal grain insect pests, although very small quantities of grain held in small pots or gourds could be shaken sufficiently violently for this purpose. Bruchid pests of stored pulses may be particularly susceptible to control by physical disturbance. In the bean bruchid A. obtectus this may be because the hatched first instar larvae require an approximately 24 hr period to penetrate the bean testa (Quentin et al., 1991). These workers have shown that twice-daily 'tumbling' of small lots of stored beans, in partly-filled cylindrical containers, reduced bean bruchid populations by 97%. Daily sieving on a fair-sized coarse mesh can achieve similar results (M N Silim; private communication). However, the practicability of these proposed techniques, for routine use by farmers or traders in developing countries, is unproven.
Insect behaviour patterns may affect pest status and pest control. Examples have been given of the ways in which adult oviposition behaviour and larval feeding behaviour can affect pest status in the grain moth and pest control in bruchids. The development of infestations (pest population development) may also be affected by, for example, the diapause habit which characterizes several storage insects including, most notably, the khapra beetle T. granarium. Diapause may postpone population development, usually in unfavourable conditions, and it may also impair the effectiveness of control measures; including fumigation and the use of contact insecticides as surface sprays for 'clean-up' treatments in empty storages. Locomotory avoidance behaviour, especially in the flour beetle T. castaneum (Wildey, 1987), is also of considerable interest.
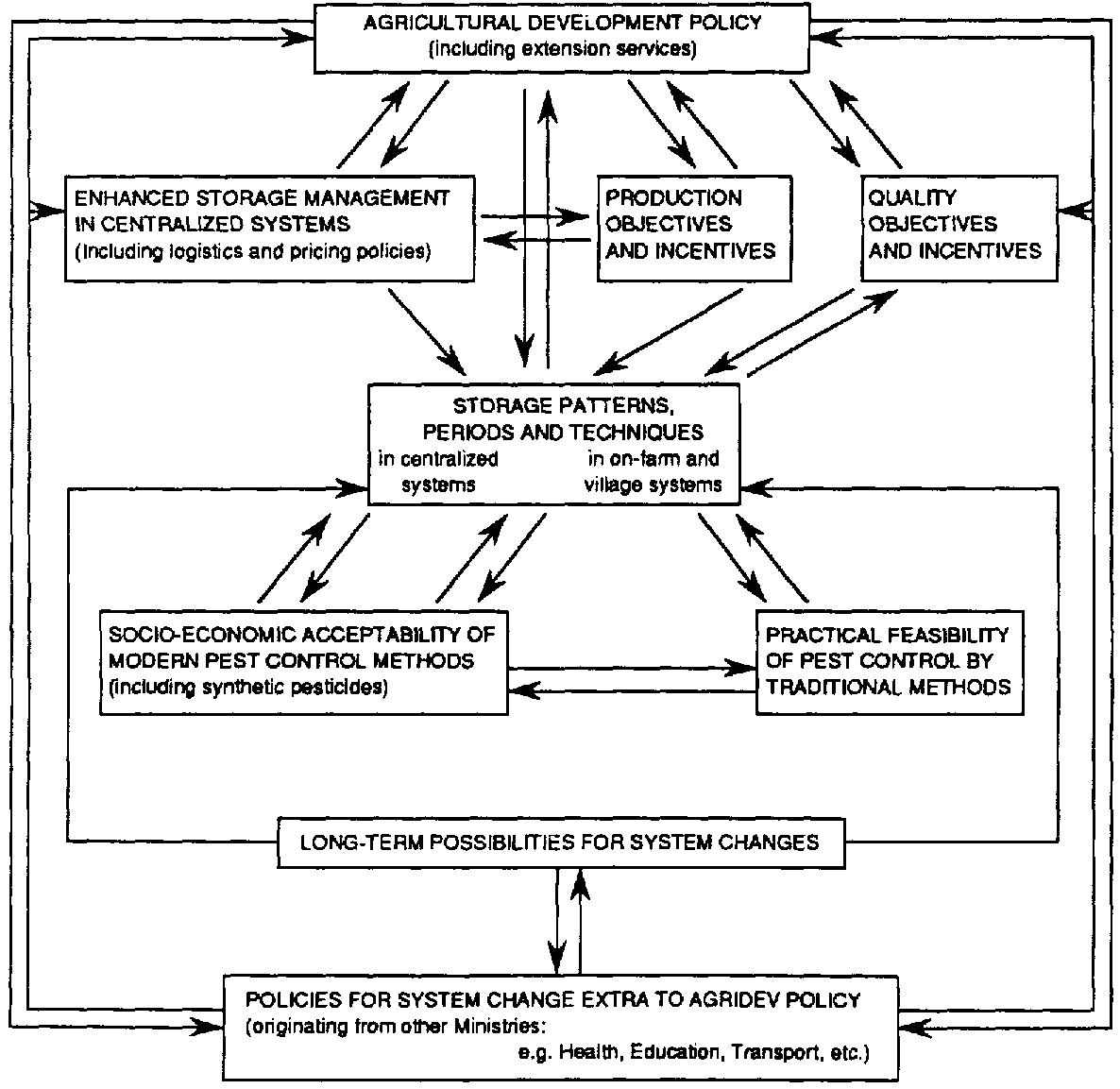
Storage management, which may be described as the science of cost-effective storage organisation (McFarlane, 1988b), greatly influences pest development and control. It encompasses decisions upon the location of stores, storage periods and the quality control objectives for stored commodities. All of these have substantial implications for pest management and are components of the complex interactive network of factors affecting loss reductions in grain storage (Figure 8.3. Factor interactions and key issues for pest management and loss reduction in grain storage.).
Figure 8.3 draws attention also to socio-economic factors affecting, in particular, the acceptability of control measures. Modern techniques, especially those involving the application of synthetic insecticides to stored grain, are especially prone to consumer sensitivity. However, many traditional techniques face comparable problems. The use of wood ash and other supposedly non-toxic 'natural' grain protectants may not be acceptable in all circumstances, nor even to all those people at the small farm level in developing countries who are often supposed to prefer such treatments.
The insect resistance problem
The development by pests of acquired resistance or increased tolerance to pesticides is now a well-known pest management problem. It is coming to be recognised as not so much a remarkable phenomenon as an almost inevitable natural consequence of pesticide use. There is little doubt that much of the existing problem, which in recent decades has come to affect many species of storage insects and a wide range of insecticides (Champ and Dyte, 1976), stems from careless use. Resistance to phosphine fumigants in particular (Taylor, 1991) is almost certainly due in part to the not infrequent use of grossly inefficient application techniques.
There is general agreement that the rate of resistance development, in any particular pest species and to any particular pesticide, is to some extent susceptible to control (i.e. management). Possibilities for the containment of resistance include sustained improvements in application techniques, where acceptable pesticide dosage rates remain effective, and the adoption of alternative pesticides or other control measures where the degree of resistance to a particular pesticide prohibits its use. Continued monitoring of resistance, in field populations, is also necessary.
Concepts of reducing resistance by genetic control (Wool, 1975) have progressed to some extent (Wool et al., 1992) and are not implausible. However, their practicality and cost effectiveness in stored-grains pest control remain undemonstrated. Moreover, it has been shown (McFarlane, 1990) that some resistant strains of several storage insects have a reduced capacity to cause grain damage, through an impaired population growth rate. There is no reason to suppose that all resistant biotypes will be less damaging than their susceptible counterparts but the monitoring of this capacity, in field populations, is feasible and seems worthwhile. Conversely, it would seem illogical to foster a reversion to susceptibility in cases where the resistant biotype has a substantially reduced capacity to cause damage and, therefore, a considerably reduced pest status.
The possibility that insects may develop resistance to other control measures has been noted already with regard to reports of increased insect tolerance to low oxygen tensions in controlled atmosphere storage (Donahaye, 1990). As in the development of pesticide resistance, considerable genetic flexibility is to be expected. However, one may also expect that insect adaptability to massive constraints on fundamental biotic requirements, such as aerobic conditions, will be more limited than their ability to exploit the more natural genetic potential for metabolising extraneous toxicants.
{kind=link}
{kind=link}