![]()
![]()
![]()
K.R. Barker
Introduction
Components of integrated pest management
Nematode management, strategies and tactics
Integration of strategies/tactics
Conclusions and outlook
References
Plant-parasitic nematodes must be addressed in crop production and integrated pest management (IPM) systems if agriculture is to meet the world demands for increasing food and fibre production. On a worldwide basis, annual crop losses due to nematode damage have been estimated to average 12.3 percent (Sasser and Freckman, 1987), amounting to some US$77 million annually. The data available indicate that similar losses occur in the Near East (Maqbool, Hashmi and Ghaffar, 1988; Saxena, Sikora and Srivastava, 1988). For example, estimated losses for vegetable crops due to nematode-related disease complexes in Egypt amounted to some 15 percent in 1986, with losses for field crops ranging from 5 to 20 percent (Eissa, 1988). Nevertheless, the subtle nematode symptoms and signs are often confused with nutrient deficiencies and other maladies, resulting in nematodes being overlooked by agricultural scientists as well as growers.
The concept of combining compatible tactics for controlling nematodes predates that of IPM. In 1889, Atkinson discussed a range of nematode-management tactics that is surprisingly similar to those available today. Tactics discussed for controlling root-knot nematodes include: sterilization of the soil by starvation (including the use of non-host plants); the potential of trap crops; composts; nematicides; and soil amendments such as hardwood ash and potash. The early work of Cobb (1918) on sampling nematode communities provided a basis for the development of improved tactics and strategies essential for integrated pest management (Barker, 1985a). The following statement was offered by Tyler in 1933:
"A well-planned combination of practices will go much further for control of nematodes than any of the recommended treatments alone". Tyler also stated that the presence of nematodes (root-knot) in any soil can best be determined by examining the roots of susceptible plants which have been growing for at least three weeks in warm, moist conditions. For attempted eradication of root-knot nematodes, the author suggested the following:
1. Burning of residues two or three times if possible, each preceded by digging or ploughing.
2. Dry fallow with frequent ploughing.
3. One or two well-irrigated trap crops, completely destroyed two or three weeks after sprouting.
4. Moist fallow during warm weather, without weeds.
5. Resistant crops in rotation, kept free of weeds.
6. Repetition of steps 4 and 5.
Tyler also emphasized that root-knot nematodes enhance the susceptibility of crops to other diseases such as cotton wilt, black shank of tobacco and Rhizoctonia disease of groundnuts. Thus, early nematologists provided an exceptionally strong conceptual framework for the development of advanced integrated management systems.
The specific concept of IPM is a relatively recent development in pest control (Bird, 1987). The 1962 publication of Silent Spring by Rachel Carlson and the subsequent banning of DDT awakened United States agriculturists, especially entomologists, to the risks of pesticides and provided the impetus for the development of IPM (Bird, 1987). IPM consists of the development, use and deployment of pest-control strategies and tactics that result in favourable socio-economic and environmental consequences. Theoretically, IPM is a holistic systems approach to limit pest damage to tolerable levels through a combination of tactics and techniques, including parasites and predators, host resistance, cultural practices, environmental modification, and pesticides where appropriate (Bird, 1987). Unfortunately, the interest and progress of IPM programmes has not been balanced among the pest disciplines with the major emphasis often being on insects. Still, a number of national and international IPM programmes have resulted in many research and extension programmes becoming more interdisciplinary (National Academy of Sciences, 1991, 1992b).
The relative youth of the science of nematology and the availability of effective, relatively inexpensive nematicides hampered the development of integrated management systems in this discipline. During the last two decades, however, much progress has been made in characterizing damage functions and thresholds for major nematode-crop combinations, the study of general population dynamics and simulation models for key nematodes, and the development of improved management tactics (Barker, Schmitt and Imbriani, 1985; Duncan, 1991; Ferris and Noling, 1987). In addition, considerable progress has been made in elucidating the roles of nematodes in disease/pest complexes, and their interactions with symbionts.
IPM has been divided into a number of components in an operational framework including:
· biological monitoring;
· environmental monitoring;
· the decision-maker;
· the decision-support system;
· the decision;
· procedure implementation;
· the system (Fig. 6; Bird, 1987).
Biological monitoring
Dispersal patterns of plant-parasitic nematodes over space and time must be considered in characterizing populations for IPM purposes (Bridge, 1988; Duncan, 1991; Ferris and Noling, 1987). Contagious or spotty horizontal dispersal patterns in the field (Fig. 7) pose difficulties in sampling, experimentation and management. Vertical dispersal patterns also vary greatly, but these obligate parasites are generally closely associated with host roots (Barker, 1985a).
FIGURE 6: Components of integrated pest management
Source: Bird (1987 and personal communication)
Major factors that influence the seasonal fluctuations of nematode populations include: their biology (especially reproductive and survival characteristics); environmental parameters; and, especially, management practices. The temporal population dynamics of Meloidogyne incognita are typical of many nematodes. This pathogen increases to very high population densities during the growing season and then declines very abruptly after harvest. This decline, however, may be greatly affected by management practices (Fig. 8). Note, in Fig. 8, the striking decline of M. incognita numbers in plots in which residual roots were destroyed by repeated discing, in contrast to the relatively high survival in the absence of ploughing. Frequently, these declines are so severe that the population levels may become non-detectable by routine sampling.
Advice for nematode IPM programmes may be based on qualitative and/or quantitative information. Today, many management decisions are based on qualitative or, at best, semi-quantitative population thresholds. This problem is related to the low level of precision in sampling schemes, limited information on damage functions, and the effects of crop cultivar and environmental parameters on nematode/host interactions. Despite these limitations, nematode monitoring can offer much to IPM programmes for growers (Barker and Imbriani, 1984; Duncan, 1991; Schmitt et al., 1990). For example, the detection of small numbers of a highly damaging species, such as Meloidogyne arenaria on groundnuts, may require a shift to another crop or a change in management tactics. Large numbers of a cyst nematode such as Heterodera zeae may require long-term rotations or a combination of management tactics.
As suggested by Tyler in 1933, bioassays utilizing suitable host plants have much to offer in managing certain nematodes. This approach has been used successfully for Heterodera avenae on wheat in Australia (Brown, 1987). Critically timed gall ratings of crop roots, and/or North Carolina differential host plants, are very useful for identifying species and host races of Meloidogyne (Hartman and Sasser, 1985). This approach to monitoring nematode populations can be adapted readily for grower use (Duncan, 1991). Carefully and properly timed compilations of root-gall and root-necrosis indices also are useful as a basis for obtaining yield-loss estimates from these pathogens (Barker, Schmitt and Imbriani, 1985).
Much progress is being made in the development of new technologies for reliable nematode diagnostics and identification which are critical to IPM. Biochemical techniques offer promise to nematode identification and possibly population assessment (Duncan, 1991; Piotte et al., 1992; Schots, Gommers and Egberts, 1992). The topic of disease diagnosis and nematode identification is addressed by Dr Siddiqi on page 21.
Environmental monitoring
Nematode population dynamics and associated crop responses are often related to environmental parameters (Oostenbrink, 1972; Barker, Schmitt and Imbriani, 1985). Thus, such information is critical to effective IPM nematode programmes. In addition to enhancing the predictability of crop responses to given nematode species, environmental data are also helpful in achieving optimum efficacy of a number of management tactics. For example, data on expected soil temperatures should prove useful for maximizing the efficiency of solarization, bare fallow, or combinations with tillage and/or residual root destruction.
Decision-making
The focal point of IPM programmes should be on the decision-maker (Bird, 1987). Thus, the decision-maker should have an understanding of nematode biology, the system to be managed and possible effects of IPM practices on the environment. Although only recently introduced to plant pathology and nematology, expert systems have much potential in this regard (Bird, 1987; Travis and Latin, 1991). For an IPM system to be successful, it must have a decision-support system (Bird, 1987). The minimum support should include innovative research programmes and effective information delivery/transfer. Support systems must address related socio-economic issues as well as nematode damage functions/thresholds, sampling, management options, education and grower inputs. Computer communications and simulation models are becoming increasingly important in IPM and nematology (Bird, 1987; Ferris and Noling, 1987; Travis and Latin, 1991).
Management decisions are based on assessment of the problem, related economics, and availability of a range of management strategies and tactics. (Factors affecting the efficacy of given tactics will be discussed with the respective options.) In management decisions, it is important that compatible tactics be selected and that they be utilized in a proper, effective application. The key topic of nematode-damage functions and thresholds is discussed by Dr Trudgill on page 45. Where damage thresholds and related information are unavailable, nematode-hazard indices (relative probability of moderate to severe damage) might be considered (Barker and Imbriani, 1984).
Reassessment of the system through biological and environmental monitoring is the final step in IPM (Bird, 1987). For nematodes, this should include follow-up population assessments to determine the effectiveness of the management tactics deployed. Long-term monitoring and assessments of given cropping systems should facilitate the detection of shifts in nematode communities, including new species, races, or resistance-attacking biotypes.
Overall IPM constraints
The general constraints to developing effective IPM programmes vary with each country or region. Major overall constraints common to most countries concern technology development and transfer, institutional resources and related training, environmental factors, and key economic and socio-cultural components (National Academy of Sciences, 1992b). For example, growers are often excluded in the early phases of IPM development, yet they may have much to offer and their support is essential for a successful programme.
This list of constraints implies that IPM requires interdisciplinary research and extension efforts. This truly integrated approach to IPM has been achieved in too few instances to date. Nevertheless this issue is central to effective pest management because of the many pest interactions (Alston et al., 1991; Sasser, 1989; Wheelers al., 1992) and pesticide interactions (Beane and Perry, 1990; Schmitt and Nelson, 1987).
The terms strategy and tactics have been used interchangeably in some instances in nematology. In this discussion, Vanderplank's (1963) usage of these concepts will be followed, that is: strategy refers to the overall plan for pathogen (or nematode) management, and tactics denote the specific tools or weapons utilized to carry out the plans. Thus, the general strategies for managing nematodes include:
1. Exclusion (quarantine).
2. Reduction of initial inoculum density.
3. Suppression of nematode reproduction.
4. Restriction of damage to the current crop.
Some researchers combine strategies 2 and 3 under population density management (Bird, 1987). The specific tactics and relative efficacy of the respective strategies are outlined in Table 5.
Currently, most nematode management programmes focus on tactics involving the reduction of the initial nematode population density and/or the suppression of their reproduction during the season. Most nematode management tactics also have the inherent feature of limiting damage associated with the target crop. In contrast, tolerant crop cultivars may be damaged little economically while supporting relatively high nematode reproduction. With the diminishing availability of a single strategy or tactic for nematode management, a combination of two or more compatible tactics in an integrated system is becoming more critical than in recent years when nematicide usage was the primary measure for nematode control.
Exclusion or avoidance
Fortunately nematodes have a number of characteristics that limit their long-distance dispersal, including their restricted active movement, obligate parasitism, narrow host ranges of many species, sharp population declines in the absence of hosts, and survival depending on environmental conditions and crop management practices. Thus, long-distance dispersal of these parasites is largely passive and by chance. Key means of dissemination include movement of soil on equipment and plant parts, crop transplants, water, animals and contaminated containers such as burlap bags. Measures for avoiding dissemination and establishment of new nematode problems should be a component of national or regional nematode IPM programmes.
TABLE 5
Relative efficacy of nematode management strategies and tactics1
|
Strategy2 |
Tactic Relative efficacy3 |
||
|
1. Exclusion/avoidance |
Quarantine |
M,H |
|
|
2. Reduce initial population density |
a. Eliminate established foci |
L,M |
|
|
|
b. Cultural |
||
|
|
· use of clean planting stock |
H |
|
|
· crop rotation |
L-H |
||
|
· inter- and intracropping |
L,M |
||
|
· cover/trap crops |
L,M |
||
|
· soil amendments |
L,M |
||
|
· fallow |
L-H |
||
|
· time of planting/harvesting |
L,M |
||
|
· farm hygiene and general culture |
M,H |
||
|
c. Vertical resistance |
H |
||
|
d. Chemical nematicides4 |
|
||
|
|
· fumigants |
M,H |
|
|
· non-fumigants |
L-H |
||
|
e. Biological |
|
||
|
|
· natural |
L-H |
|
|
· introduced, including enineered |
L-H |
||
|
f. Physical |
|
||
|
|
· tillage, including residual root destruction |
L-H |
|
|
· heat solarization |
L,M |
||
|
· flooding |
L,M |
||
|
3. Suppress nematode reproduction |
a. Resistance horizontal/ quantitative |
L,M L,M |
|
|
4. Restrict current crop damage |
a. Tolerant cultivars |
L-H |
|
1 In part after Oostenbrink, 1972 and Roberts, 1993.
2 Strategies No. 2 and 3 after Vanderplank, 1963.
3 Low (L); moderate (M); high (H).
4 Chemical nematicides often result in high carryover population densities.
Government-regulated quarantines have had varying levels of success in different countries (Barker, 1985b; O'Bannon and Esser, 1987; Taylor, 1986). For example, the quarantines for Globodera rostochiensis and Heterodera zeae have been success stories to date in the United States (Barker, 19850). In contrast, the quarantine for the soybean-cyst nematode, Heterodera glycines, failed in that country. This failure may have been a result of the nematodes being more widely disseminated than initially realized as well as having several effective means of dispersal. Internal quarantines developed by the European and Mediterranean Plant Protection Organization have focused on Aphelenchoides besseyi for rice and strawberry, Ditylenchus destructor, G. rostochiensis and G. pallida on potato, Radopholus citrophilus on citrus, and Xiphinema americanum on a range of crops (EPPO, 1982). More recently, Bursaphelenchus xylophilis has been a focal point concerning the importation of wood products in a number of countries. Other nematodes that have limited distribution and are threats to certain crops include Meloidogyne chitwoodi and M. nataliei (Barker, 1985b). Further spread of these and other highly aggressive nematodes is undoubtedly a concern in most Near East countries.
At a more practical level, certified plant material, nematode-free planting stock and clean farm equipment should be standard IPM practices. This issue is particularly important for vegetatively propagated crops such as banana, potato, and crops for which seedlings are transplanted. In vitro propagation of banana has proved to be an effective means of providing clean planting stock and should reduce the need for nematicides (Sarah, 1989).
In addition, a number of nematodes are disseminated with crop seeds, including Anguina tritici on wheat and Ditylenchus dipsaci on alfalfa and a number of other crops.
Reduction of initial nematode population densities
Six general tactic categories may be placed under this strategy: crop eradication; rotation; vertical resistance; chemical nematicides; biological control; and physical treatments to reduce nematode population densities. This strategy includes some of the oldest and most recently developed tactics that have been used in carefully developed IPM systems.
Eradication or containment. Although eradication is a longstanding tactic for nematode management, it is very difficult to achieve success for field infestations. Fumigation or heat treatments, for example, may eliminate all nematodes in upper soil layers, but those occurring at depths of 0.3 to 1.0 m below the soil surface will generally escape and eventually move back into the plough layer. In contrast, nematode infestations in confined environments, including greenhouses and container-propagated plants, can be eliminated through standard sanitation procedures.
Cultural practices. A wide range of cultural practices have varying levels of efficacy in nematode management. These include clean planting stock, crop rotation, inter- and intracropping, cover/trap crops, soil amendments, fallow, time of planting/harvesting, and general farm hygiene and culture. Such non-chemical tactics are especially important for low-value crops (Brown, 1987).
Crop rotation. This is one of the oldest and most effective tactics for managing plant-parasitic nematodes (Nusbaum and Ferris, 1973). The goal of rotation is to bring about a striking population decline of the target nematodes that will facilitate the subsequent crop to grow and produce an acceptable yield. Crop rotation and cropping systems are similar concepts which are sometimes confused. Crop rotation is the fixed yearly sequence and spatial arrangement of crops, or the alternation of crops and fallow, on a given land area (Nusbaum and Ferris, 1973). Alternate crops in rotation may be planted or natural. In contrast, a cropping system is the sequence of growing crops and the required technologies for their production. Thus, cropping systems cover all kinds of crop sequences, including monoculture, whereas crop rotation indicates an inflexible cycle or a fixed sequence of crops (Nusbaum and Ferris, 1973).
The study of cropping systems should include quantitative analysis of the relationships among crops and pests, and general management tactics which may be deployed in that system (Noe 1986; Noe, Sasser and Imbriani, 1991). The crop system may be spatial or temporal. Thus nematode populations may respond to individual crops as well as the arrangement of the crops over space and time (Table 6). To maximize the efficacy of rotation and cropping systems in nematode management, the short- and long-term effects of cropping systems, spacing and associated interactions with biotic and abiotic environmental factors on crop yields must be better understood. Research addressing these issues should include potential effects of the general environment (Noe and Sikora, 1990), associated weeds and other plant pests in cropping systems and development of appropriate crop rotations.
TABLE 6
Types of crops and their arrangement in time and space evaluated with reference to potential development of pest problems. Some effects are high in pest potential. some intermediate and some low1
|
High pest potential |
Low pest potential |
|
General types of crops and associated pest problems | |
|
Large pest complex ® |
Small pest complex |
|
Susceptible® Tolerant cultivar ® |
Resistant pure line® Resistant multigenic |
|
Annual® |
Perennial |
|
Long-maturing® |
Short-maturing |
|
Crop arrangement in time | |
|
Monoculture® |
Crop species rotation |
|
Continuous planting® |
Discontinuous planting |
|
Asynchronous planting® |
Synchronous planting |
|
Season favourable to pest® |
Season unfavourable to pest |
|
Crop arrangement in space | |
|
Sole cropping® Row or strip® intercropping |
Mixed intercropping |
|
High plant density® |
Low plant density |
|
Large field® |
Small field |
|
Large host crop area® |
Small host crop area |
|
Host fields aggregated® |
Host fields scattered |
1 Partially after Raymundo (1985) and Trivedi and Barker, 1986.
Nusbaum and Ferris (1973) gave two key principles for crop rotation: the reduction of initial nematode population levels necessary to permit the subsequent crop to complete its growth before severe attack; and the preservation of competitive, antagonistic and/or predacious nematodes and other organisms at population densities that allow effective buffering of the pathogenic species. Shifts in nematode populations and communities may be characterized numerically through the application of the concepts that define host status and by various population models. For example, good hosts have a high equilibrium density (E) and maximum rates of reproduction (a) (Seinhorst, 1970; Nusbaum and Ferris, 1973). In contrast, these parameters would be low for poor hosts or non-hosts. The relative host sensitivity is defined or indicated by the tolerance limit (T) (Seinhorst, 1970). The utilization of these quantitative concepts for target nematode species for a primary host and associated crops is necessary for the development of effective rotation systems.
Much knowledge is essential for quantifying nematode-host interactions and the development of specific recommendations for crop-rotation systems. Such data must include nematode species and race or biotype, their host ranges, host efficiency and susceptibility of crops to be included in the rotation, associated weeds, general population dynamics, and information on the relationships between population densities and crop loss (Nusbaum and Ferris, 1973). Other critical information includes the effects of abiotic environmental parameters on nematode population dynamics and associated disease complexes and possible pesticide interactions.
The level of success in nematode management through crop rotation may vary with the year, target nematodes and associated plant pathogens, location, weed hosts present, and the nature and length of the rotation (Trivedi and Barker, 1986). Publications that give host ranges, including weeds, for various nematodes are available (Goodey, Franklin and Hooper, 1965; USDA, 1960). Details on the host ranges of given nematodes are beyond the scope of this discussion. Nevertheless, the non-hosts and poor hosts tabulated for given nematodes in Table 7 may serve as a starting point for developing effective crop rotations for IPM purposes.
TABLE 7
List of selected non-host plants useful in crop rotation1
|
Nematode species |
Non-hosts or poor hosts2 |
|
Belonolaimus longicaudatus |
Crotalaria spp., Crotalaria spectabilis hairy indigo, marigold, tobacco |
|
B. gracilis |
Crotalaria spp., tobacco, watermelon |
|
Dolichodorus heterocephalus |
Crotalaria spectabilis |
|
Helicotylenchus dihystera |
Alfalfa, maize, fescue2 |
|
Heterodera glycines |
Maize, cotton, cowpea, potato, small grains, grains, tobacco, most vegetables |
|
H. schachtii |
Alfalfa, bean, clover, maize, Hesperis matronalis, onion |
|
H. zeae3 |
Wide range of crops |
|
Globodera rostochiensis |
Maize, green beans, red clover |
|
Hoplolaimus indicus |
Cabbage, chili, eggplant |
|
Meloidogyne javanica |
Andropogon, Crotalaria spp., cotton, groudnut, sorghum, velvet bean |
|
M. hapla |
Maize, cotton, grasses, lettuce, onion, radish |
|
M. incognita |
Fescue, orchard grass |
|
Meloidogyne spp. |
Crotalaria spectabilis, Indigolera hirsuta, millet, oats, wheat2 |
|
Paratrichodorus minor |
Maize,2 Crotalaria spectabilis |
|
Pratylenchus leiocephalus |
Groundnut |
|
P. penetrans |
Alfalfa,2 beet, fescue, marigold, oats, Sudan grass, rye |
|
Pratylenchus spp. |
Lettuce, onion, radish |
|
Radopholus similis |
Crotalaria spectabilis |
|
Tylenchorhynchus mirzal |
Wheat |
|
T. brassicae |
Potato, tomato |
|
Xiphinema americanum |
Alfalfa,2 maize,2 fescue,2 tobacco |
1 See Trivedi and Barker (19S6) for specific references per nematode/crop combination.
2 Some populations of respective nematode species will reproduce rapidly on these crop plants.
3 Host range needs further study.
Non-host crops have been used for developing effective rotation programmes for nematodes such as Meloidogyne spp., Heterodera spp., Ditylenchus spp., Belonolaimus spp. and Pratylenchus spp. (Trivedi and Barker, 1986). Rotation systems that involve two to four years of non-host crops generally give excellent results managing these nematodes. For cropping systems in the Near East, this may involve several crop sequences of non-hosts between the production of a susceptible host during such a two- to four-year period. For example, groundnuts, cotton, potato and a range of vegetables or perennial crops, such as alfalfa, may be grown between crops of maize as a means of limiting damage to the latter by H. zeae. Effective crop rotations for Meloidogyne spp., with their broad host range, are more difficult to develop. However, a number of crops important in the Near East may be considered as a nucleus for rotations involving species of Meloidogyne. Cotton is a non-host for M. arenaria which attacks groundnut, and the latter is a non-host for M. incognita which attacks cotton. Some cultivars of wheat are also poor hosts for both of these nematodes. Thus, such limited information can be used as a starting-point for rotation systems for these important crops where sufficient land is available.
Much information on the distribution of major nematodes, their host ranges and potential management options, including rotation, is becoming available for Near East countries (Al-Hazmi, 1988; Maqbool, Hashmi and Ghaffar, 1988; Luc, Sikora and Bridge, 1990; Ibrahim and Rezk, 1988; Ibrahim, Rezk and Ibrahim, 1988; Saxena, Sikora and Srivastava, 1988). Also, more comprehensive data on potential weed hosts are becoming available (Montasser, 1990).
Inter- and intracropping is a widely used practice in several countries. Various types of mixed cropping and asynchronous temporal planting (Table 6) may limit nematode damage as outlined by Raymundo (1985). Definitive data on these practices are rather limited.
Cover/trap crops and antagonistic plants are useful for reducing nematode populations as well as conserving soil and often improving soil texture (Nusbaum and Ferns, 1973; Trivedi and Barker, 1986). In localities where land availability permits, the use of cover crops, especially plants that serve as trap crops or offer other suppressive effects on nematode populations, should be considered (Table 8). Carefully selected cover crops may serve as living mulches and provide multiple pest control (National Academy of Sciences, 1991).
TABLE 8
Trap crops for plant-parasitic nematodes1
|
Nematode species |
Trap crop |
Reference |
|
Meloidogyne spp. |
Crotalaria spectabilis, cowpea, English pea, periwinkle |
Patel et al., 1991; Christie, 1959; Godfrey and Hagan, 1934 |
|
Heterodera avenae |
Oat |
Stone, 1961 |
|
H. schachtii |
Hesperis matronalis |
Moriarty, 1961 |
|
Globodera spp. |
Potato |
Carrol and McMahon, 1939 |
1 In pan, after Trivedi and Barker (1986).
Organic amendments offer considerable potential for restricting populations of plant-parasitic nematodes (Stirling, 1991; Jairajpuri, Alam and Ahmad, 1990; Trivedi and Barker, 1986). Soil amendments with oilcakes, green manure, mature crop residues, chitin-containing materials, hemicellulose and farm manures may provide varying levels of nematode suppression. Leaves of neem (Azadirachta spp.) and products thereof provide nematicidal and other pesticidal activity (National Academy of Sciences, 1992a). A chitin-based preparation was recently released as a nematicide in the United States. Unfortunately, this product failed to provide acceptable levels of nematode suppression. A combination of green manure crops or other organic amendments with promising biological control agents may have potential for nematode management on a large-field basis (Abu-Laban and Saleh, 1992; Bansal, Walia and Bhatti, 1988; Trivedi and Barker, 1986).
Fallow is a simple tactic for reducing nematode populations through starvation, as suggested by Tyler in 1933. The goal with this practice is to free the soil of all vegetation for varying periods of time, often by frequent tillage. Theoretically this tactic should be more effective in hot, arid climates then in cooler regions that have high rainfall. However, a number of nematode species inhabiting dry habitats may survive for extended periods through anhydrobiosis. Also, cysts of Heterodera or Globodera spp. may be tolerant to desiccation and/or high temperatures and survive for prolonged periods in the absence of host plants. Additional disadvantages of this tactic include the problems of increased soil erosion, adverse effects on soil structure, and the fact that this practice fails to contribute directly to farm income.
The time of planting/harvesting may be utilized to exploit differential environmental effects on nematode populations versus crop growth and maturity. For example, early planting of crops such as wheat, barley, rye, chickpea and potato has restricted associated nematode damage in some instances (Brown, 1987; Duncan, 1991; Trivedi and Barker, 1986). Because of the prevailing temperatures and the conditions required for optimum growth of most crops, this approach often is impractical.
Overall farm hygiene and culture may also play a key role in limiting crop damage caused by plant-parasitic nematodes. The typical nutrient deficiencies resulting from root damage and impaired uptake effected by nematode parasitism may be offset, in part, by proper nutrition, moisture and protection from adverse conditions (National Academy of Sciences, 1968; Brown, 1987; Trivedi and Barker, 1986). Adequate potassium levels may enable a number of plants to tolerate large populations of nematodes. Ammonium sources of nitrogen have been shown to suppress a number of nematodes, including H. glycines. Based on research with D. dipsaci on alfalfa, high calcium levels may increase host resistance to nematodes. The integration of crop management and compatible cultural tactics might be viewed as the basic foundation for successful nematode control and crop production (Trivedi and Barker, 1986).
Vertical resistance. This type of resistance in plants to pathogens, including nematodes, theoretically falls under the strategy of reducing initial inoculum level (Vanderplank, 1963). This situation clearly is true for resistant cultivars that function in large part as trap crops. (The main discussion on resistance is given under the strategy of suppressing nematode reproduction below.)
Chemical nematicides. Nematicides available on the world markets (Sasser, 1989) may be classified as fumigants and non-fumigants, and considerable research is focusing on the development of novel compounds. The properties that are responsible for soil fumigants being excellent nematicides may result in their being placed under regulatory review (Bird, 1987). To date, DBCP, EDP and DD have been withdrawn from United States markets through the action of regulatory agencies. In addition, 1,3-D has been removed from current use in California, and there is a risk that methyl bromide will be removed from the United States markets. The latter compound has been implicated as an agent that damages the ozone layer. Although a range of non-fumigant nematicides have acceptable levels of efficacy, most of these compounds have been detected in groundwater (Bird, 1987). In addition, some reports have focused on the microbial breakdown of some of these materials in soil, especially under continuous usage (Wu, Stachyra and Viglierchio, 1992).
In spite of the problems associated with nematicides, they comprise an important tactic for many nematode IPM systems, especially for high-value crops. As indicated by Sasser (1989), the procedure for selecting a nematicide should include procurement of information on efficacy and capacity to enhance yields of the primary crop or crops in a specific region. Sasser (1989) and Thomason (1987) provide details on nematicide safety and environmental issues, related economics and additional problems concerning factors in selecting and using these materials, their methods of application and dosage and use on annual or perennial crops.
Biological control agents. Increasing interest in this important nematode management tactic is reflected in two recent books (Stirling, 1991; Jairajpuri, Alam and Ahmad, 1990). As we gain greater understanding of the ecology and biology of nematode antagonists, parasites and predators, they undoubtedly will become important in an integrated approach to managing plant-parasitic nematodes as suggested by Stirling (1991). This topic is discussed by Dr Kerry on page 79.
Physical. The concept of physical means of reducing nematode populations was considered by Atkinson (1889), Bessey (1911) and Tyler (1933). Postharvest ploughing or discing can result in a 90 percent or greater reduction of nematode populations (Fig. 7; Jain and Bhatti, 1989). The prevailing high temperatures in much of the Near East may add to the effectiveness of this tactic during much of the year.
Heat solarization may be an effective nematode management tactic for small land plots in some regions of the Near East. For example, Saleh, Abu-Gharbieh and Al-Banna (1989) showed that a combination of the application of solar-heated water and solarization (with a plastic cover) resulted in a decline of 96 percent in populations of Meloidogyne javanica in Jordan. With this treatment, soil temperatures at 10 to 20 cm depths reached 60° and 56°C, respectively, whereas a maximum temperature of 46°C was achieved with solarization alone. Still, the addition of a nematicide or another tactic may be needed to achieve this level of control in some regions (Greco et al., 1990). The likely differential sensitivity of different nematode species to high temperatures should be considered in using this tactic. For example, M. javanica can tolerate temperatures of 50°C up to three hours or more, whereas M. hapla and M. incognita are killed in much shorter times at this temperature. Solarization also provides some multiple pest control (National Academy of Sciences, 1991).
Flooding has been evaluated periodically for decades as a means of reducing nematode populations (Duncan, 1991; Trivedi and Barker, 1986). At high temperatures, and especially with organic amendments, this treatment can be effective in reducing nematode levels. In contrast, flooding plants such as cranberries under extremely cool conditions may have little impact on parasitic nematodes. Because of the extensive use of irrigation in much of the Near East, the potential of delivery of nematode antagonists or parasites with the water warrants careful study.
Suppression of nematode reproduction
Host-plant resistance will provide an increasingly important contribution to the solution of many nematode problems (Roberts, 1992). With the enhanced availability of germplasm that has nematode resistance genes and rapidly advancing molecular-transfer techniques, resistant cultivars should become a primary management tactic in nematode IPM. This approach will become more critical as chemical nematicides become more restricted. The striking returns from deployment of some nematode-resistant cultivars reflect the great potential of this management tactic. For example, Bradley and Duffey (1982) in an economic analysis concluded that the soybean cultivar Forrest, resistant to M. incognita and H. glycines, gave a US$400 million return over a six-year period (the cost of development of this cultivar was US$1 million).
A number of reviews on the development and deployment of host-plant resistance are available (Roberts, 1992). Except for the stem nematode D. dipsaci, most of the available host resistance is limited to species that have sedentary feeding habits. This situation probably results from three factors: the reproductive activity of these sedentary nematodes can be assessed more readily than migratory endoparasites or ectoparasites; the intimate host-parasite relationship of sedentary nematodes is more likely to be controlled genetically than those of migratory species; and the real or perceived importance of the maladies induced by sedentary nematodes results in much greater research on host resistance than for other feeding groups.
The list of highly effective host-plant resistances to given nematodes is rather limited (Roberts, 1992; Table 9). This listing is restricted to only seven nematode genera on a relatively narrow range of crops. Still, some resistance has been developed for other nematodes, including Pratylenchus spp., Naccobus aberrans, Rotylenchulus reniformis, Radopholus citrophilus, Xiphinema americanum and Criconemella xenoplax. As yet, there is
TABLE 9
Host resistance to nematodes1
|
Nematode species |
Crop |
|
Highly effective | |
|
Ditylenchus dipsaci |
Alfalfa, oat, red clover |
|
Meloidogyne spp. |
Alfalfa, Prunus (Nemaguard rootstock), |
|
Heterodera avenae |
Barley, oat |
|
Tylenchulus semipenetrans |
Citrus (Poncirus trifoliata) |
|
Meloidogyne incognita |
Cowpea, lima and common bean, soybean, tobacco |
|
Globodera rostochiensis |
Potato |
|
Heterodera glycines |
Soybean |
|
Some developed resistance | |
|
Heterodera avenae |
Barley, oat |
|
Tylenchulus semipenetrans |
Citrus |
|
Radopholus citrophilus |
" |
|
Meloidogyne incognita |
Cotton |
|
Rotylenchulus reniformis |
" |
|
Meloidogyne javanica |
Cowpea, tobacco |
|
Meloidogyne arenaria |
" |
|
Heterodera zeae |
Maize2 |
|
Globodera rostochiensis |
Potato |
|
Globodera pallida |
" |
|
Meloidogyne spp. |
" |
|
Nacobbus aberrans |
" |
|
Pratylenchus vulnus |
Prunus spp. |
|
Xiphinema americanum |
" |
|
Criconemella xenoplax |
" |
|
Heterodera glycines |
Soybean |
|
Meloidogyne spp. |
" |
|
Rotylenchulus reniformis |
" |
|
Meloidogyne hapla |
Tomato |
|
Meloidogyne incognita |
" |
|
Meloidogyne javanica |
" |
|
Meloidogyne arenaria |
" |
|
No developed resistance | |
|
Heterodera schachtii |
Sugar beet, crucifers |
|
Heterodera avenae |
Wheat |
|
Meloidogyne spp. |
Barley, carrot, maize, cucurbits, eggplant, lettuce, groundnut, sugar beet, many others |
|
Meloidogyne hapla |
Most host crops |
|
Rotylenchulus reniformis |
Tomato, many host crops |
|
Naccobus aberrans |
Pepper, sugar beet, tomato |
1 Largely after Roberts (1992) and Sasser (1989).2 Some sources identified (L.K. Krusberg, personal communication). no developed resistance for a number of important nematode-host combinations (Table 9).
The deployment of host-resistant cultivars has been accomplished largely on a regional basis (Roberts, 1992). A world survey by Sasser and Freckman (1987) indicated that at least some nematode-host resistance or tolerance is available on one or more crops in all regions surveyed. In Africa, however, only four crops were reported to have tolerance or resistance: bean, cowpea, tomato and yam. The resistant host lists for other geographic regions is somewhat longer and South America had the most extensive list with 26 plant species. The list for Asia was intermediate with 18 crop plants exhibiting some nematode resistance.
The available host resistance to nematodes also has some limitations. For example, resistance to Meloidogyne spp. in bean and tomato plants is heat-sensitive, and thus may fail in hot production regions at lower latitudes (Roberts, 1992). In addition, crop cultivars with resistance to Meloidogyne spp. are often susceptible to other associated nematode species. The latter limitation is not a serious restriction in fields that have a primary dominant or target nematode species. However, the restricted deployment of host-plant resistance in the large African continent reflects a critical regional and worldwide need for greater collaboration between nematologists and plant breeders in the development and deployment of this important tactic for reducing nematode problems.
Available host-plant resistance to nematodes should be deployed in carefully developed cropping systems. Monocultures with resistant cultivars frequently result in the appearance of biotypes of the target nematode that circumvent the resistance (Young, 1992).
Supplemental chemical nematicides have come to the forefront in some regions as a means of suppressing the increase of nematodes on high-value crops such as groundnut or citrus (Thomason, 1987). Repeated applications of low dosages through trickle irrigation may suppress nematode multiplication, resulting in the growth of a healthy crop.
Restriction of current crop damage
Tolerant cultivars of suitable host species grow and yield well in spite of nematode parasitism (Oostenbrink, 1972). Although such hosts have near-normal shoot and root growth, nematode population densities build up to levels that often severely damage subsequent susceptible crops. For example, maize may support relatively high reproduction of M. incognita or M. arenaria, and still produce near-normal yields. Thus, such crops should be examined carefully before placing them in rotations with highly susceptible crops. A few plant breeders and nematologists are working to combine host-plant resistance and tolerance to nematodes. Resistant/tolerant cultivars clearly would have an advantage in that some resistant cultivars may be severely stunted by high population densities of the target nematode.
As indicated earlier, Tyler (1933) suggested that two or more management tactics be combined to develop effective control programmes for root-knot nematodes, and also stated that the value and permanence of chemical or cultural treatment would be increased when followed by a wet fallow or resistant crop, with particular attention to the control of weed hosts. As highly efficacious, inexpensive chemical soil treatments have been removed from the market, the integration of available management tactics and the maximizing of the potential of cropping systems must receive much greater attention (Noe, Sasser and Imbriani, 1991; Roberts, 1993).
To date, most IPM nematology research has focused on population biology, host phenology, modelling, damage functions and overall pest management decision-making (Bird, 1987; Roberts, 1993). Progress in these areas has clearly facilitated the development of better management programmes as well as enhancing our understanding of nematode-host interactions. Nevertheless, the integration and assessment of combinations of potentially compatible nematode management strategies and tactics have received only limited attention. This situation has developed, in large part, from heavy reliance on the use of nematicides, rotations where practical and resistant cultivars when available. The integration of host resistance and nematicides with other compatible tactics should minimize potential problems of loss of efficacy of given compounds or the appearance of nematode biotypes that attack resistant cultivars.
Integration of nematode management may be considered at two levels: within nematology; and across pathogen/pest disciplines. Within the context of nematodes, the application of multiple management tactics may be done over time or simultaneously, or both (Roberts, 1993). The temporal approach focuses on season-to-season or year-to-year integration of tactics relevant to cropping cycles. This cycle of integration of tactics might focus on crop rotation with non-hosts, resistant cultivars, or both, with additional strategies such as nematicides, cultural practices or biological control being added as they become available. This approach could involve different control tactics on different crops for given years. The second approach described by Roberts (1993) for integrating nematode-control components, involved simultaneous use of two or more tactics. This approach requires compatibility among the tactics to be deployed.
Numerous examples of single and multiple pest/pathogen successes could be cited (Duncan, 1991; Luc, Sikora and Bridge, 1990). Combinations of host resistance and tolerance, nematicides and rotations of resistant and susceptible potatoes with non-host rotation crops have provided a successful integrated management programme for the potato-cyst nematode (Roberts, 1993; Trudgill, 1987). A rather different system focusing host resistance, crop rotation, residual host-root destruction immediately after harvest, and an appropriate nematicide where warranted, has proved to be highly successful for Meloidogyne spp. and certain fungi, bacteria, insects and weeds on tobacco in North Carolina (Melton, 1991).
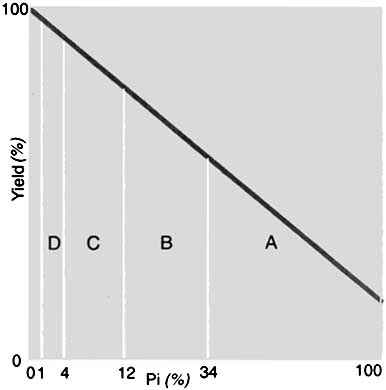
Roberts (1993) also offered a theoretical but practical framework for the integration of nematode management tactics with only partial efficacy (Fig. 9). This example depicts four hypothetical control tactics, A-D, with each having an efficacy of 66 percent. Thus, stepwise, residuals of the initial nematode population levels per successive tactic would be 34, 11.6, 2.9 and 1.3 percent, respectively. Information on the damage threshold for the target nematodes and the efficacy of the integrated management tactics deployed, would be essential to determining the number of the types of treatments necessary to achieve satisfactory crop yields. For highly aggressive nematode species such as M. arenaria on groundnut, an even greater number of these types of tactics might be required in a management programme. Clearly, more information on the potential interactions of available management tactics, whether antagonistic or synergistic, or simply additive effects, will be critical for the integration of nematode control strategies and tactics.
Much progress has been made in characterizing nematode communities and in the assessment of some management tactics in the Near East. Research opportunities remain in these areas as well as in developing related damage functions/thresholds and the integration of compatible strategies/tactics. Candidates for this integration include rotation/cropping systems, resistant cultivars, fallow/destruction of residual infected roots, solarization, nematicides and soil amendments/biological control. Carefully developed research-extension priorities, and the use of some type of data/decision-support system, should facilitate the development and deployment of effective IPM programmes. Long-term IPM goals and priorities might focus on interdisciplinary IPM (all pathogen and pest groups) and sustainable crop-production systems with critical inputs from experts in technology development/transfer, institutional resources/training, and economic/social/cultural components.
Promising new technologies for nematode diagnosis and identification, critical to IPM, are on the horizon. In addition, novel management weapons, including transgenic plants with nematode resistance and/or biological control genes, new safer nematicides and more effective biological control agents should be forthcoming. Improved data manage- ment and decision-support systems should facilitate the integration of new and traditional IPM strategies and tactics.
Abu-Laban, A.Z. & Saleh, H.M. 1992. Evaluation of animal manures for mass production, storage and application of some nematode egg-parasitic fungi. Nematologica, 38: 237-244.
Al-Hazmi, A.S. 1988. Relative host suitability of corn and alfalfa cultivars to Meloidogyne javanica. Pak. J. Nematol., 6: 101-105.
Alston, D.G., Bradley, J.R., Schmitt, D.P. & Coble, H.D. 1991. Response of Helicoverpa zea (Lepidoptera: Noctuidae) populations to canopy development in soybean as influenced by Heterodera glycines (Nematoda:Heteroderidae) and annual weed population densities. J. Econ. Entomol., 84: 267-276.
Atkinson, G.F. 1889. A preliminary report upon the life history and metamorphoses of a root-gall nematode, Heterodera radicicola (Greef) Müller, and the injuries caused by it upon the roots of various plants. Ala. Agric. Expt. Stn. Bull., n.s. 9. 55 pp.
Bansal, R.K., Walia, R.K. & Bhatti, D.S. 1988. Evaluation of some agro-industrial wastes for mass propagation of the nematode parasitic fungus Paecilomyces lilacinus. Nematol. Medit., 16: 135-136.
Barker, K.R. 1985a. Nematode extraction and bioassays. In K.R. Barker, C.C. Carter & J.N. Sasser, eds. An advanced treatise on Meloidogyne. Vol. II, p. 19-35. Raleigh, NC, USA, North Carolina State University Graphics.
Barker, K.R. 1985b. A history of the introduction and spread of nematodes. In D.R. Mackenzie, C.S. Barfield, G.G. Kennedy, R.D. Berger & D.J. Taranto, eds. The movement and dispersal of agriculturally important biotic agents, p. 131-144. Baton Rouge, Louisiana, USA, Claitor's Publishing Division.
Barker, K.R. & Imbriani, J.L. 1984. Nematode advisory programs - status and prospects. Plant Dis., 68: 735-741.
Barker, K.R., Schmitt, D.P. & Imbriani, J.L. 1985. Nematode population dynamics with emphasis on determining damage potential on crops. In K.R.
Barker, C.C. Carter & J.N. Sasser, eds. An advanced treatise on Meloidogyne. Vol. II, p. 135-148. Raleigh, NC, USA, North Carolina State University Graphics.
Beane, J. & Perry, R. 1990. The influence of certain herbicides in pelleted form on the hatch and invasion of Globodera rostochiensis, G. pallida and Heterodera schachtii. Rev. Nematol., 13: 275-281.
Bessey, E.A. 1911. Root-knot and its control. USDA Bur. Plant Ind. Bull., 217.
Bird, G.W. 1987. Role of nematology in integrated pest management programs. In J.A. Veech & D.W. Dickson, eds. Vistas on nematology, p. 114-121. Hyattsville, Maryland, USA, Society of Nematologists.
Bradley, E.B. & Duffey, M. 1982. The value of plant resistance to soybean cyst nematode: a case study of Forrest soybeans. Report No. AGE820929. Natural Resources Economic Division. Washington, DC. USDA.
Bridge, J. 1988. Analysis of methods for determining presence, distribution and importance of nematodes on a regional basis. In M.C. Saxena, R.A. Sikora & J.R Srivastava, eds. Nematodes parasitic to cereals and legumes in temperate semi-arid regions, p. 125-141. Aleppo, Syrian Arab Republic, ICARDA.
Brown, R.H. 1987. Control strategies in low-value crops. In R.H. Brown & B.R. Kerry, eds. Principles and practice of nematode control in crops, p. 351-387. Orlando, Florida, USA, Academic Press.
Carrol, J. & McMahon, E. 1939. Experiments on trap cropping with potatoes as a control measure against potato eelworm (Heterodera schachtii). J. Helminthol., 17: 101-112.
Christie, J.R. 1959. Plant nematodes, their bionomics and control. Gainesville, Florida, USA, Florida Agricultural Experiment Station.
Cobb, N.A. 1918. Estimating the nema population of soil. USDA Agricultural Circular No. 1. Washington, DC, USDA. 48 pp.
Duncan, L.W. 1991. Current options for nematode management. Annu. Rev. Phytopathol., 29: 469-490.
Eissa, M.F.M. 1988. Loss estimation for winter season cereal and legume crops due to plant-parasitic nematodes and complex diseases in Egypt. In M.C. Saxena, R.A. Sikora & J.R Srivastava, eds. Nematodes parasitic to cereals and legumes in temperate semi-arid regions, p. 147-159. Aleppo, Syrian Arab Republic, ICARDA.
EPPO. 1982. Recommendations on new quarantine measures (2nd edition). European and Mediterranean Plant Protection Organization Bulletin. Special Issue. Paris. 52 pp.
Ferris, H. & Noling, J.W. 1987. Analysis and prediction as a basis for management decisions. In R.H. Brown & B.R. Kerry, eds. Principles and practice of nematode control in crops, p. 49-85. Orlando, Florida, USA, Academic Press.
Godfrey, G.H. & Hagan, H.R. 1934. A study of the root-knot nematode trap crop under field soil conditions. Phytopatholgy, 24: 648-658.
Goodey, J.B., Franklin, M.T. & Hooper, D.J. 1965. T. Goodey's The nematode parasites of plants catalogued under their hosts (3rd ed.). Famham Royal, Bucks, UK, CAB. 214 pp.
Greco, N., D'Addabbo, Brandonisio, A. & Zweep, A. 1990. Combined effect of soil solarization and 1,3 dichloropropene for control of Heterodera carotae. Nematol. Medit., 18: 261-264.
Hartman, K.M. & Sasser, J.N. 1985. Identification of Meloidogyne species on the basis of differential host tests and perineal-pattern morphology. In K.R. Barker, C.C. Carter & J.N. Sasser, eds. An advanced treatise on Meloidogyne. Vol. II, p. 69-77. Raleigh, NC, USA, North Carolina State University Graphics.
Ibrahim, I.K.A. & Rezk, M.A. 1988. The root-knot nematodes - a major problem in crop production in Egypt. In M.A. Maqbool, A.M. Golden, A. Ghaffar & L.R. Krusberg, eds. Advances in plant nematology, p. 81-98. Karachi, Pakistan, National Nematological Research Centre, University of Karachi.
Ibrahim, I.K.A., Rezk, M.A. & Ibrahim, A.A.M. 1988. Resistance of barley and wheat cultivars to root-knot nematodes, Meloidogyne spp. Pak. J. Nematol., 6: 39-43.
Jain, R.K. & Bhatti, D.S. 1989. An integrated approach for the management of root-knot nematode (Meloidogyne javanica) in tomato. Ind. J. Nematol., 19: 159-161.
Jairajpuri, M.S., Alam, M.M. & Ahmad, I., eds. 1990. Nematode bio-control. Delhi, India, CBS Publ. and Distr. Pvt. Ltd. 152 pp.
Luc, M., Sikora, R.A. & Bridge, J., eds. 1990. Plant-parasitic nematodes in subtropical and tropical agriculture. Wallingford, UK, CAB International. 629 pp.
Maqbool, M.A., Hashmi, S. & Ghaflar, A. 1988. Problem of root-knot nematodes in Pakistan and strategy for their control. In M.A. Maqbool, A.M. Golden, A. Ghaffar & L.R. Krusberg, eds. Advances in plant nematology, p. 229-240. Karachi, Pakistan, National Nematological Research Centre, University of Karachi.
Melton, T.A. 1991. Disease management practices. In 7997 Flue-cured information, p. 81-107. Raleigh, NC, USA, North Carolina Agric. Ext. Serv. AG-187, North Carolina State University.
Montasser, S.A. 1990. New non-crop hosts of the root-knot nematode, Meloidogyne incognita in Egypt. Pak. J. Nematol., 8: 39-41.
Moriarty, F. 1961. The effects of red beet and of Hesperis matronalis L. on a population of Heterodera schachtii. Schm. Nematologica, 6: 214-221.
National Academy of Sciences. 1968. Principles of plant and animal pest control. 4. Control of plant-parasitic nematodes. Washington, DC. 172 pp.
National Academy of Sciences. 1991. Sustainable agriculture research and education. Washington, DC. 437 pp.
National Academy of Sciences. 1992a. Neem: a tree for solving global problems. Washington, DC. 149 pp.
National Academy of Sciences. 1992b. Toward sustainability: an addendum on integrated pest management, a component of sustainability research. Washington, DC. 36 pp.
Noe, J.P. 1986. Cropping systems analysis for limiting losses due to plant-parasitic nematodes. Guide to research methodology. Raleigh, NC, USA, North Carolina State University Graphics. 18 pp.
Noe, J.P., Sasser, J.N. & Imbriani, J.L. 1991. Maximizing the potential of cropping systems for nematode management. J. Nematol., 23: 353-361.
Noe, J.P. & Sikora, R.A. 1990. Effects of tropical climates on the distribution and host-parasite relationship of plant-parasitic nematodes. In M. Luc, R.A. Sikora & J. Bridge, eds. Plant-parasitic nematodes in subtropical and tropical agriculture, p. 583-597. Wallingford, UK, CAB International.
Nusbaum, C.J. & Ferris, H. 1973. The role of cropping systems in nematode population management. Ann. Rev. Phytopathol., 11: 423-440.
O'Bannon, J.H. & Esser, R.P. 1987. Regulatory perspectives in nematology. In JA. Veech & D.W. Dickson, eds. Vistas on nematology, p. 38-46. Hyattsville, Maryland, USA, Society of Nematologists.
Oostenbrink, M. 1972. Evaluation and integration of nematode control methods. In J.M. Webster, ed. Economic nematology, p. 497-514. New York, NY, USA, Academic Press.
Patel, H.R., Patel, DJ., Patel, C.C. & Thakar, N.A. 1991. Management of root-knot nematodes by Periwinkle. Nematol. Medit., 19: 65-66.
Piotte, C., Castegnone-Sereno, P., Uijthof, J., Abad, P., Bongiovanni, M. & Dalmasso, A. 1992. Molecular characterization of species and populations of Meloidogyne from various geographic origins with repeated DNA homologous probes. Fundam. Appl. Nematol., 15: 271-276.
Raymundo, S.A. 1985. Cropping systems research and root-knot nematode control. In J.N. Sasser & C.C. Carter, eds. An advanced treatise on Meloidogyne. Vol. I. Biology and control, p. 227-281. Raleigh, NC, USA, North Carolina State University Graphics.
Roberts, P.A. 1992. Current status of the availability, development and use of host plant resistance to nematodes. J. Nematol., 24: 213-227.
Roberts, P.A. 1993. Future of nematology - integration of new and improved management strategies. J. Nematol., 25.
Saleh, H., Abu-Gharbieh, W.L & Al-Banna, L. 1989. Augmentation of soil solarization effects by application of solar-heated water. Nematol. Medit., 17: 127-129.
Sarah, J.L. 1989. Nematological review - banana nematodes and their control in Africa. Nematropica, 19: 199-216.
Sasser, J.N. 1989. Plant-parasitic nematodes: the farmer's hidden enemy. Raleigh, NC, USA, North Carolina State University Graphics. 115 pp.
Sasser, J.N. & Freckman, D.W. 1987. A world perspective on nematology: the role of the society. In J.A.Veech & D.W. Dickson, eds. Vistas on nematology, p. 7-14. Hyattsville, Maryland, USA, Society of Nematologists.
Saxena, M.C., Sikora, R.A. & Srivastava, J.R, eds. 1988. Nematodes parasitic to cereals and legumes in temperate and semi-arid regions, p. 69-84. Aleppo, Syrian Arab Republic, ICARDA.
Schmitt, D.R, Barker, K.R., Noe, J.P. & Koenning, S.R. 1990. Repeated sampling to determine the precision of estimating nematode population densities. J. Nematol., 22: 552-559.
Schmitt, D.P. & Nelson, L.A. 1987. Interactions of nematicides with other pesticides. In JA Veech & D.W. Dickson, eds. Vistas on nematology, p. 455-460. Hyattsville, Maryland, USA, Society of Nematologists.
Schots, A., Gommers, F.J. & Egberts, E. 1992. Quantitative ELISA for the detection of potato-cyst nematodes in soil samples. Fundam. Appl. Nematol., 15: 55-61.
Seinhorst, J.W. 1970. Dynamics of populations of plant-parasitic nematodes. Annu. Rev. Phytopathol., 8: 131-156.
Stirling, G.R. 1991. Biological control of plant-parasitic nematodes. Wallingford, UK, CAB International. 282 pp.
Stone, L.E.W. 1961. Oats as a trap-crop for cereal root eelworm. Plant Path., 10: 164.
Taylor, C.E. 1986. Quarantine and legislation. In F. Lamberti & C.E. Taylor, eds. Cyst nematodes, p. 451-459. New York, NY, USA, Plenum Press.
Thomason, I.J. 1987. Challenges facing nematology: environmental risks with nematicides and the need for new approaches. In J.A. Veech & D.W. Dickson, eds. Vistas on nematology, p. 469-476. Hyattsville, Maryland, USA, Society of Nematologists.
Travis, J.W. & Latin, R.X. 1991. Development, implementation, and adoption of expert systems in plant pathology. Annu. Rev. Phytopathol., 29: 343-360.
Trivedi, P.C. & Barker, K.R. 1986. Management of nematodes by cultural practices. Nematropica, 16: 213-236.
Trudgill, D.L. 1987. Effects of rates of a nematicide and of fertilizer on the growth and yield of cultivars of potato which differ in their tolerance of damage by potato-cyst nematodes (Globodera rostochiensis and G. pallida). Plant & Soil, 104: 235-243.
Tyler, J. 1933. The root-knot nematode. Berkeley, CA, USA, Circular No. 330, University of California College of Agriculture, Agricultural Experiment Station. 33 pp.
USDA. 1960. Index of plant diseases in the United States. USDA Agric. Res. Ser., Agric. Handbook No. 165. Washington, DC. 531 pp.
Vanderplank, J.E. 1963. Plant diseases: epidemics and control. New York, NY, USA, Academic Press. 349 pp.
Wheeler, T.A., Madden, L.V., Rowe, R.C. & Riedel, R.M. 1992. Modelling of yield loss in potato early dying caused by Pratylenchus penetrans and Verticillium dahliae. J. Nematol., 24: 99-102.
Wu, F.F., Stachyra, A. & Viglierchio, D.R. 1992. The capacity of soil microbial isolates from non-fumigant nematicide stressed Heterodera schachtii greenhouse culture stocks to inactivate non-fumigant nematicides. Fundam. Appl. Nematol., 15: 193-200.
Young, L.D. 1992. Problems and strategies associated with long-term use of nematode resistant cultivars. J. Nematol., 24: 228-233.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}