![]()
![]()
![]()
MICHAEL H. UNSWORTH
Center for Analysis of Environmental Change, Oregon State University, Corvallis, Oregon, USA
WILLIAM E. HOGSETT
US Environmental Protection Agency, Environmental Research Laboratory, Corvallis,
Oregon, USA
Carbon dioxide
CO2 and temperature
CO2 and water availability
CO2 and ozone
CO2 and UV-B radiation
Ozone
Ozone and temperature
Ozone and water availability
Ozone and UV-B radiation
Concluding remarks
Acknowledgements
References
Our understanding of the relationships between crop growth and the atmospheric environment has developed substantially in the past few decades. Improvements in technology have enabled crop physiologists to study processes such as transpiration and carbon dioxide exchange in the field. Mechanistic understanding of the factors controlling photosynthesis and respiration has improved greatly, but we still have no mechanistic understanding of the ways in which many abiotic stresses affect these processes. Our knowledge of factors that control crop development and the partitioning of assimilate between organs is still at the descriptive stage, and poses one of the main limitations on our ability to develop fully mechanistic mathematical models of crop growth. Nevertheless, crop science has reached a stage where reliable semi-empirical simulation models of some of the major world crops have been constructed and used to explore the potential for crop growth and yield in different environments. For the reasons discussed above, these models are restricted in their application to situations where the empirical relationships in them are well defined, but they are often useful in the design of particular experiments to investigate new environments, for example, to apply them to scenarios of climatic change. It should be emphasized that, although we have a much improved understanding of the growth and yield potential of major agricultural crops, this knowledge needs to be extended to include other crops, especially those important for developing countries.
However, as we steadily improve our understanding of how crop yield is influenced by the atmospheric environment, that environment is constantly changing as a result of human activities, and the challenge today is to predict how crops will respond to the changed environments of tomorrow. Traditionally, crop scientists have used controlled environments and field-based facilities to investigate crop responses to individual factors, such as radiation, temperature, humidity and water availability. More recently, manipulation of air quality has been used to study crop responses to air pollutants, such as SO2, O3 NOx, and some of the technology developed for these experiments has been successfully applied to study responses to increased CO2. Although this 'single component' approach is a highly effective way of discovering how one atmospheric factor influences crops, in practice the atmosphere in the future will alter in several ways simultaneously.
Some of the impending changes are more certain than others. Without doubt, CO2 concentration is increasing worldwide at about 1.8 m mol/mol per year and under all likely scenarios will continue to do so. This increase will enhance the global greenhouse effect, and the consensus of scientific opinion is that it will result in global warming and in changed global distributions of rainfall and other weather components. The amount of climatic change is, however, very uncertain, because even the best computer simulation models do not adequately describe features such as ocean-atmosphere linkages, and feedbacks associated with changes in atmospheric aerosols or cloud amount and distribution. At present, expert opinion is that, when the CO2 concentration reaches double its present value of about 350 m mol/mol, global temperatures will be about 2.5°C warmer, but this figure and the associated global variability are very uncertain; even more uncertain are any changes in rainfall amount and distribution (Houghton et al., 1992).
In contrast to the uncertainty associated with computer models, there is clear evidence from satellite and from surface monitoring that stratospheric ozone is being depleted, probably because of emissions of chlorine- and bromine-containing substances, so that globally the column of ozone from the surface through the atmosphere is thinning at about 3% per decade, a rate that seems to be increasing (Houghton et al., 1992). Because the chemicals believed to be causing this thinning are long-lived in the stratosphere, this thinning process is likely to continue for several decades even though the chemicals are being phased out of production. The consequence of this thinning is that ultraviolet (UV) radiation at the ground, normally severely attenuated by stratospheric O3 will increase. There is, however, an inadequate network of surface monitoring stations to define the UV climate at the ground (i.e., taking into consideration effects of cloud, dust, etc.) or to determine whether UV has increased significantly. Even if annual mean UV exposure has not changed, there are indications that thinning of stratospheric ozone, similar but less extreme than that observed in the Antarctic, can occur episodically at temperate latitudes.
Air quality in the troposphere is also changing. In particular, in many highly populated regions, emissions of hydrocarbons and exhaust gases from motor vehicles provide the precursors for photochemical production of tropospheric ozone. There is evidence that background tropospheric ozone concentrations have increased by about 10% per decade in Europe over the past 20 years, and that the European background has approximately doubled since the beginning of the century (Houghton et al., 1992). Although there are no equivalent long-term records, it is likely that the tropospheric O3, concentration in the United States and Southeast Asia has also increased. Perhaps more important than the mean background concentration of O3 is the regular occurrence on a regional scale of high concentrations of tropospheric O3 (typically 5 to 10 times background) associated with stagnant weather systems in summer. These concentrations have been demonstrated to reduce the growth and yield of many agricultural crops.
The atmospheric environment of many world crops of the future will undoubtedly contain more CO2 than present, is likely to be somewhat warmer, and rainfall (and hence humidity) and cloudiness will probably also change, though in a direction unknown at present. Whether UV will increase much on a seasonal basis remains unclear, but episodes of increased UV at the ground, associated with patches of thinned stratospheric O3, may occur, especially in temperate latitudes. Without doubt, in regions with large population densities, episodes of high concentrations of tropospheric O3 and of other pollutants associated with fossil-fuel burning will become more common.
In this paper we review our knowledge of some of the interacting effects of these changing atmospheric conditions on agricultural crops, and we draw attention to particular gaps where further research is needed. In an attempt to structure what could otherwise be a very disparate summary, we will concentrate on two major gases, CO2 and O3, and will describe work that has investigated the effects of other stresses combined with them. In each case, we first briefly review responses to these gases alone.
Several recent reviews of responses of crops to CO2 have been published (Goudriaan and Unsworth, 1990; Lawlor and Mitchell, 1991; van de Geijn et al., 1993; chapters 4 and 5 of this publication), and there is no advantage in repeating the detail of these here. At the leaf level, the two most well-known responses to elevated CO2 are an increase in the rate of net photosynthesis, PN, and a decrease in stomatal conductance, gs. The increase in P is much larger in C3 species (50 to 100% when CO2 concentration doubles) than in C4 species (10%), but a substantial decrease in g (30 to 40%) is observed in both types of species. There is some evidence of photosynthetic acclimation to elevated CO2, so that increased rates of PN do not persist at the leaf level in long-term studies with some species, but there is no evidence of acclimation in g. It is not clear whether increased CO2 also indirectly affects plant growth, e.g., by changing rates of leaf initiation, expansion or longevity, but such effects could have a large influence on crop productivity.
Much of our understanding of effects of CO2 on plants has been gained from studies with individual leaves. There have been far fewer studies of long-term effects of elevated CO2 on canopy-scale photosynthesis and transpiration. Most of those reported have been in closed- or open-top chambers, though a few reports of field-scale exposures (FACE) are now becoming available. Drake and Leadley (1991) summarized the data available at that time and concluded that: (1) canopy photosynthesis increases in elevated CO2 when there is a sink available for the carbon; (2) the relative effect of CO2 is greatest at highest temperatures; and (3) elevated CO2 alters many interacting factors, such as canopy architecture and partitioning of assimilates, that mediate gas exchange of canopies and ecosystems.
As a consequence of increased net photosynthesis, dry matter production and yield are substantially increased by elevated CO2. Several authors (e.g., Lawlor and Mitchell, 1991) have reviewed the literature and concluded that, provided there is adequate water, nutrients and pest control, yields of C3 and C4 crops growing in about 700 m mol/mol CO2 would be about 30 to 40% and 9%, respectively, greater than present yields (350 m mol/mol CO2) if all other climatic factors were unchanged.
Table 8.1 summarizes harvest indices and yield from recent work at Nottingham, England (Clifford et al., 1993; Azam Ali, pers. comm.) in which stands of groundnut (C3) and sorghum (C4) were grown at two CO2 concentrations in a computer-controlled glasshouse system designed to control temperature, humidity and soil moisture to simulate tropical growing conditions. Although harvest index (seed or pod yield per plant) was not altered by CO2, yield increases were consistent with other reports for unstressed C3 and C4 crops.
However, in much of the world, ideal crop growing conditions, with adequate water and nutrition, are wishful thinking, and the influence of elevated CO2 on aspects such as water-use may be much more relevant. Reviews by Morison (1985) and Eamus (1991) make it clear that, on a leaf area basis, elevated CO2 improves the water-use efficiency (WUE - the ratio of weight of dry matter produced to weight of water transpired) of plants. On a whole-plant basis, the increase in leaf area resulting from enhanced photosynthesis may cause the amount of water-used per plant to remain close to that of plants growing at current levels of CO2, although the WUE as defined here increases. We return to this topic later.
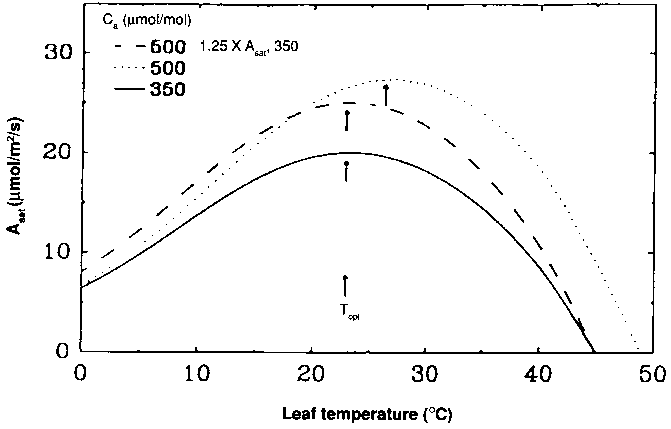
As discussed earlier, the atmospheric environment of the future is likely to include correlated increases of atmospheric CO2 concentration and temperature. Long (1991) reviewed the mechanisms by which temperature and CO2 affect photosynthesis in C3 species, and developed models of the response of leaf canopy carbon exchange to changes in these variables. He showed (Figure 8.1) that the interaction of carbon dioxide concentration and temperature causes the temperature optimum of the light-saturated rate of CO2 uptake to increase as CO2 increases. Without this interaction, an increase in global temperature of 2.5°C would increase the frequency of temperatures in temperate regions that are supra-optimal for light-saturated photosynthesis. However, if warming on this scale was coincident with a CO2 concentration increase to above 500 m mol/mol, the temperature optimum would increase, and this inhibition would not occur. Long (1991) went on to develop a simple model of canopy CO2 uptake (Ac) in response to temperature, taking into account the substantial fraction of the canopy leaf area that would be shaded and therefore not photosynthesizing at the saturated rate. Figure 8.2 shows the model sensitivity of daily canopy carbon uptake to temperature. Using a simple model of the diurnal variation of radiation and temperature at different latitudes, Long showed that the implications of Figure 8.2 are that daily net carbon assimilation responds much more to increased CO2 at low latitudes than at high latitudes. These calculations are in good agreement with observations from long-term studies reported by Oechel and Strain (1985), Drake and Leadley (1991), and Kimball (1983).
Table 8.1. Harvest indices (HI, seed or pod yield per plant/total dry mass per plant) and seed or pod yields for stands of groundnut (Arachis hypogaea L.) and sorghum (Sorghum bicolor L.) grown in controlled-environment glasshouses at 350 m mol/mol or 700 m mol/mol CO2 (from Clifford et al., 1993 and Azam Ali, pers. comm.)
|
Crop |
HI |
Pod/seed yield (g/plant) |
Percentage yield increase at elevated CO2 |
||
|
350 m mol/mol CO2 |
700 m mol/mol CO2 |
350 m mol/mol CO2 |
700 m mol/mol CO2 |
||
|
Groundnut |
0.195 |
0.212 |
9.06 |
13.29 |
30.9 |
|
±0.019 |
±0.038 |
±0.06 |
±1.70 |
±13.3 |
|
|
Sorghum |
0.333 |
0.334 |
25.78 |
27.37 |
6.2 |
|
±0.016 |
±0.015 |
±0.11 |
±0.85 |
±3.3 |
|
Idso (1990) grew crops in field chambers in Phoenix, Arizona, at 300 and 600 m mol/mol CO2. Figure 8.3 shows the ratio of net photosynthesis at the two CO2 concentrations plotted against leaf temperature, and includes values calculated from Long's theoretical relationships, showing the good agreement at the leaf level. For a number of crops grown in open-top chambers in Phoenix, Arizona, Figure 8.4 (from Kimball et al., 1993) summarizes the relative increase in growth (weekly dry matter production, comparing 650 m mol/mol CO2 with 350 m mol/mol) with mean air temperature, indicating that, at low temperatures, elevated CO2 actually decreased growth (a phenomenon predicted by Long's models).
Interactions between temperature and CO2 also need to account for effect of temperature on development. Squire and Unsworth (summarized in Goudriaan and Unsworth, 1990) pointed out that determinate crops with discrete elements to their life cycle develop faster in higher temperatures, and so the stage of seed filling is shortened, limiting the benefits of elevated CO2 Figure 8.5 illustrates results from a model of winter wheat growth that was modified for CO2 response (though in a less sophisticated manner than Long, 1991). When CO2 concentration doubled, but daily weather data from a typical year were used, the potential grain yield was 27% larger than in a control run at 340 m mol/mol CO2 (11.5 and 9.0 t/ha, respectively). The date of maturity was unchanged. When daily temperatures were increased by 3°C and CO2 was doubled, more rapid development of the crop shortened the growing season, and the potential grain yield (10.4 t/ha) was only 15% larger than in the control, but maturity occurred 30 days earlier. The wheat model used in these simulations contains a number of elements that are sensitive to temperature. Recently, Kocabas (1993) completed a detailed investigation of the model sensitivity to changes in mean temperature at various stages of crop development (Kocabas et al., 1993), and to changes in variability of temperature in various developmental phases and over the whole season. The results indicated the particularly strong dependence of yield on temperature in the phase from emergence to double ridges in wheat, but suggested that changes in temperature variability of ±20% would not have any consistent effects on yield variability. It would be useful to incorporate Long's analysis into the wheat model to allow more precise simulation of the interactive effects of temperature and CO2 on carbon assimilation and crop development.
Figure 8.3. Variation of relative CO2 assimilation rate (net photosynthesis at an intercellular CO2 concentration of 600 m mol/mol divided by net photosynthesis at 300 m mol/mol) with leaf temperature for bell pepper, tomato and cottonwood tree. The dotted line is the relationship predicted by the analysis of Long (1991) (adapted from Idso, 1990)

Indeterminate crops, such as grass and sugar beet, continue to grow and produce yield as long as the temperature is above a minimum threshold, provided that other factors such as drought and nutrition do not limit growth. Consequently, simultaneous increases in temperature and CO2 concentration increase the potential yield of such crops in temperate environments, where low temperatures at the beginning and end of the season usually limit productivity.
Perennial tree crops, which yield fruit, nuts and wood, pose complex problems in assessing the effects of CO2 and temperature interactions, because their yield depends on a phased sequence of development over at least two years. Climatic warming could disrupt this sequence, for example, by providing insufficient winter chilling to synchronize spring bud break, or by advancing ovule development and leading to poor fruit set in spring (Cannell et al., 1989). Cannell and Smith (1986) pointed out that climatic warming could either advance or delay bud burst in apples depending on the extent to which chilling requirements were met. The long-term influence of CO2 on tree growth, and its interaction with temperature responses, is very uncertain at present.
Figure 8.5. Calculations using the model ARCWHEAT (Porter, 1984), modified for CO2 responses to illustrate potential production of winter wheat based on weather data from Rothamsted, England. The straight lines correspond to total dry matter production and grain yield predicted at 340 m mol/mol CO2 and observed weather. The curves are predicted production and yield at 680 m mol/mol CO2 and increases of daily mean temperature up to 4°C (from Squire and Unsworth, 1988)

The influence of CO2 on the water-use of crops may well prove to be the most important benefit of increased CO2 concentrations for agriculture. Morison(1993) reviewed the recent literature and discussed the effect at a range of scales, pointing out that there are now many studies, including annual cereal crops, legumes and woody perennials, which show that, contrary to earlier assumptions, although the absolute amount of dry matter produced decreases at all CO2 concentrations as water availability is reduced, the relative effect of increasing CO2 on plant growth increases with decreasing water supply. There are at least two mechanisms responsible for this relative enhancement: (1) the reduction in stomatal conductance, with a consequent beneficial effect on leaf water potential and hence on leaf expansion; (2) greater allocation of carbon to roots in elevated CO2, and hence improved potential for exploring the soil volume for water and nutrients. In contrast, the increased production of leaf area in elevated CO2 acts to increase the water-use per unit ground area of crops.
It is less clear whether increasing CO2 will decrease water-use of crop canopies in the field over periods of days or months. This is not only because of the various mechanisms described above, but also because soil evaporation must be accounted for, and feedbacks between leaf temperature, atmospheric humidity and canopy evaporation apply. Consequently, the very rough estimate that a 33% increase in dry matter production combined with a 33% decrease in transpiration in response to doubling CO2 concentration would increase water-use efficiency by a factor of 2 is unlikely to apply. Goudriaan and Unsworth (1990) used a crop model with feedbacks between vegetation and the atmosphere to estimate that daily plant canopy water-use efficiency would increase by about 35% when carbon dioxide concentration doubled. In a simpler model, but including soil evaporation, Morison (1993) calculated that seasonal water-use by crops with leaf area indices between 2 and 6 would decrease by only 10 to 15% in doubled CO2 concentration. Nijs et al. (1989) grew ryegrass in 350 m mol/mol CO2 and 600 m mol/mol CO2. While the total water-use of the canopy was unaffected by growth in elevated CO2, WUE was increased by 25% on a canopy basis or 87% on a leaf area basis. Results from this experiment indicated that, because the influence of elevated CO2 on PN and gs was greatest early in the season, before the canopy closed, WUE probably varied seasonally, and this may be a general feature for annual crops.
Azam Ali (pers. comm.) used two methods to assess transpiration of crops of sorghum and groundnut growing at 350 and 700 m mol/mol CO2. Seasonal monitoring of soil water content with a neutron probe, combined with metered irrigation and subsidiary measurements of soil evaporation, enabled cumulative crop transpiration to be calculated. Porometry on individual leaves, and measurements of leaf area distribution, enabled transpiration per unit leaf area to be calculated. In groundnut but not in sorghum, elevated CO2 reduced transpiration per unit leaf area throughout the season, but more so in the earlier stages. Figure 8.6 shows the seasonal relationships between dry mass and transpired water for the two crops; the slope is water-use efficiency (WUE). For the C3 groundnut, seasonal WUE was significantly larger at 700 m mol/mol CO2 (4.4 ± 0.2 g/kg) than for 350 m mol/mol CO2 (3.0 ± 0.2 g/kg); equivalent values for sorghum (4.9 ± 0.3 and 4.2 ± 0.3 g/kg, respectively) did not differ significantly. The figure also shows that, in spite of the increased WUE in groundnut, the total amount of water-used in the season was about the same (450 mm) in both CO2 treatments.
On an even larger scale, de Bruin and Jacob (1993) estimated the influence of doubling CO2 concentration on regional-scale transpiration, allowing for interactions between transpiration, leaf temperature and the planetary boundary layer. They concluded that, for vegetation where stomatal conductance decreased by 34% in response to elevated CO2, but leaf area was unchanged, regional transpiration on a typical summer day would decrease by about 11 % for short crops such as cultivated grass and 17% for tall crops such as plantation forests. If negative feedback from the planetary boundary layer had been ignored, these values would have been overestimated (15 and 24%, respectively).
When Krupa and Kickert (1989) attempted to review the interactive responses of crops to CO2 and O3 they found no publications describing experimental work. We summarize here the few papers that have been published since then, and comment on other work known to be in progress. We briefly review the effects of ozone alone later in this paper.
Allen (1990) made an estimate of the possible scale by which CO2 might influence responses of soybeans to O3. He used published results relating O3 exposure to yield (Heagle et al., 1983) and a model of the sensitivity of stomatal conductance for soybean to CO2 (Rogers et al., 1983). According to these models, doubling CO2 concentration would reduce gs by about 30% and thus decrease the stomatal uptake of O3 by the crop. Allen estimated that this would result in a yield decrease of about 15% compared with yield in ambient air. This estimate takes no account of the large direct effects of CO2 on soybean growth and assumes no interactions between the two gases.
Scientists at the USDA Beltsville Research Center have studied soybeans growing in open-top chambers in the field and exposed to CO2 and O3. Kramer et al. (1991), in a preliminary study, showed that the yield loss attributed to ozone at the site was about 12%, but when CO2 concentrations were increased to 500 m mol/mol the yield loss was only 6.7%. Mulchi et al. (1992) reported a more complex design with three O3 treatments (charcoal-filtered, CF, non-filtered, NF, NF + 40 m mol/mol O3) and three CO2 treatments (ambient, + 50, + 150 m mol/mol). Increasing CO2 concentration increased grain yield and grain oil content, and decreased protein content. Increasing O3 reduced grain yield but did not significantly alter oil or protein content. Table 8.2 summarizes the interactive effects of CO2 and O3 on growth, yield and quality. Ozone caused a significant decrease in yield compared to the charcoal-filtered controls. When CO2 was added to the ozone treatments, it partially counteracted the negative effect of O3 so that yields in chambers with plus 150 m mol/mol CO2 and NF air were similar to those in ambient CO2 and CF air. The results suggested that most effects of CO2 and O3 were additive.
Figure 8.6. (a) Seasonal relationship between dry mass and transpired water for crops of groundnut growing in controlled-environment glasshouses at 350 m mol/mol CO2 and 700 m mol/mol CO2. Regression lines are shown dotted for 350 m mol/mol and solid for 700 m mol/mol. The slopes of the regression line are the water-use efficiencies (WUEs).

Figure 8.6. (b) As 6(a) but for a crop of sorghum (Azam Ali, pers. comm.)

Barnes and Pfirrman (1992) grew radishes in a phytotron in Munich, Germany, at two CO2 concentrations (385 and 765 m mol/mol) and two O3 concentrations (20 nmol/mol and 73 nmol/mol), and studied gas exchange, growth and productivity, and mineral composition of the crop. In clean air the typical effects of elevated CO2 on photosynthesis (increased) and stomatal conductance (decreased) were observed; the extra carbon assimilation stimulated root growth rate by 43% but there was no significant effect on shoot growth or leaf area. At ambient CO2, the high O3 treatment depressed photosynthesis by 26% (compared to the low treatment) and induced a slight reduction in gs, with the net result that water-use efficiency declined. The reduction in carbon uptake was reflected in reduced growth, with roots more affected than shoots. Interactive effects of ozone and CO2 were generally consistent with a reduction in ozone-induced responses in elevated CO2. Early in the experiment, elevated CO2 reduced the effect of O3 on photosynthesis, but later this benefit disappeared, perhaps because long-term exposure to O3 has an overall limiting effect on plant response to CO2. The effects of CO2 and O3 on gs appeared to be additive. Work is in progress in Munich on responses of grass swards to O3, CO2 and water availability (Payer et al., 1993). Preliminary results indicate that the growth depression which is observed as water is withheld in ambient CO2 is alleviated at higher CO2 levels. Responses of yield and water-use efficiency to the mixtures of CO2 and ozone seem to be additive rather than synergistic.
In Newcastle, England, work is in progress exposing winter and spring cultivars of wheat to two CO2 concentrations and two O3 concentrations (Barnes et al., 1993). Early results indicate that after 50 days' exposure there was evidence of downregulation of photosynthesis in response to prolonged exposure to elevated CO2; this was most pronounced in the winter cultivars. Long-term exposure to O3 resulted in a decrease in the light-saturated rate of CO2 assimilation, partial stomatal closure, and accumulation of water soluble carbohydrate and starch in leaves. These resulted in decreased growth, with root growth more severely affected than shoot growth. In plants exposed to elevated CO2 and O3, effects on stomatal conductance were less than additive, and CO2 enhancement effects on photosynthesis and growth were reduced by O3.
In summary, from the very limited experimental results available at the time of writing, interactions between exposures to elevated O3 and CO2 seem to result in approximately additive effects on growth and yield. Both gases decrease stomatal conductance, and this results in less uptake of O3 in elevated CO2 treatments than in ambient air. Several authors have noted that this reduces (or at least delays) the damaging effects of O3 In principle, the reverse is also true, i.e., increased O3, by reducing stomatal conductance, negating some of the benefits to growth that elevated CO2 confers. However, the O3 exposure necessary to induce stomatal closure is generally sufficient to cause visible leaf injury, and this disbenefit is likely to be far more damaging than reduced CO2 uptake through stomata. More field-based studies are necessary to assess fully the effects of the two gases in combination.
Table 8.2. Direct and interactive effects of CO2 and O3 treatments on growth, yield components and grain quality characteristics of soybean growing in open-top field chambers (from Mulchi et al., 1992)
|
Chamber treatments |
Shoot biomass 1 |
Leaf area 1 |
SLW 1 |
Pods/plant |
Seeds/plant |
Weight of 100 seeds |
Grain yield |
Grain oil |
Grain protein |
|
|
CO2 |
O3 |
(g) |
(dm2) |
(mg/cm2) |
(g) |
(g) |
(%) |
(%) |
||
|
CO2 treatment means |
||||||||||
|
Ambient |
- |
55.3 |
55.7 |
2.88 |
51.7 |
128 |
16.9 |
436 |
20.4 |
40.4 |
|
+50 m mol/mol |
- |
61.1 |
58.1 |
3.07 |
57.8 |
140 |
17.7 |
497 |
22.3 |
39.3 |
|
+150 m mol/mol |
- |
59.2 |
58.0 |
2.99 |
60.9 |
144 |
17.6 |
509 |
22.1 |
39.2 |
|
LSD(0.05) |
|
3.2 |
NS |
NS |
7.4 |
11 |
0.5 |
41 |
0.5 |
1.1 |
|
O3 treatment means |
||||||||||
|
- |
CF |
58.9 |
60.1 |
2.73 |
63.7 |
153 |
18.2 |
558 |
21.9 |
39.4 |
|
- |
NF |
62.5 |
59.7 |
3.04 |
52.1 |
133 |
17.8 |
472 |
22.1 |
39.5 |
|
- |
NF+O3 |
54.3 |
52.0 |
3.17 |
54.6 |
127 |
16,2 |
412 |
21.7 |
40.1 |
|
LSD(0.05) |
3.2 |
7.5 |
0.36 |
7.4 |
11 |
0.5 |
0.5 |
NS |
NS |
|
|
CO2 and O3 treatment |
||||||||||
|
Ambient |
CF |
57.6 |
61.4 |
2.31 |
56.2 |
147 |
17.5 |
513 |
21.4 |
40.1 |
|
|
NF |
59.2 |
57.5 |
3.07 |
50.9 |
122 |
17.6 |
432 |
21.7 |
40.4 |
|
|
NF+O3 |
49.2 |
48.2 |
3.26 |
48.2 |
117 |
15.5 |
363 |
21.2 |
40.8 |
|
+50 m mol/mol |
CF |
60.4 |
61.8 |
2.99 |
66.6 |
155 |
19.1 |
594 |
22.1 |
39.6 |
|
|
NF |
66.7 |
58.0 |
3.12 |
50.6 |
135 |
17.7 |
474 |
22.9 |
38.6 |
|
|
NF+O3 |
56.4 |
54.4 |
3.11 |
55.8 |
130 |
16.3 |
424 |
22.1 |
39.7 |
|
+ 150 m mol/mol |
CF |
58.6 |
57.1 |
2.88 |
68.4 |
157 |
18.1 |
568 |
22.4 |
38.6 |
|
|
NF |
61.7 |
63.6 |
2.94 |
54.7 |
142 |
18.0 |
510 |
21.9 |
39.4 |
|
|
NF+O3 |
57.4 |
53.3 |
3.14 |
59.6 |
134 |
16.8 |
448 |
22.0 |
39.8 |
|
|
Av. |
58.6 |
57,3 |
2.98 |
56.8 |
137 |
17.4 |
480 |
21.9 |
39.6 |
|
LSD(0.05) |
|
5.7 |
NS |
0.36 |
NS |
NS |
0.9 |
53 |
NS |
NS |
|
CV (%) |
|
5.3 |
10.7 |
9.3 |
9.9 |
6.1 |
2.4 |
7.0 |
1.6 |
2.2 |
1 Vegetative samples were collected on 15 August 1989.
NS = not significant: LSD = Least significant difference; CV = Coefficient of variance
Many studies of effects of UV-B radiation on crops are seriously flawed because UV exposures were either inadequately specified and/or were unrealistically large. Tevini (1993), in a thorough review of effects of UV-B radiation on plants, pointed out that, since the spectrum of UV from artificial light sources differs from that in the solar spectrum, and since many photobiological processes are strongly wavelength dependent, it is important to use weighting functions based on the action spectrum for specific responses before experiments in different exposure systems can be compared. Unfortunately, action spectra for many responses remain unknown. In addition, many plant responses to UV-B are larger when plants are growing at low light levels typical of growth chambers and some glasshouses than they are in full sunshine, probably because natural protective pigments are inadequately synthesized and repair processes are restricted in low light. For this reason, field-based studies of crops are most relevant for estimating responses of productivity and yield to UV-B. Even so, most field studies have used continuous UV-B exposures corresponding to 15 to 25% reductions in stratospheric O3 i.e. roughly 30 to 50% increases in UV-B above present values. Such increases are much larger than are likely to occur as long-term means in agricultural regions under any likely depletion of stratospheric ozone in the next 30 years.
Effects of UV-B on yield of crops growing in the field have produced very variable results. Work in the late 1970s and early 1980s by Biggs and colleagues in Florida, using very high UV exposures, found yield reductions in only about half the crops studied. Work in Germany, also at high UV exposures, found no UV-B effects on cabbage, lettuce and rape. More recently, a large field-based study in Beltsville, Maryland, by Teramura, has revealed a high degree of intra-specific variability among soybean cultivars. The two most sensitive cultivars were grown for five seasons in the field in two UV treatments, corresponding to 16 and 25% ozone depletion simulations. In one cultivar (Essex) yield was reduced by 20 to 25% at the high UV treatment; in the other cultivar, yield generally increased by 10 to 22% in this treatment. Neither cultivar showed a consistent significant yield change at the lower UV exposure. The conclusion from these studies must be that, under any realistic scenario of increasing UV-B radiation, crop yields are unlikely to be altered to a significant extent. We therefore turn to the question of whether UV-B exposure alters the sensitivity of crops to other atmospheric changes.
Effects of the combination of elevated CO2 and UV-B radiation on crop growth and yield have been addressed in very few studies. Teramura and his colleagues (Teramura et al., 1990) grew wheat, rice and soybean in a glasshouse experiment. Treatments in the factorial design were: ambient CO2 (350 (m mol/mol), ambient UV, elevated CO2 (650 m mol/mol) and elevated UV-B (corresponding to a 10% decrease in stratospheric O3 at the equator). Compared to the control, seed yield and total biomass increased significantly in elevated CO2 for all three species. However, when UV-B and CO2 were increased simultaneously, no increase in either seed yield (wheat and rice) or total biomass (rice) was observed compared to the control. In contrast, in soybean, the increases in seed yield and biomass induced by CO2 alone were maintained in the high CO2/high UV-B environment. Studies of leaf gas exchange indicated that UV-B reduced the apparent carboxylation efficiency in wheat and rice, but not in soybean.
Results of a CO2/UV study on seedlings of sunflower and maize reported by Tevini (1993) are confounded by temperature changes in some treatments. A 25% increase in UV-B at 340 m mol/mol CO2 changed sunflower and maize dry weights (W) by -14 and -24%, respectively (compared to ambient controls). The same UV-B increment combined with a +2°C temperature change altered W by +5 % in sunflower and +31 % in maize. Adding a doubled CO2 concentration to the elevated UV and temperature regime altered W by +19 and +32% for sunflower and maize, respectively. The incomplete experimental design limits interpretation of this study, but the results suggest that temperature and CO2 are much more important influences on growth than any likely realistic increases in UV-B.
There are a number of recent reviews of the mechanism of action of O3 on plants, and of crop responses to O3 (e.g., Tingey and Andersen, 1991; Heck et al., 1988); consequently, we will only briefly summarize the main features. The response of plants to O3 may be viewed as the culmination of a sequence of physical, biochemical and physiological events. The O3 diffuses from the air into the leaf through stomata, and these exert control on O3 uptake. Plants are able to detoxify O3 or its metabolites and can repair or compensate for O3 impacts, so visible signs of ozone injury may not occur if the rate of O3 uptake is sufficiently small. When resources have to be devoted to repair or compensation mechanisms, or when O3 uptake is too large for full repair to be achieved, growth and yield may be reduced in the absence of visible injury by O3. The principal modes of action of O3 on plants are through injury to proteins and membranes, reduction in photosynthesis (i.e., carbon gained), changes in allocation of carbohydrate, and acceleration of senescence. The main result of O3 exposure is therefore a reduction in the capacity of the plant to accumulate photosynthate through loss in photosynthetic capacity and increased foliage senescence.
Methods for exposing crops to ozone in experiments designed to develop yield/exposure responses are well established. The most commonly used technique is the open-top field chamber, which enables crop stands to be grown in either charcoal-filtered air or in air to which known amounts of O3 have been added. Coordinated programmes in the United States and Europe have investigated crop responses to O3 in open-top chambers, and relationships between yield and seasonal mean O3 concentration are available for many of the major world crops. The seasonal mean index, however, has been shown to be inadequate in relating ozone exposure to effects, primarily because of its inability to consider duration of exposure and its consideration of all concentrations as equal in their effect. Both duration and concentration are important in the effect of ozone (Hogsett et al., 1988). In particular, studies have demonstrated that ozone's effect is cumulative and that higher concentrations are more important than lower concentrations in causing an effect (Hogsett et al., 1988; Musselman et al., 1994). Consequently, the cumulative, peak-weighted indices are more appropriate because of their biological relevancy. Examples of such indices include SUM06 (in units of nmol/mol-h) which cumulates all concentrations equal to or greater than 0.06 nmol/mol, or SIGMOID which cumulates all concentrations during the season and weights all concentrations according to a sigmoid function. The US Environmental Protection Agency (EPA) Draft Ozone Criteria Document (1995) reviewed and tabulated the latest information concerning effects of ozone on crop yields using both the seasonal mean, and two of the cumulative, peak-weighted indices. The data for crop yield response are primarily from the National Crop Loss Assessment Network (NCLAN). Yield reductions in the NCLAN studies are calculated relative to a typical background O3 concentration of SUM06 of O3 or a 7-h seasonal mean of 25 nmol/mol. Yield losses of 10% or less in 50% of the crops (12 crop species, 54 studies including well-watered and drought studies) would occur with SUM06 exposure concentrations of 26.4 nmol/mol-h in three months, or 49 nmol/mol 7-h seasonal mean. Some species/cultivars were particularly sensitive; 11 % would be expected to have a yield reduction of 10% at a 7-hr seasonal mean of less than 35 nmol/mol. Similarly, 18% of the crops/cultivars are expected to have yield reductions of 10% or less at 3-month SUM06 concentrations of less than 10 nmol/mol-h. Both of these exposure values are quite low compared to the decade-average (1982-1991) exposure (3 months) from all monitoring sites across the United States (SUM06 = 29.5 nmol/mol-h; 7-h mean = 54 nmol/mol). In general, grain crops are less sensitive than others, but within-species variability and sensitivity may be greater than between species. Krupa and Kickert (1989) reviewed various published lists of the sensitivity of crop yields to O3 onion, spinach, potato, alfalfa and cotton appear to be particularly sensitive crops, although it is possible to find resistant cultivars of all these species. Legumes range between sensitive and intermediate, and cereal and grass crops tend to be relatively resistant.
There is also a very large literature describing biotic and abiotic factors that modify plant response to 0. (US EPA, 1995). We will concentrate in this paper on only a few interacting factors that are particularly relevant in terms of future changes in the atmospheric environment.
When interactions between ozone exposure and temperature were reviewed by the US EPA (1986) it was concluded that there were variable and conflicting results. More recent analysis (US EPA, 1995) suggests that many of these studies in controlled environments confounded effects of temperature with changes in vapour pressure deficit (VPD) because relative humidity was kept constant while temperature was increased. Since VPD can have profound effects on stomatal responses, evaporation rates and leaf expansion, it is not surprising that these early results have led to confusion. Recently, Todd et al. (1991) designed an experiment with tomato seedlings in which differences in VPD at different temperatures were minimized during O3 exposure. This study showed that, out of 11 growth variables measured, the only significant modifications by temperature of the effects of O3 were on stem fresh weight and specific leaf area (leaf area/leaf dry weight). The authors suggested that VPD probably plays a more important role in determining sensitivity to O3 than temperature. It is important to have more studies to clarify this issue, as many field-based studies of ozone responses use open-top chambers in which temperatures are increased by a few degrees above ambient. If either the changed temperature or the changed VPD in such chambers alters the sensitivity of the crops to O3 there would be serious consequences for the application of much of our knowledge of the O3 sensitivity of crops that has been gained from field-chamber experiments.
A second important O3/temperature interaction, particularly affecting perennial species, including trees, is winter hardiness. Several studies have shown that exposures to O3 reduce the frost-hardiness; Davison et al. (1988) reviewed the subject. Whilst most of this research has been with woody species, Barnes et al. (1988) showed that daily exposures to about 80 nmol/mol O3 for 7 days significantly reduced the survival of pea seedlings after exposure to nighttime temperatures of about -4°C. It seems likely that, as with sulphur dioxide, exposure to ozone could alter the frost hardiness of a number of agricultural crops. This response would be most likely to be significant in the event of an early autumn frost on early-sown crops such as winter cereals. There are suggestions that exposure of trees in summer to ozone alters the hardiness the following winter. Eamus and Murray (1991) pointed out that, even in severe winters, there are brief periods of mild temperatures that induce partial dehardening of woody species. They speculated that O3 increases tree predisposition to dehardening in such conditions and consequently puts trees at greater risk from low temperatures. There are likely to be significant differences between species in this phenomenon. For example, in Florida, Eissenstat et al. (1991) found that, although O3 reduced frost hardiness of citrus and avocado, the effects were small and the likelihood of significantly changing frost resistance is slight.
The availability of soil water is one of the strongest influences on crop growth and productivity. When available, irrigation is commonly used to alleviate water stress, but in conditions of large evaporative demand, even irrigated crops may experience water stress. The most immediate effects of water stress are on plant-water potential, and may lead to reductions in leaf expansion rates and altered partitioning of dry matter, with increased assimilate partitioning to roots. When stomatal closure occurs, there is a resulting reduction in transpiration and CO2 uptake and an increase in leaf temperature.
WATER STRESS AND O3 RESPONSES
There has long been a general belief that water stress reduces the magnitude of any adverse effects of O3 i.e., leaf injury, growth and yield reductions. This belief was based on observations that stomatal closure, induced by drought, reduced the rate and quantity of O3 absorbed by leaves (US EPA, 1986). However, more recent results show that the O3/water availability interaction is complex, and probably depends on the magnitude and timing of the water stress (which has seldom been adequately defined in air pollution experiments). One difficulty in quantifying the interaction is the very large effect that water stress alone may have on yield, so that any influence of O3 is sometimes masked.
Temple et al. (1985) studied the effects of O3 and water stress on cotton growing in California. In this typically hot, dry season, the water-stressed plants wilted frequently, and yielded much less than the well-watered controls; nevertheless, when compared to yields of plants in a background O3 concentration of 25 nmol/mol, and expressed as a percentage, the estimated yield loss at a seasonal mean 0, concentration of 50 nmol/mol was 7% in the well-watered plants and 2% in the water-stressed crop. Clearly the absolute yield loss from drought was much larger than the yield loss attributed to O3 in either treatment.
Heagle et al. (1983) reviewed six studies of soybean responses to O3 and water stress, and concluded that in only three were there significant interactions, i.e., the clear negative relationships between yield and O3 exposure observed with well-watered plants were much reduced with water stressed plants. More recently, Heggestad and Lesser (1990) analysed three years of data for four soybean cultivars. They concluded that, in most cases, the relationships between yield and O3 concentration had similar slopes for the water-stressed and well-watered treatments. If this conclusion applies more generally, the extra yield losses which would result from O3 exposure could be estimated if a yield response to drought alone is known. Clearly, there is still uncertainty over the influence of water stress on the form of the yield response to O3. This uncertainty is unlikely to be resolved until better experimental designs with more precise specifications of the degree of water stress are developed.
O3 AND WATER-USE EFFICIENCY
For irrigated crops where water supplies may be limited or expensive, it is important to know whether O3 would influence the water-use efficiency (WUE) and consequently the amount of water required in a season. Reich et al. (1985) exposed well-watered soybeans to 130 nmol/mol O3 for 7 hours daily, and found a 25% decrease in WUE compared with controls in 10 nmol/mol O3. Similar results were found for alfalfa (Temple et al., 1988), probably indicating that carbon dioxide uptake was reduced more by O3 than water loss. Tingey et al. (1994) also reported a significant decrease in WUE in soybeans exposed to episodic exposure regimes of 20-30 nmol/mol-h (3-month SUM06). The study found the O3 did not close stomata, but rather affected CO2 assimilation. A significant increase in leaf construction cost was reported, indicating that both carbon and water are used inefficiently by plants exposed to ozone. In contrast, Greitner and Winner (1988) found that O3 exposure increased the WUE of radish and soybeans. This disagreement probably reflects variation with exposure dynamics, stage of development, and genetic variation between and within species.
Recently Barnes and Pfirrman (1992) investigated effects of O3 and elevated CO2 singly and in combination on the WUE of radish. Both gases reduced stomatal conductance, and the combination of gases reduced conductance still further. Figure 8.7 shows that O3 reduced the instantaneous WUE, whereas CO2 increased it (as discussed earlier). In the combination of O3 and CO2, there was initially no significant effect of O3 (i.e., the response of WUE was similar to that observed in elevated CO2), but as growth progressed, the WUE was significantly reduced in the combination treatment compared to in elevated CO2 alone. This may be a response to accelerated senescence induced by O3 exposure.
In summary, relatively severe water stress may reduce the yield losses attributable to O3 in some crop species but, in general, the yield loss resulting from the water stress outweighs the benefits of the O3 protection. For well-watered crops, O3 exposure may change the water-use efficiency compared with crops in clean air, but the direction and magnitude of change probably depend on exposure and genetic factors. The influence of O3 and CO2 in combination on WUE is potentially important and merits further study.
Krupa and Kickert (1989) found no reports of O3/UV-B interactions in their review. They assessed potential risks on a geographical basis, using distributions of major crops, tropospheric ozone concentrations and UV-B irradiance. They suggested that interactions might involve episodic exposure to the two stresses, so that peaks of O3 would coincide with lower UV-B irradiance and vice versa. Runeckles and Krupa (1994) have developed this concept further, arguing that, as tropospheric O3 increases, UV-B irradiance at the surface is reduced, because the O3 absorbs some UV. In theory, this effect could be important, in spite of the relatively small contribution of tropospheric O3 to the total atmospheric O3 column, because scattering by aerosols and molecules in the troposphere increases the radiation path length (Bruhl and Crutzen, 1989). In an attempt to quantify this effect, Albar (1992) compared the measured solar spectral irradiance I (1) at the ground near Nottingham, England, on two days (12 and 18 July 1990) when the tropospheric O3 concentration was 51 and 84 nmol/mol, respectively, and stratospheric O3 was constant. Figure 8.8 shows that I (1) was about 20 to 40% greater in the UV-B waveband (280-320 nm) on the low O3 day than on the high O3 day, apparently supporting the hypothesis of Runeckles and Krupa (1994). However I (1) at longer wavelengths was also greater (by about 20 to 25%), and this increase is most probably a consequence of less aerosol (dust) being present on the low O3 day. Since this aerosol effect would also apply in the UV-B waveband, it seems likely that the UV-B increase directly attributable to reduced tropospheric O3 was no more than 10% to 15%. Variations in aerosol from day to day seem likely to be more important than variations in tropospheric O3 in modulating the intensity of UV-B at the ground, but further observations would be valuable.
Figure 8.7. Instantaneous water-use efficiency WUE (m mol CO2 assimilated (A) per mol HO transpired (E) for radish (Raphanus sativus L. cv. Cherry Belle) plants exposed in climatic chambers to CO2 and O3 Treatments were 350 m mol/mol CO2 + 20 nmol/mol O3 (first columns) or + 80 nmol/mol O3 (second columns); 750 m mol/mol CO2 + 20 nmol/mol O3 (third columns) or + 80 nmol/mol O3 (fourth columns). Error bars show lsd at p=0.01 for each of the sampling times (from Barnes and Pfirrman, 1992)

Figure 8.8. Variation of percentage increase in solar spectral irradiance ((I(l, 51)-I (l , 84))/I (l, 84)) with wavelength l between days when the tropospheric ozone concentration was 84 nmol/mol O3 and 51 nmol/mol O3 (from Albar, 1992)

As mentioned earlier, many investigations of plant responses to UV-B are seriously flawed by inadequate specification and/or excessive UV exposure. An improved field-based system at Raleigh, North Carolina, using open-top chambers for studying crop responses to UV and O3 (Booker et al., 1992a) has been used for three seasons of research on soybeans (Miller et al., 1994). The system exposed soybean crops from emergence to maturity to UV treatments (ranging from ambient to about twice ambient biologically effective UV-B) and to O3 treatments giving seasonal mean 12 h/d O3 concentrations from 14 to 83 nmol/mol.
The ozone treatments resulted in reductions of photosynthesis, accelerated senescence, and reduced yield, in agreement with many other published studies. In contrast, the UV treatments, even at this relatively large UV irradiance, did not induce any significant changes in photosynthesis or yield, and there were no UV/O3 interactions (Booker et al., 1992b).
Fiscus et al. (1994) attempted to reconcile the lack of response to UV reported in the Raleigh experiments with other reports which indicate that increased UV-B causes physiological dysfunction and reductions in crop yields. They concluded that three factors make the conclusions of other field and glasshouse studies, at best, hard to interpret and possibly misleading: a frequent failure to monitor UV-B adequately; a tendency to underestimate UV exposures when relying on model calculations which do not allow sufficiently for effects of dust in the atmosphere; and no adjustment of UV exposure for seasonal and daily weather changes (and hence a tendency towards unrealistically large exposures).
Although it would be useful to see further well-designed field studies of UV/O3 interactions using a wide range of crop and natural species, on the evidence of the careful work of Booker, Miller, Fiscus and their colleagues, it seems unlikely that UV/O3 interactions are of any importance for crop productivity, and it seems clear that O3 poses a much greater threat to yields of many crops than any likely increases in UV-B radiation.
In our introduction we emphasized the need for knowledge of how interacting environmental changes will affect crop productivity. Many examples in this paper show that misleading conclusions can be drawn by considering crop responses to single factors. However, the experimental demands of studying multiple factors are great, and it is necessary to define priorities. In our view, the following interactions should have high priority:
· Interactions of CO2 and temperature. If the benefits of increased CO2 in terms of increased potential carbon assimilation by C3 species are not achievable at low temperatures, there are important implications for agriculture in cool climates. Equally, the merits of C3 versus C4 crops may need reassessing in warmer climates in view of the direct benefits of CO2 to C3 species.· Canopy scale studies of effects of CO2 on water-use efficiency, and on the interacting effects of O3. Although it is clear that increased CO2 enhances WUE at the leaf or single plant level, there are very few studies relevant to field-grown crops. Equally, the very limited studies of effects of O3 and CO2 on WUE suggest that responses are non-additive, and this is an important issue to resolve for agriculture in Europe, the eastern United States and other regions where tropospheric O3 concentrations influence crop productivity.
It is disappointing that, in spite of considerable interest in the effects of UV-B radiation on plants, much of the research in the past decade has been seriously flawed by employing very unrealistic UV-B exposures and/or by using highly artificial growth conditions. On the evidence of the few well-designed studies of crop responses to UV-B that we have reviewed here, we do not believe that UV-B poses serious threats to crops, and have not identified any important interactions between UV-B and other stressors that would make further research of this type a particularly high priority.
We are grateful to Ossama Albar, Sayed Azam Ali, Fitz Booker, Zahide Kocabas and their colleagues for allowing us access to unpublished results, and we thank Pat Sommer for her assistance in preparing the typescript.
Albar, O.F. 1992. The Spectral Distribution of Solar Ultraviolet Radiation at the Ground. PhD Thesis. University of Nottingham, UK.
Allen, L.H. 1990. Plant responses to rising carbon dioxide and potential interactions with air pollutants. J. Environmental Quality 19: 15-34.
Barnes, J.D. and Pfirrman, T. 1992. The influence of CO2 and O3 singly and in combination, on gas exchange, growth and nutrient status of radish (Raphanus sativus L.). New Phytologist 121: 403-412.
Barnes, J.D., Reiling, K., Davison, A.W. and Renner, C.J. 1988. Interaction between ozone and winter stress. Environmental Pollution 53: 235-254.
Barnes, J.D., Bannister, G. and Ollerenshaw, J. 1993. Interactive effects of CO2 and/or O3 on wheat (Triticum aestivum L.). UK Committee for Air Pollution Effects Research Annual Meeting, Bangor, Wales (Abstract).
Booker, F.L., Fiscus, E.L., Philbeck, R.B., Heagle, A.S., Miller, J.E. and Heck, W.W. 1992a. A supplemental ultraviolet-B radiation system for open-top field chambers. J. Environmental Quality 21: 56-61.
Booker, F.L., Miller, J.E. and Fiscus, E.L. 1992b. Effects of ozone and UV-B radiation on pigments, biomass and peroxidase activity in soybean. In: Tropospheric Ozone and the Environment: II. Effects, Modelling and Control. R.L. Berglund (ed.). Air and Waste Management Association, Pittsburgh, PA. pp. 489-503.
Bruhl, C. and Crutzen, P.J. 1989. On the disproportionate role of tropospheric ozone as a filter against solar UV-B radiation. Geophysics Research Letters 16 (7): 703-706.
Cannell, M.G.R. and Smith, R.I. 1986. Climatic warming, spring budburst and frost damage on trees. J. Applied Ecology 23: 177-191.
Cannell, M.G.R., Grace, J. and Booth, A. 1989. Possible impacts of climatic warming on trees and forests in the United Kingdom: a review. Forestry 62: 337-364.
Clifford, S.C., Stronach, I.M., Mohamed, A.D., Azam-Ali, S.N. and Crout, N.M.J. 1993. The effects of elevated atmospheric carbon dioxide and water stress on light interception, dry matter production and yield in stands of groundnut (Arachis hypogaea L.). J. Experimental Botany 44: 1763-1770.
Davison, A.W., Barnes, J.D. and Renner, C.J. 1988. Interactions between air pollutants and cold stress. In: Gaseous Air Pollution and Plant Metabolism. S. Schulte-Hostede, N.M. Darrall, L.W. Blank and A.R. Wellburn (eds.). Elsevier Science, London. pp. 307-328.
de Bruin, H.A.R. and Jacob, C.M.J. 1993. Impact of CO2 enrichment on the regional evapotranspiration of agro-ecosystems, a theoretical and numerical modelling study. In: CO2 and Biosphere. J. Rozema, H. Lambers, S.C. van de Geijn and M.L. Cambridge (eds.). Kluwer Academic Publishers, Dordrecht. pp. 307-318.
Drake, B.G. and Leadley, P.W. 1991. Canopy photosynthesis of crops and native plant communities exposed to long-term elevated CO2 treatment. Plant, Cell and Environment 14: 853-860.
Eamus, D. 1991. The interaction of rising CO2 and temperatures with water-use efficiency. Plant, Cell and Environment 14: 843-852.
Eamus, D. and Murray, M. 1991. Photosynthetic and stomatal conductance responses of Norway Spruce and Beech to ozone acid mist and frost in a conceptual model. Environmental Pollution 72: 23-44.
Eissenstat, D.M., Syverson, J.P, Dean, T.J., Yelenosky, G. and Johnson, J.D. 1991. Sensitivity of frost resistance and growth in citrus and avocado to chronic ozone exposure. New Phytologist 118: 139-146.
Fiscus, E.L., Miller, J.E. and Booker, F.L. 1994. Is UV-B a hazard to soybean photosynthesis and yield?: Results of an ozone/UV-B interaction study and model predictions. In: Stratospheric Ozone Depletion/UV-B Radiation in the Biosphere. R.H. Biggs and M.E.B. Joyner (eds.). Springer-Verlag, Berlin. pp. 135-147.
Goudriaan, J. and Unsworth, M.H. 1990. Implications of increasing carbon dioxide and climate change for agricultural productivity and water resources. In: Impact of Carbon Dioxide, Trace Gases, and Climate Change on Global Agriculture. B.A. Kimball, N.J. Rosenberg and L.H. Allen, Jr. (eds.). ASA Spec. Pub. No. 53. American Society of Agronomy, Madison, Wisconsin. pp. 111-133.
Greitner, C.S. and Winner, WE. 1988. Increases in delta 13C values of radish and soybean plants caused by ozone. New Phytologist 108: 489-494.
Heagle, A.S., Heck, W.W., Rawlings, J.O. and Philbeck, R.B. 1983. Effects of chronic doses of ozone and sulfur dioxide on injury and yield of soybeans in open-top field chambers. Crop Science 23: 1184-1191.
Heck, W.W., Taylor, O.C. and Tingey, D.T. 1988. Assessment of Crop Loss from Air Pollutants. Elsevier Applied Science, London.
Heggestad, H.E. and Lesser, V.M. 1990. Effects of ozone, sulfur dioxide, soil water deficit and cultivar on yields of soybean. J. Environmental Quality 19: 488-495.
Hogsett, W.E., Tingey, D.T. and Lee, E.H. 1988. Ozone exposure indices: concepts for development and evaluation of their use. In: Assessment of Crop Loss from Air Pollutants. W.W. Heck, O.C. Taylor and D.T. Tingey (eds.). Elsevier and Applied Science, London. pp. 107-138.
Houghton, J.T., Callander, B.A. and Varney, S.K. 1992. Climate Change 1992. Cambridge University Press, Cambridge.
Idso, S.B. 1990. Interactive effects of carbon dioxide and climate variables on plant growth. In: Impact of Carbon Dioxide, Trace Gases, and Climate Change on Global Agriculture. B.A. Kimball, N.J. Rosenberg and L.H. Allen Jr. (eds.). ASA Spec. Pub. No. 53. American Society of Agronomy, Madison, Wisconsin. pp. 61-69.
Kimball, B.A. 1983. Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agronomy J. 75: 779-788.
Kimball, B.A., Mauney, J.R., Nakayama, F.S. and Idso, S.B. 1993. Effects of increasing atmospheric CO2 on vegetation. In: CO2 and Biosphere. J. Rozema, H. Lambers, S.C. van de Geijn and M.L. Cambridge (eds.). Kluwer Academic Publishers, Dordrecht. pp. 65-75.
Kocabas, Z. 1993. Sensitivity of Crop Models to Climate Variables and Modelling Techniques. University of Nottingham. PhD.
Kocabas, Z., Mitchell, R.A.C., Craigon, J. and Perry, J.N. 1993. Sensitivity analyses of AFRCWHEAT1 crop model: the effect of changes in radiation and temperature. J. Agric. Sci. (Cambridge) 120: 149-158.
Kramer, G.F, Lee, E.H. and Rowland, R.A. 1991. Effects of elevated CO2 concentration on the polyamine levels of field-grown soybean at three O3 regimes. Environmental Pollution 73 (2): 137-152.
Krupa, S.V. and Kickert, R.N. 1989. The greenhouse effect: impacts of ultraviolet-B (UV-B) radiation, carbon dioxide, and ozone on vegetation. Environmental Pollution 61: 263-393.
Lawlor, D.W. and Mitchell, R.A.C. 1991. The effects of increasing CO2 on crop photosynthesis and productivity: a review of field studies. Plant, Cell and Environment 14: 807-818.
Long, S.P. 1991. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations: has its importance been underestimated? Plant, Cell and Environment 14: 729-739.
Miller, J.E., Booker, F.L., Fiscus, E.L., Heagle, A.S., Pursley, W.A., Vozzo, S.F. and Heck, W.W. 1994. Ultraviolet-B radiation and ozone effects on growth, yield, and photosynthesis of soybean. J. Environmental Quality 23 (1): 83-91.
Morison, J.I.L. 1985. Sensitivity of stomata and water-use efficiency to high CO2. Plant, Cell and Environment 8: 467-474.
Morison, J.I.L. 1993. Response of plants to CO2 under water limited conditions. In: CO2 and Biosphere. J. Rozema, H. Lambers, S.C. van de Geijn and M.L. Cambridge (eds.). Kluwer Academic Publishers, Dordrecht. pp. 193-209.
Mulchi, C.L., Slaughter, L., Saleem, M., Lee, E.H., Pausch, R. and Rowland, R. 1992. Growth and physiological characteristics of soybean in open-top chambers in response to ozone and increased atmospheric CO2. Agriculture, Ecosystems and Environment 38: 107-118.
Musselman, R.C., McCool, P.M. and Lefohn, A.S. 1994. Ozone descriptors for an air quality standard to protect vegetation. Journal of the Air and Waste Management Association. 44: 1383-1390.
Nijs, I., Impens, I. and Behaeghe, T. 1989. Leaf and canopy responses of Lolium perenne to long-term elevated atmospheric carbon dioxide concentration. Planta 177: 312-320.
Oechel, W.C. and Strain, B.R. 1985. Native species responses to increased atmospheric carbon dioxide concentration. In: Direct Effects of Increasing Carbon Dioxide on Vegetation. B.R. Strain and J.D. Cure (eds.). US Department of Energy, Washington DC. pp. 119-154.
Payer, H., Firsching, K., Steiner, K. and Djelassi, K. 1993. Factorial effects of CO2, O3 and irrigation on the yield and water-use efficiency of experimental grass swards grown in closed chambers. UK Committee for Air Pollution Effects Research Annual Meeting, Bangor, Wales (Abstract).
Porter, J. 1984. A model of canopy development in winter wheat. J. Agric. Sci. (Cambridge) 102: 383-392.
Reich, P.B., Schoettle, A.W. and Amundson, R.G. 1985. Effects of low concentrations of O3 leaf age and water stress on leaf diffusive conductance and water-use efficiency in soybean. Physiologia Plantarum 63: 58-64.
Rogers, H.H., Bingham, G.E., Cure, J.D., Smith, J.M. and Surano, K.A. 1983. Responses of selected plant species to elevated carbon dioxide in the field. J. Environmental Quality 12: 569-574.
Runeckles, V.C. and Krupa, S.V. 1994. The impact of UV-B radiation and ozone on terrestrial vegetation. Environmental Pollution 83: 191-213.
Squire, G.J. and Unsworth, M.H. 1988. Effects of CO2 and Climate Change on Agriculture. Report to U.K. Department of the Environment. University of Nottingham, England.
Temple, P.J., Taylor, O.C. and Benoit, L.F. 1985. Cotton yield responses to ozone as mediated by soil moisture and evapotranspiration. J. Environmental Quality 14: 55-60.
Temple, P.J., Benoit, L.F., Lennox, R.W., Reagan, C.A. and Taylor, O.C. 1988. Combined effects of ozone and water stress on alfalfa growth and yield. J. Environmental Quality 17: 108-113.
Teramura, A.H., Sullivan, J.H. and Ziska, L.H. 1990. Interaction of elevated ultraviolet-B radiation and CO2 on productivity and photosynthetic characteristics in wheat, rice, and soybean. Plant Physiology 94: 470-475.
Tevini, M. 1993. UV-B Radiation and Ozone Depletion: Effects on Humans, Animals, Plants, Microorganisms, and Materials. Lewis, Boca Raton, Florida.
Tingey, D.T. and Andersen, C.P. 1991. The physiological basis of differential plant sensitivity to changes in atmospheric quality. In: Ecological Genetics and Air Pollution. G.E. Taylor, J.L.F. Pitelka and M.T. Clegg (eds.). Springer-Verlag, Berlin. pp. 209-235.
Tingey, D.T. and Taylor, G.E.J. 1982. Variation in plant response to ozone: a conceptual model of physiological events. In: Effects of Gaseous Air Pollution in Agriculture and Horticulture. M.H. Unsworth and D.P. Ormrod (eds.). Butterworth Scientific, London. pp. 113-138.
Tingey, D.T., Hogsett, W.E., Rodecap, K.D., Lee, E.H. and Moser, T.J. 1994. The impact of O3 on leaf construction cost and carbon isotope discrimination. In: Immissionsokologische Forschun im Wnadel der Zeit: Festschrift fur Robert Guderian. W. Kuttler and M. Jochimsen (eds.). Westarp Wissenschaften: Essener Okologische Schriften. pp. 195-206.
Todd, A.G., Ormrod, D.P., Hale, B.A. and Goodyear, S.N. 1991. Temperature effects on tomato response to ozone at constant vapor pressure deficit. Biotronics 20: 43-52.
US EPA. 1986. Effects of ozone and other photochemical oxidants on vegetation. In: Air Quality Criteria for Ozone and Other Photochemical Oxidants. Research Triangle Park, North Carolina. Environmental Criteria and Assessment Office. pp. 6/1-6/298.
US EPA. 1995. Air Quality Criteria for Ozone and other Photochemical Oxidants. Draft edition. Research Triangle Park, North Carolina. Environmental Criteria and Assessment Office.
van de Geijn, S.C., Goudriaan, J. and Berendse, F. (eds.). 1993. Climate Change; Crops and Terrestrial Ecosystems. CABO-DLO, Wageningen. 144 p.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}