![]()
![]()
![]()
Objective and analysed information
In an evolutionary biology context, adaptation is a process, adaptedness is the level of adaptation of plant material to a given environment, and adaptability is the ability to show good adaptedness in a wide range of environments (Tigerstedt, 1994). In a plant breeding context, the first two terms relate to a condition rather than a process, indicating the ability of the material to be high-yielding with respect to a given environment or given conditions (to which it is adapted) (Gallais, 1992; Cooper and Byth, 1996). In breeding for wide adaptation (i.e. adaptability), the aim is to obtain a variety which performs well in nearly all environments; in breeding for specific adaptation, the aim is to obtain a variety which performs well in a definite subset of environments within a target region. The adaptive response of a variety is assessed with respect to other genotypes and tends to undergo modification when better-performing germplasm becomes available.
Breeding for wide adaptation and for high yield stability and reliability have sometimes been considered one and the same, insofar as the latter two terms indicate a consistently good yield response across environments. Some authors, however, have applied the yield stability concept with respect to consistency in time of genotype performance, using the adaptation concept in relation to consistency in space (Barah et al., 1981; Lin and Binns, 1988; Evans, 1993). It has also been widely acknowledged (Ghaderi et al., 1980; Becker, 1984; Lin and Butler, 1988; Bowman, 1989; Annicchiarico, 1992, 1997b; Romagosa and Fox, 1993; Piepho et al., 1998) that only genotype × location (GL) interaction, rather than all kinds of GE interaction, is useful for depicting adaptation patterns, as only this interaction can be exploited by selecting for specific adaptation or by growing specifically adapted genotypes. For example, the knowledge of specific adaptation to past years, as shown by positive genotype × year (GY) interaction effects, cannot be exploited in future years, since the climatic conditions that generate year-to-year environmental variation are not known in advance. This view implies that analysis of adaptation - and its implications for the definition of adaptation strategies for breeding programmes and domains of cultivar recommendation for extension services - may concern only responses to locations, geographic areas, farming practices or other factors that can be controlled or predicted prior to sowing. In particular, the analysis of multi-environment yield trials should focus primarily on GL interaction, with the characteristics of the locations depending on climatic, soil, biotic (pests and diseases) and crop management factors. The remaining interactions of genotype with the time factor (year for annual crop, crop cycle for perennials) should be dealt with in terms of yield stability. In some cases, the concept and the analysis of adaptation may concern the genotype responses to a set of management practices that have a crucial impact on GE effects, rather than the responses to locations (e.g. Annicchiarico and Piano, 1994).
The above definition of GL interaction implies that only GL effects repeatable in time are of practical importance. These effects can be either:
exploited, by growing specifically adapted germplasm; or
minimized, by growing widely adapted material.
Non-repeatable GL effects contribute to the second-order interaction (genotype × location × year [GLY] for annuals, and genotype × location × crop cycle for perennials). They represent the error term for (repeatable) GL interaction in analysis of variance (ANOVA) models where the time factor (considered random) is crossed with the location factor. In ANOVA models holding the time factor nested into location, non-repeatable GL effects are included in the GY interaction within site, which acts as the error term for GL interaction (see Section 4.1). The size of the GL effects is also relevant: in particular, if the GL interaction variance component (although statistically significant) is small compared to other components, particularly the genotypic one, it reduces the possible advantage of breeding for specific adaptation.
For annual crops in particular, multilocation trials should be repeated in time to distinguish between repeatable and non-repeatable GL interaction effects. A number of reports from various countries (Talbot, 1984; Léon and Becker, 1988; Atlin and McRae, 1994; Weber and Westermann, 1994; Sneller and Dombek, 1995; Annicchiarico, 1997a), providing estimates of variance components or investigating the consistency of GL interaction patterns across individual years, have shown that a reliable assessment of GL effects is not possible with just one year’s data, because the estimation is inflated by non-repeatable effects. The lack of repeatability is due mainly to the year-to-year variation in climatic factors within locations.
The emphasis on GL interaction effects is justified even when analysis of adaptation relates directly to genotype responses to environmental factors (as with the statistical models considered in Section 5.4). Specifically adapted genotypes can be targeted to respond well under the environmental conditions prevailing in a given area, provided that these conditions are not highly variable from year to year. Important information may not be obtained if the focus is on GE rather than GL effects, because the impact on the GL interaction of an environmental factor that strongly affects the GE interaction (impact which determines the scope for specific adaptation) may be large or limited, depending on the extent of the between-location variation relative to the within-location, year-to year variation for the factor (Fig. 2.1). For example, genotype variation in response to rainfall may appear important when analysing GE interaction and negligible when analysing GL interaction, when test locations have similar mean value and large year-to-year variation for the environmental variable.
By concentrating on GL interaction in analysis of adaptation, the process is also simplified, because adaptation patterns which are remarkably complex when evaluated on a GE basis (requiring three or more dimensions for a convenient multivariate representation) become relatively simple on a GL basis (requiring only one or two dimensions), mainly as a consequence of the larger size of the appropriate error term. This is shown in Table 2.1 for different data sets, in which the number of dimensions are represented by the number of significant GE or GL interaction principal component axes (according to an analytical model described in Section 5.3). Reducing the number of dimensions has practical implications, because one- or two-dimensional models offer additional and simplified procedures for the identification of best-yielding material for variety recommendation and to help breeding programmes define adaptation strategies (see Sections 5.3, 5.4 and 5.8).
While earlier studies on GE interaction concentrated on GL interaction in the assessment of genotype adaptation and zoning of locations (Yates and Cochran, 1938; Horner and Frey, 1957; Abou-El-Fittouh et al., 1969), recent developments and applications of statistical methods have frequently focused on the interaction of genotypes with generic environments (Crossa et al., 1995; DeLacy et al., 1996a; Gauch and Zobel, 1997). However, most methods proposed for GE interaction analysis can easily be adapted to the investigation of GL effects.
TABLE 2.1 - Complexity of adaptation patterns as depicted by the number of significant PC axes in the analysis of GE and GL data matrices of six data sets
| Data set |
No. sites |
No. years |
No. genotypes |
Significant PC axesa |
|
| GE |
GL |
||||
| Bread wheat |
31 |
3 |
18 |
5 |
2 |
| Durum wheat 1 |
6 |
3 |
9 |
4 |
2 |
| Durum wheat 2 |
5 |
2 |
15 |
4 |
1 |
| Durum wheat 3 |
6 |
2 |
12 |
4 |
1 |
| Maize 1 |
11 |
3 |
13 |
4 |
0 |
| Maize 2 |
11 |
3 |
11 |
3 |
1 |
a P < 0.01 according to FGH2 test (Cornelius, 1993), in an AMMI analysis.
Source: Annicchiarico, 1997a.
Subregions
Adaptation patterns with respect to individual locations are of limited interest per se, as the sample of sites is very small compared to the large number of locations in any target region. Specific breeding, in particular, can only be directed to areas and cannot realistically be so fine-tuned as to exploit positive interaction effects of genotypes with individual locations. However, sites that are similar in terms of genotype response can be grouped by different methods (discussed in Chapter 5), and each group may identify a cropping area that is relatively uniform because GL interaction effects are limited or negligible. Such areas (possibly the object of specific breeding) have been termed by different authors as subregions, subzones, subareas, macro-environments or mega-environments (Horner and Frey, 1957; Seif et al., 1979; Ceccarelli, 1989; CIMMYT, 1989). Subregions may also be defined for variety recommendation: each subregion then coincides with a recommendation domain, grouping those sites with the same best-performing genotype(s) (Gauch and Zobel, 1997). The definition of subregions is not just geographical, but may also encompass farming practices (e.g. irrigated or rainfed cropping).
Subregions have sometimes been defined on the basis of site similarity for environmental factors that are supposed to be important but are, in fact, chosen without a definite assessment of their impact on GL interaction (e.g. Pollak and Corbett, 1993). The arbitrariness of this procedure makes its results less reliable than those for site similarity for GL effects (Gauch, 1992[3]). However, additional information on the climatic, soil, biotic and crop management variables closely related to the occurrence of GL interaction may help locate geographic boundaries for subregions (see Section 5.8), besides contributing to the understanding of causal factors for the interaction (Eisemann et al., 1990; Bidinger et al., 1996; van Eeuwijk et al., 1996). The zoning process should produce subregions that can be defined on a geographical basis or by other means, such as climatic factors or management practices, in order to be useful for breeding or cultivar recommendation. In reality, this is not always the case: for example, subregion definition for maize variety recommendation in Italy revealed a leopard skin pattern as a result of the inconsistency between geographic proximity and similarity for GL interaction effects of the sites, which could not be accounted for by any of the available environmental variables (Annicchiarico et al., 1995). In this case, recommendations concerned the whole region. In other instances, individual locations or small groups of sites (apparently distinct, in terms of GL effects, from a larger group of geographically close locations) may be added to the larger group to form a unique subregion for breeding or recommendation (e.g. Annicchiarico and Perenzin, 1994).
Setting adaptation strategies for breeding programmes and defining recommendation domains for cultivars are distinct objectives. As such, they may require partly different analytical approaches and provide different results with regard to the definition of subregions. The same data set may be analysed with both objectives in mind. However, the adaptation strategy objective focuses on the responses of a set of genotypes to obtain indications and generate predictions relative to future breeding material that may be produced from the genetic base of which the tested genotypes are assumed to be a representative sample.
Assessing the value of a specific adaptation strategy, implying a distinct selection programme for each subregion rather than a unique selection programme for the whole target region, is of obvious interest to globally-oriented breeding programmes of large seed companies or international research centres, where the target region may include more than one country and very different environments. In this case, each subregion may include several countries. Specific adaptation, however, may also prove a valuable target for national breeding programmes, for which the yield gain derived from exploitation of GL interaction effects within the country can also help face the increasing competition exerted on local seed markets by international seed companies. For public institutions, the breeding of diversified, specifically adapted germplasm can be a major element of a research policy enforcing sustainable agriculture (Bramel-Cox et al., 1991; Ceccarelli, 1996) by:
maximizing the potential of different areas by fitting cultivars to an environment instead of altering the environment (possibly with costly or environment-unfriendly inputs, such as pesticides, fertilizers and irrigation) to fit widely adapted cultivars; and
safeguarding crop biodiversity by increasing the number of varieties under cultivation, with positive implications for the stability of production at national level.
Furthermore, specific breeding may facilitate the technological adaptation of varieties by fixing characteristics of specific interest to subregions (for small-grain cereals, short straw for intensive cereal farming and long straw for extensive cereal-livestock systems; for cereal or food legume crops, different grain quality characteristics etc.).
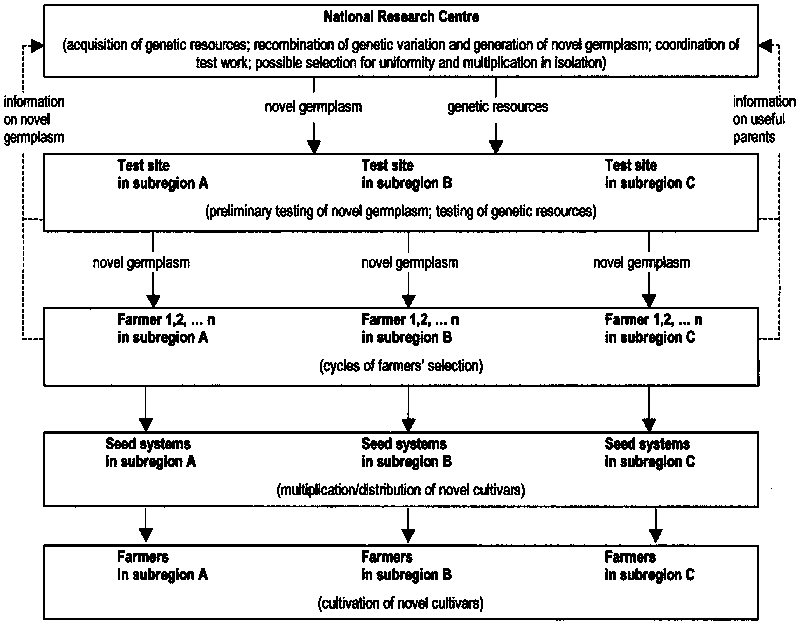
In general, breeding for specific adaptation tends to imply greater genetic gains associated with increased costs relative to those for a wide adaptation strategy. The genetic gains are derived from exploitation of GL interaction effects via useful adaptive traits (Bidinger et al., 1996), as well as increased heritability of yield as a consequence of decreased GL interaction (Kang, 1998). The relatively high costs may be due to increased field testing rather than to duplication of breeding stations, because crossing and hybridization operations can be centralized in a single national station providing each subregion with novel germplasm for local selection (and, possibly, genetic resources for local testing to identify parent material of specific interest). Figure 2.2 presents a hypothetical situation including a phase of farmers’ selection in a participatory plant breeding scheme (which may or may not be present).
A comparison of wide vs. specific adaptation strategies may be based on:
yield gains predicted for the different options according to selection theory, using the same yield data already used in the preliminary definition of subregions; or
actual yield gains provided by different strategies, following further research work (e.g. Singh et al., 1992; Ceccarelli et al., 1998).
In fact, more complex scenarios (including indirect selection gains in one subregion, derived from direct selection in another subregion) can also be envisaged (see Section 6.1). The comparison between adaptation strategies based on predicted yield gains may underestimate the potential advantage of a specific strategy, when material with markedly specific adaptation is under-represented among the tested varieties or breeding lines for various reasons (e.g. previous selection for wide adaptation by local breeding or wide representation in a sample of foreign, widely adapted material). Likewise, the potential gain of specific breeding for unfavourable areas may be underestimated when most tested genotypes have been selected in favourable environments. Specific adaptation could be more advantageous if it also implied the use of a distinct genetic base for each subregion. Although not negligible, the positive effects of specific breeding on the biodiversity of cultivated material are difficult to quantify.
For the definition of adaptation strategies, the comparison of yield responses across environments managed for contrasting levels of a given stress (e.g. drought or nutrient deficiency, dependent on the level of irrigation or fertilization, respectively) is a valid alternative to the current approach (based on the analysis of regional trials managed according to ordinary practices in each area), provided that the managed environments can represent actual geographic areas or farming systems within the target region. The use of managed environments is currently considered mainly for genotype selection within a given adaptation strategy (see Section 6.2).
Even if a wide adaptation strategy is preferred, the provisional identification of subregions can help locate crucial test sites for germplasm selection (Abou-El-Fittouh et al., 1969; Lin and Butler, 1988; Annicchiarico, 1992). In this case, a few sites representative of different subregions and capable of reproducing the mean responses of genotypes across the region can be used for selection of material either contemporarily (Brennan et al., 1981) or in turn (Calhoun et al., 1994). Identifying crucial test sites can also be a valuable objective for the routine evaluation of genotypes carried out by public institutions, such as those committed to the definition of lists of recommended varieties, or those responsible for the assessment of the Value for Cultivation and Use of newly released germplasm.
Decisions on the adaptation strategy, which can have a considerable and lasting effect on the organization of a plant breeding programme, should be based on the analysis of more data sets if available, and verified after a reasonable period of time on the basis of new data (DeLacy et al., 1994).
Already in the 1920s, studies of plant material (Turesson, 1922) leading to the introduction of the term “ecotype” highlighted the occurrence of specific adaptation to certain areas and environmental conditions. During the same period, pioneer breeders in Great Britain (Engledow, 1925) and Strampelli, Italy (Lorenzetti, 2000) advocated the importance of understanding and exploiting specific adaptation effects in order to raise crop yields in their respective countries. However, breeding programmes in the second half of the century mostly concentrated on the improvement of yield potential (Bramel-Cox et al., 1991; Evans, 1993) using - also in less developed countries - favourable environments for selection (Simmonds, 1991). Also the concept of plant ideotype to select for has been developed for different species to increase yields under favourable cropping conditions (Donald, 1968; Mock and Pearce, 1975). This trend has been favoured by numerous factors:
the perspective of rapid yield gain offered by high input levels;
the greater profitability of targeting seed markets in more productive areas; and
the common belief that selection in favourable areas produces a substantial yield increase also in less favourable areas.
This belief has been challenged by theoretical work (Jinks and Connolly, 1973; Falconer, 1990; Simmonds, 1991) and by mounting experimental evidence concerning improvement of crops for environments suffering from drought stress (Nageswara Rao et al., 1989; Ceccarelli and Grando, 1991; Ud-Din et al., 1992; Byrne et al., 1995; Muñoz et al., 1998; Ceccarelli et al., 1998), nutrient deficiency (Atlin and Frey, 1989; Bänzinger et al., 1997) or both (Cooper et al., 1997). These works have highlighted that selecting under low stress conditions implies a specific adaptation target towards favourable areas.
In reality, favourable, high-yielding environments for selection are not necessarily so because of natural climatic or soil factors. Optimal management at research stations, including abundant fertilization, accurate soil preparation, timely sowing, irrigation and chemical control of weeds, pests and diseases, can often result in higher mean yields than in neighbouring farmers’ fields. Such differences in yield may also imply remarkable GE interaction between these environments (Pederson and Rathjen, 1981; Ceccarelli, 1994). Spaced planting or low density procedures adopted for selection may also result in improved cropping environments, because they increase the availability of resources, such as light, water and nutrients (Rotili and Zannone, 1975).
In recent decades, various programmes run at international research centres have modified their breeding strategy to produce germplasm suitable for cropping in less favourable areas. A specific adaptation strategy has been pursued for barley improvement at the International Centre for Agricultural Research in Dry Areas (ICARDA), with selection under conditions similar to those in the target environment (favourable or drought-stressed), and with implications extending also to elements of the breeding strategy, such as the choice of genetic resources and variety type (Ceccarelli, 1994). A “shuttle breeding” procedure - alternate selection in drought-stressed (unfavourable) and irrigated (favourable) environments - was established for the selection of widely adapted germplasm at CIMMYT (Edmeades et al., 1989; Calhoun et al., 1994). However, the possibility of GE interaction between environments of similar ecological potential (based on crop mean yield) may lead to the definition of different subregions also within unfavourable and moderately favourable areas. For this reason, barley breeding at ICARDA began producing specific material for different sets of drought-prone countries (Ceccarelli, 1996); rice breeding at IRRI defined specific plant ideotypes for several different ecosystems (Fischer, 1996); and wheat breeding at CIMMYT is attempting a compromise between a wide adaptation prospect and the opportunity to breed specifically for 12 different mega-environments (Braun et al., 1996).
Different subregions may be identified not only within large or transnational regions (e.g. Crossa et al., 1991; DeLacy et al., 1994) but also within relatively small regions, as suggested by results for:
northern Syria (Ceccarelli, 1996);
Italy (Annicchiarico, 1997a) and northern Italy (Annicchiarico, 1992, 2002);
New South Wales (Seif et al., 1979; Basford and Cooper, 1998);
Queensland (DeLacy et al., 1996c);
southwest Canada (Saindon and Schaalje, 1993); and
Ontario (Yan et al., 2000).
Therefore, the choice between a wide and a specific adaptation strategy may be a key question for national breeding programmes.
The specific adaptation strategy is receiving increasing attention in developing countries, sometimes in combination with participatory plant breeding schemes. The participation of farmers can: i) support the multilocational selection work; ii) allow for exploiting possible specific adaptation effects even within subregions; iii) contribute to enhance the biodiversity of material under cultivation, thereby improving production stability; and iv) facilitate the seed supply to farmers via local seed systems (McGuire et al., 1999; Weltzien et al., 1999; Ceccarelli et al., 2000). For small farmers in relatively poor countries, such systems may be far more important than formal seed systems (Almekinders et al., 1994).
Agricultural techniques, following an assessment of their economic performance in regional yield trials, can be recommended: either widely over the target region or specifically for one subregion (Perrin et al., 1976; Shaner et al., 1982). For specific recommendation, the recommendation domain of a given technique may be defined on the basis of geography alone, or also on the basis of farming practices (e.g. irrigated or rainfed cropping) or socio-economic constraints. In all cases, the information obtained from previous testing is exploited for predicting yield responses in coming years and, most frequently, in new locations.
The definition of recommendations for varieties is generally simpler than for other techniques. Usually there is little difference in seed price among cultivars. In this case, variable costs do not need to be taken into account in the assessment. Statistical analysis concerns yield response and, where the genotypes differ for crop value, gross benefit. In either case, different domains (i.e. subregions which are the object of distinct recommendation) may be identified on the basis of repeatable GL interaction effects when varieties have different adaptation patterns. The domains are characterized by different top-yielding, “winning” genotypes (Gauch and Zobel, 1997; Ebdon and Gauch, 2002). For each location, the advantage of specific vs. wide recommendation can be assessed in terms of the difference in predicted yield between: i) the top-ranking genotype(s) in the relevant subregion; and ii) the top-ranking genotype(s) over the region (Gauch and Zobel, 1997).
More than one recommended genotype can be a sensible choice for either scenario, particularly for wide recommendation, as it may limit the risk of disasters arising due to the unforeseen susceptibility of the only cultivar recommended in a vast area to a biotic or abiotic stress. This is particularly the case for the cultivation of genetically homogeneous variety types (pure lines, clones) (Simmonds, 1979[4]). Possible differences in yield stability between cultivars can also be taken into account making recommendations (see sections below and Chapter 7).
Favouring the cultivation of specifically adapted germplasm is generally convenient for maximizing regional yields and increasing the biodiversity of cultivated material. However, subregions with very limited extension or negligible advantage of specifically adapted cultivars may be merged with larger, relatively similar subregions when the additional costs of multiplying and marketing specifically adapted germplasm are likely to outweigh the expected benefits (Gauch, 1992[5]).
Targeting of genotypes is also a concern of public and private seed companies who wish to verify the area of adaptation and the agronomic value of novel germplasm. This information, necessary for planning proper marketing and advisory schemes, is especially warranted when breeding has not contemplated a definite adaptation target. In this case, just one top-yielding genotype, across the region or in a distinct subregion, is usually identified for promotion to commercial variety status. Techniques envisaged herein for defining variety recommendations can also be useful for this purpose.
Yield stability
High yield stability usually refers to a genotype’s ability to perform consistently, whether at high or low yield levels, across a wide range of environments. As discussed more thoroughly in Section 7.1, most stability measures relate to either of two contrasting concepts of stability: “static” (Type 1) and “dynamic” (Type 2) (Becker and Léon, 1988; Lin et al., 1986). Static stability is analogous to the biological concept of homeostasis: a stable genotype tends to maintain a constant yield across environments. The term “environmental sensitivity” has also been used in this respect, where greater sensitivity corresponds to lower stability (Falconer, 1990; Dyke et al., 1995). Dynamic stability implies for a stable genotype a yield response in each environment that is always parallel to the mean response of the tested genotypes, i.e. zero GE interaction. The measure of dynamic stability depends on the specific set of tested genotypes, unlike the measure of static stability (Lin et al., 1986). Lin and Binns (1991) defined a Type 4 concept of stability that is strictly related to the static concept. Type 4 stability relates to consistency of yield exclusively in time, i.e. across years (or crop cycles) within locations, whereas Type 1 stability relates to consistency both in time and in space, i.e. across environments belonging to the same or different sites.
As with GL effects, the GE effects contributing to yield stability can be either:
exploited, by breeding and growing genotypes that are stable according to the static concept (i.e. with a better response in unfavourable environments or years); or
minimized, by using material that is stable according to the dynamic concept.
Static stability may be more useful than dynamic in a wide range of situations, especially in developing countries (Simmonds, 1991).
From a farmer’s point of view, location is a constant - not variable - factor, and yield consistency over time is the only relevant component of a genotype’s yield stability. It was therefore proposed (Barah et al., 1981; Lin and Binns, 1988) to evaluate yield stability with regard to GY interaction effects within locations. In reality, yield consistency in space also deserves consideration in the presence of sizeable GL interaction, since a selected or recommended genotype should be stable-yielding both across years and across locations in its area of adaptation or recommendation (Piepho, 1998). This is particularly so when there is a prospect of wide adaptation or recommendation, because in the context of a specific adaptation or recommendation the GL effects are minimized by the division of the target region into subregions. In a wide prospect, the assessment of yield stability in relation to genotype responses to environments provides a simple means for considering all possibly relevant GE interaction effects. In a specific prospect, assessment based on GY interaction effects within locations can be recommended, especially for genotype recommendation (see Section 7.2). Whatever the adaptation strategy, breeding for high yield stability can be considered a useful target when the relevant GE interaction variation is wide.
Yield reliability
High yield stability may be associated with low mean yield (or low stability with high mean yield), which complicates genotype selection or recommendation. As an extreme example of high stability associated with low yield, consider a hypothetic genotype that yields just above zero in all environments (greatest stability according to its static concept), or that is consistently the least-yielding (greatest stability according to the dynamic concept). Obviously, a less stable, higher-yielding genotype would be preferable. The practical interest of combining high levels of mean yield and yield stability has led to the development of the yield reliability concept (Eskridge, 1990; Kang and Pham, 1991; Evans, 1993). A reliable genotype is characterized by consistently high yield across environments. The use of a yield reliability index facilitates genotype selection or recommendation, as the mean yield and the yield stability traits are combined into a unique measure of genotype merit. Considering yield stability in conjunction with mean yield may also provide a more sensitive comparison of genotypes than when using only mean yield, owing to the reduction in Type 2 error rates (Kang, 1993). The stability measure contributing to the index of yield reliability may relate to the static or the dynamic concept of stability (see Section 7.2).
Genetic improvement
Despite its potential interest, increased yield stability has tended to be a minor objective in breeding programmes worldwide (Romagosa and Fox, 1993). A number of studies reviewed by Becker and Léon (1988) and Brancourt-Hulmel et al. (1997) confirmed the early indication by Allard and Bradshaw (1964) that variety types where the genetic structure implies high levels of heterozigosity and/or heterogeneity are less sensitive to environmental variation and are, therefore, more stable- yielding. Unfortunately, such types may sometimes offer fewer opportunities for maximizing the yield potential.
Within a given variety type, breeding successfully for this trait relies on the adoption of a heritable or repeatable stability measure as a selection criterion. There is extensive evidence (see Section 7.1) that heritability or repeatability values tend to:
vary from moderate to low, depending on the situation;
be higher when the assessment is done over two or more years or crop cycles; and
be somewhat higher for measures that relate to the static (Type 1 or Type 4) concept of stability.
Given the high sampling error, the assessment of yield stability requires numerous test environments (at least eight) to guarantee reliability (Kang, 1998; Piepho, 1998). Therefore, direct selection for yield stability may be limited by high costs and can be recommended, even when it has high priority, only for elite material in the final testing stages. The choice of parental germplasm with recognized yield stability and of, if possible, a convenient variety type, can play a major role in breeding for more stable crop yields. In addition, indirect selection for higher yield stability may be attempted by selecting for morphophysiological traits that have proved to be strictly associated with this character (see Section 6.3).
Adaptation strategy and yield stability targets
The main analytical steps involved in the definition of an adaptation strategy and yield stability targets on the basis of regional trial data are summarized in Figure 2.3 for the general case of experiments repeated also in time. There are six possible conclusions, implying a wide or specific adaptation strategy and, in both cases, the inclusion or exclusion of increased yield stability as a breeding target. Within the wide adaptation strategy, indications may or may not urge the choice of selection locations that contrast for GL interaction effects.
As a preliminary step, it is useful to estimate the variance components for: genotype; GE interaction across environments (location-year or location-crop cycle combinations); and the two determinants of GE interaction variance, i.e. the lack of genetic correlation among environments for genotype values, and the heterogeneity of genotypic variance among environments (see Section 4.3). Adaptive responses and yield stability of genotypes as affected by heterogeneity of genotypic variance have no practical importance for breeding, since they relate to GL and other GE effects originating from a scale effect of the environment and implying no change in relative response of genotypes. The larger variance of this determinant relative to the lack of genetic correlation variance definitely suggests its reduction through a suitable data transformation (as discussed, in particular, with respect to analysis of adaptation - see Section 5.6). In any case, if the lack of genetic correlation variance component is low (e.g. below 25-30%) compared to the genotypic variance component, it reveals the limited extent of GE interaction effects relevant to breeding, and supports (without further analyses) the selection for wide adaptation with no regard for yield stability (Fig. 2.3). To simplify the analysis (see Section 5.6), this preliminary step may be replaced by the estimation of the genotypic and GE interaction components of variance only, together with the verification of the relationship between location mean yield and within-location phenotypic standard deviation of genotype yields. If no correlation is found and the GE interaction variance component is larger than or about as large as the genotypic variance component (e.g. > 80-100%), the next analytical step can be considered.
Note: Environment as location-year or location-crop cycle combination (see Section 2.6 for comment of steps).
The next step involves the estimation of different genotypic and genotype-environmental variance components through an appropriate model of combined ANOVA (see Section 4.1). An analysis of adaptation may already be justified if the GL interaction variance component is significant and only moderately large (e.g. > 30-35% relative to the genotypic variance) (Fig. 2.3), also because the variance of some of its components as defined in the analysis of adaptation (e.g. the heterogeneity of genotype regressions component) may prove larger (see case study in Chapter 8). Following an appropriate modelling of the GL effects or the site classification based on similarity for these effects (see Chapter 5), subregions may be provisionally identified lending themselves to a practical definition based on geography, environmental factors or farming practices. Those that cannot be characterized as distinct from each other may be merged at this stage. Likewise, subregions which are too small to be of practical interest may be merged with larger ones.
Wide and specific adaptation strategies can be compared in terms of yield gains predicted from original yield data of the same data set (see Section 6.1). In the case of several candidate subregions, specific adaptation may contemplate other possibilities besides targeting each subregion (e.g. merging of some subregions, neglecting subregions of minor importance). In most cases, specific breeding would not contemplate more than two or three subregions. A final decision on the adaptation strategy may be made at this stage or, especially when there are no clear-cut indications, further research may be necessary to compare the best options on the basis of actual yield gains.
A wide adaptation strategy may arise from two events:
low GL interaction variance (paths 1 and 2 in Fig. 2.3); or
high GL interaction variance with no clear advantage of specific breeding (paths 3 and 4 in Fig. 2.3).
When the latter event occurs, the final selection stages may be devised across contrasting locations capable of reproducing GL interaction effects and, thereby, the genotype mean responses across the target region. Analysis of adaptation helps in the identification of these locations, as well as of tests of special interest for early stages of selection in the context of the adopted adaptation strategy (see Section 6.2). The selection for wide or specific adaptation may also be attempted through artificial or managed selection environments capable of reproducing the genotype responses across the target region (see Section 6.2).
Decisions regarding yield stability depend essentially on the size of other GE interaction variance components, which may only be estimated if the trials are repeated also in time. In particular, the GY and GLY interaction components, or the GY interaction within locations component (as estimated from the adopted model of combined ANOVA - see Section 4.1), are relevant to decisions in the context of a wide adaptation strategy (options 1 vs. 2, or 3 vs. 4, in Fig. 2.3). Breeding for yield stability is justified when the overall variance accounted for by the relevant GE effects is large relative to the genotypic variance component (e.g. > 150 or 200%). In a specific adaptation prospect (options 5 vs. 6), decisions may differ for individual subregions, suggesting yield stability as a valuable target in some cases only. They may be based on the size of the estimated GE interaction variance component relative to that of the genotypic variance in the relevant subregion, following separate ANOVAs limited to test environments of each subregion (this simple procedure includes the GL interaction variance in the assessment, but its size within the subregion is limited due to the zoning process). Selection for yield stability (based on an index of yield reliability) requires a higher number of selection environments (Fig. 2.3) and the definition of the stability concept to breed for (see Chapter 7).
Cultivar recommendation
For definition of an adaptation strategy and yield stability targets, the contribution of any of the tested genotypes (considered as a sample of the relevant genetic base) is equally important. Emphasis is therefore placed on the estimation of genotypic and genotype-environmental components of variance, and location similarity is assessed on the basis of adaptation patterns for all genotypes. Conversely, the most important information for variety recommendation concerns the response of, and comparison between, high-yielding genotypes. The main analytical steps of interest in this context are summarized in Figure 2.4. There are four possible conclusions, implying a general recommendation for the target region or a specific recommendation for distinct subregions and, in both cases, the inclusion or exclusion of yield stability in the assessment of genotype merit.
Specific recommendation for different subregions can conveniently be envisaged when, in the presence of significant GL interaction in a combined ANOVA, the GL effects modelled through an analysis of adaptation imply the rank inversion of top-yielding genotypes between locations. The subregions are characterized by different best-performing genotypes (see Chapter 5).
For trials repeated also in time, the definition of subregions and recommendation domains based on mean yield (general recommendation) or adaptation patterns (specific recommendation) of the best-yielding genotypes may change if yield stability is also taken into account by means of a yield reliability index. The presence of highly significant GE interaction for effects other than GL effects supports the interest of the yield stability characteristic. However, yield stability needs to be taken into account only if an entry from the best-performing set (on the basis of mean yield, following an appropriate comparison of means) shows significantly lower yield stability than other genotypes (Fig. 2.4). It is usually preferable to estimate yield stability and reliability values with reference to all GE interaction effects (GL included) for general recommendation, and average GY interaction within locations for specific recommendation (in which GL effects are already accounted for by the modelled adaptation patterns). For specific recommendation, varieties and possible subregions (i.e. recommendation domains) can be identified by modelling the yield reliability (rather than the mean response) of genotypes across locations (exploiting also the information generated by analysis of adaptation: see Section 7.2). Subregions that are barely distinguishable on a geographical basis or according to other characteristics may be merged and their information combined, thus defining a unique recommendation domain. Likewise, small subregions may be merged with larger ones. When appropriate, gross benefit (rather than yield) may be analysed following the steps reported in Figure 2.4. In this case, adaptation, stability and reliability concepts all relate to this economic variable.
Note: Environment as location-year or location-crop cycle combination (see Section 2.6 for comment of steps).
Information on environmental variables (including farming practices) closely related to the occurrence of GL interaction can facilitate the characterization of subregions for breeding or genotype recommendation, as well as the assignment of new locations (see Section 5.8). This spatial scaling-up of results may be combined with a temporal scaling-up, by re-assigning test locations to subregions depending on long-term values of climatic or biotic variables (thereby compensating for the effect of unusual years on the location response for GL interaction effects).
|
[3] Ibid., p.
220 [4] Ibid., p. 267 and p.361 [5] Ibid., p. 220 |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}