![]()
![]()
![]()
The concentration of dissolved gases in water is of prime importance in considering the quality of water along with the other physico-chemical characteristics.
Ambient concentration of dissolved oxygen, carbon dioxide, nitrogen and ammonia are of greater importance, eventhough depending on the premises hydrogen sulfide, sulfur dioxide, chlorine etc, can also be of significance in aquaculture.
In view of the importance of gas content (in solution) of a mass of water we shall recall the gas laws, which you must be familiar with: Henry's law, which states that “the concentration of saturated solution of a gas is proportional to the pressure at which the gas is supplied” and Dalton's law of partial pressure, which states that, “the pressure exercised by a gas is proportional to the component of its concentration in the mixture, and the total pressure of the gas is equal to sum of its components”.
Thus the amount of consituent gases in atmosphere that will dissolve in water depends on the proportion of each gas in the air and the solubility of each constituent gas. A given volume of atmospheric air contains 78.084% nitrogen, 20.946% oxygen, and 0.934% argon. The remaining 0.036% contains carbon dioxide, noble gases and trace gases such as helium, krypton, neon and xenon plus water vapour. The partial pressure in the liquid phase that would be in equilibrium with measured concentration is called the dissolved gas pressure (TGP) (EIFAC, 1986) under natural conditions any of the three states can occur in water:
| TGP | = | BP (water is supersaturated - BP Barometer pressure in mm) |
| TGP | = | BP (In equilibrium) |
| TGP | = | BP (water is undersaturated) |
Percentage saturation can be indicated in terms of the constituent gases, or in terms of the total dissolved gases:

At an atmospheric pressure of 760 mm Hg (NTP) the partial pressure of oxygen (pO2) will be 159 mm Hg i.e.

At 100% air saturation solubility of oxygen at 30°C is 7.54 mg/l (see Table 1). If one records 6 mg/l at the same temperature then the corresponding pO2 value will be:

or 126.5/159 (6.0/7.54) = 79.6% saturation.
For presentation of values in different units of expression please see Fig. 2. To give an example, for conversion mg/l of oxygen to ml/1, multiply by 0.7 (i.e. 22.4/32 or mole volume/mole weight).
The sources of dissolved oxygen (D.O.) in natural waters is from atmosphere and photosynthesis of plants.
Oxygen is absorbed in water by direct diffusion and by surface-water agitation. Solubility of oxygen in water is so small and by diffusion process alone in still water, it was culculated that it would take 6 years for oxygen to diffuse from surface to a depth of 6 meters in quiet water. Absorption of water is very minor, that almost all the oxygen enrichment of natural waters takes place by agitation of water.
Photosynthesis of plants supply oxygen considerably - but this limited to the zones to which light penetration takes place. Photoperiod (day light duration), diurnal and seasonal, would decide the amount of oxygen produced by photosynthesis. Daily pulses of oxygen could be seen clearly in waters with high concentration of plankton - with an afternoon maximum and early morning drop, in some cases taking D.O. values close to zero. This can be seen in our tropical waters, where phytoplankton blooms occur. This will be referred to again in our discussion.
In this group the following can be listed:
Respiration of fauna and flora; decomposition of organic matter, reduction due to other gases - gases such as methane, CO2 and others which accumulate in the bottom bubble up and wash out the oxygen dissolved in water (cf. excess CO2 released through water in the volcanic lake in Cameroun in 1986, release of oxygen from super-saturated surface waters - when upper waters full with O2 warm up the upper waters become supersaturated and release O2 to the air, inflow of subterranean waters of low O2 content (this is negligible under fish pond conditions), presence of iron - oxidation of iron to form soluble ferric hydrate consumes oxygen in iron-rich waters.
All these may combine together and act at times to reduce oxygen completely, especially in shallow plankton-rich waters in high ambient temperatures such as in the fish pond.
As indicated earlier, the atmospheric air has 20.9% (209 ml/1) of oxygen. Relatively the amount (volume) of DO in water is much less, 5.3 ml in a litre of air-saturated water at 30°C i.e. about 0.5% - by weight the proportion is still less - 7.6 mg in a litre or kg of water (see Fig. 2).
Since finfish and shellfish depend on primarily oxygen dissolved in water for sustenance (the air-breathing fishes are exceptions, but their capacity for tolerance of hypoxic conditions can be made use of in aquaculture e.g. fish culture in sewage oxidation ponds) the relative handicaps in having water as respiratory medium as compared with air can be pointed out.
Since the content of oxygen in water is very much less in water than in air, the fish has to pump through its gills relatively a larger amount of the medium, which is much heavier too, to extract sufficient oxygen to sustain itself. An air-breather in this connection has relatively a greater saving of energy and it is yet to be established if the air-breathing fish fares better in energy conversion and growth because of this advantage but the air-breathing fish spends considerable energy in surfacing frequently especially if it is in deep water. It is interesting to explore this aspect further. Water appears to be a risky medium when compared with air and the variety of adaptations of aquatic life to ambient oxygen bear testimony to this.
The low diffusion of oxygen in water referred to already and the stratification of gases even in small bodies of water, unlike atmospheric air which is uniform, make each water body distinct in its nature. This has significance in that each water bodythas to be therefore carefully studied for choosing it for aquaculture.
The solubility of oxygen in water values with temperature, the DO content reducing sharply with increase in temperature. The solubility value for oxygen at various temperature in vogue for long time were those given by Treusdale et al (1955); these were re-estimated using improved Henry coefficients (Benson & Krauz, 1980), discussed in Cole (1983). The values of solubility of oxygen in water at various temperatures, from moist air at 760 mm Hg at 0.0 ppt salinity, taken from EIFAC (1986) is given in Table II. The values have been compiled with specific reference to the need of the aquaculturist and hence presented here in detail. The EIFAC values are slightly different from those given in APHA (1985) or Cole (1983), but for most practical purposes in the field (unless working with very sensitive high intensity systems) these latter are also acceptable.
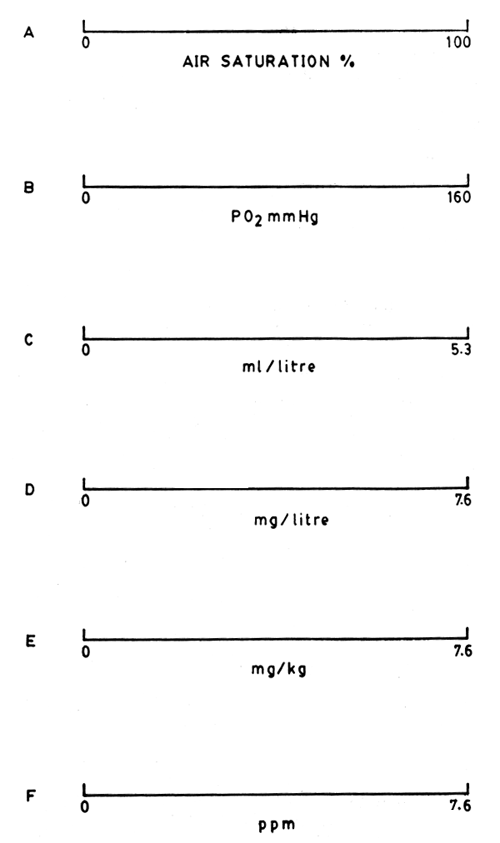
Fig. 2. Units of expression of dissolved oxygen concentration in water. The values shown are for air-saturated freshwater at 30°C at BP 160mm Hg. A. Percent air saturation; B. Partial pressure of O2, pO2 would change slightly with temperature; C. ml O2/litre i.e. volume of oxygen per litre of water; D. mg O2/litre i.e. weight of oxygen per litre of water; E. mg O2/kg i.e. weight of oxygen per kg of water; F. ppm - parts per million. Note E & F are same, but D could be different from E & F slightly with temperature. See also figure, density/temperature changes with water (Chap. 8, this manual).
A simple method of estimating saturation values for specific temperature is to aerate, using a diffuser, a litre of water you are studying in a flask over night, maintaining it at the temperature required, and then taking a sample and estimating DO in it by Winkler titration (see APHA, 1985; also our handout on “Water Analysis” given separately). This air saturated water can also be used for standardising the DO electrode (eg. YSI DO Analyser Model 51B - this analyser has in addition a simpler method of calibration exposing the electrode to atmospheric air). It must be pointed out here that several field kits available commercially give only gross values of DO. These kits use titrants, a drop of which equals 1 mg/L of DO, under the conditions of test. Increasing the size of sample can improve this accuracy, but it must be borne in mind that the accuracies we are concerned with in sensitive culture systems (in view metabolic requirements) are fractions of a mg (see Fig. 3.). Thus the kits are permissible only for quick and rough field uses.

Fig. 3. Distribution of oxygen concentrations (mgO2.L-1) in a circular fish tank: stocking density 62.3 kg.m-3, water flow rate 6.3 L.kg:h-1; oxygen supply through attached aeration pockets. Shaded area = approximate distribution of fish during the observation period, n = number of observations at each point of the vertical profile. (Source, EIFAC, 1986).
TABLE I
Solubility of oxygen in mg/L as a function of temperature (mount air barometric pressure = 760 mm Hg, salinity = 0.0 ppt) (Source: EIFAC, 1986)
| Temp (°C) | 0.0 | 0.1 | 0.2 | 0.3 | 0.4 | 0.5 | 0.6 | 0.7 | 0.8 | 0.9 |
| 0 | 14.602 | 14.561 | 14.520 | 14.479 | 14.438 | 14.398 | 14.358 | 14.318 | 14.278 | 14.238 |
| 1. | 14.198 | 14.159 | 14.120 | 14.081 | 14.042 | 14.004 | 13.969 | 13.927 | 13.889 | 13.851 |
| 2 | 13.813 | 13.776 | 13.738 | 13.701 | 13.664 | 13.627 | 13.591 | 13.554 | 13.518 | 13.482 |
| 3 | 13.445 | 13.410 | 13.374 | 13.338 | 13.303 | 13.268 | 13.233 | 13.198 | 13.163 | 13.128 |
| 4 | 13.094 | 13.060 | 13.025 | 12.991 | 12.957 | 12.924 | 12.890 | 12.857 | 12.823 | 12.790 |
| 5 | 12.757 | 12.725 | 12.692 | 12.659 | 12.627 | 12.595 | 12.563 | 12.531 | 12.499 | 12.467 |
| 6 | 12.436 | 12.404 | 12.373 | 12.342 | 12.311 | 12.280 | 12.249 | 12.218 | 12.188 | 12.158 |
| 7 | 12.127 | 12.097 | 12.067 | 12.037 | 12.008 | 11.978 | 11.949 | 11.919 | 11.980 | 11.861 |
| 8 | 11.832 | 11.803 | 11.774 | 11.746 | 11.717 | 11.689 | 11.661 | 11.632 | 11.604 | 11.577 |
| 9 | 11.549 | 11.521 | 11.493 | 11.466 | 11.439 | 11.412 | 11.384 | 11.357 | 11.331 | 11.304 |
| 10 | 11.277 | 11.251 | 11.224 | 11.198 | 11.172 | 11.145 | 11.119 | 11.093 | 11.068 | 11.042 |
| 11 | 11.016 | 10.991 | 10.995 | 10.940 | 10.915 | 10.890 | 10.865 | 10.864 | 10.815 | 10.791 |
| 12 | 10 766 | 10.741 | 10.717 | 10.693 | 10.669 | 10.645 | 10.620 | 10.597 | 10.573 | 10.549 |
| 13 | 10.525 | 10.502 | 10.478 | 10.455 | 10.432 | 10.409 | 10.386 | 10.363 | 10.340 | 10.317 |
| 14 | 10.294 | 10.271 | 10.249 | 10.226 | 10.204 | 10.182 | 10.160 | 10.137 | 10.115 | 10.094 |
| 15 | 10.072 | 10.050 | 10.028 | 10.007 | 9.985 | 9.964 | 9.942 | 9.921 | 9.900 | 9.879 |
| 16 | 9.858 | 9.837 | 9.816 | 9.795 | 9.774 | 9.753 | 9.733 | 9.712 | 9.692 | 9.672 |
| 17 | 9.651 | 9.631 | 9.611 | 9.591 | 9.571 | 9.551 | 9.531 | 9.512 | 9.492 | 9.472 |
| 18 | 9.453 | 9.433 | 9.414 | 9.395 | 9.375 | 9.356 | 9.337 | 9.318 | 9.299 | 9.280 |
| 19 | 9.261 | 9.242 | 9.224 | 9.205 | 9.187 | 9.168 | 9.150 | 9.131 | 9.113 | 9.095 |
| 20 | 9.077 | 9.058 | 9.040 | 9.022 | 9.004 | 8.987 | 8.969 | 8.951 | 8.933 | 8.916 |
| 21 | 8.989 | 8.881 | 8.863 | 8.846 | 8.829 | 8.812 | 8.794 | 8.777 | 8.760 | 8.743 |
| 22 | 8.726 | 8.709 | 8.693 | 8.676 | 8.659 | 8.642 | 8.626 | 8.609 | 8.583 | 8.576 |
| 23 | 8.560 | 8.544 | 8.528 | 8.511 | 8.495 | 8.479 | 8.463 | 8.447 | 8.431 | 8.415 |
| 24 | 8.400 | 8.384 | 8.368 | 8.352 | 8.337 | 8.321 | 8.306 | 8.290 | 8.275 | 8.260 |
| 25 | 8.244 | 8. 29 | 9.214 | 8.199 | 8.184 | 8.168 | 8.153 | 8.139 | 8.124 | 8.109 |
| 26 | 8.094 | 8.079 | 8.065 | 8.050 | 8.035 | 8.021 | 8.006 | 7.992 | 7.977 | 7.963 |
| 27 | 7.949 | 7.934 | 7.920 | 7.906 | 7.892 | 7.878 | 7.864 | 7.850 | 7.836 | 7.822 |
| 28 | 7.808 | 7.794 | 7.780 | 7.766 | 7.753 | 7.739 | 7.725 | 7.712 | 7.698 | 7.685 |
| 29 | 7.671 | 7.658 | 7.645 | 7.631 | 7.618 | 7.605 | 7.592 | 7.578 | 7.565 | 7.552 |
| 30 | 7.539 | 7.526 | 7.513 | 7.500 | 7.487 | 7.475 | 7.462 | 7.449 | 7.436 | 7.424 |
| 31 | 7.411 | 7.398 | 7.386 | 7.373 | 7.361 | 7.348 | 7.336 | 7.324 | 7.311 | 7.299 |
| 32 | 7.287 | 7.274 | 7.262 | 7.250 | 7.238 | 7.226 | 7.214 | 7.202 | 7.190 | 7.178 |
| 33 | 7.166 | 7.154 | 7.142 | 7.130 | 7.119 | 7.107 | 7.095 | 7.083 | 7.072 | 7.060 |
| 34 | 7.049 | 7.037 | 7.026 | 7.014 | 7.003 | 6.991 | 6.980 | 6.969 | 6.957 | 6.946 |
| 35 | 6.935 | 6.924 | 6.912 | 6.901 | 6.890 | 6.879 | 6.868 | 6.857 | 6.846 | 6.835 |
| 36 | 6.824 | 6.813 | 6.802 | 6.791 | 6.781 | 6.770 | 6.759 | 6.748 | 6.738 | 6.727 |
| 37 | 6.716 | 6.706 | 6.695 | 6.685 | 6.674 | 6.664 | 6.653 | 6.643 | 6.632 | 6.622 |
| 38 | 6.612 | 6.601 | 6.591 | 6.581 | 6.570 | 6.560 | 6.550 | 6.540 | 6.530 | 6.520 |
| 39 | 6.509 | 6.499 | 6.489 | 6.479 | 6.469 | 6.460 | 6.450 | 6.440 | 6.430 | 6.420 |
| 40 | 6.410 | 6.400 | 6.391 | 6.381 | 6.371 | 6.361 | 6.352 | 6.342 | 6.333 | 6.323 |
TABLE II.
Solubility of oxygen, nitrogen, and argon (mg/L) as a function of depth (moist air, T = 20°C, salinity = 0.0 ppt, p = 760 mm Hg)
(source: EIFAC, 1986)
| Depth m | N2+Ar | O2 | Total |
| 0.00 | 15.43 | 9.08 | 24.51 |
| 0.10 | 15.58 | 9.17 | 24.75 |
| 0.20 | 15.74 | 9.26 | 24.99 |
| 0.30 | 15.89 | 9.35 | 25.24 |
| 0.40 | 16.04 | 9.44 | 24.48 |
| 0.50 | 16.19 | 9.53 | 25.72 |
| 0.60 | 16.35 | 9.62 | 25.96 |
| 0.70 | 16.50 | 9.71 | 26.21 |
| 0.80 | 16.65 | 9.80 | 26.45 |
| 0.90 | 16.80 | 9.89 | 26.69 |
| 1.00 | 16.96 | 9.98 | 26.93 |
| 1.10 | 17.11 | 10.07 | 27.18 |
| 1.20 | 17.27 | 10.16 | 27.42 |
| 1.30 | 17.41 | 10.25 | 27.66 |
| 1.40 | 17.57 | 10.34 | 27.90 |
| 1.50 | 17.72 | 10.43 | 28.15 |
| 1.60 | 17.87 | 10.52 | 28.39 |
| 1.70 | 18.02 | 10.61 | 28.63 |
| 1.80 | 18.18 | 10.69 | 28.87 |
| 1.90 | 18.33 | 10.78 | 29.11 |
| 2.00 | 18.48 | 10.87 | 29.36 |
| 2.10 | 18.64 | 10.96 | 29.60 |
| 2.20 | 18.79 | 11.05 | 29.84 |
| 2.30 | 18.94 | 11.14 | 30.08 |
| 2.40 | 19.09 | 11.23 | 30.33 |
| 2.50 | 19.25 | 11.32 | 30.57 |
| 2.60 | 19.40 | 11.41 | 30.81 |
| 2.70 | 19.55 | 11.50 | 31.05 |
| 2.80 | 19.70 | 11.59 | 31.30 |
| 2.90 | 19.86 | 11.68 | 31.54 |
| 3.00 | 20.01 | 11.77 | 31.78 |
| 3.10 | 20.16 | 11.86 | 32.02 |
| 3.20 | 20.31 | 11.95 | 32.27 |
| 3.30 | 20.47 | 12.04 | 32.51 |
| 3.40 | 20.62 | 12.13 | 32.75 |
| 3.50 | 20.77 | 12.22 | 32.99 |
| 3.60 | 20.92 | 12.31 | 33.24 |
| 3.70 | 21.08 | 12.40 | 33.48 |
| 3.80 | 21.23 | 12.49 | 33.72 |
| 3.90 | 21.38 | 12.58 | 33.96 |
| 4.00 | 21.53 | 12.67 | 34.21 |
NOTE: Values of nitrogen and argon are important when supersaturation values are being considered.
It becomes obvious from the values of solubility of oxygen that the warm tropical and semitropical waters have ordinarily contain much less oxygen than these of the temperate, but fish production in the tropric does not appear to be limited by this relative lesser availability of oxygen as we have indicated elsewhere the rate functions which enhance growth are higher at the warmer tropical temperature (unless it is too high) and generally the tropical fishes have a better capacity to extract oxygen from oxygen deficient waters, by means of adaptations of haemoglobin. We shall be reverting to this when species relations are considered.
Solubility of oxygen in water changes markedly with depth of water body, increasing with the increasing pressure with depth (see discussion under pressure and depth, elsewhere, “Physical features of waters”). Table II taken from EIFAC (1986), again demonstrates this aspect very clearly. At 20°C oxygen solubility content increases from a value of 9.08 mg/L at the surface to 9.98 mg/L at m depth (Table II). Proportionate reductions in values could be expected for other temperatures as well.
Solubility of any gas decreases markedly with increase in dissolved solids. This is not obvious for DO in ordinary freshwater, but is marked when the salinity increases. The dissolved oxygen content (saturation values) for waters of different chlorinities are given in APHA (1985). The DO value changes from 7.6 in freshwater (0 ppt. S.) to 6.8 at 27 ppt.S. and 6.4 in sea water (35 ppt.S.). This reduction in oxygen availability should be considered, when aquaculture practices are planned in brackish water and sea water conditions.
In the above discussion we have referred to the oxygen content in air-saturated water only i.e. the solubility of oxygen in water. The oxygen content of natural waters can vary widely depending on the physical, chemical and biological processes in the water body as already indicated. Running waters generally have higher oxygen content than stagnant waters. As explained the major source of oxygen in natural waters is the atmospheric air. Oxygen enters water by the process of diffusion and higher the mixing of air and water due to surface-water agitation such as by wave action and turbulence in running water or due to artificial process of aeration by compressed air diffusers and use of agitators, in aquaria and ponds, the higher and oxygen saturation. Apart from diffusion from the air, water bodies gain oxygen through photosynthesis of chorophyll bearing aquatic organisms i.e. higher plants, phytoplankton and photosynthetic bacteria.
Oxygen is lost from the water body by the respiration of animals and plants and also by decomposition of organic matter. Excess oxygen will also be released from water to air by diffusion. Evolution of other gases (N2, H2S, CH4, CO2) from the bottom also can cause loss of oxygen by their stripping action. (The latter principle is used in stripping oxygen from water by bubbling nitrogen in the laboratory e.g. Gay-Lussac column).
In larger water bodies where thermal stratification takes place, there is often a stratification in oxygen also evident from the oxygen profile of the water. If the water body is not very productive (oligotrophic) then while the temperature falls markedly with depth the oxygen content does not change, showing ‘orthograde’ distribution of oxygen, but if the water body is productive or rich (eutropnic) then the oxygen concentration falls steeply with depth in the hypolimnion (heterograde distribution). This lack of oxygen is caused due to the lack of production of oxygen by the darker hypolimnetic zone and also due to the utilization of the available oxygen by decomposing plant and animal materials which rain from the upper rich zone. Oxygen level becomes often zero in the bottom waters. This change is of great significance in the large water bodies by its influence on the soil-water interface - a thin layer of bottom soil, referred to as oxidized microzone layer becoming reduced (see Section 15). The nature of the water body and its oxygen profile are thus very important in considering fish stocking and management.
The major types of oxygen profiles recognized in stratified natural water bodies in the temperate region are described well in limnology textbooks. The four commonly recognized profiles described by Hutchinson (1957) are indicated in Fig. 4. Stratification of lakes with reference to temperature and physico-chemical conditions, though mainly recognized in temperate conditions, are also seen in tropical water bodies in a modified form (see discussion under ‘temperature’); these are of special importance for aquaculturists working to stock open waters, which is also part of the present course. (“Stocking of Open Waters”.)

Fig. 4. Types of vertical-oxygen profiles in lakes typically of a stratified condition (generalised). (After Hutchinson, 1957)
In shallow waters there is likely to be little difference in the oxygen concentration at the surface and the bottom, but in deeper water bodies, as just referred to, this difference is likely to increase. In tropics where the water is already warm the oxygen content is less, any factor causing reduction of oxygen is important. A peculiar feature is the diel rhythm in oxygen in rich water bodies such as fertilized ponds. In the tropical fish pond supplied with high nutrients (an extreme case is the pond supplied with oxidized sewage) in the mid-afternoon the oxygen concentration would go high, often much higher than the saturation level owing to the high oxygen production by the phytoplankton and subsequently in the night towards morning, there will be a sharp deadline in oxygen level often reaching near complete depletion. In ponds supplied with organic wastes and small ponds with algal blooms in the tropics, with temperature around 30°C or above, the daily maximum in oxygen can be 2 – 3 times the saturation level. The subsequent depletion of oxygen in the absence of light in the night by the combined respiration of the large biomass (plants and animals) often takes the oxygen level in fish ponds to lethal levels of fishes. Those which live at the bottom (cf. prawns in brackish water ponds) more frequently are more susceptible even in shallow ponds, for the oxygen depletion would take place initially in the bottom. The time course of oxygen depletion can be predicted by making a plot of DO values obtained during the phase of DO during the late day/night against time of day and extrapolating a line, drawn through the DO value to zero DO on the time axis as explained in Boyd (1979) (Fig. 5).
The diel cycle of oxygen, was studied by Ali (1986) in ponds at ARAC, Aluu, Plots of Ali's data for Pond ‘9’ are shown in Fig. 6. As shown in the figure, the DO values hardly exceed saturation value in pond before fertilization, but the values are pronounced and reach high levels in the afternoon in ponds when phytoplankton blooms have developed consequent to fertilization (these will be discussed separately).
The observations of Ali with specific reference to local tropical conditions are similar to those of Boyd (1979). Ali's observations in a nursery pond at Aluu (Fig. 7), clearly shows how clear (sunny) weather and cloudy (overcast) weather affect the DO cycle. It is signigicant that the DO values which peak at the afternoons values close to zero towards dawn. The effect of consequent cloudy weather days will be to cause more serious low oxygen problems as indicated by Boyd (1979) (see also Fig. 5 indicating night time DO projection).

Fig. 5. Estimating DO decline during the night (After Boyd, 1979)
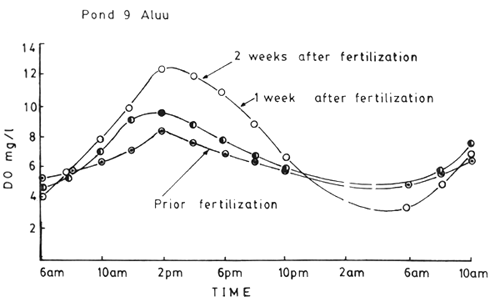
Fig. 6. Diel cycle of DO in pond 9 at Aluu before and after fertilization. (Max. temp at 2 p.m. was 33–35°C and Min at 6 a.m. was 28–29°C). (From Ali, 1986).
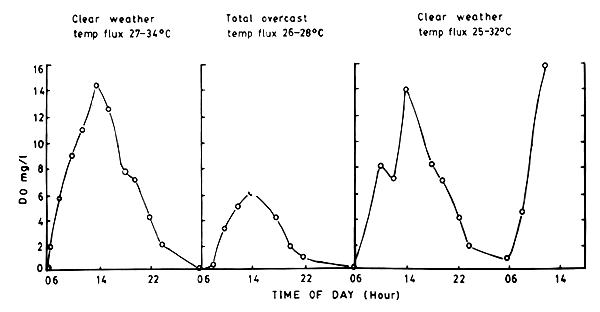
Fig. 7. Diel changes in DO in nursery pond ‘2’ at Aluu during clear weather and overcast (cloudy) days. (From Ali, 1986).
Depthwise oxygen profile at different times of day in a fish pond (pond 9) where phyto-plankton bloom was there was also show by Ali. His observations are shown in Fig. 8. It is interesting to see that at dawn while there was minimum oxygen in the pond (see also Ali's temperature date) the maximum DO was not at the surface but below (cf metalimnetic maximum for stratified lakes, Fig. 4), but as the day progresses, towards afternoon a clear supersaturation pattern develops in the pond, with a DO minimum of 6 mg/L (28°C) at the bottom and a DO maximum of 15 mg/L (34°C) at 14.00 hrs. What is the effect of fluctuating DO in fish growth? Tsadik (1984) studied the effect of fluctuating DO on Oreochromis niloticus and found that fluctuating DO reduces growth rate almost close to that caused by low DO itself (1 mg/L). Thus the DO changes will have great influence on the fish stocked in the pond. See also Wickins (1981)
Oxygen in depleted water can be increased by giving more chance for water and air to mix - fish pond owners in Asia often mix the surface water by beating with sticks and hands when they suspect oxygen depletion; mixing of water by electrical agitators and compressed air supply can also be effective. Water entering the pond can be made to cascade and splash down into the water for picking up more oxygen. Fertilizer application and blooms must be controlled so as to avoid the night time low of oxygen in ponds. Artificial lighting can cause increased photosynthesis and higher DO levels, but this is possible only in small often closed systems.
Increased water movement i.e. turbulence and mixing of water and any oxygen can be provided in the pond (especially shrimp ponds) by the following ways:
Changing of water - letting in new water as referred to may have to be done by pumping; if it takes hours for the tide to fill new warer the shrimp may die; using aeration equipment, such as air blower or compressor with air stones, electric paddle wheel; orienting the pond axis (during construction) with that of prevailing wind - wind may also cause erosion of the dyke - by wave action; providing greater sweep of wind across pond by construction of large ponds, decreasing depth of water to accentuate wind action; having low dykes which would not obstruct wind and avoid planting trees on dykes.
Relative merits of these provisions will have to be considered, as obvious, with the overall need of the pond under the prevailing condition of all factors.
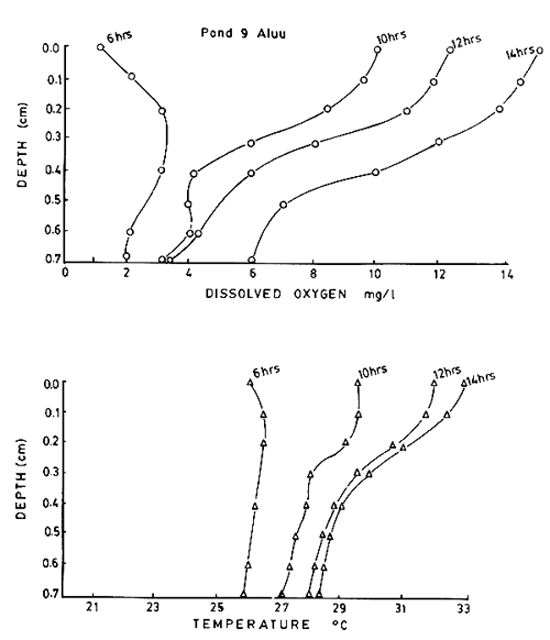
Fig. 8. Vertical distribution of dissolved oxygen and temperature at different times of day in pond 9 (fertilized, plankton development at peak) at Aluu. (From Ali, 1986).
Ambient oxygen acts as a limiting factor of metabolism and growth. Therefore while often oxygen level in the water body may be quite above the lethal point oxygen might be restricting (limiting) activity and growth.
With increase in level of ambient oxygen, metabolism increases up to a point (dependent zone) and beyond the critical point metabolism and growth are independent of oxygen - this relation will be evident only when maximal metabolism (active) is measured. When oxygen level is reduced to a point which can sustain only the basal requirement of the fish, it cannot afford any further physical activity at the specific oxygen concentration - hence referred to as the level of no excess activity of Fry (1947, 1971). Still below this level oxygen becomes inadequate and the fish dies due to asphyxiation (lethal or asphyxial level of oxygen) (Fig. 9). Do effect on growth is shown in Fig. 10.
Both the critical (limiting) level and the lethal level of oxygen vary markedly with fish species. Swimming activity and therefore migration of fishes and even feeding activity is restricted by critical oxygen concentrations. Salmonids (salmon and rainbow trout) stop swimming below 2–5 ppm oxygen but tilapia and carps swim at O2 level of 1–2 ppm (Kutty 1968; Kutty & Saunders, 1972). Generally tropical fishes have a high tolerance to low oxygen. Fishes which live in cold waters such as salmonids have a high lethal level of oxygen, but tropical fishes such as tilapias and carp tolerate oxygen levels even less than 1 ppm - these tolerance levels would change with temperature. There are some cases where fishes are known to survive in complete absence of oxygen. Blazka (1958) describes the crucian carp which survives under anoxic conditions in low temperature and Coulter records, certain cichlid fishes in deep African lake waters where oxygen content is nil or negligible. Many fishes such as carps have capacity to extract oxygen even at very low ambient oxygen levels.
Oxygen supersaturation also causes problems for fish culturists. When oxygen levels go high than saturation level, gas embolism or trapping of gas bubble in the blood stream takes place, causing death. It is said that minute particles of oxygen released by phytoplankton rather than the larger bubbles released by macrophytes cause more serious problem to fishes. The values of DO increase of oxygen solubility with depth of water (Table II) are thus useful for reference.
Oxygen becomes in some cases lethal, when too high, and acts as a depressant of metabolism. It is said that oxygen excess inhibits the activity of the respiratory enzyme, “succinic dehydrogenase”.
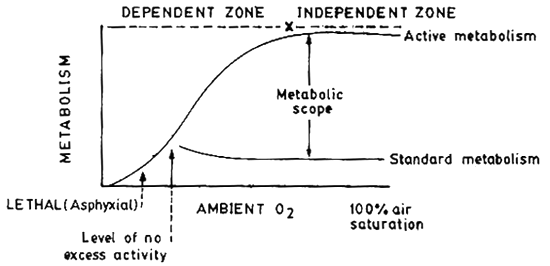
Fig. 9. Influence of ambient oxygen on metabolism of fish. The cross indicates the critical oxygen concentration below which the metabolism and growth of fish will be limited by the level of oxygen in water.
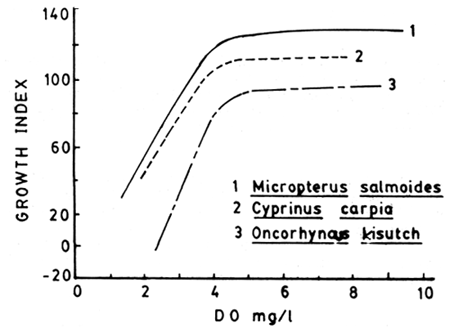
Fig. 10. Influence of ambient oxygen on the growth of 1) Micropterus salmoides. 2) Cyprinus carpio and 3) Oncorhyncus kisutch. (After Brett, 1979).
Dissolved oxygen requirements of fishes have been reviewed and discribed elaborately by Doudoroff and Shnmway (1970). We shall have occasion to refer to this again under “Species selection”.
For shrimp growth in brackish water ponds a minimum level of 2 ppm has been suggested. In Penaeus japonicus, according to Egusa (1961), stress is indicated by burrowing (burying in sand). In P. schmitti, most shrimps began swimming at the water surface when oxygen was reduced to 1.2 ppm. Subsequently (after 10 minutes) the shrimps began jumping out of water and fell to the bottom and became immobile. When the immobile shrimps were transferred to aerated water about £0% survived. Thus for this shrimp 1.2 ppm O2 appears to be the critical (lethal) level.
When exposed for long term in low oxygen (also thermal extreme) white areas of degenerated tissue develop in the tail muscle of shrimp, Penaeus aztecus which when kept in well-aerated water survived and the white areas disappeared. P. merguiensis also responded similarly in culture ponds.
In general it appears that in tropical shrimp ponds oxygen levels should not go below 3 ppm and below 1.2 ppm immediate death would occur. But in combination with other stress factors death would occur at higher oxygen. At 2.7 ppm Shigueno (1975) recorded die-off in the night time. Shigueno (1975) estimated oxygen consumption of various organisms and also bottom sand and water in shrimp pond and reported percentage consumptions:-
| CONSUMER | Percent O2 | consumed (%) |
| Penaeus japonicus | 8.6 | |
| Other shrimps | 0.5 | |
| Fish | 6.7 | |
| Bottom sand | 14.8 | |
| Water (bacteria/organic matter) | 69.4 | |
| TOTAL | 100.0 | |
Thus it could be seen that shrimps themselves account for only a small portion of the oxygen reduced, maximum share being by microorganisms and organic matter in the water and bottom sand and mud. Thus if the old water is drained and new water added at least temporarily much of the oxygen lack can be solved.
Heavy rains could cause stratification of water, the lighter freshwater would float and oxygen depletion could take place in the lower saline water. Therefore after heavy rains some provision for mixing water should be made, especially if there is not much wind.
![]()
![]()
![]()