![]()
![]()
![]()
por
M.R. Bowen 1, P. Howland 1, F.T. Last 2
R.R.B. Leakey 2 y K.A. Longman 2
INTRODUCCION
A juzgar por la manera en que se ha explotado comercialmente, Triplochiton scleroxylon (“obeche”) ha constituido un recurso natural valioso para muchos países del Africa occidental, desde Zaire, pasando por Camerún, Nigeria, Ghana y Costa de Marfil, hasta Sierra Leona y Guinea, donde se encuentran ejemplares apartados (Figura 1).
Como de costumbre, la corta indiscriminada raras veces se ha visto contrarrestada por esfuerzos enérgicos para lograr la regeneración natural o artificial. Con todo, hay que reconocer que en el pasado estos esfuerzos habrían tropezado con dificultades técnicas aparentemente insuperables, tales como, por ejemplo, la irregularidad de la fructificación y la imposibilidad de prolongar la viabilidad de las semillas almacenadas por mucho tiempo, requisito indispensable para mantener la continuidad y un programa de plantación año a año. No obstante lo dicho, los forestales, siguiendo el ejemplo de los horticultores, actualmente están considerando cada vez más el empleo de estacas arraigadas en sus programas de plantación (Hinds y Krugman, 1974; Kleinschmit y Sauer, 1976). Pero el impacto de la reproducción vegetativa no se limita a la producción directa de material de plantación, ya que puede influir mucho en nuestro método de selección y esto inevitablemente plantea la necesidad de clarificar los métodos que nos permiten distinguir entre las respuestas geno y fenotípicas.
El arraigo es reflejo del equilibrio hormonal y se supone que también lo es la floración. ¿Si se logra que las estacas de T. scleroxylon arraiguen, es posible inducir después la floración precoz en árboles relativamente pequeños en los que puedan hacerse fácilmente cruces controlados y en los que la incidencia de plagas y patógenos pueda minimizarse eficazmente? - Esto es virtualmente imposible en el terreno, donde la floración sólo suele tener lugar cuando los árboles han alcanzado 20 o más metros de altura.
A raíz de las observaciones hechas por nuestro ex colega Norman Jones, quien logró hacer arraigar algunas estacas, el Ministerio de Desarrollo de Ultramar del Reino Unido preparó un programa extensivo de investigación sobre T. scleroxylon, con la cooperación cordial del D.E. Iyamabo y del Dr. S.T. Olatoye, Directores sucesivamente del Instituto de Investigación Forestal de Nigeria (ex Departamento Federal de Investigación Forestal). Con su aliento, con la ayuda de sus colegas y con la facilitación de invernaderos y parcelas experimentales se ha venido desarrollando en los últimos 6 años un programa dividido en cuatro partes, a saber:
Figura 1: AREA DE DISTRIBUCION APROXIMADA DE TRIPLOCHITON SCLEROXYLON EN AFRICA OCCIDENTAL
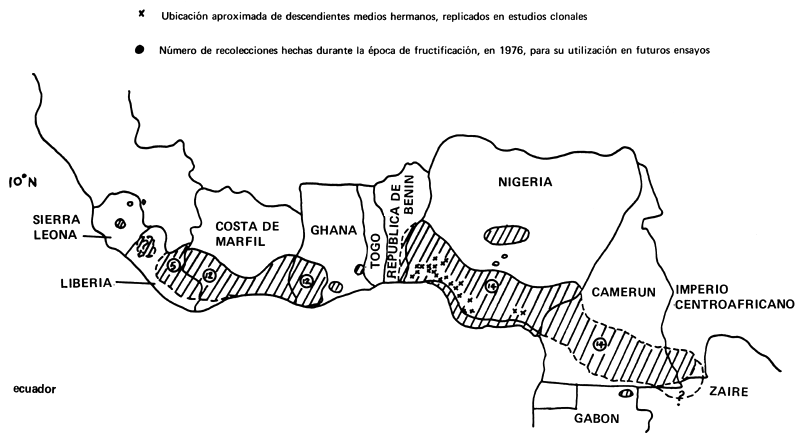
Nota: Cuando en el contorno de la parte sombreada hay una línea de trazos, la ubicación de T. scleroxylon es dudosa.
Cuadro 1: Efectos del almacenamiento a diferentes temperaturas de frutos verdes no secados sobre el tiempo que se demoran las semillas de T. scleroxylon en perder su capacidad de germinar.
| Temperatura, °C | ||||
| __________________________________________________^__________________________________________________ | ||||
| 0° | 10° | 20° | 30° | 40° |
| Tiempo (número de días) que se demoran los lotes de semillas en perder completamente su capacidad de germinar | ||||
| 41 | 26 | 20 | 12 | 6 |
Nota: 93 por ciento de las semillas eran viables al comienzo.
Cuadro 2. Germinabilidad de las semillas de T. scleroxylon tomadas de frutos secados hasta 14 por ciento de humedad y mantenidos a diferentes temperaturas de almacenamiento por 18 meses.
| Origen de las semillas | Porcentaje de germinación antes de su almacenamiento | Porcentaje de germinación antes de su almacenamiento a, °C | |||
| -18 | 0° | 6° | 25° | ||
| Arbol A | 82 | 78 | 51 | 27 | 0 |
| Arbol B | 76 | 72 | 27 | 11 | 0 |
| Promedio | 79 | 75 | 39 | 19 | 0 |
1) Viabilidad de las semillas almacenadas
En Nigeria las plantaciones de T. scleroxylon se han visto limitadas forzosamente a los años fructíferos, pero en los últimos años también se han recolectado semillas para programas experimentales destinados a determinar los efectos de diferentes factores operativos durante el almacenamiento. Disminuyendo la temperatura de 40°C a 0°C se retrasa de manera apreciable, 6 a 41 días, la pérdida de la germinabilidad (Cuadro 1), si bien experimentos posteriores revelan que estos plazos se pueden prolongar muchísimo, a saber, desde algunas semanas hasta varios años. La viabilidad se podría prolongar minimizando los ataques de los gorgojos (Apion spp.) y de los hongos (Mycosyrinx sp.), cosechando las semillas cuando los frutos están en sazón y haciendo descender lentamente su contenido de humedad hasta un 8 por ciento aproximadamente, para lo cual se recomienda una pérdida de peso durante el secado de 1 por ciento por hora. Preparadas de esta manera, el 39 por ciento de las semillas conservaron su capacidad de germinar después de estar almacenadas 18 meses a 0°C, pero a menos 18°C no se detectó ninguna pérdida con respecto al 79 por ciento de germinabilidad que tenían al iniciarse el experimento (Cuadro 2, Bowen in litt.).
2) Arraigo de estacas y manejo de las plantitas
Muy pocas estacas de T. scleroxylon arraigaban antes de 1972 (Britwum, 1970; Okoro, 1974), pero posteriormente han arraigado en gran número, en Ibadán y, en invernadero, en Edimburgo, estacas “tiernas” con hojas, con un solo nudo, de plantas jóvenes, mantenidas en atmósfera húmeda, empleando nebulizadoras automáticas o cubriéndolas con polietileno.
Se logró producir más de 60 000 estacas con raíces, para: (i) una serie de experimentos en el terreno y ensayos clonales con réplicas y (ii) el establecimiento de un banco de genes, pero es comprensible que todavía quede margen para mejorar y comprender mejor la fisiología de la iniciación de las raíces adventicias. ¿Por qué es necesario dejar algo de tejido de lámina de hoja (por ejemplo, 5 cm2) para lograr la supervivencia y el arraigo?
Un sustituto químico eficaz, la identificación de un co-factor (Hess, 1969), facilitaría el empleo de estacas sin hojas, que requieren un espaciamiento menor y favorecería también el arraigo de estacas leñosas “difíciles”, de árboles maduros.
Los clones de T. scleroxylon difieren en cuanto a su capacidad de arraigo. Los de algunos lotes de semillas invariablemente arraigan con mayor dificultad que otros. Es interesante observar que el número de estacas que arraigan, al parecer, no se ve afectado por la aplicación de hormonas, en Ibadán, pero en Edimburgo aumenta apreciablemente, habiéndose detectado interacciones significativas clon x hormonas (Howland, 1975; Leaky, Chapman y Longman in litt.). Por ejemplo, el clon 8036 respondió de manera semejante al ácido indol-butírico (AIB) y al ácido naftalén-acético (ANA), pero el clon 8021 sólo respondió al primero (Figura 2).
Los horticultores conocen de sobra la importancia del manejo de las plantitas. Las diferencias de manejo pueden explicar la diferencia entre las respuestas de Ibadán y las de Edimburgo. Un programa eficaz de reproducción depende esencialmente de la abundancia de estacas producidas uniformemente. En esta especie forestal que tiene un fuerte dominio apical, el desarrollo de brotes axilares se puede fomentar de muchas maneras y la más sencilla consiste en decapitar el tallo principal. La decapitación estimula el desarrollo de brotes axilares, pero debido a la reimposición del dominio, el rendimiento en material idóneo para estacas no aumenta mucho.
En el manejo de las plantitas hay que buscar el justo medio entre el empleo de plantitas tiernas que producen pocas estacas de fácil arraigo, y el de plantas de más edad que dan más estacas que arraigan con menor facilidad. Cuando se permite que las plantitas de semilla de T. scleroxylon crezcan sin obstáculos para obtener más estacas, se puede detectar fácilmente una pérdida de la uniformidad y de la arraigabilidad (véase Komissarov, 1969). En efecto, cuando se cortaron estacas algunas semanas después de la decapitación vertical de plantitas de semillas de un metro de altura, se observó que la arraigabilidad de los brotes axilares basales era de alrededor del 70 por ciento en comparación con 15 por ciento de las de los brotes apicales (Figura 3). La diferencia de arraigabilidad de los brotes basales y apicales disminuyó mucho cuando las plantas decapitadas se hicieron crecer horizontalmente o a un ángulo de 45°, pero una proporción significativa de los brotes apicales con raíces creció plagiotropicalmente (Leaky y Longman, 1976).
Es claro que el manejo es un asunto complejo y están por investigarse muchos aspectos, tales como la poda repetida para dar forma de arbusto con muchos troncos, como mata de té, los posibles efectos de cambio de fase y de envejecimiento que se producen a través de una sucesión de estacas jóvenes, los efectos de la nutrición … pero la aplicación de técnicas hortícolas uniformes (Howland 1975) ya ha permitido lograr progresos significativos. Estas técnicas se están aproximando a una etapa en la que podría estudiarse la posibilidad de hacer plantaciones comerciales, pero ¿es posible prever qué resultado darán las estacas de plantitas de semillas jóvenes en plantaciones maduras?
3) Selección clonal
El recurso a la reproducción vegetativa puede producir efectos trascendentales para la actividad forestal, porque el poder disponer de plantitas de vivero genéticamente uniformes, sin que se mezclen los carácteres como en la reproducción sexual, permite escoger los genotipos más idóneos para determinado lugar o para una aplicación en particular (Longman, 1976), si bien la pérdida de variación podría revelarse arriesgada. ¿Cómo respondería un bosque genéticamente uniforme a las presiones de la competencia cuando la luz y/o los nutrientes llegan a ser limitantes? ¿El empleo de un solo clon aumentaría el peligro de ataque por parte de los patógenos? En caso afirmativo ¿conviene hacer una plantación intercalada de muchos clones diferentes? La investigación de T. scleroxylon está en pañales, pero en un sitio de 30 ha, en Onigambari, cerca de Ibadán, ya se han plantado 230 clones de 27 lotes de semillas de medios hermanos empleadas en los siguientes experimentos:
ensayo de lotes de semillas (proveniencias) recolectadas en diversos sitios que representan diferentes zonas pluviales de Nigeria y
comparación de plantitas de semilla y de estacas derivadas de lotes de semillas iguales o diferentes. En esta primera etapa, en algunos casos, hay una diferencia sorprendente entre clones de un mismo genitor, pero en algunos casos quizá ésta pueda atribuirse a efectos “c” (Burdon y Sweet, 1976). En su conjunto estos experimentos forman un complejo de plantaciones de latifoliadas, probablemente único en el trópico, activo cuyo valor seguirá aumentando con el crecimiento de las plantas, hasta que ofrezcan una oportunidad más crítica para:
determinar la gama de variaciones dentro de T. scleroxylon,
estudiar en qué medida las primeras características de crecimiento pueden servir para prever la forma madura y
comparar el crecimiento de los clones y el de las plantitas de semilla.
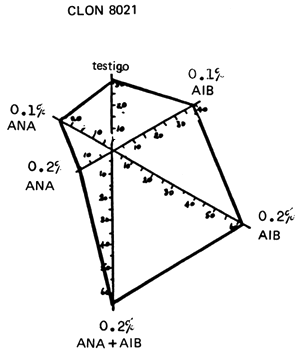 | 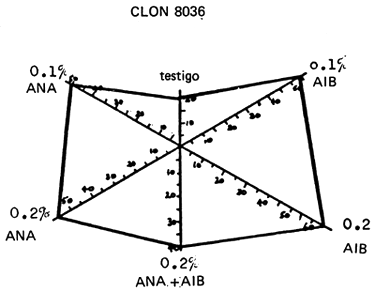 |
FIGURA 2: Diferencia entre las respuestas de arraigo (%) de 2 clones (8021 y 8036) de Triplochiton scleroxylon a diferentes cantidades de ácido indol-butírico (AIB) y de ácido naftalén-acético (ANA).
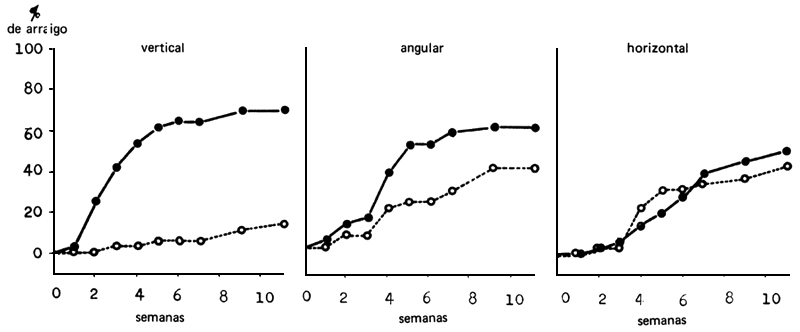
FIGURA 3: Arraigabilidad (%) de los brotes axilares de plantitas de semillas de Triplochiton scleroxylon hechas crecer horizontalmente, verticalmente, o a 45° (angularmente) después de decapitadas. (Los brotes axilares se dividieron en dos grupos - basales (●) y apicales (o).)
Estos experimentos requieren mucha mano de obra, ya que hay que hacer continuamente observaciones detalladas sobre muchos aspectos, en los árboles marcados. No sólo se trata meramente de las determinaciones tradicionales de la altura y del diámetro sino también de observaciones detalladas sobre el número de ramas, su posición y su dimensiones, la longitud de los entrenudos, la incidencia de las plagas … Ya se han hecho planes para suplementar el material de Nigeria con recolecciones hechas en Liberia, Costa de Marfil, Ghana y Camerún, y, además, se están preparando comparaciones críticas entre clones selectos y plantas de semilla no seleccionadas.
Los experimentos en el terreno son indispensables, pero suelen requerir largos plazos. Los experimentos hechos con plantitas de semillas parecen indicar que los tratamientos de decapitación provocan una respuesta fuertemente hereditaria y pueden tener relación con los resultados obtenidos en el terreno. Se están investigando estas respuestas, en las que el número de yemas axilares que producen brotes activamente después de la decapitación, difieren mucho de un clon a otro.
4) Floración inducida
La floración en los bosques raras veces se produce en árboles que tienen menos de 20 m de altura, o sea, cuando ya no resulta práctico proyectar programas extensivos de polinización controlada, si bien cabe abrigar esperanzas, ya que se han producido yemas de flores, en forma relativamente imprevista es cierto, en varios árboles jóvenes (de no más de tres años), nacidos de semilla, en Edimburgo. Esto ha permitido por lo menos identificar los clones que tienen propensión a florecer. Mientras tanto, en Ibadán, ocho injertos en maceta hechos con vástagos de árboles maduros florecieron prolíficamente y produjeron casi 1 000 flores sucesivamente en cuestión de meses, como sucede en los bosques. Se ha logrado avanzar algo hacia la floración controlada en Edimburgo, donde se han estimulado “brotes reproductivos” empleando varias técnicas combinadas.
Contrariamente a lo previsto, las flores de T. scleroxylon parecen ser autoestériles. Con los injertos floridos de Ibadán fue posible preparar una serie de polinizaciones autogámicas y cruzadas. Las autogámicas se malograron, pero muchas de las cruzadas produjeron semillas viables.
Con: (i) la identificación de clones precoces, (ii) la posibilidad de obtener flores de algunos injertos de árboles maduros y quizá también de plantas jóvenes y (iii) la casi certeza de que T. scleroxylon depende de la polinización cruzada, muchos de los aspectos esenciales de un programa de mejora genética están haciéndose menos problemáticos, pero todavía queda mucho por hacer antes de poder producir flores capaces de reproducirse a voluntad. Empleando material que se sabe tiene propensión a florecer, es indispensable hacer un estudio crítico de los factores ambientales y endógenos que provocan la floración.
CONCLUSIONES
Para tratar de: (a) obtener fácilmente estacas uniformes con raíces, (b) conservar el acervo genético de T. scleroxylon que aún queda y (c) emplear estacas y plantitas de semilla para la mejora genética, se decidió llevar T. scleroxylon al “laboratorio”. Aplicando métodos comprobados de horticultura, fue posible reproducir vegetativamente estacas tiernas con hojas, pero la necesidad indispensable de tejido de lámina sigue siendo inexplicable. Las estacas leñosas son todavía problemáticas. Se están comparando actualmente los resultados de las estacas tomadas de plantas nacidas de semillas de muchas recolecciones hechas en Africa occidental, y se espera además lograr comprender mejor las relaciones juvenil/maduro. Reconociendo los posibles peligros del empleo de poblaciones de estacas homogéneas, que por cierto se minimizarían plantando clones mezclados, el grupo está estudiando la posibilidad de lograr la mejora genética a través de una polinización controlada. Estas posibilidades se han materializado gracias a seis de años de estudio intensivo de T. scleroxylon. ¿Hay posibilidades semejantes en otras especies tropicales muchas de las cuales han arraigado en Edimburgo y en Ibadán (Cuadro 3)?
Cuadro 3: Arraigo de estacas tiernas con hojas en Ibadán, Nigeria y/o en Penicuik, Escocia, 1973–76
| Especies | Edad de la planta productora de estacas | Arraigo |
| Achras sapota | plantas de semilla de muy corta edad | fácil |
| Cedrela odorata | plantas de semilla de corta edad | fácil |
| Chlorophora excelsa | plantas de semilla de corta edad y brotes de árboles volteados | fácil |
| C. regia | brotes de árboles volteados | fácil |
| Cordia alliodora | planta de semilla de muy corta edad | muy fácil |
| Eucalyptus deglupta | planta de semilla de corta edad | fácil |
| Gmelina arborea | plantas de semilla de corta edad y brotes de árboles volteados | fácil |
| Khaya grandifoliola | plantas de semilla de corta edad | relativamente fácil |
| K. senegalensis | plantas de semilla de corta edad | relativamente fácil |
| Nauclea diderrichii | plantas de semilla de corta edad | fácil |
| Swietenia mahagoni | plantas de semilla de corta edad | fácil |
| Shorea albida | plantas de semilla de corta edad | fácil |
| Tamarindus indica | plantas de semilla de corta edad | muy fácil |
| Tectona grandis | plantas de semilla de corta edad | moderadamente fácil |
| Terminalia ivorensis | plantas de semilla de corta edad | fácil |
| T. superba | plantas de semilla de corta edad | fácil |
| Triplochiton scleroxylon | plantas de semilla de corta edad y plantas productoras de estacas, manejadas | fácil la mayoría de los clones |
BIBLIOGRAFIA
Bowen, M.R. (in litt.). Physiological studies for tropical tree improvement. I.
Britwum, S.P.K. 1970 Vegetative propagation of some tropical forest trees. Tech. Newsletter, F.P.R.I., Kumasi, Ghana, 4, 10–15.
Burdon, R.D. y Sweet, G.B. 1976 The problem of interpreting inherent differences in tree growth shortly after planting. In Tree Physiology and Yield Improvement, ed. by M.G.R. Cannel and F.T. Last, Academic Press, London, pp. 483–502.
Hess, C.E. 1969 Internal and external factors regulating root initiation. In Root Growth, ed. by W.J. Whittington, Plenom Press, New York, pp. 42–52.
Hinds, H.V. y Krugman, S.L. (Eds.). 1974 Vegetative propagation. (Proc. IUFRO mtg. Rotorua, 1973). N.Z.J. Forestry Sci., 4 (2), 119–458.
Howland, P. 1975 a - Vegetative propagation methods for Triplochiton scleroxylon K. Schum.;
b - Variation in rooting of stem cuttings of Triplochiton scleroxylon K. Schum;
c - Current management techniques for raising Triplochiton scleroxylon
K. Schum. Proc. Symp. on Variation and Breeding of Triplochiton scleroxylon
K. Schum, Ibadan, Nigeria.
Kleinschmit, J. y Sauer, A. 1976 Variation in morphology, phenology and nutrient content among Picea abies clones and provenances and its implications for tree improvement. In Tree Physiology and Yield Improvement, ed. by M.G.R. Cannell y F.T. Last, Academic Press, London, pp. 503–517.
Komissarov, D.A. 1976 Biological basis for the propagation of woody plants by cuttings. Israel Programme Sci. Transl., Jerusalem.
Leakey, R.R.B., Chapman, V.R. and Longman, K.A. (in litt.). Physiological studies for tropical tree improvement. II. Factors affecting root initiation in cuttings of Triplochiton scleroxylon K. Schum.
Leakey, R.R.B. y Longman, K.A. 1976 Root and Bud Formation in West African Trees. 2nd Annual Report, 1975.
Longman, K.A. 1976 Conservation and utilization of gene resources by vegetative multiplication of tropical trees. In Tropical Trees: Variation, Breeding and Conservation, ed. by J. Burley and B.T. Styles, Academic Press, London, pp. 19–24.
Okoro, O.O. 1974 A preliminary investigation of rooting of stem cuttings of Triplochiton scleroxylon. Fed. Dept. Forest Res., Ibadan, Res. Paper (Forest Series) 28, pp. 5.
 |  |
| Foto I: Arbol solitario de Triplochiton scleroxylon. | Foto II: Estaca juvenil de T. scleroxylon bien arraigada. |
 |  |
| Foto III: Flor y frutos inmaduros de T. scleroxylon. | Foto IV: Frutos maduros de T. scleroxylon. |
![]()
![]()
![]()