![]()
![]()
![]()
Skipjack are sexually mature in the Eastern Atlantic at a size of 45 cm (Cayré et Farrugio, 1986), that is, very early after their recruitment in the fisheries. They appear to have opportunistic reproductive behavior, very rapidly taking advantage of good feeding conditions when encountered in order to reach maturation and spawn (Cayré, 1985). This indicates that it is very difficult to distinguish between trophic and genetic migrations for Eastern Atlantic skipjack. The determinant of migration seems to amount to a permanent search for areas rich in food, and if these are also sufficiently productive and warm, they will also be spawning areas. The more or less regular periodic character of migration would be in fact induced by the cyclic availability of food (pelagic or mesopelagic micronecton, small pelagic fish…) in certain zones and certain seasons.
One can then interpret the east-west equatorial migration as the pursuit of high productivity zones suitable for the presence of micronecton, the food for skipjack (chapter 7). High primary productivity zones in the Gulf of Guinea are stimulated by a series of coastal upwellings staggered in time from Cape Lopez to Cape Palmas (paragraph 3.3). In regards to the Liberia zone, which is a particularly active reproductive zone from October to April (Cayré et Farrugio, 1986), the micronecton enrichment mechanism is not yet well identified. It could be linked to a subsurface primary productive mechanism specific to the intertropical Atlantic (Herbland, et al., 1983).
Otherwise, seasonal migrations in subtropical and tropical zones are apparently linked to the heating of surface waters that takes place during boreal or austral summers. Thermal fronts off Senegal and Angola (figure 3.2) play a role in determining skipjack concentration. However complex links are possible with certain coastal upwellings such as the Senegal-Mauritanian upwelling (Cayré and Roy, 1986).
From the examination of fishing data and from tagging conclusions, it is possible to develop a skipjack migration model in the Eastern Atlantic that takes into account both previously discussed determinants of migration. This model utilizes a series of compartments (figure 5.20) which are based on fishing data:
Strong or weak recruitment.
Presence of skipjack by size range, in large or small quantities.
Migrations established from tagging and fishing are indicated by arrows according to the following criteria:
In all cases, the continuous emigration toward the offshore areas, likely to occur from the middle of the exploitation phase, are not indicated in this figure to avoid overcrowding. The unit of time is one quarter of a year. Depending on the origin of recruitment and the rate of growth, skipjack can pass one (rapid tropical growth) to three years (slow equatorial growth) in the fisheries. The principal skipjack movements are organized according to a maximum cycle of three years, a cycle including the five major recruitments previously discussed. The most important of these recruitments takes place in the second quarter at Cape Lopez; it has been evaluated by Bard (1986) to be 90 million recruits. It seems in fact that these recruits appear in the first quarter in Angola (Cayré, 1985) where they are only exploited in certain years. Three complementary recruitments take place in Senegal, Cape Verde and the Liberia zone in the third and fourth quarter respectively.
The succession of concentrations, induced by coastal upwellings clearly marks the appearance of the large east-west migration, fed in large part by the recruitment of Cape Lopez. These skipjack measure between 40 and 50 cm, migrating to Liberia during the quarters 2, 3 and 4. In this last zone, an important reproductive activity takes place in the fourth quarter. Then these skipjack, measuring 50 to 60 cm, migrate in two ways:
One part, probably the most important, returns to Cape Lopez during the quarters 5 to 6 and restarts the same migration of the preceding year, toward the west in the quarters 6 to 8.
The other part migrates from the Liberia zone toward Senegal waters and probably more to the north toward the Canary Islands, during the quarters 5, 6 and 7. At the end of the year, they migrate in the opposite direction toward the south returning to the Liberian zone in quarter 8.

Figure 5.20 Migration model for skipjack in the Eastern Atlantic between seven compartments as a function of the quarters of three years. There is a large recruitment at Cape Lopez in the second quarter, a population which may be followed for 9 quarters. There is also a strong recruitment in Angola in the first quarter, which may be related to that of Cape Lopez, and three moderate recruitments the fate of which are not well known (particularly for Cape Verde).
Although a large part of skipjack from 40 to 50 cm migrate toward the west in the quarters 2 to 4, it is also probable that a smaller component stays in the Gulf of Guinea or migrates in the opposite direction (in quarters 3 and 4) toward Angola. At the time of quarters 6 to 8 they may rejoin the large migration toward the west along the African coasts from Angola or simply stay in the interior of the Gulf of Guinea between Cape Lopez and Cape Three Points. They may also reproduce in this zone.
In any case, after quarter 8, skipjack measuring more than 60 cm become rare in catches and emigrate toward the Central Atlantic. However another migratory cycle may be produced during the third year for a residual contingent of large skipjack that is still found at Cape Lopez.
The general pattern of skipjack migration is represented in figure 5.21. It should be noted that the bulk of skipjack catches in the Gulf of Guinea seems sustained by the recruitment off Cape Lopez.
The fate of complementary recruitment in the Senegal-Cape Verde and Liberia zones is less clear. Skipjack recruited off Senegal, tagged in the third quarter have returned to the Liberia zone in the following quarter, but no subsequent recaptures have occurred (Cayré et al., 1986). It is therefore possible that a part of these skipjack take the large migratory circuit of the Gulf of Guinea, to the migratory intersection in the Liberia zone. It is equally possible that they emigrate directly toward offshore areas after a brief stay in this zone. Skipjack recruited at the Cape Verde Islands seem to emigrate directly toward the west after a brief stay in this fishing zone during the third quarter. Some might, however, stay in the Liberia zone. The recently noted Liberia recruitment was the object of special tagging in 1984 (Bard and Amon, 1986). These skipjack first migrated toward the west and were never subject to long term recapture. It seems that this contingent emigrates toward the west, as was thought, after a brief stay in the fishery limited to the first quarter.

Figure 5.21 Pattern of migration of skipjack in the Eastern Atlantic. In this figure, the periods of passage are indicated by the rank of the quarters, counting arbitrarily from recruitment in the Angola zone in the first quarter. As there is an alternative course of migration, the periods corresponding to the second course are indicated in the dotted circles (after Cayré, 1985 and Bard, 1984).
No transatlantic migration of skipjack has yet been observed. However a gross calculation shows that if one takes into account the number of tags released, area of release (Gulf of Guinea, Senegal), and fishing intensity for skipjack in the Western Atlantic, there would be practically no chance for an Eastern skipjack to be recaptured in the Western Atlantic fisheries. However, exchange of large skipjack might exist between the Eastern Atlantic fisheries and those of the Western Atlantic, but at an unknown rate.
Skipjack migrations have been studied in considerable detail which contrasts to the gaps in yellowfin and bigeye studies. This contrast is due to international research effort specifically on this species. Thus a series of conclusions useful for the evaluation of catch potential for skipjack in the Eastern Atlantic are available is available.
These conclusions are:
Eastern Atlantic skipjack from a single stock exploited from Senegal to Angola for a maximum of three years. These skipjack are recruited starting at 35 cm and remain in the fisheries until a maximum size of 70 cm. However they have a precocious tendency to emigrate toward the central Atlantic, varying according to recruitment. This tendency is very marked for skipjack whose size is greater than 60 cm.
There are five principal recruitments currently identified in equatorial and tropical zones. The principal recruitment takes place at Cape Lopez in the second quarter of the year. These skipjack are those exploited for the longest period of time in the Eastern Atlantic. Their migratory movements are cyclical along the coasts of Africa between Senegal and Angola. As a result of the duration of their stay in the fisheries, this recruitment seems to furnish the majority of skipjack catches in the Eastern Atlantic.
The other recruitments concern skipjack that stay only for a short time in the fisheries and disappear toward the west. They constitute a fraction of stock which is probably lightly exploited.

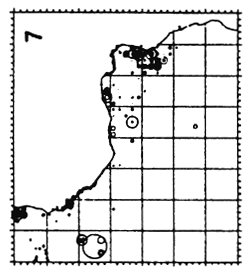
Figure 5.22 Migratory trajectories of all bigeye tagged from 1970 to 1984 in the Eastern Atlantic. Only displacements greater than 300 miles in a straight line for times at liberty greater than 30 days have been considered.
Bigeye is a large size tuna reaching 200 kg at around 200 cm. Its growth is rapid (chapter 6) and its habitat varies depending on the different phases of its development, from the equatorial zone for juveniles to subtropical latitudes for large individuals. One can consider a priori bigeye to be a very mobile tuna, probably the most migratory of tropical tuna.
Bigeye are exploited in the Atlantic by purse seiners and pole and line boats for small and medium sizes, and by longliners for large sizes (chapter 4). Biologically speaking, information on this species seems very limited (chapter 6). The three size classes that will be distinguished for studying apparent migrations of Atlantic bigeye, will be based on size frequencies observed in catches rather than on actual biological criteria. The limits of these classes might be somewhat modified by better knowledge of biological events marking the life cycle of bigeye.
The three size classes that will be used are:
The juvenile phase from 30 to 70 cm (1.5 to 8 kg). These bigeye are fished along with juvenile skipjack and yellowfin in the Gulf of Guinea.
The preadult phase from 70 to 100 cm (8 to 20 kg), composed of individuals fished primarily in the north subtropical and tropical zones.
The adult phase, greater than 100 cm (greater than 20 kg) composed of individuals caught in all of the tropical and subtropical Atlantic.
Table 5.4 Distances traveled by tagged bigeye in 100 mile classes as a function of time at liberty (in 3 month
classes). The average gross speed of displacement is 2.81 miles per day and the corresponding
dispersion coefficient 6,741 miles2 per day.
Number of bigeye originally marked = about 8,000. Number of recaptures = 544.
| DISTANCE | 100 | 200 | 300 | 400 | 500 | 600 | 700 | 800 | 900 | 1000 | >1000 | |
| TIME | ||||||||||||
| 0–3 | months | 267 | 76 | 20 | 40 | 4 | 5 | 2 | 3 | 1 | 3 | |
| 4–6 | months | 20 | 27 | 15 | 5 | 4 | 1 | 1 | 1 | 1 | 4 | |
| 7–9 | months | 1 | 5 | 1 | 1 | 1 | 1 | 3 | ||||
| 10–12 | months | 1 | 1 | 2 | 1 | 1 | 2 | |||||
| 13–15 | months | 4 | 4 | 1 | 1 | 3 | ||||||
| 16–18 | months | 4 | 1 | 1 | ||||||||
| 19–21 | months | |||||||||||
| 22–24 | months | 1 | 1 | |||||||||
| 25–27 | months | 1 | ||||||||||
| 28–30 | months | |||||||||||
| +30 | months | 1 | ||||||||||
As for yellowfin, it is interesting to first examine maps of the gross distribution of rectilinear displacements by all tagged bigeye in the tropical Atlantic, without regard to size (figure 5.22). Of around 8,000 tagged bigeye, 587 have been recaptured (7.5%) after a variable period at sea. With exception of only 3 individuals, all recaptures have been made in the tropical or equatorial Atlantic between 20° N and 20° S and to the east of 30° W. Examination of distances covered in relation to time at liberty (table 5.4) shows rapid long distance displacements (up to 1300 miles in 2 years). The average displacement speed of bigeye is somewhat higher than that of yellowfin and comparable to that of skipjack.
Bigeye tagging results could not until now be studied in relation to fishing effort due to a lack of data concerning fishing effort directed to this species by purse seiners or pole and line boats and because of small number of recaptures (n=587).
As with yellowfin and skipjack, bigeye are recruited between 30 to 40 cm in the purse seine and pole and line fisheries of the Gulf of Guinea. They are then fished communally in mixed schools with juvenile skipjack and yellowfin. Bigeye are in general poorly identified in tuna boat log books as well as in commercial unloading statistics. The geographic distribution of bigeye catches of various size classes can only be determined from sampling.
The geographic distribution of monthly catches (average from 1979 to 1983) of juvenile bigeye (figure 5.24) is similar to that observed for skipjack and juvenile yellowfin with some specific differences:
juvenile bigeye are present all year in the equatorial zone and more seasonally in boreal and austral summer in the north and south tropical zones respectively.
large catches take place in concentrations which are particularly well defined in time:
the Cape Lopez concentration from April to September,
the Liberia concentration from November to February.
a zone between 5° N and 10° N off Sherbro Island seems quite specific to catching juvenile bigeye, more than any other species, between April and May and from September to November.
seasonal catches of juvenile bigeye are also observed off Senegal and Mauritania. These catches, quite modest compared to those of yellowfin or skipjack, are limited to the estival season (July to November) and closely linked to a thermal front that stabilizes in summer between 20° and 21° N (Cayre and Diouf, 1986).
juvenile bigeye catches have also been observed in Angola where they appear in scientific samples from Angolan pole and line boats (although the bigeye are absent from statistical declarations of this country).

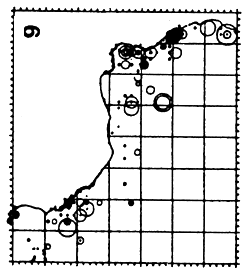
Figure 5.23 Migratory trajectories of juvenile bigeye recovered at less than 70 cm for all tagging conducted from 1970 to 1984 in the Eastern Atlantic. Only displacements greater than 30 miles in a straight line for times at liberty greater than 30 days have been considered.
The observed gross trajectories of tagged and recaptured juvenile bigeye during 1972 – 1974 (figure 5.23) suggests a displacement pattern similar to that observed for skipjack and juvenile yellowfin with:
large lateral migrations that follow the African coast from Cape Lopez to Senegal passing the zone situated off Liberia.
some migrations toward Angola
Two major migrations emphasize the very migratory character of this species even at these small sizes:
a very rapid migration (10 miles/day) from Liberia to the Azores
a very long migration by a bigeye from the Congo to a point situated in the central Atlantic (5° N to 44° W) where it was recaptured by a longliner.
These two observations seem to indicate that bigeye may emigrate precociously out of the Gulf of Guinea. This has not been observed for either yellowfin or skipjack of similar size.
In conclusion, juvenile bigeye seem to move rapidly from Cape Lopez in the second quarter to the Liberia zone in the fourth quarter. Some may also reach the Sherbro zone then Senegal in the third quarter. The two very distant recaptures observed in the Azores and in the central Atlantic suggest a hypothesis of precocious emigration (in the first years of their life), of a group of bigeye out of the Gulf of Guinea. This trend of precocious emigration of bigeye can be related to the high growth rate of this species, a rate superior to that of yellowfin or skipjack (chapter 6). To confirm this migratory schema, it would be necessary to analyse existing tagging results in more detail in relation to fishing effort and especially to carry out new tagging capable of generating a significant number of recaptures. Moreover supplementary tagging in certain zones, such as the one situated off Liberia, seem equally necessary.
 |  |  |
 |  |  |
 |  |  |
 |  |  |
Figure 5.24 Monthly geographic distribution of juvenile (less than 70 cm) bigeye by FIS and Spanish purse seiners and baitboats from 1979 to 1983 in the Eastern Atlantic.
As shown in the average monthly geographic distribution map of bigeye catches (figure 5.25), the catches come from concentrations limited in time and space:
Cape Lopez concentration from April to September
Liberia concentration from November to January
The distribution of preadult bigeye catches by purse seiners is, in general, comparable to that of preadult yellowfin; these catches seem however to be more scattered. For juvenile bigeye, there are a certain number of occasionally large catches that seem to be characteristic of this species. The preadult bigeye catches by pole and line boats of Dakar are quite distinctive. They are mixed with yellowfin of the same size between May and November, in the sectors close to the coasts of Senegal and Mauritania and seem tied to the south-north displacement of a thermal front that may reach Cape Blanc.
The track of the gross migratory trajectories (rectilinear and without a fishing effort correction) of bigeye, tagged generally in the juvenile phase but recaptured in preadult phase (figure 5.26), shows two types of migrations:
long distance migrations that link bigeye from the Gulf of Guinea with those of Senegal, Sherbro and Angola. This confirms the highly migratory tendency of this species already demonstrated for juvenile bigeye. The recapture by a longliner in the south of Angola (2° S, 11° W) of a fish tagged south of Cape Lopez is particularly notable.
a series of short trajectories by tagged bigeye in the Cape Lopez and Senegal zones.
Although the short trajectories observed in the Senegal zone seem to correspond well with local movements of the thermal front already indicated, those observed in the Cape Lopez zone are harder to interpret. Eight of these trajectories, in particular, were executed by bigeye recaptured one year after tagging. It is therefore possible that these trajectories correspond to an annual cyclic migration or they simply result from a seasonal exploitation of this sector by tuna boats.
Preadult bigeye appear to migrate like juveniles between Cape Lopez, Angola, Sherbro Island and Senegal. Emigrations toward the north and south tropical zones appear likely, even if one group of preadult bigeye seems to have a cyclic migration that carries them to Cape Lopez after a year. All these detailed observations require confirmation and more detailed analyses linked to supplementary tagging are necessary.
As was mentioned in paragraph 5.2.3.1., catches of large bigeye are made mostly by longliners for which fishing effort is distributed throughout the Atlantic. However, purse seiners and pole and line boats of Dakar catch large adult bigeye occasionally.
Figure 5.28 shows the average monthly geographic distribution (1979-83) of these catches by FIS and Spanish purse seiners and pole and line boats; their distribution is as follows:
most of the catch is taken off Cape Lopez from May to July by purse seiners only and off Senegal and Mauritania from April to December by both pole and line boats and purse seiners.
smaller catches take place in September off Angola.
as with the two other bigeye size classes (juveniles and preadults), large and isolated catches of large bigeye are occasionally observed in various places of the fishery and notably off the Côte d'Ivoire.
Catches of large bigeye by pole and line boats of the Canary Islands, Madeira and the Azores are coastal and seasonal. The major fishing season of the Canary Islands extends from March to May, although there are catches all year round (Santos, 1977). At Madeira, fishing is also practiced all year, but with a maximum in May (Carvalho et al., 1983). In the Azores, fishing is actually seasonal from April to July (Pereira, 1983).
 |  |  |
 |  |  |
 |  |  |
 |  |  |
Figure 5.25 Monthly geographic distribution of preadult (from 71 to 100 cm) bigeye by FIS and Spanish purse seiners and baitboats from 1979 to 1983 in the Eastern Atlantic.
 |  | ||
| Figure 5.26 | Migratory trajectories of juvenile bigeye recovered at the sizes of preadults (71 to 100 cm) for all tagging conducted from 1970 to 1984 in the Eastern Atlantic. Selection criteria identical to those of figure 5.23. | Figure 5.27 | Migratory trajectories of juvenile bigeye recovered at greater than 100 cm for all tagging conducted from 1970 to 1984 in the Eastern Atlantic. Selection criteria identical to those of figure 5.23. |
Asian longliners have searched preferentially for bigeye for a dozen years (1975), as evidenced by more and more frequent use of longliners especially known for deep fishing (250–300m). The annual geographic distribution of the longline catches (figure 6.24) as follows:
a general concentration of catches in the east part of the Atlantic with two zones of particularly large catches situated respectively between 5°N and 20°N from the coast to 35°W, and between 5°S and 20°S from the coast to 5°W.
three zones with low catch rates situated respectively from one end of the equator to the other between 5°N and 5°S, in the north subtropical zone between 30°N and 45°N, and in the south subtropical zone between 25°S and 40°S. The Gulf of Guinea is part one of the large zones of very moderate catches.
The quarterly distribution of catches in different zones (chapter 4) shows that the high density fishing zones of the north tropical areas exists all year round, but oscillates between 10°N and 20°N according to season. The south subtropical zone between 25°S and 40°S, where fishing is moderately intense, exists only in austral summer. These different observations lead to the conclusion that adult bigeye make large seasonal migrations.
With regard to tagging, adult bigeye recaptures are rare and can hardly be subjected to detailed analysis (figure 5.27). The four recaptures observed off Cape Lopez and the one observed off Senegal correspond to bigeye tagged at preadult sizes and recaptured around a year after tagging. The small distances apparently covered during this year of liberty may confirm the hypothesis of an annual cyclic migration, a hypothesis previously suggested for preadult bigeye. The absence of adult bigeye recaptures by longliners must be emphasized.
 |  |  |
 |  |  |
 |  |  |
 |  |  |
Figure 5.28 Monthly geographic distribution of adult (greater than 100 cm) bigeye catch by FIS and Spanish purse seiners and baitboats from 1979 to 1983 in the Eastern Atlantic.
It is currently very difficult to link the small catches of adult bigeye by surface gear in the Gulf of Guinea with the large amount of fishing by longliners in the rest of the Atlantic. It seems in fact that a majority of adult bigeye have emigrated at the juvenile or preadult state from the interior of the gulf toward the tropical and subtropical zones. Longline fisheries show a very wide distribution of these adult bigeye that have probably undertaken very large seasonal migrations on an Atlantic scale. However these adults must return to the equatorial zone to reproduce, as this zone is the largest spawning zone known at this time (paragraph 6.1.1.). The continued recent development of deep longline fishing should permit more exact studies on this subject. Tagging of large bigeye would also be very useful.
If one accepts that bigeye of less than one meter are frequently sexually immature, the observed migrations of these individuals from the Gulf of Guinea toward the north and south tropical zones are very probably linked to a search for food. One can then compare these migrations to those of skipjack or juvenile yellowfin that seem linked to the successive development of different coastal or equatorial upwellings. The bigeye migration in the north tropical zone in boreal summer, follows the same pattern with an amplitude greater than that observed for yellowfin since young bigeye may at times go all the way to the Bay of Biscay (40°N).
The spawning areas of adult bigeye determined by analysis of gonado-somatic indices and larval distribution, seem situated in a vast equatorial zone (15°N – 15°S) crossing the entire Atlantic. Moreover bigeye larvae are found jointly with those of yellowfin in waters where the temperature is equal or superior to 24° C (Caverivière and Suisse de Sainte Claire, 1980). Finally adult bigeye caught at the Azores Islands are always totally immature (Pereira, pers. com.)
It seems probable that adult bigeye, in order to reproduce, make major migrations between north and south subtropical latitudes and the equatorial reproductive zone. However the frequency as well as the fraction of the Atlantic bigeye population which undertake these migrations is totally unknown. Furthermore the existence and magnitude of exchanges between adults from north and south zones into the equatorial zone is also totally unknown. These different points are essential in managing the fisheries.
After examination of all of the available data, one can only propose a very schematic migration model for this species; this model implies several more or less well founded hypotheses.
The analysis of catch and tagging data nevertheless supports the hypothesis that there is a single unique stock of bigeye in the Atlantic Ocean. The principal facts which favor this hypothesis are:
existence of a unique nursery area for juvenile bigeye in the Gulf of Guinea.
emigration of bigeye away from this nursery to the north as well as to the south, at occasionally very long distances (Azores and south of Angola).
It is not possible to conclude with certainty as to the unidirectional or cyclic character of juvenile and preadult bigeye emigrations from the Gulf of Guinea in the direction of high north or south latitudes. Taking into account different hypotheses and uncertainties, it has only been possible to establish a schematic representation of bigeye migrations in the Atlantic (figure 5.29).

Figure 5.29 Migratory pattern of Atlantic bigeye.
Bigeye migrations remain poorly known which is paradoxical as they seem to be more important than those observed for other tropical tuna. The reasons for this lack of knowledge is due in large part to the fact that, for many years, bigeye was considered as an accessory species, often confused with yellowfin by fishermen as well as by scientists; as a result, for a long time there was a large inaccuracy in statistical data concerning this species. This situation is not unique to the study zone, since in almost all the fishing zones in the world, the problem of identification of young bigeye is poorly resolved and tagging has been very rare. Only 1000 bigeye have been tagged outside of the Atlantic according to Hunter et al. (1986). For some years, an increase in bigeye fishing in the Atlantic has been observed, notably by longliners. The renewed economic interest in this species should logically stimulate research effort on bigeye.
Migration of small tuna and related species in the zone has been studied very little. The classic methods for determining fish displacements are either used very little for these species (tagging), or are insufficient because of the mediocre quality of data bases (data linked to the exploitation and biology of the species). Nevertheless observations on geographic distribution of the species by different stage of development and analysis of seasonal variations of declared catches by species, allows clarification of the principal periods of presence of each species in different zones. The only tagging results available in the study zone concerns spotted tunny and Atlantic bonito, but these results only concern coastal stocks exploited essentially in Senegal by artisanal fisheries.
Distribution of larvae and juveniles
Larvae are distributed from Mauritania to Angola. They are more coastal but can also be frequently collected in the high seas. Collected at the surface during the day, they can be collected at depths of 50 m or more during the night (Zhukova, 1971). Juveniles, under 30 cm are very coastal and fished by beach purse seines in the Côte d'Ivoire (Marchal, 1963) and Senegal (Diouf, 1980).
Distribution of adults
Adult spotted tunny present a distribution that seems more or less continuous from Morocco to Angola. Various observations made of these individuals show that adults are fished at the surface, on and slightly beyond the continental shelf. They can descend to 150 m in depth (Postel, 1955). Preadults under 40 cm are coastal. Large spotted tunny may approach the coasts of the Côte d'Ivoire (Marchal, 1963) and Senegal (Diouf, 1980) during certain seasons.
Tagging
Tagging programs in Senegal since 1981 have released 1190 individuals (Diouf, 1985). The first recaptures, around 30 individuals, have demonstrated displacements on either side of Cape Verde in the warm season (May-October). Champagnat et al., (1979) have recaptured in January, at Dakar, a spotted tunny tagged in November at Nouakchott. There seems to be a displacement of spotted tunny in both directions Mauritania - Senegal. Rey and Cort (1980) have shown that spotted tunny move from the western Mediterranean toward the north-east Atlantic by passing through the Strait of Gibraltar.
Sasonal variations of catches
In Mauritania, available statistics show that spotted tunny are abundant in September. In Senegal, the spotted tunny is present all year round in the Cape Verde zone, but on the continental shelf as a whole, the most important fishing takes place from January to June (Diouf, 1980). Marchal (1963) notes that catches in the Ivory coast are significant in February but the spotted tunny are present all year on the coast. In Benin, tuna in general are present from January to March and from September to December. The analysis of data transmitted to the ICCAT shows that in Ghana, the catches can be significant; in Angola the maximum catches are from February to May. Data obtained on French purse seiners (Stretta, com. pers.; Bard and Kothias, 1984) and on Spanish purse seiners (Diouf and Rey, 1985) derived from the Atlantic coast of Africa show that spotted tunny is present all year from Mauritania to Angola; these data do not permit the study of the seasonal variations of this distribution.
Analysis of size frequencies
Chur (1975) has studied the size variation of spotted tunny caught from Mauritania to Angola. He found moderate sizes, the largest from the south of Cape Blanc to Sierra Leone. At the south of this zone, individuals have a medium size under 50 cm. Diouf (1980, 1985) has shown that large spotted tunny (over 60 cm) are present in Senegal from January to June; around Cape Verde, individuals of intermediate size (50 – 60 cm in FL) are poorly represented in catches while they are present in larger proportions in Sierra Leone and the Côte d'Ivoire (Chur, 1975; Marchal. 1963)
Distribution of larvae and juveniles
Auxid larvae are very abundant in the eastern Atlantic. They are collected at the surface, near the coasts, and also along the edge of and over the continental shelf. There are very few data on juveniles. However, in Senegal, individuals under 30 cm are fished in the Baie de Gorée in the warm season (July-August), Diouf (1985).
Distribution of adults
Auxid adults are present from Mauritania to Angola. In Senegal, individuals between 30 – 35 cm are caught near the coasts beginning in May, but the spatial distribution of auxid catches in the Eastern Atlantic seems to indicate that adults are also oceanic (Diouf, 1985).
Tagging
Very little tagging has been done in the zone. Off Dakar, only 8 individuals have been released and not one recapture has been recorded.
Seasonal variation in catch
On the African coasts of the Atlantic, auxids are fished all year round (Diouf and Rey, 1985). The species is very common in purse seiner landings, but catches in different zones are very variable from one year to another and do not permit the clear definition of fishing seasons. In Angola however, catches are essentially taken in the first and fourth quarter. Although essentially coastal several years ago (1979), catches seem to extend progressively toward the high seas, as for yellowfin and other major tuna.
Analysis of size frequency
Sizes of auxids fished in the eastern tropical Atlantic vary from 30 to 50 cm. Sizes of the most numerous individuals in the catches are between 35 and 45 cm. In the Gulf of Guinea it is mostly individuals of more than 35 cm that are exploited; off Cape Lopez, individuals over 40 cm are predominant in the catches. The individuals trolled near Dakar are between 30 and 35 cm (Diouf, 1985).
Distribution of larvae and juveniles
Little information is available in the Eastern Atlantic. Rodriguez Roda and Dicenta (1980) have found Atlantic bonito larvae in the north-east Atlantic on the Morrocan coasts at distances from a few miles to around 10 miles from the coast. Elsewhere in the Eastern Atlantic, there have been no reports of larvae collection.
Distribution of adults
In Senegal, the Atlantic bonito is fished at the surface on the continental shelf where it scarcely passes the 120 m isobath. It has never been reported in the high seas by tuna purse seiners operating in the Eastern Atlantic from Mauritania to Angola. In Senegal, from June to November, individuals seem to return to the coast (Diouf, 1980). Preadults under 40 cm are more coastal.
Tagging
Tagging programs in Senegal in May-June 1985 released 394 Atlantic bonito (Diouf, 1985). Some recaptures occurred in the vicinity of the tagging zones in June and to the south of the Mauritanian coasts in September.
Rey et al., (1980) showed that there is mixing between Atlantic bonito populations from the western Mediterranean and from the north-east Atlantic. This species can have migrations of large amplitude within distribution. The average distance covered is around 2.6 km/day but can vary from 0.2 to 35.6 km/day, (Rey et al., 1980).
Seasonal variation in catch
The massive appearances of Atlantic bonito on the Senegalese continental shelf take place from January to April. From May-June the species becomes rare. It disappears until October then certain isolated individuals reappear starting in November (Diouf, 1980). In Ghana, Atlantic bonito fishing has been reported without other information. In Angola, declarations made to the ICCAT report large catches in the first and fourth quarter.
Analysis of size frequencies
Size frequency data are only available in Angola and Senegal. Sizes of individuals caught by Angolese pole and line boats range from 30 to 60 cm. The most numerous classes are between 40 and 50 cm. The smaller classes arrive in the fisheries in the fourth quarter. In Senegal, sizes caught range from 30 to 65 cm. As in Angola, individuals of the 45 – 50 cm classes are more numerous in the samples from January to May. Beginning in June, small individuals of average size 35–40 cm are landed. Trollers exploit individuals 30–45 cm from January to June near the coasts.
Distribution of larvae and juveniles
Little information is available in the zone but it shows that larvae, essentially coastal, can be caught in the high seas, probably carried by the currents. Larvae are collected at night at depths of around 100 m or during the day in the surface layer to around 50 m in depth (Zhudova, 1969). The distribution of post-larvae and juveniles from 40 to 220 mm is very unknown. However, a significant number of these individuals is caught by beach purse seine in Senegal during the warm season (Diouf, 1980).
Distribution of adults
The species is present in the coastal zone from Mauritania to Angola: In Senegal, catches of Western African Spanish mackerel are made at the surface mainly at depths under 40 m (Diouf, 1980). Zhudova (1969), estimates that adults can abandon their coastal habitat for the high seas at certain periods.
Tagging
No data in the study zone.
Seasonal variations of catches
Off the Senegalese coast, the largest catches are made in the warm season (Diouf, 1980). In Mauritania, available statistics reveal that the catches reach a maximum in March (Bergerard et al., 1981). In the Gulf of Guinea information is quite insufficient, but relatively large surface catches are declared in Ghana and off the Eastern Atlantic coast by the USSR.
Analysis of size frequencies
Size data on exploited individuals is only available for Senegal within the distribution zone. The majority of catches occur at the depths from 20 to 25 m and are composed mainly of fish between 40 and 70 cm. Beach purse seines catch juveniles (10–20 cm at FL) and preadults under 30 cm during the warm season near the coasts.
In Senegal, massive appearances of Spanish mackerel occur in the warm season; however some catches are in the south of Senegal in waters where the temperature is between 18°C and 21°C (Diouf, 1980). Spanish mackerel is fished abundantly by beach purse seines from July to November, the period during which a relatively important local desalinization (to the order of 34‰) is registered in the Senegalese coastal fringe. Postel (1955) notes catches in Guinea in waters where the salinity is very low (31.8‰). The species is present in the Côte d'Ivoire lagoon (Aldrin et al., 1972) and in the Lagos lagoon (Fayade and Olaniyn, 1974). Postel (1955) notes that “the rising of cold waters that characterizes the north littoral of the Spanish Sahara from one end of the region to Walvis Bay on the other, imposes barriers that S. tritor can not circumvent in the high seas because of its constraint to strictly coastal zones. This explains its absence in regions like Morocco where conditions necessary for its presence occur during summer.” Zhudova (1969), from the collection of small larvae (3.5 to 5.8 mm) more than 1000 miles from the coast, suggests that adults of this species may, at certain periods, abandon their coastal habitat for the high seas.
Different observations show that the spanish mackerel is an essentially coastal species presenting a wide salinity tolerance, but appears sensitive to temperature which must be above 17°C.
The plain bonito is a coastal species forming schools around 20 m in diameter inside the 30 m isobath (Postel, 1956). In Senegal, adult plain bonito fishing is carried out from December to June, while the younger individuals are fished in August-September (Maigret, 1973). Adults are fished in waters where the temperature is under 20°C, while the young seem more tolerant to this factor.
There is no information even though the species is present and exploited in the zone.
Migrations of small tuna in the study zone for the most part remain unknown; the simple description of catches by size is very incomplete because of mediocre fishing statistics; tagging and recapture data are totally insufficient to evaluate the possible migrations of individuals exploited by coastal fisheries of the region.
![]()
![]()
![]()