![]()
![]()
![]()
The theoretic and analytical bioeconomic framework for fisheries management developed in Chapter 2 assumed the exploitation of a single stock by a homogeneous fishing fleet. However, sharp changes in the abundance of targeted and nontarget species, and also in the relative species composition of exploited communities, have been detected worldwide, as a result of a growing fishing intensity (see examples in Mercer, 1982; May, 1984; Pauly & Murphy, 1982). Overfishing has been observed, with a progressive decrease of the most valuable species commonly corresponding to the highest levels of the trophic chain (Pauly, 1982; Bustamante & Castilla, 1987). This has led to recognize that, in practice, fishing affects a multispecific resource (May et al., 1979), harvested incidental or intentionally by fleets with different size, fishing power and gears (Gulland & Garcia, 1984; Alverson et al., 1994; Anonymous, 1995; Hall, 1996, 1998).
In the last two decades, fishery scientists have taken a more holistic approach to management (Caddy & Sharp, 1986; Ströbele & Wacker, 1991; Seijo et al., 1994c). For this purpose, research has been directed to evaluating the ecological and technological interdependencies, as well as the influences of the physical environment. An ecological interdependence is defined as occurring when two stocks have a competitive or a predator-prey relationship (Anderson, 1975a, b; Mitchell, 1982; Seijo & Defeo, 1994a; Mesterton-Gibbons, 1996). It can also account for intraspecific interactions (e.g. recruits and adults: Defeo, 1998). A technological interdependence occurs when fleets with different fishing power (e.g. artisanal and mechanized fleets) and/or gear types, operate on different components of a single stock or on different target species, affecting their abundance in a dissimilar form (see e.g., FAO, 1978; Clark, 1985; Huppert, 1979; May, 1984). The fleet concept is here used in a wide sense, including hand-gathering techniques in artisanal coastal fisheries.
Technological interdependencies can arise from the activity of fishing fleets with different fishing power and even different effort costs, over a single stock. The sustainable yield of two fleets (e.g. artisanal and industrial) can be represented as follows:

where Y1(t) and Y2(t) are, respectively, the yield of the fishing fleets 1 and 2 in time t.
Under equilibrium conditions dΒ /dι=0. Then:

Thus, the sustainable yield of fleet 1 can be expressed as:

From Chapter 2 we know that:
Y1=q1f1 B (3.4)
Y2=q2f1 B (3.5)
Solving for B in equation (3.2) and substituting Y1 and Y2 by expressions (3.4) and (3.5):

Substituting (3.6) in (3.4) and (3.5), the yield for each fleet will be:
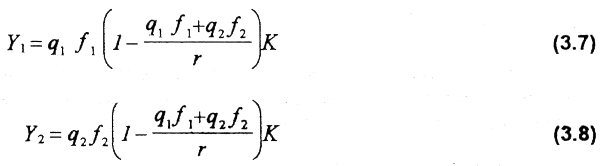
and the corres[ponding economic rent:
π1 = pY1 - c1f1 (3.9)
π2 = pY2 - c2f2 (3.10)
The bioeconomic equilibrium BE for e.g. fleet 1occurs when π=0. Substituing (3.7) in (3.9):
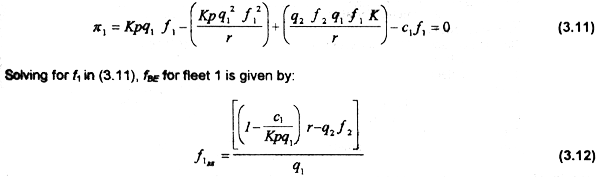
The MEY for fleet 1 can be ontained by taking the first derivative of net revenues;
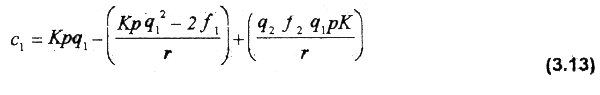
Thus, the unit cost of fishing effort is obtained:

The same reasoning can be applied to fleet 2. From equations (3.11) and (3.14), it is easy to demonstrate that fMEY is half fBE.
The dynamic behavior of the stock and fishing effort can be performed by numerically solving the model. The following example illustrates the dynamic behavior of fishery performance variables of two fleets with different fishing power.
Example 3.1. Fleets with different fishing power
Given a pelagic fishery with parameters as in Table 3.1, the dynamic behavior of population biomass, yield and fishing effort exerted by an artisanal and a mechanized fleet can be described as follows:
| Parameter/Variable | Value |
|---|---|
| Intrinsic growth rate r | 0.36 |
| Carrying capacity of the ecosystem K | 4000000 t |
| q artisanal fleet | 0.0002 |
| q industrial fleet | 0.00035 |
| Species price | 60US$/tonne |
| Unit cost of artisanal effort | 15000 US$/trip |
| Unit cost of industrial effort | 45000 US$/trip |
| Artisanal fleet dynamics parameter | 0.00001 |
| Industrial fleet dynamics parameter | 0.00001 |
Results of the simulation run for a time horizon of 50 years show an expansion of the industrial fleet until biomass reaches 2 million tonnes (Fig. 3.1a,c). Beyond this threshold, the economic rent for this fleet became negative (Fig.3.1e), leading to the exit of vessels from the fishery and a release of biomass that can be subsequently captured by the artisanal fleet (Fig.3.1d).
Figure 3.2 shows the dynamic behavior of biomass and fishing effort exerted by both fleets. A marked decrease of stock biomass is observed during the first 10 years of the fishery, as a result of a continuous increase in fishing effort, particularly by the artisanal fleet. Both variables show long-term cycles with a time lag of 5 years, determined by the delay in investment according to stock fluctuations (Fig. 3.2a). Amplitude oscillations of both variables tends to decrease until reaching a long-run bioeconomic equilibrium (TR = TC) in biomass and in the number of vessels in the industrial (7) and artisanal (1250) fleets. In the long-run, the artisanal fleet tends to exclude the industrial one, as a result of lower operative costs.
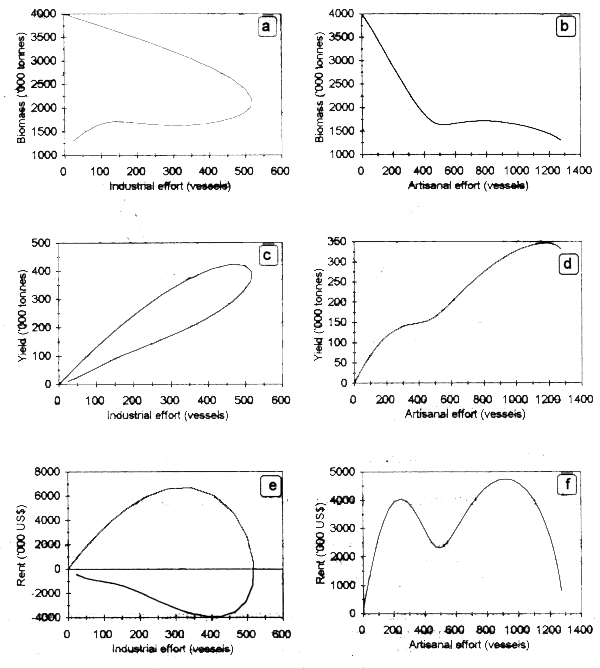
Figure 3.1. Dynamic behavior of stock biomass (a, b), yields (c, d) and economic rent (e, f) by fleet type.
This example can also be analyzed as a sequential fishery (see below), but in this case the effect of fishing fleets on population structure must be considered.
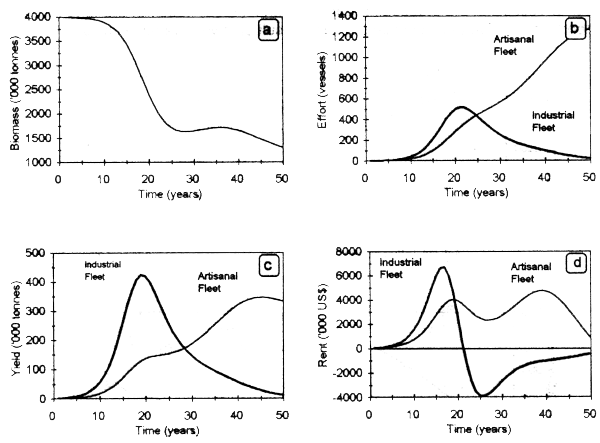
Figure 3.2. Dynamic behaviour of biomass (a), effort (b), yeild (c) and economic rent (d) obtained by the artisanal and industrial fleets.
Consider a fishery composed of 2 fleets harvesting 2 species: fleet 1 targets on species 1 and takes incidentally obtains species 2, which in turn is the target of fleet 2. The corresponding bioeconomic parameter set is given in Table 3.2.
Figure 3.3 shows the results of the bioeconomic dynamic model that represents the technological interdependence. Biomass of both species decreases as a result of fishery development (Fig 3.3a). Fleet 1, with higher fishing power, obtains important yields of both species during the first 15 years, determining a biomass reduction of species 1 to 1/4 of its carrying capacity after this period. Fleet 2 reaches maximum yields after 8 years of fishery development (Fig. 3.3b), diminishing onwards. The economic rent of fleet 1 becomes negative at t = 15 years due to a sustained increase in fishing effort, which determines diminishing biomass and yield (Figs. 3.3c, d). In the long run, both fleets reach the fBE at a number of vessels close to 500. The fishery will tend to show cycles of variable amplitude in abundance, yeild and revenues, according to the magnitude of fishing effort exerted by both fleets. In an ideal situation, effort level by fleet 1 should be expanded until the marginal revenues of both fleets equal the marginal cost of fishing effort in fleet 1. The same holds true for fleet 2. Effective management interventions in cyclic fisheries should be made, particularly, during the phase of increasing stock abundance in order to mitigate the effect of over-investment (see Caddy & Gulland, 1983 for useful discussions on the subject).
| Parameter/Variable | Value |
|---|---|
| r species 1 | 0.36 |
| r species 2 | 0.30 |
| K species 1 | 4000000 tonnes |
| K species 2 | 2000000 tonnes |
| q species 1 fleet 1 | 0.0004 |
| q species 1 fleet 2 | 0 |
| q species 2 fleet 1 | 0.00015 |
| q species 2 fleet 2 | 0.0025 |
| Price species 1 | 60 US$/tonne |
| Price species 2 | 45 US$/t |
| Unit cost fleet 1 | 60000 US$/yr |
| Unit cost fleet 2 | 15000 US$/yr |
| Fleet dynamics parameter (1) | 0.00001 |
| Fleet dynamics parameter (2) | 0.000015 |
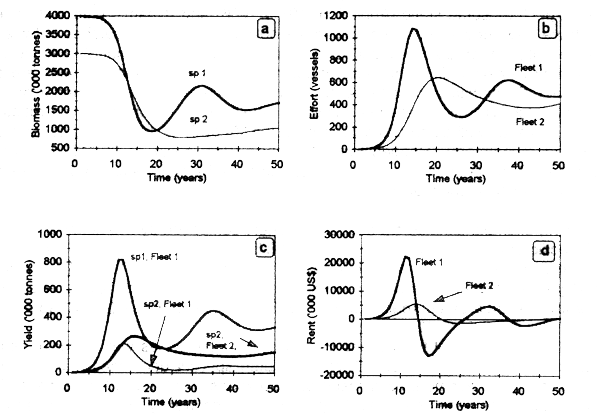
Figure 3.3. Bioeconomics of technologically interdependent fleets: (a) species biomass;(b) fishing effort; (c) yield; and (d) economic rent.
This happens when two fleets obtain incidental catches of the target species of the other fishery. Population sizes and yiels of stocks X and Y are a function of the fishing effort exerted by both fleets. However, economic revenues of one fleet will depend on the effort level exerted by the other one. For fleet 1 (target species X):
π1(t)=PxYx1(f1,f2)+PyYy1(f1f2)-c1f1 (3.16)
whereas for fleet 2 (target species Y):
π2(t)=PyYy2(f1f2)+PxYx2(f1f2)-c1f2 (3.17)
As one fleet increases its fishing effort, the equilibrium yield of the other will decrease and thus also the corresponding effort level under open access. The analysis of this case is very similar to the previous one. Because both fleets are fishing on both stocks, the operation of one will have a direct effect on the other.
In sequential fisheries, 2 fleets spatially segregated fisheries (e.g. coastal artisanal and industrial at open seas) affect different components of the life cycle of one or more species (e.g., shrimps: see Willmann & Garcia, 1985). Thus, competition between different resource users is expected. Some static (Anderson, 1977, 1986) and dynamic (see text) bioeconomic models have been developed to determine the optimal allocation of catches between 2 fleets that sequentially fish on a single stock (Geen & Nayar, 1988). Even though a single population equilibrium curve exists, different levels of catches, prices, costs and revenues are estimated for each fleet. Competitive coexistence or exclusion of fleets might occur under open access. An optimal catch allocation between fleets will depend on the corresponding ratios between prices and costs, as well as on characteristics of the life cycle of the species (longevity, growth, survival and recruitment) (Hoppensteadt & Sohn, 1981; seijo et al., 1994b). The explicit consideration of the population age structure, and the differential effect of both fleets on it, is critical for modelling sequential fisheries.
The following example considers a sequential fishery composed by two fleets and two species. A multispecific bioeconomic dynamic model is built to simulate fishing and resource behaviour through time. Input values for the model parameters are shown in Table 3.3. Selectivity and catchability parameters are estimated by the selection ogive method (sparre et al., 1989) and the Baranov's swept area approach (Caddy, 1975, 1979), respectively (see Chapter 4).
Figure 3.4 shows temporal fluctuations in species abundance, as well as yield, fishing effort and economic rent for both fleets. Biomass of both species decreases markedly during the phase of fishery development as a result of the expansion of the industrial fleet (Fig.3.4a). Catches of this fleet peak near year 20 (Fig. 3.4b), concurrently with a maximum number of vessels (Fig. 3.4c); from this year on, the fleet decreases gently until the end of the simulated time horizon (50 yr). In contrast, the artisanal fishery shows a steady increase in catches and in the number of vessels. As a result of lower operation costs, this fleet could exclude the industrial one in the long run. Increasing effort by the artisanal fleet could cause recruitment overfishing, with the consequent reduction of stock availability for the mechanized fleet. Concerning the latter, wide margins of revenues from years 15 to 20 determine an increase in the number of vessels, followed by an over-investment phase with subsequent economic losses (between years 20 and 25), being close to the BE from years 35 to 50. On the other side, the artisanal fleet presents low profits throughout the simulated period (Fig. 3.4d).
Example 3.3. Sequential fishery
| Parameter/Variable | Value |
|---|---|
| Maximum age species 1 (yr) | 20 |
| Maximum age species 2 (yr) | 12 |
| Age at first maturity species 1 (yr) | 3 |
| Age at first maturity species 2 (yr) | 2 |
| Sex ratio (both species) | 0.50 |
| Maximum observed recruitment species 1 | 50000000 |
| Maximum observed recruitment species 2 | 100000000 |
| Natural mortality rate species 1 | 0.4/yr |
| Natural mortality rate species 2 | 0.45/yr |
| Average fecundity species 1 | 1200000 |
| Average fecundity species 2 | 1500000 |
| Curvature parameter species 1 | 0.12/yr |
| Curvature parameter species 2 | 0.19/yr |
| Growth parameter t0species 1 | -0.55 yr |
| Growth parameter t0species 2 | 0.50 yr |
| Maximum length species 1 (mm) | 920 |
| Maximum length species 2 (mm) | 600 |
| Maximum weight species 1 (g) | 12500 |
| Maximum weight species 2 (g) | 2400 |
| Minimum price species 1 | 1500 US$/tonne |
| Minimum price species 2 | 1000 US$/tonne |
| Slope of price curve species 1 | 50 |
| Slope of price curve species 2 | 5 |
| Unitary cost fleet 1 | 55000 US$/vessel/yr |
| Unitary cost fleet 2 | 5000 US$/vessel/yr |
| Fleet 1 dynamic parameter | 0.00001 |
| Fleet 2 dynamic parameter | 0.0001 |
| Area swept by the gear fleet 1 (km2) | 2.50 |
| Area swept by the gear fleet 2 (km2) | 0.18 |
| 50%–75% length gear retention sp.1, fleet 1 | 450, 650 |
| 50%–75% length gear retention sp.1, fleet 2 | 300, 450 |
| 50%–75% length gear retention sp.2, fleet 1 | 300, 450 |
| 50%–75% length gear retention sp.2, fleet 2 | 200, 275 |
| Distribution area, both stocks (km2) | 5000 |
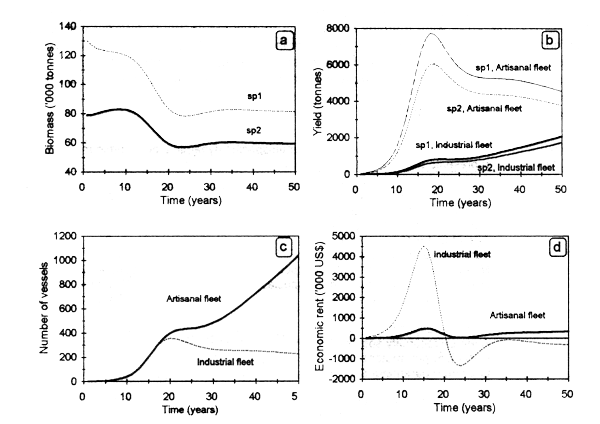
Figure 3.4. Sequential fishery. Dynamic fluctuations of: (a) biomass; (b) yield; (c) effort; (d) economic rent generated by the industrial and artisanal vessels.
It commonly occurs that 2 independent fisheries harvest 2 stocks that interact through competition or predation. In these cases, the magnitude of effort exerted in one fishery will produce an indirect effect on the performance of the other. An optimal harvesting strategy will depend on the interaction strength between species under competition, and the dietary width of the predator in predator-prey relationships. Under open-access, the bioeconomic equilibrium in one fishery will depend on the amount of fishing effort exerted in the other.
Figure 3.5 shows the dynamic trajectories of biomass of 2 competing species, as well as the corresponding yields and economic rent generated by a single fishing fleet. In the absence of harvesting, the dominant competitor has a higher biomass than the subordinate one. As fishing effort by the dominant competitor increases (suppose that it is more valuable in the market than the subordinate), its abundance decreases, thus releasing an ecological limiting resource (e.g., habitat, food). This in turn allows the biomass and yield curves to shift up for the subordinate species, until reaching an effort level above which catch also tends to decrease. The open-access bioeconomic equilibrium is reached at high effort levels that determine a low abundance of the ecologically dominant species. In this context, if the price of the subordinate species is high enough, the dominant one can be effectively driven to extinction if it is more vulnerable than the subordinate species. The economic rent is maximized at an effort level of 292 vessels.
It is worth emphasizing that the exact position of the BE in a fishery composed by two competing stocks depends upon the biological (e.g., growth rates) and economic (e.g., market prices) parameters, as well as on the interaction strength between species.
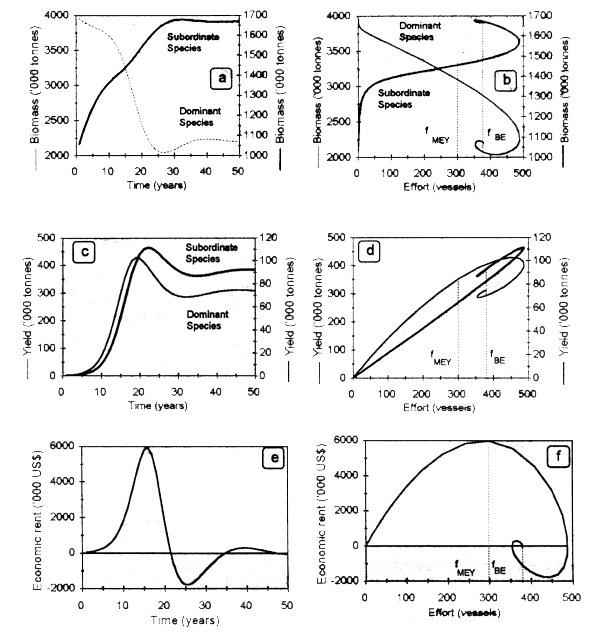
Figure 3.5. Dynamic trajectories of biomass (a, b), catch (c, d), and economic rent (e, f) generated by a fishery composed by two competing species.
In this case, an increase in fishing effort directed to the prey will decrease its abundance and thus the magnitude of resource available for the predator (assume for simplicity that the predator is specialist in its trophic behavior). Thus, increasing effort over the prey will also decrease the yield and revenues generated by predator harvesting. The reverse trend will occur for the prey: its corresponding levels of bioeconomic equilibrium and maximum yield will depend on the magnitude of fishing effort exerted over the predator. Predator behavior is critical in this context. A generalist predator feeds on several prey items and thus is able to switch according to the concentration profiles and abundance of different preys. Thus, the dynamic trajectories shown in Fig. 3.6 should be modified to account for the switching behavior of the predator.
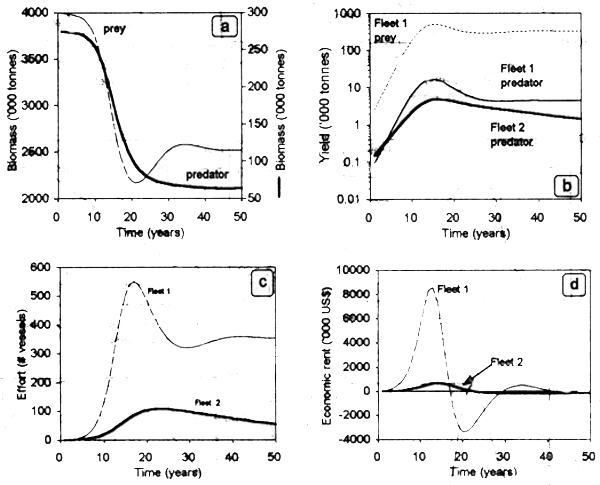
Figure 3.6. Predator-prey relationship in ecologically interdependent fisheries. Dynamic trajectories of (a) prey and predator biomass, (b) effort, (c) yield, and (d) economic rent generated by two fleets.
A techno-ecological interdependence occurs when two technologically interdependent fleets harvest two interacting stocks. The multiple nature of the interaction makes difficult to analyze the dynamics of this kind of fisheries.
The Lotka-Volterra (Volterra, 1926; Lotka, 1932) and Gause & Witt (1935) models are generally used to model competition between species. In the latter, the differential equations used are:
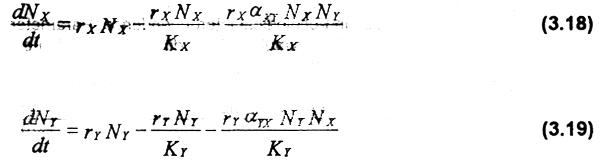
where NX and NY are the abundance of species Χ (dominant) and Υ (subordinate) respectively; rx and ry are the intrinsic rates of population growth; KX and KY are the respective carrying capacities of the environment when both species growth in isolation, αXYis a constant that measures the per capita competitive effect on species X of species Y, and αYX is a constant that measures the effect on species Y of X. Thus, the population increment of one species is limited by intra and interspecific effects. Here again, the dynamic behavior of one fishery depends upon the level of effort in the other, and also on the interaction strength between competitors.
The predator-prey model used to model this ecological interdependence is based on the logistic equations described by Leslie & Gower (1960). Let Χ be the prey and Y be the predator:
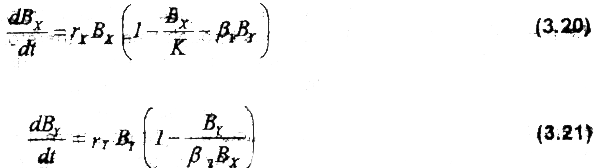
In this set of equations, prey growth is limited by the carrying capacity of the system, K, and also by the predation term β1ΒY. Growth of the predator (species Υ) population is a function of its intrinsic growth rate and prey number. Constant β2 relates the number of preys with the maximum biomass that the predator is able to reach, constrained by K.
The following example is based on a computer model (Seijo et al., 1996) that simultaneously considers a technological and an ecological (predation) interdependence (Table 3.4). Dynamic fluctuations in prey and predator biomass shown here are not the classic ones found in the ecological literature (Begon et al., 1990), because the fishing fleets are technologically interrelated. Both stocks decrease with the increasing fishing effort applied during the first 15 years of fishery development (Fig 3.7a). Fishing effort exerted by fleet 1 on prey also impacts predator biomass during this initial period, declining afterwards (Fig. 3.7b). The marked increase of fleet 1 is a response to the high yields achieved both on prey (target) and predator (incidental) (Fig. 3.7c). As a result, the economic rent for fleet 1 during the former 15 years is much higher than for fleet 2, which targets exclusively on the predator and always works close to the bioeconomic equilibrium (Fig. 3.7d). Biological and economic parameters are crucial to define the dynamic trajectories of the interdependencies. In this case for example, depleting a prey can reduce the abundance of predators, but not necessarily vice versa.
| Parameter/Variable | Value |
|---|---|
| r prey | 0.36 |
| r predator | 0.15 |
| K prey | 4000000 tonnes |
| K predator | 275000 tonnes |
| q prey: fleet 1 | 0.0004 |
| q prey: fleet 2 | 0.0 |
| q predator fleet 1 | 0.0002 |
| q predator fleet 2 | 0.0004 |
| Unit price of prey | 60 US$/tonne |
| Unit price of predator | 275 US$/tonne |
| Unit cost fleet 1 | 60000 US$ |
| Unit cost fleet 2 | 9000 US$ |
| Fleet 1 dynamic parameter | 0.00001 |
| Fleet 2 dynamic parameter | 0.000015 |
| β1 (predation constant) | 0.00000001 |
| β2 (constant) | 0.069 |
Knowledge of the magnitude of the ecological and technological interdependencies facilitates the prediction of biomass fluctuations through time and therfore the net revenues derived from fishing. The response of a community to fishing pressure depends on its complexity (sensu Cohen, 1987), that is, the interaction strength between species and the functional role of the exploited species (Sainsbury, 1988; Lubchenco et al., 1995; Botsford et al., 1997). For example, changes in community structure might be different according to the trophic spectrum (specialist or generalist) and efficiency of an exploited predator, and the magnitude of interactions between preys (if they compete or not and if the competition is symmetrical or not). Moreover, harvesting a dominant competitor will tend to increase the divrsity of a community, while the exploitation of a keystone species could lead to a simplification of the community and decrease in biodiversity (Paine, 1966; Durán & Castilla, 1989). According to exploitation levels, the magnitude of the interdependencies could be modified and the corresponding management schemes altered.
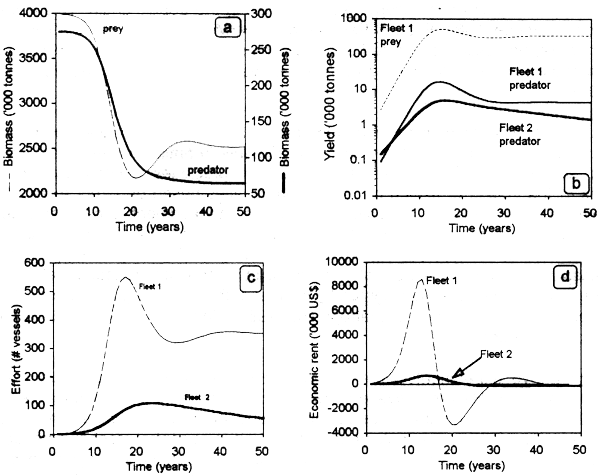
Figure 3.7. Techno-ecological interaction: stocks with a predator-prey relationship captured by 2 technologically interdependent fleets.
At least for sedentary or sessile shellfishes, improved understanding of the effects of fishing on ecological communities comes from experimental manipulation of populations and fishing effort in the field (Jamieson & Caddy, 1986; Cobb & Caddy, 1989; Defeo, 1993a, 1996; 1998; Castilla, 1994). Experimental management constitutes a useful way to determine patterns of shellfish response in relation to magnitude of fishing pressure and of linkages and strengths of ecological interactions (Defeo & de Alava, 1995). These species are particularly amenable for experimentation: the low or null mobility allows performig localized experiments with different levels of stock abundance and fishing intensity. Moreover, easy design and manipulation features of experimental plots facilitate the evaluation of population responses that might accompany dissimilar fishing levels. The low exchange of organisms among experimental replicates is an additional useful feature.
In the above context, recent experimental studies have demonstrated that the structure of harvested communities of rocky shores drastically differs from pristine ones, and that varying harvest intensity on shellfishes of different trophic levels could produce different community structures. Intertidal benthic communities protected or with minor human interference have been compared with those where man has exerted a persistent influence in the system (Castilla & Durán, 1985; Hockey & Bosman, 1986; Bosman et al., 1987; Bustamante & Castilla, 1990). Results indicated that the effect of man was not only evident in targeted species, but also indirectly in unharvested ones occupying lower levels in the trophic chain. This is known as a cascade effect (Paine, 1984; Estes & Palmisano, 1974; Moreno et al., 1984; 1986), and mainly occurs when: (a) harvesting is directed to a keystone species (sensu Paine, 1980), usually top predators; and (b) trophic links between species are very strong (Duran et al., 1987; Lubchenco et al., 1995).
Even though experimental manipulation in the field is a top priority for invertebrates, it is highly difficult and complex to perform large-scale experiments in finfish communities. Large spatial scales of events such as dispersal and migration, precludes experimentation and even predictions of the impact of fishing on finfish communities based on direct extrapolation from small-scale field experiments. Moreover, large-scale management experiments tend to be expensive and risky in comparison to some management policies favored by risk-averse policy-makers or environmental interest groups (see Walters, 1997 for a detailed discussion). Nevertheless, the need for an empirical and manipulative approach to test the effects of fishing on fish communities has been highlited by several authors (see e.g. Sainsbury, 1987, 1988; Sugihara et al., 1984; Garcia & Demetropoulos, 1986; Mapstone et al., 1996; Walters, 1997 and references therein).
Most of the above experiments were carried out in the context of adaptive management (Walters, 1984; 1986; Hilborn & Walters, 1987), that is, to manage through experimentation, changing deliberately fishing effort levels to understand the dynamics of the community and the fishery. This strategy allows the implementation of short-term control mechanisms, according to a continuous flow of information about the stock and the fishery. Management policies are improved, and uncertainties about the dynamics and structure of the finfish community are reduced, as additional information becomes available (Sainsbury, 1987, 1988). This approach is particularly useful in cases when the overall increase in fishing activity has not been accompanied by a corresponding increase in scientific and fishery data, and the relative absence of demographic and fisheries studies may lead to inadequate fishery management advice.
The impact of fishing in the structure of an ecological community might depend also on the number of species that are marketable, and on the corresponding unit prices. For example, the net revenues derived from fishing could be maximized by deliberately overexploiting one species in order to produce the increase of a subordinate competing species with higher market value. Existing market conditions could also lead to depletion of the most commercially profitable prey species to such a low population size that the fishery for the other interdependent (predator) stock could be economically unfeasible. Recent investigations do not integrate humans as highly specialized predators but rather they consider their generalist behavior, as closed seasons/areas and other regulatory measures are being imposed to the traditionally exploited resources (Smith & McKelvey, 1986).
In summary, the fishery, the ecological interactions, the physical environment and the dynamic development of the fishing sector have differential effects on fish stocks. Thus, basic biological and experimental studies, conducted at different spatial and temporal scales, could provide the only valid alternative to predict the bioeconomic effects derived from ecological and technological interdependencies.
![]()
![]()
![]()