![]()
![]()
![]()
Genetic improvement programmes for livestock in the developed countries have advanced to the present stage, chiefly because the infrastructural elements necessary for planned breeding programmes such as intensive performance recording and AI are available. In addition advances have also been made in population genetics and statistics which have made it possible to develop genetic improvement schemes applicable on a national scale.
In contrast, infrastructural facilities for national breeding programmes are lacking or are inadequate in most parts of the tropics. The need for such facilities has also not arisen because the traditional methods of livestock husbandry practised in these areas, e.g. nomadism or backyard farming, make it impossible to operate a planned breeding scheme as is done in the developed countries. As an alternative, state-sponsored livestock farms have been set up with a view to effecting genetic improvement in these herds or flocks and passing the improved stock, chiefly males, to farmers.
There are many arguments in favour of selection programmes for indigenous breeds of livestock.
They are adapted to the tropical environment. Among the more important adaptational qualities are disease and parasite resistance, e.g. to trypanosomes and ticks, the ability to survive and reproduce in hot, humid, environments and the ability to exist on low quality feed and limited water supply. Selection for increased productivity should therefore raise their potential for productive traits without serious detriment to their adaptational qualities.
Germ plasm of exotic breeds is imported into many tropical countries either as live animals or as frozen semen. Crossbreeding of these exotic breeds with indigenous stock poses a danger of extinction of the latter types. In these situations, one can ensure the survival of the local strains only by raising their genetic merit so as to minimize the competition (Cunningham, 1979).
Selection among indigenous breeds is an integral part of some crossbreeding programmes such as rotational crossbreeding where exotic and indigenous breeds are used in rotation.
In formulating selection programmes in institutional herds in the tropics, it is important to ensure that they are simple in design and that the traits selected are those of real economic value. Among these, adaptability traits should always be emphasized.
It is also important that the population under selection should be exposed to environmental conditions similar to those in which the offspring are expected to perform. For example, if the objective of selection in an experimental herd of beef cattle is to supply genetically superior bulls to farmers who keep their animals on range without supplementary feeding, it would be inappropriate to keep the experimental animals on high levels of concentrates. This is important to avoid errors in selection caused by genotype-environment interaction. The same argument also applies to experimental herds that observe stringent parasite control measures such as frequent treatment with acaricides or anthelmintics. Such considerations should, however, not lead to the other extreme of imposing excessively harsh conditions that would mask the expression of genetic variation.
Improvement programmes in many institutional farms in the tropics are at present inefficient, as they are confined to small or medium-sized government herds and are often hampered by a lack of continuity in the pursuit of selection objectives. Their effectiveness should be increased by organizing integrated programmes that embrace all government farms of the same breed of livestock and also private herds above a certain level of management. Establishing links with private farmers is important not only to extend the genetic improvement to their herds but also to increase the effective size of the population under selection and to ensure that testing of animals is also done under field conditions.
The two major selection procedures used in ruminant livestock are performance testing and progeny testing. The former is used for characters of high heritability and easily measured in both sexes, e.g. growth rate. For characters that have a low heritability or are measureable in only one sex (e.g. milk yield) or after slaughter (e.g. carcass quality), progeny tests are employed. Progeny testing, when based on adequate information, gives an accurate estimate of breeding value and is therefore preferred when an animal is to be used widely for breeding as in AI. Information on other relatives is often used as a selection aid, particularly in choosing young males for progeny testing, e.g. dairy cattle. For this purpose, the information that is most valuable is that of the sire and dam. The additional gain in accuracy by using information from ancestors further back in the pedigree is relatively small.
The above principles of genetic improvement form the basis of different types of selection schemes. These may operate within individual herds, among a group of cooperating farmers, or nationally. Some of the important ones are described below.
Testing stations: Animals from different herds are assembled in testing stations and measured under a uniform environment. The influence of the herd environment is therefore minimized and the population under comparison is vastly increased. It is particularly suitable for situations where a large number of small herds are involved each of which does not have the capacity to carry out proper withinherd comparison. Testing stations can be used either for performance testing, e.g. growth rate, or for progeny testing, e.g. milk yield.
Progeny testing schemes: The progeny test depends for its accuracy on having a large progeny group per sire. Where a number of herds are involved in testing, the maximum efficiency is achieved when each sire has equal number of progeny in each of the herds. The scheme usually operates by having a central station from where semen of the different sires on test is distributed to a network of AI centres which carry out the inseminations. A modification of this scheme called the reference sire scheme enables comparisons to be made also among bulls that are used only within individual herds. The reference sires, which are usually progeny tested animals and available only by AI, are used alongside the individual herd's own bulls in all herds. The breeding values of home-bred bulls are then estimated in relation to the reference sires. Comparisons among bulls in different herds are made through the linkages established by the reference sires. Thus sire replacements can be made from among all herds.
Nucleus or group breeding scheme: This scheme is based on the principle that in each herd there is a small number of genetically very superior animals which if brought together will form a nucleus whose average genetic merit is far greater than that in any of the contributing herds (Nicoll, 1976). The important element in this scheme is therefore for a group of farmers to agree to pool their highperforming animals. Once the nucleus herd is assembled, an efficient system of recording and selection is implemented. The best males are kept for breeding in the nucleus while other selected males are given to the base herds for breeding. By this means, improvements are quickly spread throughout the group. The nucleus may remain open to animals from the base herds, the best females from the latter being admitted periodically and compared with those in the nucleus. Usually, only females are transferred from the base to the nucleus since sire selection will not be practicable in base herds due to managerial reasons. The main advantage in the nucleus scheme is that the genetic superiority of sire replacements coming into the base herds from the nucleus is far greater than what is achievable in each of the base herds. It is particularly attractive in situations where within-herd selection programmes are ineffective due to small population size or inadequate technical skill.
Since the nucleus breeding scheme shifts the onus of operating the breeding programme from the farmer to the nucleus herd, it seems an attractive method for the tropics because of the limitations discussed earlier. However, the organization of the scheme may have to be under government control because cooperative ventures among farmers may not always be practicable.
No serious attempt has hitherto been made with national selection programmes to improve the dairy merit of indigenous cattle. This is partly due to the belief that the limiting factor to increased production is not the genetic merit of the animals but rather the adverse environment, and that improvements in husbandry and veterinary care should precede the setting up of breeding schemes. But when one considers the performance of most indigenous breeds, a remarkable similarity is noticed in their late age at maturity, short lactation, long dry period, low average daily milk yield and lack of dairy temperament. Most of these characters are expressed even under conditions of improved husbandry, thus demonstrating that poor productivity is attributable to low genetic merit. Breeding schemes to raise their genetic potential are therefore fully justified.
The maximum rates of genetic gain in milk yield achievable by selection even in temperate breeds is of the order of 2.0 percent per annum. This low rate of improvement has discouraged implementation of selection schemes for indigenous breeds, since the overall increase in production attainable by selection in cows yielding 500–1 000 kg is not of any great magnitude. It should, however, be recalled that even European breeds were as unproductive as the tropical breeds before the application of selection programmes and it is the application of planned selection programmes that has brought the advance that we see today (Cunningham, 1979). Since the techniques for genetic improvement are even more developed today than when they were first used on European cattle, it is not inconceivable that similar achievements could be made in tropical dairy breeds.
Where positive selection efforts have been made to improve tropical breeds, the results have been encouraging. Annual genetic gains of approximately 2.5 percent of the herd mean yield have been reported by Acharya and Lush (1968) in Haryana (zebu) cattle in India and by Franklin et al. (1976) in the Australian Milking Zebu (Jersey x zebu cross). The high rate of genetic gain in both populations was partly attributed to the high coefficient of variation for milk yield in the populations, which is characteristic of most dairy cattle in the tropics (Mahadevan, 1966). It is also clear that, in populations that failed to show significant gains, it was the inefficient selection procedures that were responsible, e.g. East African zebu (Kimenye, 1979).
In formulating a breeding programme for dairy cattle in the tropics, the ultimate objectives of the programme and the environment and production system under which these objectives are to be realized should be defined. Thus, the objectives should clarify whether it is milk alone or milk and beef or milk and draught or all three that are to be improved. Further, depending on market demands, the importance to be attached to compositional quality should also be clarified. Similarly, the production system, which can range from nomadism through small sedentary herds to large commercial farms, will determine the importance given to adaptation traits, particularly those concerned with disease resistance and reproduction.
Milk yield is the most important trait considered in selection programmes for dairy cattle, though fat percentage is also measured routinely in several countries. Since the negative genetic correlation between milk yield and fat percentage may result in the latter declining to low levels, it is usually monitored to eliminate undesirable animals. Bull fertility is important, particularly in AI bulls, while in the female reproductive performance is considered in culling, this being done more for economic reasons than for genetic improvement.
The principal selection method employed is progeny testing where the breeding value of bulls is assessed on the performance of their daughters. The test depends for its efficiency on a large progeny group per sire and on having each sire represented in several herds. - AI is necessary to separate environmental from genetic effects. Milk yield records are usually restricted to those from first-calving heifers since this gives the largest group of unselected daughters, although some countries, e.g. USA, consider records of older animals as well. Young bulls that enter the progeny test are bred from the best progeny-tested sires and selected dams. Each young bull is used on about 500 cows in milk-recorded herds to ensure that first lactation records of at least 80 daughters are obtained. Matings are done at random to ensure that the dam contribution to progeny genotype is similar for all sire progeny groups.
Though this type of selection programme appears to require a great deal of infrastructural facilities, an efficient testing programme could operate even with a simpler organization if recording is limited to that required for progeny testing. Thus, Ireland operates a scheme which tests 40 bulls annually on approximately 75 daughters per bull. Only 3 000 daughter records are necessary, though the scheme serves a population of approximately one million cows.
Institutional herds have an important role in the tropics since these may be the only herds where genetic improvement of indigenous stock can be carried out. Institutional facilities are also necessary in selection programmes for exotic breeds which are required in pure or crossbreeding programmes.
Artificial insemination should be employed for breeding even in institutional herds so that large sire progeny groups are obtained for accurate progeny tests. Where the institutional herd is too small, it is necessary to enlist the support of the progressive farmers in the area to participate in the progeny testing scheme. In situations where only natural mating can be employed, progeny testing loses much of its efficiency since the number of bulls required is so large that the selection intensity as well as the accuracy of the test are much reduced. Selection of bulls on their dams' records will then be equally efficient provided that bulls are replaced annually to minimize the generation interval (Buvanendran, 1981).
An example of a successful breeding programme involving an institutional herd is that used in the Sahiwal herd in Naivasha, Kenya. The breeding plan given below is based on the description by Meyn and Wilkins (1974) whose schematic diagram is also reproduced in Figure 4.1.
The herd consists of about 500 cows. Breeding is by AI with semen from the best two progeny tested bulls. Female selection is almost entirely on milk yield during the first two lactations. About 50 percent of the heifers that enter the milking herd annually are culled at the end of the first lactation and a further 50 percent after the second lactation. Thus only 25 percent of the heifers freshening annually are kept to the third and higher parities. These animals constitute the elite herd consisting of about 180 animals. They are bred only by progeny tested bulls while the balance of the herd (320 animals) consisting of heifers and first calvers is used for progeny testing young bulls.
Figure 4.1 Breeding plan at the National Sahiwal Stud, Naivasha, Kenya.
(Source: Meyn and Wilkins, 1974)

Bulls are selected from among those born in the elite herd. They are first performance tested for growth rate on pasture to 2 years of age and 15 bulls (out of the 70 born annually) are chosen on an index that combines growth rate with dam's milk yield. A further five animals are removed for poor physical conformation and semen quality and the remaining 10 are progeny tested. They are bred to females outside the elite group (about 320 animals) and each bull is evaluated on the first lactation yields of about 14 daughters. The best two bulls are chosen for use in the elite herd and national AI scheme. The genetic progress with this type of programme is estimated at 3–4 percent per year.
In programmes involving crossing of exotic and indigenous stocks for developing a new breed, selection has an important role to play after the initial crossings are made to establish the desired gene pool. Selection can be not only for the improvement of production traits but for adaptability traits as well.
Selection for milk yield: The procedures used in selection for milk yield involve progeny testing and are similar to those outlined in Section 4.2.3. Testing of bulls is, however, not possible in the early generations of new breed formation (F1 or backcross depending on the composition of the new breed (see Section 6.1.1) since young bulls are required to mate with heifers of thier own age and generation. Further, even from a genetic viewpoint, selection should commence only after segregation and recombination of the gene combinations of the parental breeds have occurred. The opportunity for this to happen will arise after inter se mating starts. Progeny testing of bulls should therefore commence with bulls born after this stage.
Private farmer participation is important in sire evaluation for reasons given earlier but complications may arise if the dams mated to the sires differ in their genetic make-up. This is usually the case in situations where crossbreeding programmes in private herds have been under way for a number of years before selection schemes commence. The crossbred dams may then differ in respect of both breed and grade of exotic blood and bias the progeny test due to their different contributions to the progeny. This was the case in Brazil when progeny testing of crossbred bulls was initiated in 1977 (Madalena, 1981a). The approach adopted to minimize the bias in this programme was to use all bulls on all farms to avoid confounding of dam breed type and bull. Further, within farm and year, dams were balanced among bulls according to percent European blood and progeny lactations were expressed as deviations from herd-year-season means corrected for age and grade of European blood. This method was suitable, because herd sizes were large (40–700 cows). It is clearly unsuitable where herd sizes are small. In such situations, the concept of the herd as a unit may have to be abandoned and comparisons done within groups of herds, grouped on the basis of average grade and production level.
Adaptability traits: Selection for adaptability traits should wherever possible proceed simultaneously with improvement for milk yield. Measurements for these traits may, however, have to be limited to experimental herds because of the facilities required. This was done in the development of the Australian Milking Zebu (AMZ) - a breed derived from the Indian zebu and Jersey. While progeny tests of bulls were done in both experimental and outside herds, selection for adaptability was confined to the experimental herd at Badgery's Creek in New South Wales (Hayman, 1972). Young bulls were first screened for heat tolerance and tick resistance and only 1 in every 6 or 8 bulls screened was finally progeny tested.
The heat tolerance test was designed to select bulls that maintained their body temperature under heat stress while simultaneously having a high energy conversion rate (Allen and Donegan, 1974). Animals were tested in climatic chambers and measurements recorded of physiological changes (rectal temperature, sweating rate and respiration rate) and feed and water intake when they were moved from low to high temperatures (from dry and wet bulb of 21° and 18°C to 38° and 24°C respectively). Body temperature and respiration rate were given a negative score and sweating rate and feed conversion given a positive score. The best third of the animals chosen on this test were put through tick challenges. This was done by artificially infesting each animal with 40 000 larvae of Boophilus microplus and counting the number surviving to adult stage from the 18th to the 22nd day after infection. The best 50 percent of the animals that had the least number of engorged ticks were finally chosen for progeny testing after examination of semen quality.
In the absence of AI and milk recording programmes, institutional herds offer the only opportunity to carry out recording and selection. Cunningham (1979) has proposed a kind of open nucleus breeding scheme (Figure 4.2) that may be suitable for this situation. The plan operates around a central herd which should preferably be under government control and carry out proper recording and breeding practices. The supporting base population is the village herds which provide cows to replace about 10 percent of the cows in the central herd annually. Selection from village herds will be by simple procedures involving judgement by eye, milking ability, size, conformation and condition. Where feasible, test milkings may also be done. The selection of these animals will be done by officers who would tour many villages to select the best animals from as wide a population as possible.
Figure 4.2 Open nucleus breeding scheme (Cunningham, 1979)

Bulls are bred from the best cows and selected bulls in the central herd. They are then evaluated on their own growth rates and dams' and milk yields and the best are chosen as sires. The lowest yielding 10 percent of the cows are replaced with new animals drafted from village herds. The latter animals are recorded during the following year with the remainder of the central herd and re-evaluated.
The benefits to the cooperating farmers from this scheme will accrue from the sale of selected bulls from the central herd. Assuming that the central herd has 200 cows and that the best third of the bulls born annually are chosen, about 20–25 bulls will be available for sale to farmers after meeting the central herd's requirements. This may satisfy the requirements of the cooperating farmers but will have hardly any impact among farmers outside the scheme. This is a serious shortcoming of programmes involving central herds where supporting AI services are inadequate.
Selection of beef animals was practised in developed countries even before the genetic mechanisms of growth were known. In the early days, selection was based on visual appraisal (type and conformation score) for desirable growth and carcass characteristics and was directed towards stocky, early maturing animals. This resulted in an increase in the ability of the animals to fatten at younger ages but there was no increase in growth rate or efficiency. In the last few decades, selection has been intensified for growth characteristics, chiefly on the basis of performance tests.
The breeding programmes in developed countries have as their chief objective, the improvement of growth rate, feed conversion and carcass quality. The heritabilities of these characters are relatively high and therefore performance testing is used, sometimes combined with sib testing for characters that can be measured only after slaughter.
The limitation of faster growth rate as a selection objective is that it results in a nearly proportional increase in mature cow size and hence in the cost of maintenance. Therefore, there is little net reduction in total cost per unit of beef marketed from faster growing animals. This is particularly true in suckler beef production, since two-thirds of the total feed used in the system is consumed during the breeding phase and only one-third in the final growing phase. Selection for growth rate, however, is important for terminal sire breeds used for crossing, where the antagonism due to the higher dam size does not arise.
Similar arguments have also been put forward to show that larger breeds of beef cattle are not intrinsically more efficient than small breeds. This fact is relevant to the situation in some tropical countries where cattle breeds differ greatly in size (such as zebu and trypanotolerant breeds in West Africa) and the smaller breeds are often considered inefficient. Such comparisons based on size alone can be very misleading since efficiency of production depends on many other factors. FAO/ILCA/UNEP (1980) have proposed breed comparisons on the basis of a cow productivity index (Table 4.1) that combines reproductive rate, cow and calf viability, milk yield, calf growth and cow weights. The index is an estimate of the total weight of a one-year-old calf plus the liveweight equivelant of milk produced per unit weight of cow per year. The merit of such an index lies in relating all the important production characters to the weight of breeding cows which determines maintenance costs.
Table 4.1
CALCULATION OF PRODUCTIVITY INDICES
| Parameter | Code | Calculation |
| Cow mortality during year (%) | A | |
| Calving percentage (%) | B | |
| Calf mortality to 1 year (%) | C | |
| Percent of calves reaching 1 year (%) | D | B (100-C)÷100 |
| Calf weight at 1 year (kg) | E | |
| Annual milked-out lactation yield (kg) | F | |
| Percent of cows completing a lactation (%) | G | 100 - (C÷2)1 |
| Total liveweight equivalent of milked-out yield (kg) | H | F (G÷100)÷9 |
| Total weight of 1-year-old calf produced per cow (kg) | I | E (D÷100) |
| Weight of 1-year-old calf plus liveweight equivalent of milk produced per cow maintained (kg) | J | (I+H)÷(100÷A÷2) ÷ 1002 |
| Average cow body weight (kg) | K | |
| Weight of 1-year-old calf plus liveweight equivalent of milk produced per 100 kg of cow maintained annually (kg) | J×100÷K |
1 A cow whose calf dies during a lactation period is considered on average to have produced milk during half the period.
2 Cows dying during the year are considered on average to have been maintained for half a year.
When large zebu breeds and the small trypanotolerant breeds were compared by this index, virtually no difference was found in terms of productivity per 100 kg cow maintained per year, or in feedlot performance estimated as weight gain per 100 kg body weight maintained. Small breeds are therefore equally efficient for beef production as the larger breeds, but the optimum weight of slaughter for maximum efficiency will be lower for smaller breeds. Smaller breeds may also be at an advantage under conditions of extreme fluctuations in feed supply. They seem to have the ability to lay down fat on a lower plane of nutrition which probably explains their ability to remain in a better condition than larger breeds during the dry season when feed is scarce.
In spite of the shortcomings of growth rate as a selection criterion, it continues to be used as the major selection objective in many developed countries. Some of the important breeding schemes in these countries are discussed and their relevance to the tropical situation examined later.
Central performance tests: Male calves with above average weaning weights in their herds are usually brought in groups at 150–190 days of age for performance testing. At the test station, they remain for a period of 7 months till they are 400 days old during which period they are given a test ration and various measurements (see Section 2.3.2) are recorded. Even though herd influences are supposed to be eliminated in such central tests, the carry-over effects of pre-test management can bias test results due to compensatory growth. Such influences can be minimized by (i) allowing animals that enter the test at 5–6 months to undergo an adjustment period of 3–10 weeks on the test ration before beginning the official test which should last 5–6 months or (ii) starting the performance test at a younger age of about 90–120 days and terminating at about 400 days of age. The type of feed and the method of feeding vary between test stations. In general, roughage is offered ad libitum, and the amount of concentrates varied according to the weight of the animal so as to keep the roughage: concentrate ratio constant irrespective of growth rate. The test period should be long enough to bring out the genetic variation between animals. This depends on the type of ration. Thus, while 5–6 months is adequate for a high concentrate ration, it should be longer if the ration is high in roughage. For animals that are only on pasture, a period of 12 months is advisable.
Selection within herds: This scheme operates by identifying cows with superior mothering ability and calves with superior growth rates. Calves are weighed at ages close to 100, 200, 400 and 500 days of age. The 100- and 200-day weights are used as indicators of mothering ability while the 400- and 500-day weights are used in selection of bull and heifer calves.
Nucleus breeding scheme: The principles of this scheme have been discussed earlier.
Progeny testing: Progeny testing is used only to a limited extent in beef cattle because, for improvement of growth rate, the performance test is cheaper, quicker and can be applied to many bulls. However, progeny testing of beef bulls may be carried out when they are to be used in AI, for traits such as ease of calving and carcass quality. Bulls to be progeny tested are first selected on the basis of a performance test and then mated to cows in commercial herds. Information is collected on dystocia, calf mortality, liveweight gain, carcass weight and carcass quality. In such tests, where the performance test precedes the progeny test, over 80–90 percent of the genetic gain in growth rate is attributable to the performance test (Allen and Kilkenney, 1980).
Reference sires: Most beef herds use natural service for management reasons. The reference sire scheme may therefore be more relevant in beef than dairy cattle, since only the small number of cows required for insemination need to be observed for oestrus.
The environment to which beef cattle are exposed differs in most cases from that of dairy animals. The latter are often shielded from environmental stress by provision of shade, improved pasture, supplementary feed and veterinary care. Beef cattle, on the other hand, are usually kept on land of low potential. Their main function is to convert pastoral vegetation into meat while being exposed to the total impact of heat, parasites, infectious diseases, periodic malnutrition and shortage of water. The most important trait required is therefore the ability to survive and reproduce in this environment. Thus, the indigenous breeds of beef cattle which possess some of these characteristics will have to be relied on to make the major contribution to beef production. As stressed earlier (Section 4.3.2), size is not the only criterion influencing efficiency of beef production. Local breeds should be evaluated on their overall productivity before any hasty decisions are made to substitute them by imported temperate or large zebu breeds.
Among the important adaptability traits of tropical breeds are disease resistance qualities. The ability of zebu breeds to resist tick infestations and the tolerance shown by certain African breeds such as the N'Dama to trypanosomiasis are well documented. The relationship between these traits and other performance traits is, however, not known. For example, all trypanotolerant breeds are small compared to their susceptible counterparts which suggests that small size is an integral part of tolerance. If an antagonism exists between large size and trypanotolerance, selection of these breeds for growth rate may result in increased susceptibility to trypanosomiasis.
The production system used in most parts of the tropics is one of pure breeding (suckler beef). In this system, most of the female offspring enter the herd as cow replacements and only surplus males are used for fattening. The calves usually suckle their dams for a long period (6–12 months) and are later fattened on low-quality pasture. The sole output from the cow is thus the weaned calf. The important traits in this system are, therefore, reproduction, milking and mothering ability of the cow and growth potential of the calf. These traits are considered below:
Fertility: The reproductive rate in most tropical breeds of cattle is low. Yet, this trait is very important not only in the economics of beef production but also in genetic improvement because of its influence on selection intensity. Though the heritability of the various measures of reproduction is low, rigid culling to eliminate animals with low fertility has improved fertility in zebu cattle (Plasse, 1974). However, as most of the improvement resulting from such selection may be non-genetic, fertility may decline in subsequent generations unless the practice of culling on reproductive performance is sustained.
Mothering ability: As important as fertility in tropical rangelands is mothering ability which determines survival rate and preweaning growth rate of the calf. Increased milk yield may increase weaning weight but it may be detrimental to the overall economics of beef productivity if the subsequent conception of the cow is delayed. To avoid such antagonisms, a cow index that combines reproductive rate, calf survival and weaning weight may be preferred for selection purposes. The heritability of this index, however, has not been estimated.
Growth rate: Selection for growth-rate is usually done on the basis of the final weight at a constant age such as 18 or 24 months. This is preferable to selection on the basis of gain from weaning to a constant age because of inaccuracies that may arise from compensatory growth as discussed earlier (Section 4.3.3). The problem of concurrent increase in mature size consequent to selection for growth rate is equally applicable to tropical cattle. One of the approaches that has been proposed for temperate breeds to minimize the increase in mature size is selection on the basis of relative growth rate (RGR). It is measured as the daily gain relative to the average size over the time interval tested
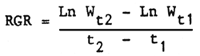
Where Ln denotes the natural logarithm of weight W at time 1 (t1) or time 2 (t2). The effectiveness of this parameter in changing the growth pattern has still to be determined.
Longevity: The length of productive life is important since the relative cost of raising a calf decreases with the increase in its productive life. However, as selection for longevity increases generation interval, males may be selected at younger ages on their own structural soundness and the longevity of close relatives.
A clear definition of the environment in which offspring of the selected populations are expected to perform is necessary. This is important because of the wide variation in nutritional level and environment in beef cattle production systems which can range from grazing on natural grasslands to feedlot fattening. Recent Australian work (Frisch and Vercoe, 1978) has shown the importance of genotype-environment interactions in growth rate. On a low plane of nutrition, the response to selection is chiefly through a reduction in maintenance requirement which results in low appetite and reduction in inherent growth potential. Conversely, on a high plane of nutrition, selection increases growth rate principally through increased appetite. Thus, animals selected under either of the extreme conditions of nutritional deprivation or high concentrate feeding will be unsuitable if conditions are reversed. A practical approach when the environment cannot be defined, would be to select animals on a moderate plane of nutrition. This may be grazing on natural or improved pasture during the rainy season and minimum supplementation with conserved forages during the dry season.
Genetic improvement programmes similar to those used in developed countries are suitable in the tropics if the herds are large and facilities for performance recording are available. However, in most cases, either herd sizes are too small or where they are large, recording may be impossible, e.g. nomadic herds. Some alternative schemes are discussed below.
Large organized ranches or institutional herds
Many tropical countries have government-owned herds with the objective of producing superior stock for distribution. Some of the important ingredients for the success of a breeding programme in such herds are:
A reasonable population size to permit adequate selection intensity without appreciable increase in inbreeding. A suitable size for a breeding unit that uses about 8–10 bulls per annum selected at an intensity of 1 in 10 is about 200–300 cows.
Short generation intervals: The use of old bulls simply because they are impressive and heavier compared to younger bulls is genetically unsound. If a breeding programme operates efficiently, a bull of superior genetic merit should sire better performing sons and therefore use of a bull for more than one year is likely to reduce the rate of improvement in the long run. Bulls should therefore be replaced rapidly to minimize generation interval. Selected young bulls should be used for breeding as early as possible.
Above average genetic merit of the population. This is important if a closed herd selection scheme is adopted. If the herd is to be opened, selected outside bulls may be introduced to a small part of the herd and their progeny compared with those sired by home-bred bulls.
A selection scheme for beef cattle can operate without the paternity of the calves being known. Where sire information is necessary (to increase the accuracy of estimation of breeding values), well fenced paddocks are required to facilitate single-sire matings.
A limited breeding season of about 8 weeks is an advantage particularly in mediumsize herds (200–300 cows), since calves are raised under uniform conditions and adjustments for environmental effects may be unnecessary. Where calvings occur all the year round, comparisons should be made only among contemporaries if reasonably large groups are available or data are adjusted for environmental effects.
Calves should be weaned at about 6 months and their weight and date of weaning recorded. Cows may be culled at this time on reproductive criteria, soundness, age and calf performance. The weaned calves should be placed on a nutritional regime which, while being adequate for expression of genetic variation, is not too different from commercial practice. They should be weighed at about 18 months of age and ranked on the basis of the adjusted weights. Tests for heat tolerance and tick resistance may be incorporated at this stage using similar procedures as in dairy cattle (Section 4.2.4). Selection in females is made on the basis of 18-month weight but animals that fail to conceive during the breeding season (if a limited season is used), or within a specified period after they reach a suitable breeding weight, should also be culled. Matings among sibs, and if possible, among more distant relatives should be avoided.
Over 80 percent of the genetic progress in this type of selection scheme is due to selection of bulls. This can be achieved without much record keeping. The only data required are date of birth, dates of weighing and weaning and 18-month weights. No records are accumulated on cows, these being culled on their current performance only. Ignoring age-of-dam effects may not cause any serious bias in the ranking of bulls on 18-month weight except perhaps in bulls produced by primiparous dams.
Nucleus breeding scheme
This scheme seems attractive for the tropics because of the limited scope for recording. In most instances, the only data available may be those obtained from questioning of farmers. Thus animals selected from the base herds may be chosen on the maternal criteria of regularity of calving and calf survival. An “open nucleus” approach may be used in selection for disease resistance such as trypanotolerance since the degree of exposure to disease is greater in base herds. Animals chosen from them may therefore be genetically superior to those in the nucleus, if the degree of disease challenge in the latter is low.
Central testing stations
A test station in the tropics to measure growth rate can consist simply of a grazing area with the necessary weighing scales, a cattle crush and other ancillary equipment. In many tropical countries, these can be established without much capital outlay by converting those government breeding stations that are too small in terms of herd size to be useful for selection programmes. Test stations should preferably be set up in areas representative of the environment in which animals are kept. This is important in breeds selected for disease resistance so that productivity can be measured under conditions of natural disease challenge. The problem of inbreeding is severe in some tropical herds because not only is one bull used at a time but entry of outside bulls is also not permitted. Farmers should therefore be encouraged to exchange their tested bulls.
Dairy breeds are sometimes selected for both milk and beef as in many European countries where dairy animals are an important source of beef. In testing for both traits, the dairy progeny test is preceded by the performance test and between a third and a half of the young bulls are culled. The remaining bulls are then progeny tested for milk yield by the usual procedure. There is no major genetic antagonism between milk yield and growth rate (Mason, 1964) but selection for increased growth rate would increase mature size. The advantage in terms of growth rate has therefore to be offset against the increased maintenance requirement of the larger dairy animal. The breeding scheme described in Section 4.2.3 for the Sahiwal herd in Kenya is suitable for a dual testing programme in institutional herds in the tropics.
All breeds of sheep are likely to produce surplus animals for meat, but with some the main emphasis is on wool, with some on milk, while others again are dual- or even triple-purpose.
Genetic improvement work in sheep in developed countries in the early days concentrated on growth and meat conformation, or on wool weight and quality. It has been realized, however, that reproductive rate is the most important factor, whether wool or meat is the main product and that selection on high wool or body weight without adjustment for litter size at birth could lead to selection against twins and so a lowering of reproductive rate (Turner, 1961). This realization has led to selection programmes aimed at increasing the reproductive rate of pure breeds (Turner, 1977) and to crossing programmes with prolific breeds such as the Finn (Donald et al., 1968).
The principles and the methods used are similar to those for beef cattle.
Within-flock selection: One of the well operated schemes for selection within flocks is the Sheeplan in New Zealand (Clark and Rae, 1977). It involves a recording scheme in which measurements are taken of four important traits of productivity - number of lambs born, weaning weight, weight at 15–16 months of age and fleece weight at the same age. The first trait is mandatory for all users while the others are optional. The data are adjusted for environmental effects and selection lists made up for each flock. The criteria used are weaning and 15–16 month weights for meat breeds used in terminal crossing while for dual-purpose breeds all four traits are used. They may be combined into an aggregate index by methods discussed earlier.
Central testing stations: Performance testing stations are not used to the same extent in sheep as in beef cattle. Where they are used, ram lambs are tested and ranked for final body weight and fleece weight. Some testing stations are also used for progeny testing, e.g. in the Federal Republic of Germany. Progeny groups are brought in as lambs at 60 days of age and assessed for weight gain, feed conversion and carcass quality.
Progeny testing: Improvement of milk yield in dairy sheep and goats requires progeny testing. It is used in some countries, e.g. France and Israel, but it is only where AI is widespread that sufficient progeny records can be obtained for a satisfactory progeny test. Progeny testing by a novel procedure described as the “ram circle” and specially designed for small flocks is used in Norway for improvement of growth rate, carcass quality and wool. Several neighbouring farms cooperate to form a ram circle of about 300–400 ewes in which about 10 rams are progeny tested. Each flock gets one ram on the first mating day and starting from the second day, the rams are circulated each day till the end of the mating season. By this means a fairly accurate estimate of the breeding values of rams bred in different flocks is obtained.
Nucleus breeding schemes: These schemes, also known as cooperative schemes, are becoming more widespread in New Zealand and are also being tried out in Australia. Some comparative studies of the genetic progress between nucleus programmes and conventional selection schemes made in New Zealand seem to bear out the superiority of the former, as expected on theoretical grounds.
Tropical sheep breeds grow a coat of hair with little or no wool, or they produce coarse, carpet-wool fleeces containing medullated fibres and kemp. They are exploited for wool, meat, skins, pelts and sometimes milk. The market for fat lamb is poor, but there is a growing demand for mutton from older animals. Lean meat is often preferred though in some countries the fat tail of indigenous sheep is still considered a delicacy. The coarse fleece is used for making carpets, rugs, blankets, tents or even local clothing. There is a high demand for carpet wool in world markets which warrants the keeping of these tropical carpet breeds and improving their productivity. Skins from tropical sheep are used for manufactured goods (e.g. gloves), while the pelt from one particular breed, the Karakul, is highly prized as Persian lamb.
An important characteristic of tropical sheep is their adaptation to the environment. The harsh, arid tropical environment exerts a profound influence on the productivity of sheep. Nutritional deficiencies and climatic extremes impose a restriction on production during much of the year. Thus, the most important economic characteristic required in tropical sheep kept under current management systems is adaptability to the environment. Tropical breeds have acquired this characteristic through a long period of natural selection and therefore sheep improvement programmes in the tropics should rely largely on genetic improvement of adapted breeds.
In formulating breeding schemes, certain flock characteristics inherent in the production systems in the tropics should be recognized. In general, flock sizes are large in breeds kept extensively, but recording in these flocks is often impractical. On the other hand, the flock sizes of breeds kept semi-intensively in backyards are too small for effective selection procedures. Some of the most prolific breeds of sheep (Mason, 1980a) are kept under these systems, e.g. D'man of Morocco, Priangan of Java, Barbados Blackbelly of tropical America. Breeding programmes should therefore give consideration to selection in institutional flocks or group breeding schemes.
Among the selection objectives for sheep, reproductive rate has the highest priority, as previously discussed, whether animals are raised for meat, wool or skins. Reproductive rate may not be as important where milk is the main product and lambs are removed early, except that a higher reproductive rate contributes to increased selection intensity.
Reproductive rate can be increased either by raising the number of offspring at a parturition, or by decreasing the period between parturitions. In tropical countries, where breeding seasons are long, and nutritional levels frequently low, the second may be the more practical course, as a dam might find difficulty in rearing more than one lamb.
Where multiple births are considered, selection for these is more likely to increase reproductive rate than selection against barrenness. Traits associated with reproduction are usually considered to be lowly heritable (Young and Turner, 1965), but in spite of this, response to selection has been obtained (Turner, 1978a).
Failure to lamb at the first lambing in some sheep breeds (e.g. the Australian Merino) may not indicate poor performance; selection should be based on more than one record. On the other hand, ewes which produce more than one lamb at their first lambing are likely to be good performers. Turner (1978a) used a selection procedure for the Merino based on the first three lambings, and Owen (1976) on the first two.
The simplest method of selection for reproductive rate is to select rams from among the multiple-born lambs on the basis of other criteria such as growth rate or wool weight. In the case of ewe lambs, all lambs born in multiple births are chosen and the balance required is made up from single lambs. Ear notching or tattooing may be done to identify multiple-born lambs if they are not individually identified. When full lambing and pedigree records are available, lambs are selected on the basis of a dam index which combines data of all her lambing records (see Section 3.3.3).
The genetic correlation between prolificacy and growth rate is positive and therefore selection for both traits can be done simultaneously without any danger of antagonism. Where selection is for weaning weight, the latter should be adjusted for the effects of age at weaning, type of birth and age of dam. However, when the trait selected is weight at a later age, such as at 15–16 months, adjustments are unnecessary as these weights are not much influenced by the above factors. A selection scheme for prolificacy and growth rate is discussed in more detail below.
The breeding scheme to be described is suitable for institutional and other flocks that have at least 100 breeding females. Even in flocks of this size, it is necessary to have a short lambing season to avoid the need for adjustment factors which cannot be properly estimated if lambing is year round.
A minimum of eight rams should be used if the flock is to remain closed. To minimize the generation interval, rams should be chosen as soon as they reach puberty and replaced every mating season. Assuming that the number of breeding females is about 100 and a percentage of lambs weaned to ewes joined of 100 (a figure likely to be achieved with about 20 percent twinning), the number of lambs of each sex available at weaning is about 50. The first selection of lambs should be on weaning weight. Though selection of all ram lambs from twins is desirable, a few singles may also be chosen to put some selection pressure on growth rate. Thus, at weaning, all twin and about 4–6 single ram lambs are retained and at 15–16 months of age, when the final selection of 8 rams is made, 6 could be twins and 2 singles. Selection of ewe lambs is less intense because of the need to maintain flock size. Thus, all twin-born ewe lambs are retained and the balance made up from singles based on 15–16 month weight.
The nucleus flock should aim to have about 500 ewes. Such a size is necessary not only to make sizeable selection gains but also to meet the requirements of farmers outside the nucleus scheme.
Selection of animals for the nucleus should be based on observation and interviews in the absence of objective data. Thus, ewes selected should at the time of selection be rearing twin lambs and have a history of twinning in one or more earlier lambings. Ram lambs should be selected from among multiple-born lambs on size and constitution. Where the choice of base flocks is wide, it is preferable to select rams from flocks that do not contribute ewes, in order to minimize inbreeding.
The selection schemes outlined so far have been confined largely to meat production. However, prolificacy is as important in wool as in meat production. The selection procedures in wool breeds would be essentially similar except that fleece weight and quality at 15–16 months of age should be substituted for weight as a selection criterion.
The procedures of selection for dairy sheep and goats are similar to those for dairy cattle.
Goats have received less attention in genetic improvement programmes, even though they are more numerous than sheep in some countries. Some of the reasons lie in the difficulties associated with raising goats in large numbers in limited grazing areas as in institutional farms. When kept in this manner, they are more susceptible than sheep to parasites and infectious diseases. The selection schemes proposed for improvement of prolificacy and growth or milk in sheep are also applicable to goats.
![]()
![]()
![]()