![]()
![]()
![]()
Christophe Destombe, Remi Wattier, Davis Bulke and M. Valero,
Laboratoire de Génétique et Evolution des Populations Vegetates,
URA CNRS 1185, SN2,
59655 Villeneuve d'Ascq Cedex, France.
ABSTRACT
Species of Gracilaria are some of the most useful algae in the world, combining the production of valuable agar with a fast growth rate, ease of vegetative reproduction and other attributes favouring its cultivation. When planning to farm these red algae in new areas, it is essential to identify the species correctly and be aware of the biology of the species under consideration. In agarophytes, delimitation of species is very difficult using morphological characteristics and the biology of the species generally depends on local adaptation. In this study, molecular comparison of the Rubisco spacer allowed the identification of the species Gracilaria verrucosa. This method can be easily transferred to other species of Gracilariaceae.
The level and pattern of population genetic structuring and the possibility for local adaptation in Gracilaria verrucosa were analysed by RFLP methods of cytoplasmic DNA. Mitochondrial and plastidial DNA of Gracilaria verrucosa are polymorphic. The analysis of this polymorphism shows that genetic differentiation occurs between close populations of Gracilaria verrucosa even at short distances (about 100m). Consequently, it is very important before planning cultivation, to be aware of the origin of the Gracilaria verrucosa inoculum to use in breeding experiments and farming.
1. INTRODUCTION
When planning to farm red algae in new areas, it is essential to identify the species and to be aware of the biology of the species under consideration. In the economically important red algal agarophytes, the genera Gracilaria and Gracilariopsis include over 170 species (Goff et al. 1994). Identification of genera and species in the family Gracilariaceae is problematic because of their great morphological similarities. Recent approaches to the taxonomy of the Gracilariaceae using traditional criteria such as morphological characteristics and reproductive anatomy led Fredericq and Hommersand (1989a; 1989b; 1990; and Hommersand and Fredericq, 1990) to conclude that Gracilariopsis and Gracilaria are distinct. The determination of species remain, in some cases, very difficult or impossible because the diagnostic characters are absent (e.g. reproductive organs are absent in some strains of Gracilaria). The use of genetic molecular markers could be an additional tool to facilitate species recognition.
For optimal exploitation of a crop species, efficient seeding, a fast growth rate and genetic improvement are of prime importance to the farmer. Growth rates can be affected by the life history phase and the season, while genetic improvement and seeding from spores can be achieved only if the methods of reproduction are properly understood (for review of Gracilariaceae see Kain and Destombe, 1995). Moreover, the biology of the species generally depends on local adaptation therefore on genetic differentiation between populations. The amount of gene flow is a parameter of great evolutionary importance in determining the level and pattern of genetic differentiation of populations. Then, in the same species, different populations (with different growth rates, physiological or reproductive characteristics) can be found.
This paper investigates:
the use of molecular tools to delineate the species Gracilaria verrucosa using the comparison of the RUBISCO spacer sequences; and
the level and pattern of population genetic structuring and the possibility for local adaptation in Gracilaria verrucosa. Genetic differentiation between populations was estimated using the analysis of cytoplasmic DNA by RFLP with the Southern probes method.
2. MATERIALS AND METHODS
2.1 Delimitation of the species
The sources of algae used in the interpopulation study are listed in Table 1. Except for the isolate from Hawaii, the algae is Gracilaria verrucosa (Hudson) Papenfuss (Gracilariales, Rhodopyhta) on the basis of the morphology and reproductive anatomy which have previously been ascribed to this species (Rice and Bird, 1990; Bird and Rice, 1990). For intrapopulation studies, the spacer was sequenced from five individuals from Cap Gris-Nez and Audresselles, Pas de Calais in the North of France.
Table 1: Sources of algal strains used in this study.
| Strain designation | Geographical location | Date of collection |
| BRZ | Brazil: Sao Paulo | 1988 |
| FR3 | France: Cap Gris-Nez | Jan 1988 |
| NOR | Norway: Oslofjord | Dec 1987 |
| WAL | Wales: Haverfordwest, Dyfed | Aug 1987 |
| ARG | Argentina: Puerto Madryn, Chbut | Jul 1986 |
| NC | USA: Beaufort, North Carolina | Sept 1987 |
| HI | USA: Oahu, Hawaii | May 1988 |
| Five FR isolates | France: Cap Gris-Nez | June 1990 |
| France: Audresselles | June 1990 |
Total DNA was isolated according to the method described by Rice and Bird, (1990) and Doyle and Doyle, (1990). Double-stranded DNA of the spacer region was amplified by polymerase chain reaction (PCR) (Saiki et al. 1988). The primers used were described by Destombe and Douglas (1991) and Destombe (1992). Both strands of the amplified DNA were directly sequenced using a modification (Bachman et al. 1990) of the dideoxy chain termination procedure developed by Sanger et al. (1977) and the DNA sequencing kit “Sequenase 2.0” (U.S. Biochemicals, Cleveland, Ohio).
2.2 Population genetic structuring
The alga used for this study was Gracilaria verrucosa from Europe. The individuals consist of a perennial holdfast fixed to the rock, bearing branched spaghetti-like thalli. Samples of Gracilaria verrucosa were taken from two European countries: France and Norway (Figure 1). Samples were taken from three French localities: Northern France, Brittany and Etang de Thau, a lagoon connected to the Mediterranean Sea. Each locality is about 1,000 km from the others. Within each locality individuals were sampled and mapped in four populations located at two sites. For example in northern France (Pas-de-Calais, on North Sea and Channel coasts), two sites were chosen: Audresselles and Cape Gris-Nez. At each of these two sites, two populations were chosen at the two extremes of the intertidal range of the species, to maximise their environmental difference.
Figure 1: Map of Europe showing where populations of Gracilaria verrucosa were sampled.
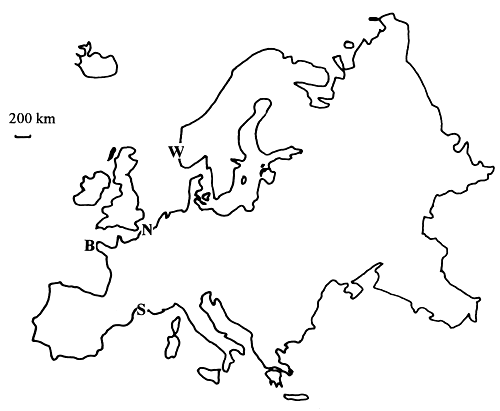
| N: | Northern France region, | B: | Brittany Region in western France, |
| S: | Southern France, Languedoc Region, | W: | Norway. |
High and low populations were noted as Gh and Gl in Gris-Nez, and as Ah and Al in Audresselles, respectively. Individuals of the same population are SO cm to 25 m apart, high and low populations of the same site are approximately 100 m apart (50 to 70 m in Gris-Nez and 150 m in Audresselles due to the difference of slope of the shore in both sites), and the two sites (Audresselles and Gris-Nez) are 3 km apart. Within each population at least five individuals were sampled.
Ecological conditions
In France, ecological conditions differ among the three regions. In Northern France, populations were located in rocky pools. Individuals were fixed to the substratum and range from the lower to mid intertidal level of the shore (Destombe et al. 1989). High and low populations are subject to different tidal and light conditions. In Brittany, individuals from Roscoff are fixed to small pebbles and drift with strong tidal movements, thus experiencing different tidal level conditions during their lifetime and individuals from Island of Batz are fixed in rocky pools. In Etang de Thau, individuals are attached to the substrate just beneath the water surface in the lagoon. They are deprived of tidal fluctuations and are subjected to higher light intensity, temperature, and salinity levels than the two other sites. In Norway and England, individuals were fixed on rocky substrate.
RFLP cytoplasmic methods
The method of RFLP (Restriction Fragment Length Polymorphism) of cytoplasmic DNA (chloroplastic and mitochondrial) was used to study genetic population structuring. Total DNA was isolated essentially as described by Dellaporta et al. (1983) from fresh or frozen material. Total DNA was digested with the restriction endonucleases Ecorl and EcorV in single enzyme digests used according to the instructions of the manufacturers. In addition, the restriction reaction buffer was supplemented with spermidine to 2.5 mM. After separation of the restriction fragments on 0.8% agarose gels, the DNA was transferred to nylon membranes using the vacuum-blot system of Pharmacia. After transfer the DNA was UV cross-linked to the nylon support. The DNA was hybridised with two plastidial DNA probes (CpCh and Ep23) isolated from Chondrus crispus and one mitochondrial DNA probe (MtCh) isolated from Chondrus crispus according to the techniques previously described by Saumitou-Laprade et al. 1993. The RFLP technique allows the detection of structural rearrangements and restriction of site mutations in the region of DNA used.
To estimate the degree of differentiation between populations and regions the Fst method of Weir was applied to haploid data (Weir, 1990). This method gives a Fst coefficient and allows the estimation of genetic differentiation between groups.
3. RESULTS AND DISCUSSION
3.1 Delimitation of the species
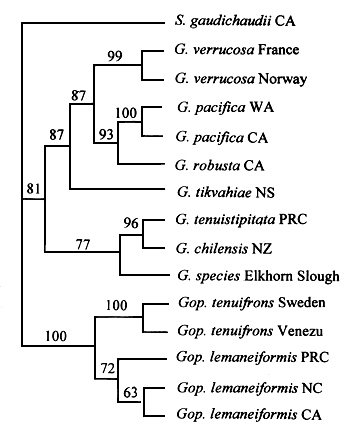
The five individuals from the population from Northern France contain an identical Rubisco spacer region. Within the limitations of our sampling, this method does not reveal any variation among individuals of a population. In contrast, sequence comparisons of the spacer regions from seven isolates from different geographical locations demonstrate the occurrence of four different groups of algae. This finding is confirmed by phylogenetic analysis using parsimony (Figure 2) (Destombe and Douglas, 1991). The first group is composed of a mixture of populations of Gracilaria verrucosa from Western Europe (France, Norway, Wales) and Southern America (Argentina) in agreement with the RFLP analysis and interfertility study conducted by Rice and Bird (1990). The three other groups are distinct both from one another and from the European “core” group. The lengths of the Rubisco spacer vary, in Gracilaria sp. from Hawaii, the length is short (65 nucleotides) (Table 2). In contrast in algae designated originally as Gracilaria verrucosa the lengths of the spacer vary from 106 nucleotides (Brazil) to 131 nucleotides (North Carolina).
Table 2: Sizes (in bp) of Rubisco spacer in Gracilaria.
| Organism | Country | Length |
| Gracilaria verrucosa | Brazil | 106 |
| Europe and Argentina | 115 | |
| North Carolina | 131 | |
| Gracilaria sp. | Hawaii | 65 |
Our results indicate that the sequence and the length of the Rubisco spacer are good additional characters that complete the morphological description of the species. These results were confirmed recently by Goff et al. (1994) who separated Gracilaria and Gracilariopsis populations, species and genera using Rubisco spacer and flanking regions (Figure 3). In this study, the difference between Gracilaria verrucosa from Norway and France is due to only a single insertion/deletion that occurs in the rbcS region.
Figure 2: Phylogenetic tree of isolates of Gracilariaceae constructed using parsimony from aligned plastid Rubisco spacer. A single most parsimonious tree consisting of 89 steps was found. The isolate from Hawaii was used as the outgroup.
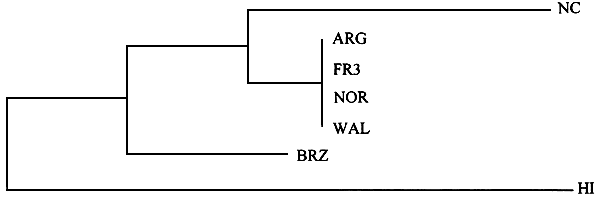
Figure 3: Single most parsimonious tree from aligned plastid Rubisco spacer and flanking regions sequence data (Goff et al 1994). Gracilariopsis sp. are abbreviated “Gop”. Values above branch points are boostrap values (100 replicated). This tree required 82 steps. Saradiotheca gaudichaudii was used as an outgroup.
A study of the length of the Rubisco spacer using PCR is a very easy and very fast method. The main advantage of this approach is that it requires only minute amounts of relatively unpurified template DNA (Goff and Moon, 1993). These authors amplified DNA from herbarium collections.
Population genetic structuring
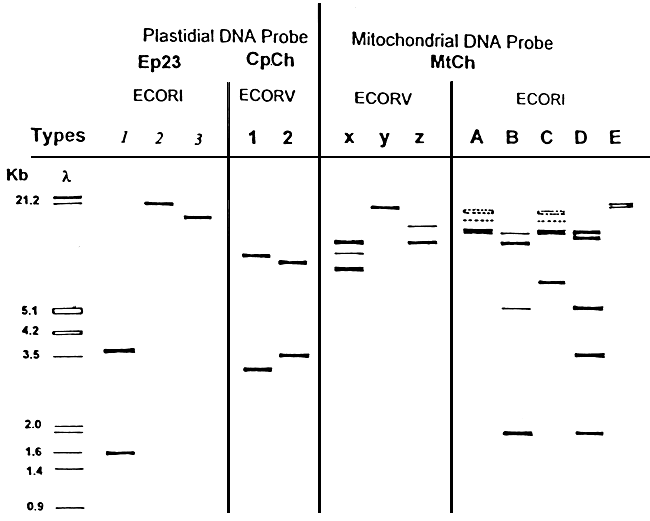
Our results indicate that plastidial and mitochondrial DNA are polymorphic in Gracilaria verrucosa. Four enzyme-probe combinations appeared to be variable (Table 3). The polymorphism is low for the combination Ep23/Ecorl and for Mtch/Ecorl the polymorphism is higher. For a given enzyme-probe combination, there are different profiles. The different profiles obtained are visualised in Figure 4. For each combination, one profile is common and the others are more or less rare.
Table 3: Polymorphism of the 4 enzyme-probe combinations
| Probes | Enzymes | N | Profiles frequencies | |||||
| 1 | 2 | |||||||
| Plastidial | Cpch | EcoRV | 89 | 0.89 | 0.11 | |||
| DNA | 1 | 2 | 3 | |||||
| EP23 | EcoRI | 73 | 0.95 | 0.04 | 0.01 | |||
| x | y | z | ||||||
| Mitochondrial | Mtch | EcoRV | 96 | 0.88 | 0.11 | 0.01 | ||
| DNA | A | B | C | D | E | |||
| Mtch | EcoRI | 86 | 0.82 | 0.04 | 0.01 | 0.12 | 0.01 | |
N: sample size.
Cpch, Ep23 and Mtch: probes isolated from Chrondrus crispus. (C. Boyen Roscoff).
To estimate the degree of differentiation between populations and between regions, the Fst method of Weir (1990) was applied to haploid data. The results are given in the Table 4. Between populations, the F is significantly different to 0 for three combinations (Cpch-EcoRI, Mtch-EcoRV and Mtch/Ecorl). These results indicate that there is genetic differentiation between populations. Between regions, the Fst is significantly different to 0 only for one combination (Ep23-EcoRI). The result obtained for this combination could be due to the small size of the sample and to the low polymorphism of this combination.
Table 4: Genetic differentiation between regions and populations measured by Fst (Weir, 1990).
| Cpch/EcoRV | Ep23/EcoRI | Mtch/EcoRV | Mtch/Ecorl | Mean over loci | ||
| Among | Fst | 0.67** | 0.60ns | 0.56** | 0.46** | 0.57 |
| populations | N | 89 | 70 | 95 | 86 | 4 |
| Among | Fst | 0.06ns | 16* | 0.01ns | 0.02ns | 0.03ns |
| Regions | N | 89 | 73 | 96 | 86 | 4 |
ns: not significant;
*: p<0.05;
** p<0.01.
N: sample size.
For all the combinations (mean over loci), the Fst indicate that genetic differentiation occurs between populations, but that genetic differentiation is not significant between regions.
The Mtch/Ecorl enzyme probe combination is the more informative. The distribution of the 5 haplotypes of this combination between populations and regions were detailed in Table 5. The haplotype A is found in almost all the populations (excepted ROSII and BRI) and the haplotype D is found also in each region. The haplotype B is an endemic type of the island of Batz. The type D is not very frequent but when it is found in one population, it is at a quite important frequency within the population. This explains why there is a high genetic differentiation between populations.
Figure 4: Enzyme-probe combination profiles of Gracilaria verrucosa obtained by
Genetic differentiation occurs between close populations in this species. These results are in accordance with the previous dispersal and crossing experiments in Gracilaria verrucosa (Destombe et al. 1990, 1992 and Richerd et al. 1993). Contrary to the intuition that dispersal in the sea is large, data on recruitment (Santelices, 1990) dispersal potentialities (Destombe et al. 1992), fertilisation distances (Destombe, et al. 1990) and heterosis effects in crosses (Richerd et al. 1993) show that dispersal is leptokurtic and limited to the order of 100 m. This corresponds to the distance between high and low populations in each site that should then be able to diverge genetically. This divergence is linked to differential adaptation, since tidal level has important selective effects on algae.
In conclusion, in future aquaculture applications it is necessary to attempt to determine and study wild populations. This genetic differentiation between populations is important in breeding studies and genetic potential.
Table 5: Distribution of the 5 haplotypes of the Mtch/Ecorl combination between populations and regions.
| Haplotypes | |||||||
| Populations | A | B | C | D | E | N | |
| Gris Nez Site | GNvl | 3 | - | - | 2 | - | 5 |
| GNl | 6 | - | - | - | - | 6 | |
| GNh | 5 | - | - | - | - | 5 | |
| GNvh | 5 | - | - | - | - | 5 | |
| Audresselles site | AU1 | 5 | - | - | - | - | 5 |
| AUh | 3 | - | - | - | - | 3 | |
| Northern region | 27 | - | - | 2 | - | 29 | |
| Island of Batz site | GB | 5 | - | - | - | - | 5 |
| GBm | 6 | 1 | - | - | - | 7 | |
| GBp | 4 | 1 | - | - | - | 5 | |
| Bil | 3 | 1 | - | - | - | 4 | |
| Roscoff site | ROSI | 3 | - | - | 1 | - | 4 |
| ROSII | - | - | 1 | 3 | - | 4 | |
| Brittany Region | 21 | 3 | 1 | 4 | - | 29 | |
| Le Barrou site | BRI | - | - | - | 3 | - | 3 |
| BRII | 9 | - | - | - | - | 9 | |
| Bouzigues site | BZI | 2 | - | - | 2 | & | 5 |
| BZII | 7 | - | - | - | - | 7 | |
| Languedoc Region | 18 | - | - | 5 | 1 | 24 | |
| Norway | 4 | - | - | - | - | 4 | |
REFERENCES
Bachmann, B., Luke, W. and Hunsmann G. 1990. Improvement of PCR amplified DNA sequencing with the aid of detergents. Nucleic Acids Research 18:1309.
Bird, C. J. and Rice, E. L. 1990. Recent approaches to the taxonomy of the Gracilariaceae (Gracilariales, Rhodophyta) and the Gracilaria verrucosa problem. Hydrobiologia 204/205:118–119.
Dellaporta S. L., Wood, J. and Hicks, J. B. 1983. A plant DNA mini preparation: Version II. Plant Molecular Biology Report 1:19–21.
Destombe, C, Valero, M., Vernet, P. and Couvet, D. 1989. What controls haploid-diploid ratio in the red alga Gracilaria verrucosa? Journal of Evolutionary Biology 2: 67–101.
Destombe, C, Godin, J. and Remy, J. M. 1990. Viability and dissemination of spermatia of Gracilaria verrucosa (Gracilariales, Rhodophyta). Hydrobiologia 204/205: 219–223.
Destombe C. and Douglas, S. 1991. Rubisco spacer sequence divergence in the rhodophyte alga Gracilaria verrucosa and closely related species. Current Genetics 19:395–398.
Destombe, C. 1992. Erratum Current Genetics 22:173.
Destombe, C, Godin, J., Lefebvre, C. L., Dehorter, O. and Vernet, Ph. 1992. Differences in dispersal abilities of haploid and diploid spores of Gracilaria verrucosa (Gracilariales, Rhodophyta). Botanica Marina 35: 93–98.
Doyle J.J. and Doyle J.L. 1990. BRL FOCUS
Fredericq, S. and Hommersand, M. H. 1989a. Proposal of the Gracilariales ord. nov. (Rhodophyta) based on an analysis of the reproductive development of Gracilaria verrucosa. Journal of Phycology 25:213–227.
Fredericq, S. and Hommersand, M. H. 1989b. Comparative morphology and taxonomic status of Gracilariopsis (Gracilariales, Rhodophyta). Journal of Phycology 25:228–241.
Fredericq, S. and Hommersand, M. H. 1990. Diagnoses and key to the genera of the Gracilariaceae (Gracilariales, Rhodophyta). Hydrobiologia 204/205: 173–178.
Goff, L. J. and Moon, D. A. 1993. Amplification of nuclear and plastid genes from algal herbarium specimens and algal spores. Journal of Phycology 29:381–384.
Goff, L. J., Moon, D. A. and Coleman, A. W. 1994. Molecular delineation of species relationships in the red algal agarophytes Gracilariopsis and Gracilaria (Gracilariales). Journal of Phycology 30:521–537.
Hommersand, M. H. and Fredericq, S. 1990. Sexual reproduction and cystocarp development. In: K. M. Cole and R. G. Sheath [eds.], Biology of the red algae, 305–345. Cambridge University Press, Cambridge.
Kain J. M. and C. Destombe, 1995. A review of the life history, reproduction and phenology of Gracilaria. Journal of Applied Phycology (in press)
Saiki R. F., Scharf, S., Faloona F., Mullis, K. B. Horn, G. T., Erlich, H. A. and Arnheim, N. 1988. Science 230:1350–1350.
Sanger F., Nicklen, S. and Coulson, A. R. 1977. Proc. Natl. Acad. Sci. USA 74:5463–5467.
Santelices, B. 1990. Patterns of reproduction, dispersal and recruitment in seaweeds. Oceanography and Marine Biology Annual Review 28: 177–276.
Saumitou-Laprade P, Pannenbecker G, Boutin-Stadler V., Michaelis G, and Vernet, P. 1991. Plastid DNA diversity in populations of Beta maritima showing additional variations in sexual phenotype and mitochondrial DNA. Theoretical Applied Genetics 533–536.
Rice, E. L and Bird, C. J. 1990. Relationships among geographically distant populations of Gracilaria verrucosa (Gracilariales, Rhodophyta) and related species. Phycologia 29:501–510.
Richerd, S., Destombe, C, Cuguen, J. and Valero, M. 1993. Variation of reproductive success in a haplo-diploid red alga, Gracilaria verrucosa: Effects of parental identities and crossing distance. American Journal of Botany 80:1379–1391.
Weir, B. S. 1990. Genetic data analysis. Sinauer Ass.
A. Q. Hurtado-Ponce,
Aquaculture Department, Southeast Asian Fisheries Development Center,
Tigbauan, Iloilo 5021, Philippines.
ABSTRACT
Research in seaweed from 1988–1994 focused on the agar producing Gracilaria. Studies in the following areas were also conducted: inventory of the seaweed resources; production ecology; farming systems; and agar characterisation. Six species of Gracilaria and one Gracilariopsis abound in the Western Visayas. There were monthly variations in biomass and agar quality of G. blodgettii, G. manilaensis and G. heteroclada collected at different places.
The bulk of the studies were carried out on Gracilariopsis heteroclada because of its wide distribution, fast growth characteristics and good quality agar. Its reproductive state was seasonal with tetrasporophyte abundant in May and carposporophyte in January. Seventy-five percent harvesting of the available biomass was sufficient for the next cropping season. Harvesting the seaweed using “araña” was not appropriate in places where the biomass was exposed to air during the lowest tide. The addition of nutrients to the stock increased the growth rate of the seaweed and gel strength of the agar. G. heteroclada grown at lower stocking density in hapa nets, both in floating cages and in ponds, gave higher growth rates and production than at higher stocking densities. This species, when polycultured with P. monodon at lower stocking density combinations, gave the highest growth rate and income. Likewise, this species when cultured vertically in ropes inside a floating cage showed encouraging results. G. heteroclada, when grown at 24–25 ppt under tank conditions, produced the highest gel strength.
1. INTRODUCTION
During the Aquaculture Development in Southeast Asia (ADSEA) Seminar-Workshops in 1987, 1991 and 1994, the member countries of the Southeast Asian Fisheries Development Center (SEAFDEC) recommended Gracilaria as the number one priority for seaweed research in the following areas: (1) basic biology and production ecology; (2) refinement of culture techniques; (3) screening and characterisation of natural products; (4) product utilization; and (S) biofiltration. The Seaweed Unit of the Department was re-activated in 1988 and studies along these lines have been conducted.
1.1 Taxonomy
In order to determine the existing resources of the region, an inventory of the macrobenthic algae of Panay was conducted. Among the species identified, seven were Gracilaria, namely: Gracilaria arcuata Zanardini; G. changii (Xia et Abbott) Abbott, Zhang and Xia; G. coronipofolia J. Agardh; G. eucheumoides Harvey; G. manilaensis (= G. “verrucosa”) Yamamoto et Trono sp. nov.; G. salicornia (C. Agardh) Dawson; and Gracilariopsis heteroclada (= Gracilaria heteroclada) (Zhang et Xia) Zhang et Xia (Hurtado-Ponce et al 1992a). Occurrence of these species ranges from full seawater on rocky-coral substrate to brackishwater on sandy-muddy substrate.
Among the species identified in western Visayas, G. heteroclada was the most widely distributed from mudflats, estuarine, rivers, drainage canals to ponds. Consequently, several studies were conducted on this particular species.
1.2 Production ecology
Reproductive state
Studies on production ecology are necessary to have a better understanding of the basic biology and ecology of an organism so that management schemes may be formulated for its commercial farming. One study was done on the reproductive state of G. heteroclada to determine the monthly occurrence of fertile plants. Plants originating from spores are more viable sources of new plants. Results showed that tetrasporophyte (64%) and carposporophyte (48%) were more abundant in May and January, respectively. Percentage occurrence of tetrasporophyte and carposporophyte did not significantly correlate with water temperature, salinity, and turbidity (Luhan, unpubl. data).
Stock assessment
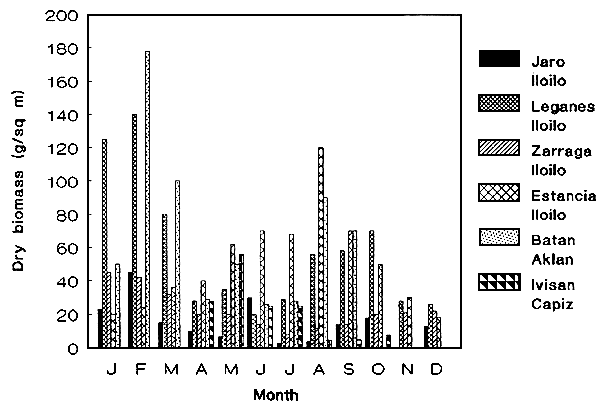
Studies on the monthly biomass of wild Gracilaria species were assessed to determine the amount of available biomass that could possibly support an industry. Variations due to culture months and geographical locations were factors which influenced differences in biomass (Figure 1) (de Castro et al 1991, Pondevida unpubl. data). Among the species studied, G. heteroclada gave the highest dry biomass and it is available throughout the year (Figure 2).
Regeneration capacity
The amount of harvest left after the first cropping is important in determining the amount of biomass available for the next cropping season. A 75% harvest provides an appropriate amount of “seeds” for the next cropping season compared with 25, 50, and 100% (Hurtado-Ponce, 1993). The amount of biomass to be harvested during each harvest regime should not exceed the amount of biomass available for cropping.
Harvesting tools
The use of harvesting tools in the gathering of wild Gracilaria in places where it is convenient and effective improves the regeneration capacity of the seaweed and increases the harvest (Santelices et al. 1984). Such tools (araña) were not proven to be effective in places where the seaweed is exposed to air during the lowest tide (Hurtado-Ponce, 1994). Raking the seaweed as reported by Hurtado-Ponce et al. 1992) brings excessive disturbance on the substrate and to the population (Luxton, 1981). Gathering with bare hands is a much better means although it is tedious.
The addition of nutrients to the seaweed is one of the management schemes to increase its growth characteristics. This was substantiated when G. heteroclada was cultured in tanks at different stocking densities. The addition of nutrients was significant at lower stocking density (500 g m-2) than at higher density (1000 and 2000 g m-2) (Chavoso and Hurtado-Ponce, unpubl. data). The enriched stock gave higher growth rates than the unenriched ones.
Salinity tolerance
Salinity tolerance studies on 3 species of Gracilaria was done to formulate a management scheme for cultivation. Each species tolerated a wide range (15–35 ppt) of salinity, however, it is species-specific. Optimum growth was obtained at 15 ppt in G. tenuistipitata, 24–25 ppt in G. changii and G. heteroclada, and 35 ppt in G.firma.
Environmental parameters
Production ecology of the seaweed is incomplete without considering environmental factors that influence the growth of the species both cultivated and in natural beds. The single and interactive effect of the different factors were analysed and correlated with one another to determine the relationship of the seaweed with its environment. Correlation analysis showed that temperature and pH had no effect on biomass; salinity is negatively correlated with biomass; while rainfall is inversely correlated with the biomass of wild G. heteroclada (de Castro et al 1991, Pondevida, unpubl. data). The same environmental factors had no effect on the growth rate and production of G. heteroclada when grown in net cages inside a brackishwater pond (de Castro and Guanzon, 1993).
Figure 1: Stock assessment of wild G. blodgetti and G. manilaensis.
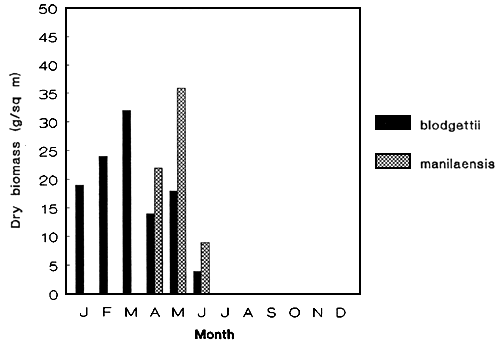
Figure 2: Stock assessment of wild G. heteroclada.
1.3 Farming systems
Indiscriminate or uncontrolled gathering of Gracilaria from the wild leads to the depletion of the resources. Gathering of agarophytes (like G. blodgetti, G. coronopifolia, G. eucheumoides, G. salicornia and Gracilariopsis heteroclada) during the dry season has been reported (Hurtado-Ponce et al. 1992b). Although commercial cultivation of G. heteroclada both in ponds and drainage canals has been reported (Hurtado-Ponce 1992c), other farming techniques need to be developed both land-based and in open sea, while existing techniques need to be refined. These are all necessary in order to have a sustainable industry.
Attempts were made to cultivate G. heteroclada using different systems to develop an economically feasible technique.
Cage culture
A study was done on G. heteroclada using vegetative fragments to determine the effect of spacing interval on growth. Results show that significant differences in growth and monthly yields were observed when the seaweed was cultured on vertical ropes at 3 spacing intervals (10, 15 and 20 cm) inside a floating cage. Maximum yields were obtained in April at 10 and 15 cm and in February at 20 cm (Hurtado-Ponce, 1990).
Another study was carried out to test the effect of the presence of sea bass as biological control to grazers of G. heteroclada. The interactive effect of the presence of sea bass, water depth and month of culture significantly affected the growth G. heteroclada (Hurtado-Ponce 1994a). Highest and lowest growth rates were recorded with seabass at 25 and 100 cm below the water surface, respectively.
Manipulating the stocking density of a species will yield different growth rates and production (Santelices et al. 1993). To confirm this statement, a study was done on G. heteroclada using hapa nets installed to a floating cage. The results showed that lower stocking rates (400 and 500 g m-2) yields higher growth rates and production than higher stocking densities (600 and 700 g m-2) (Guanzon and de Castro 1992). Highest production was obtained during the dry season.
Pond culture
When the same stocking densities were tried in ponds using the same species, similar results were obtained when G. heteroclada was cultured in net cages inside a brackishwater pond. Higher growth and production was obtained at lower stocking densities (400 and 500 g m-2) than at higher stocking densities (600 and 700 g-2). Higher production was obtained during the dry season than during the wet culture months (de Castro and Guanzon, 1993). To ascertain whether G. heteroclada and Penaeus monodon can be cultured together in brackishwater ponds using the “rice planting” and long line methods, a five-month study was done at different stocking combinations. A lower stocking density combination of G. heteroclada (250 g m-2) and P. monodon (2,500 pcs ha-1) gave higher growth rates and production of the seaweed but a lower production of shrimp. Income above variable cost was also highest at this stocking density combination. Growth rates and production of the seaweed was significantly affected by the culture month and stocking density (Hurtado-Ponce, 1995). Higher growth rates and production were obtained at 24–29 ppt salinity confirming the results of an earlier study by Hurtado-Ponce (1994b) in tanks.
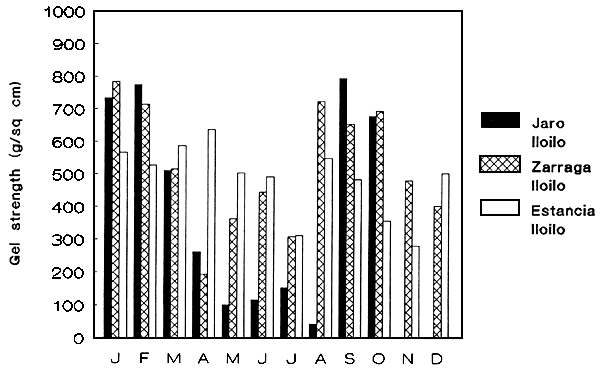
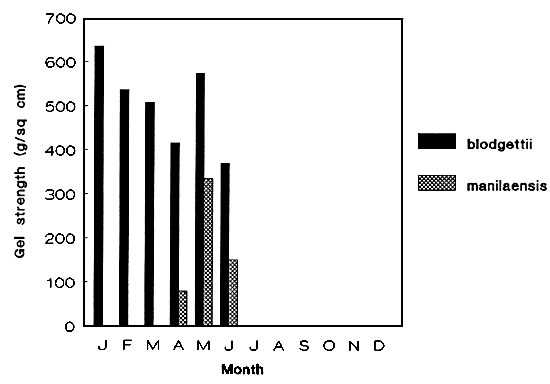
Figure 3: Gel strength of wild seaweed populations.
a. G. heteroclada
b. G. blodgettii and G. manilaensis
Drainage canal
Maximising use of drainage canals for the cultivation of seaweeds will increase production. A study on G. heteroclada using long-line, a deviation from the existing practice, revealed a 4.5% day-1 growth, a yield of 3,440 kg/ha, an annual production cost of 4,120 Pesos, annual net returns of 17,552 Pesos, a return on investment of 566% and a pay back period of 2 months (Hurtado-Ponce et al, unpubl. data).
1.4 Agar characterisation
Attempts to screen several species of Gracilaria as source of good quality agar were made both from the wild (Figures 3a-b) and cultivated strains. The effect of pre-alkali (NaOH) treatment (concentration and time) to the seaweed was species-specific (Table 1). G. heteroclada cultured at 24–25 ppt under tank conditions gave the highest gel strength (807–850 g cm-2). Among the species tested for agar both wild and cultured, G. heteroclada is the best source.
2. RECOMMENDATIONS
As mandated during ADSEA 94, the Aquaculture Department of SEAFDEC will continue to conduct research studies on Gracilaria. As the number one priority commodity among the seaweeds, studies on the following areas will be conducted:
inventory of other species, selection and development of highly productive cultivators with high quality agar;
bioremediation (biofilter) for water quality control in aquaculture;
monoculture and polyculture in ponds;
refinement of production methods;
product utilization;
village municipal processing (semi-refined);
socio-economic studies;
development of hatchery technology; and
genetic studies and creation of seedbank.
REFERENCES
de Castro, T. R. 1993. Evaluation of agar from three species of Gracilaria from Panay and Guimaras Islands. Philipp. Scientist. 30: 41–47
de Castro, T. R. and N. G. Guanzon Jr. 1993. Growth of Gracilaria sp. (Gracilariales, Rhodophyta) in brackishwater ponds at different stocking densities. Israeli J. Aqua.-Bamidgeh. 45: 89–94.
de Castro, T. R., N. G. Guanzon Jr., and Ma. R. J. Luhan. 1991. Assessment of stocks of a natural Gracilaria population on Panay Island, Philippines. Bot. Mar. 34: 383–386.
Guanzon, N. G. Jr and T. R. de Castro. 1992. The effects of different stocking densities and some abiotic factors on cage culture of Gracilaria sp. (Rhodophyta, Gigartinales). Bot. Mar. 35: 239–243.
Hurtado-Ponce, A. Q. 1995. Polyculture of Gracialriopsis heteroclada and Penaeus monodon in brackishwater ponds. In 15th Internal. Seaweed Symposium, Valdivia, Chile (Abstract only).
Hurtado-Ponce, A. Q. 1994. Seaweed: Gracilaria. In: F. Lacanila, R. M. Coloso and G. F. Quinitio (eds.) Proceedings Aquaculture Development in Southeast Asia and Japan and Prospects for Seafarming and Searanching. 46–50.
Hurtado-Ponce, A. Q. 1994. Agar production from Gracilariopsis heteroclada (Gracilariales, Rhodophyta) grown at different salinity levels. Bot. Mar. 37: 97–100.
Hurtado-Ponce, A. Q. 1993. Harvesting Gracilariopsis heteroclada (Gracilariales, Rhodophyta) in Iloilo, Philippines. Philipp. J. Sci. 122: 413–423.
Hurtado-Ponce, A. Q. 1990. Vertical rope cultivation of Gracilaria (Rhodophyta) using vegetative fragments. Bot. Mar. 33: 477–481.
Hurtado-Ponce, A. Q., Luhan, Ma. R. J. and Guanzon, N. G. Jr. 1992a. Seaweeds of Panay. SEAFDEC/AQD Monograph, Tigbauan, Iloilo.
Hurtado-Ponce, A. Q., Luhan, Ma. R. J. and Guanzon, N. G. 1992b. Gathering of economically important seaweeds in western Visayas, Philippines. Philipp. Scientist 29: 40–47.
Hurtado-Ponce, A. Q., G. PB. Samonte, Luhan, Ma. R. J. and Guanzon, N. G. 1992c. Gracilaria (Rhodophyta) farming in Panay, western Visayas, Philippines. Aquaculture, 105: 233–240.
Luhan, Ma. R. J. 1991. Assessment of the natural beds of Gracilaria heteroclada (Zhang et Xia) in Jaro, Iloilo. Master of Aquaculture Thesis, University of the Philippines in the Visayas, Miag-ao, Iloilo.
Luhan, Ma. R. J. 1992. Agar yield and gel strength of Gracilaria heteroclada collected from Iloilo, Central Philippines. Bot. Mar. 35: 169–172.
Luxton, D. M. 1981. Experimental harvesting of Gracilaria in New Zealand. Internal. Seaweed Symp. 10: 693–698.
Pondevida, H. B. 1993. Influence of some environmental factors on growth rates and agar quality of selected Gracilaria species found in Iloilo. Ph. D. Dissertation, University of the Philippines, Institute of Biology, Diliman, Quezon City.
Santelices, B., Westermeier, R. and Bobadilla, M. 1993. Effects of stock loading and planting distance on the growth and production of Gracilaria chilensis in rope culture. J. Appl. Phycol. 5: 517–524.
Suwalee Chandrkrachang,
Asst. Professor, Biopolymer Research Unit Department of Chemistry, Faculty of Science,
Srinakarinvirot University, Prasanmitre, Sukhumvit, Bangkok.
ABSTRACT
Seaweeds are a major coastal resource which are valuable to both the economy and environment of many countries. The uses of the seaweeds as raw materials for phycocolloid production has become a major world-wide industry involving food, cosmetics, drugs and biotechnology. The most valuable product of phycocolloids is the agar, which is produced from red seaweeds of the family of Gracilaria, involving different species. Agar consists of heterogeneous biopolymers containing galactose units and their derivatives. The different market prices of the agar results from different grades, which depend on the qualities of gel strength, clarity and the quantities of charged particles in the agar composition. Different techniques of agar extraction are reviewed in this paper. Samples of seaweed and agar from NACA member countries participating in the regional study were sent for analysis at BRU. The results will be used as guidelines for the development of both quality and quantity of the raw seaweed materials and their agar products, which are essential information for research study and public services.
1. INTRODUCTION
Seaweeds, or marine macroalgae, are valuable coastal resources with respect to marine ecology, environment and human consumption. The utility of seaweed has been known since ancient times, varying with time and geographic location. There are so many variations in seaweed species, their habitats, availability, methods of harvesting and processing, chemical properties and product utilization, it is quite impossible to consider the seaweed industry as a homogeneous entity. Processing is dominated by relatively few major commercial enterprises who keep their production methods and figures confidential as the market for seaweed colloids is highly competitive.
Presently, the extraction of valuable products is the most rapidly developing area of seaweed utilization, which will lead to world-wide commercialisation of seaweeds.
1.1 Phycocolloids
Phycocolloids are chemical polysaccharides or carbohydrates which have the ability to give viscosity, gel strength and stability to aqueous mixtures, solutions and emulsions. With such properties, they play increasingly important roles in the production of processed foods, dairy products, cosmetics, drugs, fertilisers, animal feeds and other valuable commodities. Agar, a high-priced phycocolloid, is a galactan-type biopolymer produced from red seaweeds of different families in the Class Rhodophyceae. Other strongly negatively charged polymers of galactan, derived from different families of red seaweeds, are carrageenans and furcellarans. The most important colloid product from brown seaweeds is the alginate which is widely used in the textile, printing, paint and pharmaceutical industries.
1.2 Agar
Agar, a traditional gelling agent, may be defined as a hydrocolloid of ancient origin. Chemically, agar consists of neutral and negatively charged polymers of galactan. Agar is soluble in hot water but becomes water-insoluble at room temperature. One of its unique characteristics is gel formation at very low concentration. With the substitution of the negative charge in the polymer chain of galactan, agar is considered as one of the anionic biopolymers from marine resources.
The traditional concept of agar being comprised of neutral agarose and negatively charged agaropectin, is now regarded as an over simplification. Many research results indicate that agar is a complex mixture of polysaccharides all with the same backbone of linkages but substituted to a variable degree with negatively charged groups of some sulphate, pyruvate and methoxy substitutions. The agarose has been suggested as the polymer containing the agarobiose structure of 1,3-linked β-D-galactophyranose and 1,4-linked 3,6-anhydro-α-L-galactopyranose. However, the naturally-occurring material is variously substituted with half sulphate groups, pyruvic acid, ketal and methyl esters which refer to the negative charged agaropectin in the composition of the agar. Dockworth and Yaphe (1971), recommended a practical definition of agarose:
“ --- not a neutral polysaccharide obtained by fractionating it away from the charged polymer agaropectin, but rather that mixture of agar molecules with lowest charge content and, therefore, the greatest gelling ability fractionated by a whole complex of molecules, called agar, all differing in their extent of masking with charged groups.”
The basic repeating unit of agarose, agarobiose, is shown in Figure 1.
Figure 1: Chemical structure of “agarobiose” unit
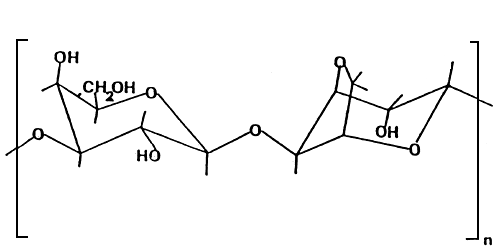
The chemical nature of agar which, affects its quality, varies according to the seaweed raw materials, the environment where the seaweeds grow and on the methods of extraction of the agar. Originally, Gelidium was the major source of agar, because it gave superior quality of agar gel even by the traditional simple method of extraction. However, availability of Gelidium is now limited. Since 1950, Gracilaria spp, which is abundant along coastal areas particularly in the Asia-Pacific region, have become important sources of agar production. However, further modification of alkali treatment of Gracilaria spp. to improve the gelling ability needs to be done during the extraction process. Different alkali treatment techniques are now commonly used in agar factories. The most common concentration (pre-alkali treatment) is between 3 to 5 % NaOH at a temperature of 80–90°C for about one hour.
An alternative technique developed by the Biopolymer Research Unit (BRU) research team is strong alkali treatment of agar at relatively low temperatures after traditional extraction. With this technique, the low grade crude agar can be refined to obtain high quality agar.
2. AGAR EXTRACTION TECHNIQUES
Since seaweed raw materials are different depending on the species and environmental conditions during growth, optimum conditions for extraction vary from source to source. Furthermore, the purity of the harvested seaweed as well as storage and handling conditions prior to processing, also affect the quality and quantity of the agar product. Laboratory tests are essential to optimise suitable conditions for industrial agar production as the seaweeds need to be carefully evaluated for their agar quality. Samples of the seaweed are taken and tested for moisture content, purity, agar yield and other qualities. The most common method involves the following steps:
Cleaning and washing to remove impurities.
Chemical treatment - some hard seaweeds need acid modification but most of the Gracilaria species require the alkali treatment for desulphation to improve agar gel strength. Some seaweeds do not need any treatment.
The extraction step is carried out after washing out the acid or alkali.
Filtration and gelation. Filtration techniques depend on the scale of extraction. In small scale production, the simple pressing tool and the pressure filter column can be used. Most industrial scale productions need plate and flame pressure filter units. During filtration, a hot and clear solution should be obtained and then cooled to set the gel at room temperature or below.
Dewatering of the gel - the hydraulic pressing machine is the most common piece of equipment used in the agar industry. The freezing and thawing processes are done naturally during winter time for strip-agar production in some countries.
In order to obtain a good quality agar product, the seaweeds are generally sorted by hand and then thoroughly washed with fresh water, followed by sun-drying for natural bleaching. The industrial methods of agar extraction by traditional and pressure techniques, are shown below:
| Traditional method | Pressure method |
| (Gelidium + Gracilaria) | ( Gracilaria ) |
| Hard Seaweed | Seaweed |
| ↓ | ↓ |
| Boiling with water | Treating with alkali |
| ↓ | ↓ |
| Adding soft seaweed | Washing |
| ↓ | ↓ |
| Extraction | Extraction in open system |
| ↓ | ↓ |
| Filtration | Filtration |
| ↓ | ↓ |
| Coagulation | Coagulation of agar |
| ↓ | ↓ |
| Cutting to Square or Strip | Mechanical dehydration or Freeze - Thawing |
| ↓ | |
| Dehydration by natural freezing and thawing for 7–8 days | ↓ |
| Drying by heat | |
| ↓ | |
| ↓ | Crushing |
| Packing as square or string agar | ↓ |
| Packing as powder agar |
2.1 Small-scale agar production
The sequence of processes involved in the extraction of agar-agar from seaweed on a small-scale is summarised in the following flow diagram. The resultant product is classified as crude agar. It is normally used in the preparation of desserts, or sold as a raw material to be refined into higher-grade products. The yield is about 20–25% by weight of the initial dried seaweed.
Small-scale processing of seaweed for the extraction of agar
| Dry seaweed |
| ↓ |
| Wash with water |
| ↓ |
| Sun dry |
| ↓ |
| Repeat the wash/dry process several time |
| ↓ |
| Cleaned dry seaweed |
| ↓ |
| Soak in water |
| ↓ |
| Boil |
| ↓ |
| Grind |
| ↓ |
| Boil |
| ↓ |
| Add filter aid |
| ↓ |
| Boil |
| ↓ |
| Filter by pressing |
| ↓ |
| Cool to set |
| ↓ |
| Crude agar-agar gel |
| ↓ |
| Cut into smaller pieces |
| ↓ |
| Press to dewater |
| ↓ |
| Sun dry |
| ↓ |
| Grind to powder |
| ↓ |
| Crude agar powder |
3. APPLICATIONS OF AGAR RELATED TO ITS QUALITY
Agar has wide ranging applications in many areas of the food, Pharmaceuticals and biotechnology related industries, as detailed below:
Food industry: Agar is used in the food industry as a stabilising, thickening and gelling agent. It is used directly in different kinds of desserts in Thailand. It functions as a texture-improving, setting and stabilising agent for the bakery and candy industry. Agar of high gel strength is always preferred for food preparation in order reduce the quantity required and costs.
Tissue culture: The use of agar as a medium for tissue culture is very important to improve the consistency and standard methods for propagation of orchids and other ornamental plants, vegetables and other agricultural crops. Agar with high gel strength is preferred because of the low concentrations of agar required and low cost.
Microbiology and pharmaceutical uses: An important application of agar is as a culture medium for micro-organisms, bacteria and fungi. Agar has the unique characteristic of gel firmness and stability and it is not easy to metabolise. Agar for bacterial culture needs to be clean, clear and low contamination. The temperature for gel setting should be equal or lower than human body temperatures (37°C). Agar with low gelling temperatures are preferred for bacteriological culture mediums.
Preparation of agarose: Agar is an essential source for agarose which is defined as a neutral biopolymer. Agarose is the high value added bio-product used as a medium in biotechnology. Agarose gel media are used in electrophoresis for separation of nucleic acids, specific proteins, virus and other genetic materials. Agarose is a biologically inert material with low ionic properties. Agar for agarose production needs to have a low sulphate content and relatively high gel strength. Agarose beads, prepared from agarose gel, are used in gel filtration for molecular weight determination and separation of specific protein mixtures and other biological materials. Agarose is used extensively in immunology techniques of identification such as electroimunoassay immuno-electrophoresis and counter electrophoresis, including the modern techniques of separation and identification of genetic materials.
4. INTERNATIONAL AGAR SPECIFICATIONS (Chandrkrachang and Chinadit, 1988)
The most important properties to be considered when determining the quality of agar for food, tissue culture or bacteriological use are gel strength, gelation and melting temperatures, sulphate and methoxyl content, clarity of the solution and ash content (Tables 1 and 2). An agar of high gel strength is always preferred for food. For bacteriological media, a gel strength of 300–400 g/cm at a 1% concentration is acceptable. Agar, when dissolved in hot water, should form a clear solution, the clarity of which can be determined by measuring the transmittance of the solution.
Table 1: Major parameters of the Japanese specification for processed agar.
| Criteria | Grade | |||
| Special | 1 | 2 | 3 | |
| Gel strength (g/cm2) | ≥600 | ≥350 | ≥250 | ≥150 |
| Water content (%) | ≤22 | ≤22 | ≤22 | ≤22 |
| Crude protein content (%) | ≤1.5 | ≤1.5 | ≤2 | ≤3 |
| Solids insoluble in hot water (%) | ≤0.5 | ≤2 | ≤4 | ≤4 |
| Crude ash (%) | ≤4 | ≤4 | ≤4 | ≤4 |
Table 2: Agar specifications of the United States Pharmacopeia (USP) and Food
| Criteria | USP | FCC |
| Microbial limit (Salmonella) | negative | - |
| Maximum water (%) | 20 | 20 |
| Maximum total ash (%) | 6.5 | 6.5 |
| Maximum acid-insoluble ash (%) | 0.5 | 0.5 |
| Maximum foreign inorganic matter (%) | 1 | - |
| Maximum foreign insoluble matter (%) | 1 | 1 |
| Arsenic (ppm) | 3 | 3 |
| Lead (ppm) | 10 | 10 |
| Heavy metals (ppm) | 40 | 40 |
| Foreign starch | negative | negative |
| Gelatin | negative | negative |
| Maximum water absorption (ml) | 75 | 75 |
Higher grade agar is white in colour, while light yellow is allowed for lower grades. Consistent quality and size are also important criteria.
6. ANALYSIS OF AGAR QUALITY FROM GRACILARIA OF PARTICIPATING COUNTRIES
The participating countries used the same methods of agar extraction together with some analysis of seaweed quality to allow an evaluation to be made of their local Gracilaria species. Some of the seaweed and agar samples were sent to BRU for analysis. The results of the analysis are shown in Table 3. Some important properties which reflect the quality of agar and the process of agar extraction are gel strength, sulphate content and the clarity of the agar solution. Since the seaweeds are of many variations in different geographic locations and seasons, it is impossible to consider and make an evaluation in the same, or similar, directions. Each country needs to develop and improve extraction techniques which are suitable to their seaweed raw materials. The analytical results are important guidelines which will lead to the improvement of the process as well as product development.
REFERENCES
Chandrkrachang, S. and Chinadit, U. 1988. Seaweed production and processing - A new approach. INFOFISH Int. (1988), 4: 22–25.
Chandrkrachang, S. and Chinadit, U. Chandayot, P. and Supasiri, T. 1991. Profitable spin-offs from shrimp seaweed polyculture. INFOFISH Int. (1991), 6: 26–28.
Dockworth, M. and Yaphe, W. 1971. The structure of agar Part I. Fractionation of a complex mixture of polysaccharides. Carbohydrate Research. 16: 189–197.
Singh, T. 1992. Agar and agar production. INFOFISH Technical Handbook 7. 26 pages
Table 3: BRU Analysis of the seaweed and agar samples from participating countries.
| Country | Agar | Seaweed | % Yield | Clarity at 520 mm (%) | Gel strength gm-cm | Sulphate content (%) | Remarks |
| China | |||||||
| - | G. verrucosa | 12.75 | 26 | 233 | 2.89 | ||
| Strip | - | - | 82 | 357 | 1.55 | ||
| India | |||||||
| - | G. edulis | 26.5 | 50 | 245 | 2.57 | IND(MA)06 | |
| MA-01-06 | - | - | 2.0–2.5 | 23–54 | - | IND(MA)01-06 | |
| Indonesia, | |||||||
| West Java | - | Gracilaria sp | 19.75 | 78 | 174 | 2.64 | SW010/IND |
| Kalianda | - | Gracilaria sp | 23.8 | 62 | 99 | 2.36 | SW 001/IND |
| Kalianda | - | Gracilaria sp | 13.8 | 45 | 267 | 0.76 | SW/002/IND |
| Malaysia | |||||||
| Midale | MSW-007 | G.fastigiata | - | 2.0 | 89 | - | |
| Kedah | MSW-008 | G. changii | - | 10.0 | 174 | - | |
| Philippines | Two people trained in BRU for 1 month under FAO support | ||||||
| SW-0050 | - | - | 66.0 | 139 | 0.12 | ||
| SW-0078 | - | - | 87.0 | 214 | 0.23 | ||
| Thailand | |||||||
| Songkhla | No. 5 | G. tenuistipitata | - | 72 | 768 | 1.81 | The extraction of agar and analysis were done at BRU |
| Pattani | No. 4 | G. fisheri | - | 57 | 758 | 2.01 | |
| No. 3 | G.firma | - | 66 | 692 | 2.04 | ||
| Vietnam | |||||||
| Haiphong | - | G. verrucosa | 25 | 60 | 298 | 2.18 | |
| Haiphong | - | G. blodgetti | 21 | 32 | 157 | 2.33 | |
| Haiphong | No. 1 | - | - | 76 | 813 | 0.91 | High quality agar, Haiphong |
Annette Alfsen. Directeur de Recherche,
CNRS, France. 45 rue des Saints Peres. 75006 Paris.
ABSTRACT
The research currently carried out in co-operation with Dr. Suwalee Chandrkrachang from Bangkok (Srinakarinvirot University) deals with the protein content of different samples of Gracilaria spp. growing or cultivated in Thailand. Amino acids are analysed to select the best species for preparation of proteins on an industrial scale from these algae. Such proteins could be used in human and animal food with the same advantages as vegetal proteins, at low cost to countries lacking in animal proteins in Asia. The extraction of proteins has to be done after the extraction of agar, adding value to the production of cultivated Gracilaria spp. Research and extraction of lipid with polyunsaturated fatty acids essential in children's nutrition, are also planned from Gracilaria spp. Further, the preparation of enriched endocytic vesicles, cell organelles previously isolated and studied in other algae, has to be set up from these algae to develop their use in pharmacology particularly with reference to using such materials as drug carriers.
1. INTRODUCTION
The most widely used material obtained from algae such as Gracilaria, is agar. Extraction of good quality agar has been set up in different countries of the region. In the laboratory of Dr Suwalee Chandrkrachang, Biopolymer Research Unit, Sri Nakarinviroj University, the procedure used allows agar to be obtained on an industrial scale with well defined characteristics. Such a product may permit the use of the phycocolloids for a large variety of applications.
1.1 Biotechnology
Recently, algal polysaccharides have found new uses in the field of biotechnology which must be considered as important directions of research in Asian countries because of their high value on the world market. Agars have been used for a long time as solid media for the growth and identification of all types of microorganisms. Agar derived from Gracilaria spp. has functional properties which need to be defined for the different types of uses. One of the new uses is as material for immobilisation of biocatalysts. Immobilisation of an enzyme in its soluble state can be achieved by either cross linking or binding the protein to the polysaccharide. This technique, however, presents some problems and more recently whole cells immobilised on the same material have been used. The procedure should differ from the one used for enzymes. Gel entrapment or micro-encapsulation are the most widely used techniques for immobilising living cells, which can be animal or plant cells. The gel-entrapped cells could then be used as biocatalysts for a lot of commercial processes going from production of ethanol by yeast cells to the production of monoclonal antibodies from hybridoma cells. Since high temperatures have to be avoided for the handling of living cells, the characteristics of each agar have to be well known. Agars with low setting and melting temperatures are the ones to be used.
Techniques to obtain spherical gel beads from agarose/agar to encapsulate living cells have been described, but they have to be studied in each country according to the type of agar obtained from each Gracilaria spp. Immobilisation of living cells in gel beads has found numerous applications due to the easier handling of the beads compared to fragile cells. Plant protoplasts have been entrapped in agarose gels for use in plant cell studies or reproduction, as well as pregerminated seeds or somatic embryos for agricultural uses.
These new uses of agar and agarose in the field of biotechnology make the production, processing and study of agar/agarose from Gracilaria spp. grown and cultivated in south east Asia a process of high economic value. Gracilaria spp, like many other red algae, contain (besides the phycocolloids) proteins which can enter human or animal food.
1.2 Source of protein
A procedure to prepare protein extracts without destroying the colloids has been set up and a study of the content of these proteins in the different amino acids has been undertaken on Gracilaria sp. obtained from Dr Chandrkrachang's lab. Preliminary results suggest that Gracilaria can be an important source of vegetal proteins for the different countries of Asia which lack animal proteins. Like any other protein extracted from vegetal origin they are rich in the essential amino acids and can be obtained on an industrial scale by low cost processing. The proteins are lipo-proteins associated with cell membranes, chlorophyll pigments and soluble proteins from the cytoplasm. The procedure used for extraction of proteins from plant leaves, mainly “luzerne” has to be adapted to the algal extract. Because of the similarity between the luzerne protein extracts and the one obtained from different red algae, namely Gracilaria, the studies recently published by the European Association “Leaf extract and nutrition” can be applied to algae.
Projects to supplement children's food with vegetal proteins in India, Sri Lanka and different countries in Africa have been very successful, with the children showing weight increases and regular growth. The suppression of anaemia and improvements in vitamin A carency and good tolerance are other advantages. Since red algae like Gracilaria are commonly used in the food of the Asian population, the use of algal protein extracts for human food should not bring any problems and may greatly help the nutrition of children in all countries with low resources in animal proteins.
The project for using Gracilaria as a source of vegetal proteins should encompass: research on protein supplementation techniques; the training of local people; and a follow-up of the project among the population receiving supplementation, with a series of control tests.
1.3 Food supplements
Further advantages of algal extracts for food supplementation are their content of different vitamins and minerals. It has also been shown that some lipids with polyunsaturated fatty acyl chains which are essential in child nutrition, are present in algae. Future research on Gracilaria should therefore bring more details on the lipid content.
2. FUTURE RESEARCH
In a more distant future, research on Gracilaria should deal with the use of some cell organelles as drug carriers, according to the results obtained with unicellular algae Dunaliella. This would be a very important field of research and development for the aquaculture and processing of red algae. Pharmacology could benefit from research on the use of algae which are naturally rich in useful molecules and could be the source of cell organelles with low antigenicity . These cell organelles can be enriched in different molecules used as drugs and further used directly as drug carriers since they are natural liposomes with more specificity. Such a field of research requires:
the training and education of personnel, possibly in foreign laboratories;
the establishment of small scale laboratories for biotechnology in different countries;
the regular exchange of information; and
continuous follow up of the progress in each country.
NACA should be the co-ordinator of the research and applications and should be able to control future development, taking into consideration environmental protection.
Mr. Kanit Chaiyakam,
National Institute of Coastal Aquaculture,
Songkhla, Thailand, 90000.
ABSTRACT
Experiments on using seaweed, Gracilaria fisheri for biological wastewater treatment from shrimp pond effluent were conducted by static bioassay with indoor tanks 200 litres in size. The first trial, used 168 ± 2 g of seaweed and the second trial used 340 ± 2 g.
The results of the first experiment (Table 1) showed decreases in the measured parameters, especially after 48 hours had elapsed.
Table 1: Decrease in water quality parameters (%) in tanks with seaweed (Trial 1).
| Hours | BOD | COD | Total Ammonia | Nitrate | Suspended solids | Chlorophyll a |
| 6 | - | 20.4 | 12.5 | 12.5 | 9.1 | - |
| 12 | - | 23.4 | 14.4 | 14.4 | 9.1 | 6.7 |
| 24 | 5.1 | 17.3 | 3.6 | 3.6 | 6.1 | 13.7 |
| 48 | 36.8 | 36.0 | 54.9 | 54.9 | 35.6 | 42.4 |
The second experiment showed more pronounced results (Table 2) due to the higher biomass of seaweed.
Table 1: Decrease in water quality parameters in tanks with seaweed (%).
| Hours | BOD | COD | Total Ammonia | Nitrate | Suspended solids | Chlorophyll a |
| 6 | 41.2 | 25.6 | - | 42.1 | 8.1 | - |
| 12 | 68.7 | 52.1 | - | 100 | 5.1 | - |
| 24 | 39.5 | 55.8 | - | 97.4 | - | - |
| 48 | 73.9 | 20.1 | 66.7 | 100 | - | - |
The utilization of the seaweed Gracilaria for reclamation of shrimp pond effluents requires aeration to encourage water movement and stop sediment attachment to the thallus of seaweed, which otherwise inhibits thallus growth.
Experiments have also been conducted using Gracilaria in combination with green mussel (Mytilus sp.) for biological water treatment. There have also been some field trials in Lake Songkhla for reducing nutrient loads in proximity to finfish cages. The results are very encouraging and confirm the phytosanitation value of Gracilaria.
![]()
![]()
![]()