5.1. Sensory changes
5.2. Autolytic changes
5.3. Bacteriological
changes
5.4. Lipid
oxidation and hydrolysis
Sensory changes are those perceived with the senses, i.e., appearance, odour, texture and taste.
Changes in raw fresh fish
The first sensory changes of fish during storage are concerned with appearance and texture. The characteristic taste of the species is normally developed the first couple of days during storage in ice.
The most dramatic change is onset of rigor mortis. Immediately after death the muscle is totally relaxed and the limp elastic texture usually persists for some hours, whereafter the muscle will contract. When it becomes hard and stiff the whole body becomes inflexible and the fish is in rigor mortis This condition usually lasts for a day or more and then rigor resolves. The resolution of rigor mortis makes the muscle relax again and it becomes limp, but no longer as elastic as before rigor. The rate in onset and resolution of rigor varies from species to species and is affected by temperature, handling, size and physical condition of the fish (Table 5.1).
The effect of temperature on rigor is not uniform. In the case of cod, high temperatures give a fast onset and a very strong rigor mortis. This should be avoided as strong rigor tensions may cause gaping, i.e., weakening of the connective tissue and rupture of the fillet.
It has generally been accepted that the onset and duration of rigor mortis are more rapid at high temperatures, but observations, especially on tropical fish show the opposite effect of temperature with regard to the onset of rigor. It is evident that in these species the onset of rigor is accelerated at 0°C compared to 10°C, which is in good correlation with a stimulation of biochemical changes at 0°C (Poulter et al., 1982; Iwamoto et al., 1987). However, an explanation for this has been suggested by Abe and Okuma (1991) who have shown that onset of rigor mortis in carp (Cyprinus carpio) depends on the difference in sea temperature and storage temperature. When the difference is large the time from death to onset of rigor is short and vice versa.
Rigor mortis starts immediately or shortly after death if the fish is starved and the glycogen reserves are depleted, or if the fish is stressed. The method used for stunning and killing the fish also influences the onset of rigor. Stunning and killing by hypothermia (the fish is killed in iced water) give the fastest onset of rigor, while a blow on the head gives a delay of up to 18 hours (Azam et al., 1990; Proctor et al., 1992).
The technological significance of rigor mortis is of major importance when the fish is filleted before or in rigor. In rigor the fish body will be completely stiff; the filleting yield will be very poor, and rough handling can cause gaping. If the fillets are removed from the bone pre-rigor the muscle can contract freely and the fillets will shorten following the onset of rigor. Dark muscle may shrink up to 52 % and white muscle up to 15 % of the original length (Buttkus, 1963). If the fish is cooked pre-rigor the texture will be very soft and pasty. In contrast, the texture is tough but not dry when the fish is cooked in rigor. Post-rigor the flesh will become firm, succulent and elastic.
Table 5.1 Onset and duration of rigor mortis in various fish species
| Species | Condition | Temperature °C | Time from death to onset of rigor (hours) |
Time from death to end of rigor (hours) |
| Cod (Gadus morhua) | Stressed | 0 |
2-8 |
20-65 |
| Stressed | 10-12 |
1 |
20-30 |
|
| Stressed | 30 |
0.5 |
1-2 |
|
| Unstressed | 0 |
14-15 |
72-96 |
|
| Grouper (Epinephelus malabaricus) | Unstressed | 2 |
2 |
18 |
| Blue Tilapia (Areochromis aureus) | Stressed | 0 |
1 |
|
| Unstressed | 0 |
6 |
||
| Tilapia (Tilapia mossanibica) small 60g | Unstressed | 0-2 |
2-9 |
26.5 |
| Grenadier (Macrourus whitson) | Stressed | 0 |
<1 |
35-55 |
| Anchovy (Engraulis anchoita) | Stressed | 0 |
20-30 |
18 |
| Plaice (Pleuronectes platessa) | Stressed | 0 |
7-11 |
54-55 |
| Coalfish (Pollachius virens) | Stressed | 0 |
18 |
110 |
| Redfish (Sebastes spp.) | Stressed | 0 |
22 |
120 |
| Japanese flounder (Paralichthys olivaceus) | 0 |
3 |
>72 |
|
5 |
12 |
>72 |
||
10 |
6 |
72 |
||
15 |
6 |
48 |
||
20 |
6 |
24 |
||
| Carp (Cyprinus carpio) | 0 |
8 |
||
10 |
60 |
|||
20 |
16 |
|||
| Stressed | 0 |
1 |
||
| Unstressed | 0 |
6 |
SOURCES: Hwang et al., 1991; Iwamoto et al., 1987; Korhonen et al., 1990;Nakayama et al., 1992; Nazir and Magar, 1963; Partmann, 1965; Pawar and Magar, 1965; Stroud, 196; Trucco et al., 1982
Whole fish and fillets frozen pre-rigor can give good products if they are carefully thawed at a low temperature in order to give rigor mortis time to pass while the muscle is still frozen.
The sensory evaluation of raw fish in markets and landing sites is done by assessing the appearance, texture and odour. The sensory attributes for fish are listed in Table 5.2. Most scoring systems are based upon changes taking place during storage in melting ice. It should be remembered that the characteristic changes vary depending on the storage method. The appearance of fish stored under chilled condition without ice does not change as much as for iced fish, but the fish spoil more rapidly and an evaluation of cooked flavour will be necessary. A knowledge of the time /temperature history of the fish should therefore be essential at landing.
The characteristic sensory changes in fish post mortem vary considerably depending on fish species and storage method. A general description has been provided by the EEC in the guidelines for quality assessment of fish as shown in Table 5.2. The suggested scale is numbered from 0 to 3, where 3 is the best quality.
The West European Fish Technologists' Association has compiled a multilingual glossary of odours and flavours which also can be very useful when looking for descriptive words for sensory evaluation of freshness of fish (Howgate et al., 1992 (Appendix C).
Changes in eating quality
If quality criteria of chilled fish during storing are needed, sensory assessment of the cooked fish can be conducted. Some of the attributes for cooked fish and shellfish are mentioned in Table 5.2. A characteristic pattern of the deterioration of fish stored in ice can be found and divided into the following four phases:
Table 5.2 Freshness ratings: Council Regulation (EEC) No. 103/76 OJ No. L20 (28 January 1976) (EEC, 1976)
Criteria |
||||
Marks |
||||
| Part of fish inspected | 3 |
2 |
1 |
0 |
Appearance |
||||
| Skin | Bright, iridescent pigmentation, no
discoloration Aqueous, transparent, mucus |
Pigmentation bright but not lustrous Slightly cloudy mucus |
Pigmentation in the process of becoming
discoloured and dull Milky mucus |
1Dull pigmentation Opaque mucus |
| Eye | Convex (bulging) Transparent cornea Black, bright pupil |
Convex and slightly sunken Slightly opalescent cornea Black, dull pupil |
Flat Opalescent cornea Opaque pupil |
1Concave in the centre Milky cornea Grey pupil |
| Gills | Bright colour No mucus |
Less coloured Slight traces of clear mucus |
Becoming discoloured Opaque mucus |
1Yellowish Milky mucus |
| Flesh (cut from abdomen) | Bluish, translucent, smooth, shining No change in original colour |
Velvety, waxy, dull Colour slightly changed |
Slightly opaque | 1Opaque |
| Colour (along vertebral column) | Uncoloured | Slightly pink | Pink | 1Red |
| Organs | Kidneys and residues of other organs should be bright red, as should the blood inside the aorta | Kidneys and residues of other organs should be dull red; blood becoming discoloured | Kidneys and residues of other organs and blood should be pale red | Kidneys and residues of other organs and should be brownish in colour |
Condition |
||||
| Flesh | Firm and elastic Smooth surface |
Less elastic | Slightly soft (flaccid), less elastic Waxy (velvety) and dull surface |
1Soft (flaccid) Scales easily detached from skin, surface rather wrinkled, inclining to mealy |
| Vertebral column | Breaks instead of coming away | Sticks | Sticks slightly | 1Does not stick |
| Peritoneum | Sticks completely to flesh | Sticks | Sticks slightly | 1Does not stick |
Smell |
||||
| Gills, skin abdominal cavity | Seaweed | No smell of seaweed or any bad smell | Slightly sour | 1Sour |
1 Or in a more advanced state of decay.
A numbered scale may be used for the sensory evaluation of cooked fish as shown in Figure 5.1. The scale is numbered from 0 to 10, 10 indicating absolute freshness, 8 good quality and 6 a neutral tasteless fish. The rejection level is 4. Using the scale in this way the graph becomes S-shaped indicating a fast degradation of the fish during the first phase, a slower rate in phase 2 and 3 and finally a high rate when the fish is spoiled.
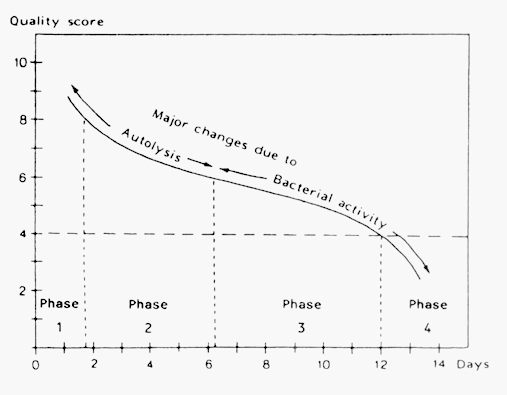
Figure 5.1 Changes in the eating quality of iced (0°C) cod (Huss, 1976)
Other scales can well be used and can change the shape of the graph. It is, however, important to understand the kind of results desired from the sensory analysis in order to ask the right questions to the sensory assessors.
Autolysis means "self-digestion". It has been known for many years that there are at least two types of fish spoilage: bacterial and enzymatic. Uchyama and Ehira (1974) showed that for cod and yellowtail tuna, enzymatic changes related to fish freshness preceded and were unrelated to changes in the microbiological quality. In some species (squid, herring), the enzymatic changes precede and therefore predominate the spoilage of chilled fish. In others, autolysis contributes to varying degrees to the overall quality loss in addition to microbially-mediated processes.
Production of energy in post mortem muscle
At the point of death, the supply of oxygen to the muscle tissue is interrupted because the blood is no longer pumped by the heart and is not circulated through the gills where, in the living fish, it becomes enriched with oxygen. Since no oxygen is available for normal respiration, the production of energy from ingested nutrients is greatly restricted. Figure 5.2 illustrates the normal pathway for the production of muscle energy in most living teleost fish (bony finfish). Glycogen (stored carbohydrate) or fat is oxidized or "burned" by the tissue enzymes in a series of reactions which ultimately produce carbon dioxide (CO2), water and the energy-rich organic compound adenosine triphosphate (ATP). This type of respiration takes place in two stages: an anaerobic and an aerobic stage. The latter depends on the continued presence of oxygen (O2) which is only available from the circulatory system. Most crustaceans are capable of respiring outside the aquatic environment by absorption of atmospheric oxygen for limited periods.
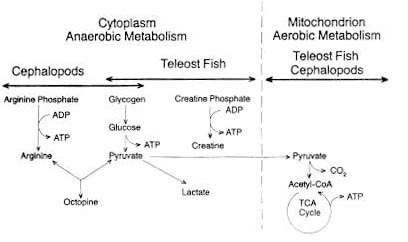
Figure 5.2 Aerobic and anaerobic breakdown of glycogen in fish muscle
Figure 5.2 also illustrates that, under anaerobic conditions, ATP may be synthesized by two other important pathways from creatine phosphate or from arginine phosphate. The former source of energy is restricted to vertebrate muscle (teleost fish) while the latter is characteristic of some invertebrates such as the cephalopods (squid and octopus). In either case, ATP production ceases when the creatine or arginine phosphates are depleted. It is interesting to note that octopine is the end-product from the anaerobic metabolism of cephalopods and is not acidic (unlike lactate), thus any changes in post mortem pH in such animals are not related to the production of lactic acid from glycogen.
For most teleost fish, glycolysis is the only possible pathway for the production of energy once the heart stops beating. This more inefficient process has principally lactic and pyruvic acids as its end-products. In addition, ATP is produced in glycolysis, but only 2 moles for each mole of glucose oxidized as compared to 36 moles ATP produced for each mole of glucose if the glycolytic end products are oxidized aerobically in the mitochondrion in the living animal. Thus, after death, the anaerobic muscle cannot maintain its normal level of ATP, and when the intracellular level declines from 7-10 µmoles/g to £ 1.0 µmoles/g tissue, the muscle enters rigor mortis. Post mortem glycolysis results in the accumulation of lactic acid which in turn lowers the pH of the muscle. In cod, the pH drops from 6.8 to an ultimate pH of 6.1-6.5. In some species of fish, the final pH may be lower: in large mackerel, the ultimate rigor pH may be as low as 5.8-6.0 and as low as 5.4-5.6 in tuna and halibut, however such low pH levels are unusual in marine teleosts. These pHs are seldom as low as those observed for post mortem mammalian muscle. For example, beef muscle often drops to pH levels of 5.1 in rigor mortis. The amount of lactic acid produced is related to the amount of stored carbohydrate (glycogen) in the living tissue. In general, fish muscle contains a relatively low level of glycogen compared to mammals, thus far less lactic acid is generated after death. Also, the nutritional status of the fish and the amount of stress and exercise encountered before death will have a dramatic effect on the levels of stored glycogen and consequently on the ultimate post mortem pH. As a rule, well-rested, well-fed fish contain more glycogen than exhausted fish. In a recent study of Japanese loach (Chiba et al., 1991), it was shown that only minutes of pre-capture stress resulted in a decrease of 0.50 pH units in 3 hours as compared to non-struggling fish whose pH dropped only 0.10 units in the same time period. In addition, the same authors showed that bleeding of fish significantly reduced the post mortem production of lactic acid.
The post mortem reduction in the pH of fish muscle has an effect on the physical properties of the muscle. As the pH drops, the net surface charge on the muscle proteins is reduced, causing them to partially denature and lose some of their water-holding capacity. Muscle tissue in the state of rigor mortis loses its moisture when cooked and is particularly unsuitable for further processing which involves heating, since heat denaturation enhances the water loss. Loss of water has a detrimental effect on the texture of fish muscle and it has been shown by Love (1975) that there is an inverse relationship between muscle toughness and pH, unacceptable levels of toughness (and water-loss on cooking) occurring at lower pH levels (Figure 5.3).
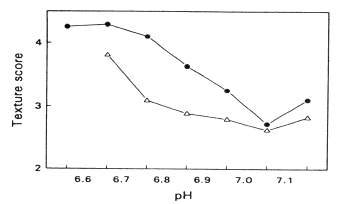
Figure 5.3.Relationship between cod muscle texture and pH, adapted from Love (1975). Black spots refer to fish caught from St. Kilda, Atlantic Ocean, whereas triangles refer to fish caught on Fells Bank, Davis Strait
Autolysis and nucleotide catabolism
As mentioned earlier, rigor mortis sets in when the muscle ATP level drops to £ 1.0 µmoles/g. ATP is not only a source of high energy which is required for muscle contraction in the living animal, but also acts as a muscle plasticizer. Muscle contraction per se is controlled by calcium and an enzyme, ATP-ase which is found in every muscle cell. When intracellular Ca+2 levels are 1 µM, Ca+2 - activated ATP-ase reduces the amount of free muscle ATP which results in the interaction between the major contractile proteins, actin and myosin. This ultimately results in the shortening of the muscle, making it stiff and inextensible. A fish in rigor mortis cannot normally be filleted or processed because the carcass is too stiff to be manipulated and is often contorted, making machine-handling impossible (see also section 3.2 on bleeding and section 5.1 on sensory changes).
The resolution of rigor is a process still not completely understood but always results in the subsequent softening (relaxation) of the muscle tissue and is thought to be related to the activation of one or more of the naturally-occurring muscle enzymes, digesting away certain components of the rigor mortis complex. The softening of the muscle during resolution of rigor (and eventually spoilage processes) is coincidental with the autolytic changes. Among the changes, one of the first to be recognized was the degradation of ATP-related compounds in a more-or-less predictable manner after death. Figure 5.4 illustrates the degradation of ATP to form adenosine diphosphate (ADP), adenosine monophosphate (AMP), inosine monophosphate (IMP), inosine (Ino) and hypoxanthine (Hx). The degradation of ATP catabolites proceeds in the same manner with most fish but the speed of each individual reaction (from one catabolite to another) greatly varies from one species to another and often progresses coincidentally with the perceived level of spoilage as determined by trained analysts. Saito et al. (1959) were the first to observe this pattern and to develop a formula for fish freshness based on these autolytic changes:

where [ATP], [ADP], [AMP], [IMP], [Ino] and [Hx] represent the relative concentrations of these compounds in fish muscle measured at various times during chilled storage.
The K or "freshness" index gives a relative freshness rating based primarily on the autolytic changes which take place during post mortem storage of the muscle. Thus, the higher the K value, the lower the freshness level. Unfortunately, some fish species such as Atlantic cod reach a maximum K value well in advance of the shelf life as determined by trained judges, and K is therefore not considered reliable as a freshness index for all marine finfish. Also, the degradation of nucleotide catabolites is only coincidental with perceived changes in freshness and not necessarily related to the cause of freshness deterioration since only Hx is considered to have a direct effect on the perceived bitter off-flavour of spoiled fish (Hughes and Jones, 1966). It is now widely accepted that IMP is responsible for the desirable fresh fish flavour which is only present in top quality seafood. None of the nucleotide catabolites are considered to be related to the perceived changes in texture during the autolytic process except of course ATP whose loss is associated with rigor mortis.

Figure 5.4 Postmortem ATP degradation in fish muscle. Enzymes include: 1. ATP-ase; 2. myokinase; 3. AMP deaminase; 4.IMP phosphohydrolase; 5a. nucleoside phosphorylase; 5b. inosine nucleosidase; 6,7. xanthine oxidase. Source: Gill (1992)
Surette et al. (1988) followed the autolysis of sterile and non-sterile cod as indicated by the ATP catabolites. The rates of formation and breakdown of IMP were the same in both sterile and non- sterile samples of cod tissue (Figures 5.5a and 5.5b), indicating that the catabolic pathway for the degradation of ATP through to inosine is entirely due to autolytic enzymes.
The conversion of Iino to Hex was accelerated by about 2 days for the non-sterile samples, suggesting that bacterial nucleoside phosphorylase (enzyme 5.a in Figure 5.4) plays a major role in the postmortem production of Hx in refrigerated cod (see also section 5.3). It is interesting to note that Surette et al. (1988) were not able to recover nucleoside phosphorylase from freshly killed cod, but Surette et al. (1990) later went on to isolate and purify this enzyme from a Proteus bacterium recovered from spoiled cod fillets. As mentioned earlier, large variations can be expected in the patterns of nucleotide degradation from one species to another. The variations in Hx among various types of fish are shown in Figure 5.6. It is clear therefore that Hx determination would likely not be useful for such species as swordfish and redfish.
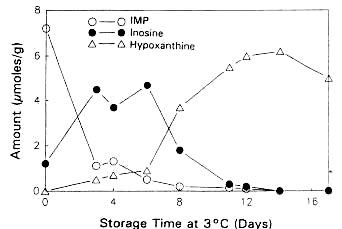
Figure 5.5a Changes in IMP, Ino and Hx in sterile cod fillets at 3°C adapted from Gill (1990)
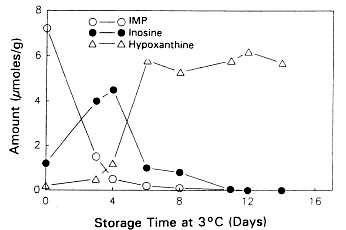
Figure 5.5b Changes in IMP, Ino and Hx in non-sterile cod fillets at 3°C adapted from Gill (1990)
There is little doubt that physical handling accelerates the autolytic changes in chilled fish. Surette et al. (1988) reported that the breakdown rate of the nucleotide catabolites was greater in sterile fillets than in non-sterile gutted whole cod. This is perhaps not surprising since many of the autolytic enzymes have been shown to be compartmentalized in discrete membrane-bound packages which become broken when subjected to physical abuse and result in the intimate mixing of enzyme and substrate. Crushing of the fish by ice or other fish can seriously affect the edibility and filleting yields even for fish which have a relatively low bacterial load, demonstrating the importance of autolytic processes. Iced fish should never be stored in boxes deeper than 30 cm and it is equally important to be sure that fish boxes are not permitted to "nest" one on top of the other if autolysis is to be minimized. Systems for conveying fish and for discharge from the vessels must be designed so as to avoid physical damage to the delicate tissues.
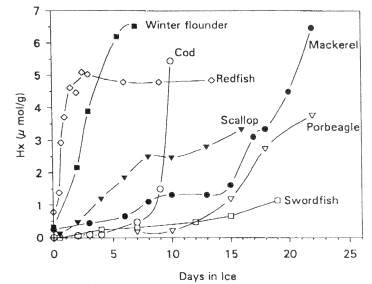
Figure 5.6 Variation in the rate of Hx accumulation of several species during storage in ice. Adapted from Fraser et al. (1967)
Several rapid methods have been developed for the determination of individual nucleotide catabolites or combinations including the freshness index. Two recent reviews should be consulted (Gill, 1990, 1992).
Autolytic changes involving proteolytic enzymes
Many proteases have been isolated from fish muscle and the effects of proteolytic breakdown are often related to extensive softening of the tissue. Perhaps one of the most notable examples of autolytic proteolysis is the incidence of belly-bursting in pelagic (fatty fish) species such as herring and capelin. This type of tissue softening is most predominant in summer months when pelagics are feeding heavily, particularly on "red feed" consisting of copepods and euphausiids. The low molecular weight peptides and free amino-acids produced by the autolysis of proteins not only lower the commercial acceptability of pelagics, but in bulk-stored capelin, autolysis has been shown to accelerate the growth of spoilage bacteria by providing a superior growth environment for such organisms (Aksnes and Brekken, 1988). The induction of bacterial spoilage in capelin by autolysis also resulted in the decarboxylation of amino-acids, producing biogenic amines and lowered the nutritive value of the fish significantly. This is particularly important since autolysis and bacterial growth greatly lower the commercial value of pelagics used for the production of fishmeal.
Similarly, bulk-stored herring used for fishmeal has been found to contain carboxy-peptidases A and B, chymotrypsin, and trypsin; and preliminary studies have shown that proteolysis can be inhibited by the addition of potato extracts which not only slowed the proteolysis but resulted in lower microbial growth and preservation of the nutritional value of the meal (Aksnes, 1989).
More recently, Botta et al. (1992) found that autolysis of the visceral cavity (belly-bursting) of herring was related more to physical handling practices than to biological factors such as fish size, amount of red feed in the gut or roe content. In particular, it was shown that for herring, freezing/thawing, thawing time at 15°C and time of iced storage, had a far greater influence on belly- bursting than biological factors.
Cathepsins
Although several proteolytic enzymes have been discovered in the fish tissues, it has perhaps been the cathepsins which have been described most often. The cathepsins are "acid" proteases usually found packaged in tiny, submicroscopic organelles called lysozomes. In living tissue, lysozomal proteases are believed to be responsible for protein breakdown at sites of injury. Thus cathepsins are for the most part inactive in living tissue but become released into the cell juices upon physical abuse or upon freezing and thawing of post mortem muscle.
Cathepsins D and L are believed to play a major role in the autolytic degradation of fish tissue since most of the other cathepsins have a relatively narrow pH range of activity far too low to be of physiological significance. Reddi et al. (1972) demonstrated that an enzyme believed to be cathepsin D from winter flounder was active over a pH range of 3-8 with a maximum near pH 4.0, although no attempt was made to confirm the identity of the enzyme using synthetic substrates or specific inhibitors. Nevertheless, the enzyme was far less active in the presence of ATP, suggesting that such an enzyme would only be active in post mortem fish muscle. Also, the enzyme activity was inhibited strongly by the presence of salt (Figure 5.7) with virtually no activity remaining after a 25-hour incubation in the presence of 5% sodium chloride. It is therefore unlikely that Reddi's enzyme was active in salted fish products.
Cathepsin L has been implicated in the softening of salmon muscle during spawning migration. It is likely that this enzyme contributes more to autolysis of fish muscle than cathepsin D since it is far more active at neutral pH, and has been shown to digest both myofibrillar proteins (actomyosin) as well as connective tissue. Yamashita and Konogaya (1990) produced strong evidence implicating cathepsin L rather than other cathepsins in the softening of salmon during spawning. They demonstrated that electrophoresis of purified myofibrils treated with cathepsin L resulted in patterns which were almost identical to patterns of proteins recovered from muscle from spawning fish. Furthermore, the cathepsin L autolytic activity correlated well with the texture of the muscle as measured instrumentally. The linear correlation between cathepsin L activity and breaking strength of the muscle was excellent; r = 0.86 and -0.95 for fresh and frozen/thawed tissue, respectively. It is interesting that, in all cases, the autolytic ability as measured by cathepsin L activity was higher in frozen/thawed tissue than in fresh tissue. Freezing and thawing often break down cell membranes allowing autolytic membrane-bound enzymes to react with their natural substrates. The enzyme and its naturally occurring inhibitor were further studied by the same authors (Yamashita and Konogaya, 1992). Cathepsin L has also been associated with the production of a jelly- like softening of flounder (Toyohara et l., 1993 a) and the uncontrollable softening of Pacific hake muscle which has been parasitized by Myxosporidia (Toyohara et al., 1993 b).
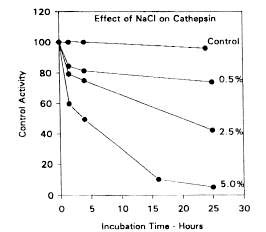
Figure 5.7 Effect of NaCl on the catheptic activity. Adapted from Reddi et al. (1972)
The tissues of such infected fish have little commercial value, but at present it is not known if it is the parasite or the host which secretes the proteolytic enzymes which autolyze the muscle.
In addition to their detrimental effect on texture, catheptic enzymes induce intentional autolytic changes in fermented fish products. For example, cathepsins are believed to be responsible for major textural changes during the fermentation of salted preserved Japanese squid and Crucian carp (Makinodan et al., 1991, 1993).
Calpains
A second group of intracellular proteases called "calpains" or "calcium activated factor" (CAF) has recently been associated with fish muscle autolysis and is found in meats, finfish and crustaceans. Tenderness is probably the most important quality characteristic of red meat. It has been known for nearly a century that post mortem aging of red meat results in the tenderization process. Calpains have been found primarily responsible for the post mortem autolysis of meat through digestion of the z- line proteins of the myofibril. Although toughness is seldom a problem with unfrozen fish muscle, softening through autolysis is a serious problem limiting the commercial value. The calpains are intra-cellular endopeptidases requiring cysteine and calcium; µ-calpain requiring 5-50 µM Ca2+ , m-calpain requiring 150-1000 µM Ca2+ . Most calpains are active at physiological pH, making it reasonable to suspect their importance in fish-softening during chilled storage.
Studies have shown that in crustacean muscle, calpains are associated with moltinduced textural changes to the muscle and carry out non-specific generalized digestion of the myofibrillar proteins. However, vertebrate muscle calpains have been shown to be very specific, digesting primarily tropinin- T, desmin, titin and nebulin, attacking neither vertebrate actin or myosin (Koohmaraie, 1992). In contrast, fish calpains digest myosin (specifically the myosin heavy chain) to form an initial fragment with approximate molecular weight of 150 000 Da (Muramoto et al., 1989). The same authors demonstrated that fish calpains were far more active at low temperatures than were mammalian calpains and that the rates of cleavage were species-specific, being most active against myosins with lowest heat stabilities. Thus, fish species adapted to colder environmental temperatures are more susceptible to calpain autolysis than those from tropical waters. Although calpain has been identified in several fish species including carp (Toyohara et al., 1985), tilapia and shrimp (Wang et al., 1993), as well as tuna, croaker, red seabream and trout (Muramoto et al., 1989) to name a few, little work has to date demonstrated a "cause and effect" relationship between calpain activity and instrumental measurements of texture.
Collagenases
To this point, all of the post mortem autolytic changes described have involved changes within the muscle cell per se. However, the flesh of teleost fish is divided into blocks of muscle cells separated into "flakes" or myotomes by connective tissue called myocommata (Figure 3.3). Each muscle cell or fibre is surrounded with connective tissue which attaches to the myocommata at the ends of the cells by means of fine collagenous fibrils. During chilled storage, these fibrils deteriorate (Bremner and Hallett, 1985). More recently, it was shown that instrumental measurements of texture of chilled trout muscle decreased as the amount of type V collagen was solubilized, presumably due to the action of autolytic collagenase enzymes (Sato et al., 1991). It is these enzymes which presumably cause "gaping" or breakdown of the myotome during long-term storage on ice or short term storage at high temperature. For Atlantic cod, it has been shown that upon reaching 17°C, gaping is inevitable presumably because of degradation of the connective tissue and rapid shortening of the muscle due to high temperature rigor.
The relatively short shelf life of chilled prawns due to softening of the tissue has also been shown to be due to the presence of collagenase enzymes (Nip et al., 1985). The source of the collagenase enzymes in prawn is thought to be the hepatopancreas (digestive organ).
Autolytic changes during frozen storage
The reduction of trimethylamine oxide (TMAO), an osmoregulatory compound in many marine teleost fish, is usually due to bacterial action (section 5.3) but in some species an enzyme is present in the muscle tissue which is able to break down TMAO into dimethylamine (DMA) and formaldehyde (FA):
(CH3)3 NO
(CH3)2NH + HCHO
It is important to note that the amount of formaldehyde produced is equivalent to the dimethylamine formed but is of far greater commercial significance. Formaldehyde induces cross- linking of the muscle proteins making the muscle tough and readily lose its water holding capacity. The enzyme responsible for formal dehyde-induced toughening is called TMAO-ase or TMAO demethylase and is most commonly found in the gadoid fishes (cod family). Most of the TMAO demethylase enzymes reported to date weremembrane-bound and become most active when the tissue membranes are disrupted by freezing or artificially by detergent solubilization. Dark (red) muscle has a higher rate of activity than white muscle whereas other tissues such as kidney, spleen and gall bladder are extremely rich in the enzyme. Thus, it is important that minced fish is completely free of organ tissue such as kidney from gadoid species if toughening in frozen storage is to be avoided. It is often difficult to ensure that the kidney is removed prior to mechanical deboning since this particular organ runs the full length of the backbone and is adherent to it. The TMAO-ase enzyme has been isolated from the microsomal fraction in hake muscle (Parkin and Hultin, 1986) and the lysosomal membrane in kidney tissue (Gill et al., 1992). It has been shown that the toughening of frozen hake muscle is correlated to the amount of formaldehyde produced, and that the rate of FA production is greatest at high frozen-storage temperatures (Gill et al., 1979). In addition, it has been shown that the amount of FA-induced toughening is enhanced by physical abuse to the catch prior to freezing and by temperature fluctuations during frozen storage. The most practical means of preventing the autolytic production of FA is to store fish at temperatures < -30°C to minimize temperature fluctuations in the cold store and to avoid rough handling or the application of physical pressure on the fish prior to freezing. The autolytic changes affecting the edibility of fresh and frozen fish are summarized in Table 5.3. Generally, the most important single factor affecting autolysis is physical disruption of the muscle cells. No attempt has been made here to deal with the alkaline proteases associated with the softening of cooked surimi products. An article by Kinoshita et al. (1990) deals with the heat-activated alkaline proteases associated with the softening in surimi-based products.
Table 5.3 Summary of Autolytic Changes in Chilled Fish
| Enzyme(s) | Substrate | Changes Encountered | Prevention/Inhibition |
| glycolytic enzymes | glycogen | production of lactic acid, pH of tissue drops, loss
of water-holding capacity in muscle high temperature rigor may result in gaping |
fish should be allowed to pass through rigor at
temperatures as close to 0°C as practically possible pre-rigor stress must be avoided |
| autolytic enzymes, involved in nucleotide breakdown | ATP ADP AMP IMP |
loss of fresh fish flavour, gradual production of bitternes with Hx (later stages) | same as above rough handling or crushing accelerates breakdown |
| cathepsins | proteins, peptides | softening of tissue making processing difficult or impossible | rough handling during storage and discharge |
| chymotrypsin, trypsin, carboxy-peptidases | proteins, peptides | autolysis of visceral cavity in pelagics (belly- bursting) | problem increased with freezing/thawing or long- term chill storage |
| calpain | myofibrillar proteins | softening, molt-induced softening in crustaceans | removal of calcium thus preventing activation? |
| collagenases | connective tissue | gaping" of fillets softening |
connective tissue degradation related to time and temperature of chilled storage |
| TMAO demethylase | TMAO | formaldehyde-induced toughening of frozen gadoid fish | store fish at temperature <= -30°C physical abuse and freezing/thawing accelerate formaldehyde-induced toughening |
The bacterial flora on live fish
Microorganisms are found on all the outer surfaces (skin and gills) and in the intestines of live and newly caught fish. The total number of organisms vary enormously and Liston (1980) states a normal range of 102-107 cfu (colony forming units)/cm2 on the skin surface. The gills and the intestines both contain between 103 and 109 cfu/g (Shewan, 1962).
The bacterial flora on newly-caught fish depends on the environment in which it is caught rather than on the fish species (Shewan, 1977). Fish caught in very cold, clean waters carry the lower numbers whereas fish caught in warm waters have slightly higher counts. Very high numbers, i.e., 107 cfu/cm2 are found on fish from polluted warm waters. Many different bacterial species can be found on the fish surfaces. The bacteria on temperate water fish are all classified according to their growth temperature range as either psychrotrophs or psychrophiles. Psychrotrophs (cold-tolerant) are bacteria capable of growth at 0°C but with optimum around 25°C. Psychrophiles (cold-loving) are bacteria with maximum growth temperature around 20°C and optimum temperature at 15°C (Morita, 1975). In warmer waters, higher numbers of mesophiles can be isolated. The microflora on temperate water fish is dominated by psychrotrophic Gram-negative rodshaped bacteria belonging to the genera Pseudomonas, Moraxella, Acinetobacter, Shewanella and Flavobacterium. Members of the Vibrionaceae (Vibrio and Photobacterium) and the Aeromonadaceae (Aeromonas spp.) are also common aquatic bacteria and typical of the fish flora (Table 5.4). Gram-positive organisms as Bacillus, Micrococcus, Clostridium, Lactobacillus and coryneforms can also be found in varying proportions, but in general, Gram-negative bacteria dominate the microflora. Shewan (1977) concluded that Gram-positive Bacillus and Micrococcus dominate on fish from tropical waters. However, this conclusion has later been challenged by several studies which have found that the microflora on tropical fish species is very similar to the flora on temperate species (Acuff et al.,1984; Gram et al., 1990; Lima dos Santos 1978; Surendran et al., 1989). A microflora consisting of Pseudomonas, Acinetobacter, Moraxella and Vibrio has been found on newly-caught fish in several Indian studies (Surendran et al., 1989). Several authors conclude, as Liston (1980), that the microflora on tropical fish often carry a slightly higher load of Gram-positives and enteric bacteria but otherwise is similar to the flora on temperate-water fish.
Aeromonas spp. are typical of freshwater fish, whereas a number of bacteria require sodium for growth and are thus typical of marine waters. These include Vibrio, Photobacterium and Shewanella. However, although Shewanella putrefaciens is characterized as sodium-requiring, strains of S. putrefaciens can also be isolated from freshwater environments (DiChristina and DeLong, 1993; Gram et al., 1990; Spanggaard et al., 1993). Although S. putrefaciens has been isolated from tropical freshwaters, it is not important in the spoilage of freshwater fish (Lima dos Santos, 1978; Gram, 1990).
Table 5.4 Bacterial flora on fish caught in clean, unpolluted waters
| Gram-negative | Gram-positive | Comments |
| Pseudomonas | Bacillus | |
| Moraxella | Clostridium | |
| Acinetobacter | Micrococcus | |
| Shewanella putrefaciens | Lactobacillus | |
| Flavobacterium | Coryneforms | |
| Cytophaga | ||
| Vibrio Photobacterium Aeromonas |
Vibrio and Photobacterium are typical of marine waters; Aeromonas is typical of freshwater |
In polluted waters, high numbers of Enterobacteriaceae may be found. In clean temperate waters, these organisms disappear rapidly, but it has been shown that Escherichia coli and Salmonella can survive for very long periods in tropical waters and once introduced may almost become indigenous to the environment (Fujioka et al., 1988).
The taxonomy of S. putrefaciens has been rather confused. The organism was originally associated with the Achromobacter group but was later placed in the Shewan Pseudomonas group IV. Based on percentage of guanine+ cytosine (GC%) it was transferred to the genus Alteromonas, but on the basis of 5SRNA homology it was reclassified to a new genus, Shewanella (MacDonnell and Colwell, 1985). It has recently been suggested that the genus Aeromonas spp. which was a member of the Vibrionaceae family be transferred to its own family, the Aeromonadaceae (Colwell et al., 1986).
Japanese studies have shown very high numbers of microorganisms in the gastrointestinal tract of fish, and as such numbers are much higher than in the surrounding water, this indicates the presence of a favourable ecological niche for the microorganisms. Similarly, Larsen et al. (1978) reported up to 107 cfu/g of vibrio-like organisms in the intestinal tract of cod and Westerdahl et al. (1991) also isolated high numbers of vibrio-like organisms from the intestines of turbot. Photobacterium phosphoreum which can be isolated from the surface can also be isolated in high numbers from the intestinal tract of some fish species (Dalgaard, 1993). On the contrary, some authors believe that the microflora of the gastrointestinal tract is merely a reflection of the environment and the food intake.
Microbial invasion
The flesh of healthy live or newly-caught fish is sterile as the immune system of the fish prevents the bacteria from growing in the flesh (Figure 5.8 a). When the fish dies, the immune system collapses and bacteria are allowed to proliferate freely. On the skin surface, the bacteria to a large extent colonize the scale pockets. During storage, they invade the flesh by moving between the muscle fibres. Murray and Shewan (1979) found that only a very limited number of bacteria invaded the flesh during iced storage. Ruskol and Bendsen (1992) showed that bacteria can be detected by microscope in the flesh when the number of organisms on the skin surface increases above 106 cfu/cm2 (Figure 5.6 b). This was seen at both iced and ambient temperatures. No difference was found in the invasive patterns of specific spoilage bacteria (e.g., S. putrefaciens) and non-spoilage bacteria.
Since only a limited number of organisms actually invade the flesh and microbial growth mainly takes place at the surface, spoilage is probably to a large extent a consequence of bacterial enzymes diffusing into the flesh and nutrients diffusing to the outside.
Fish spoil at very different rates (see also section 6.5), and differences in surface properties of fish have been proposed to explain this. Skins of fish have very different textures. Thus whiting (Merlangius merlangus) and cod (Gadus morhua) which have a very fragile integument spoil rapidly compared to several flatfish such as plaice that has a very robust dermis and epidermis. Furthermore, the latter group has a very thick slime layer, which includes several antibacterial components, such as antibodies, complement and bacteriolytic enzymes (Murray and Fletcher, 1976; Hjelmland et al., 1983).
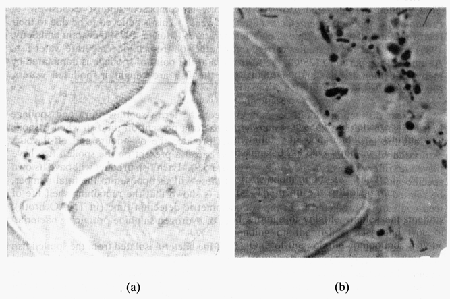
Figure 5.8 Histological section of (a) newly-caught cod and (b) cod fillets stored 12 days in ice. The section has been Giemsa-stained (Ruskol and Bendsen, 1992)
Changes in the microflora during storage and spoilage/Specific spoilage organisms
Bacteria on fish caught in temperate waters will enter the exponential growth phase almost immediately after the fish have died. This is also true when the fish are iced, probably because the microflora is already adapted to the chill temperatures. During ice storage, the bacteria will grow with a doubling time of approximately 1 day and will, after 2-3 weeks, reach numbers of 108-109 cfu/g flesh or cm2 skin. During ambient storage, a slightly lower level of 107-108 cfu/g is reached in 24 hours. The bacteria on fish caught in tropical waters will often pass through a lag-phase of 1-2 weeks if the fish are stored in ice, whereafter exponential growth begins. At spoilage, the bacterial level on tropical fish is similar to the levels found on temperate fish species (Gram, 1990; Gram et al., 1990).
If iced fish are stored under anaerobic conditions or if stored in CO2 containing atmosphere, the number of the normal psychrotrophic bacteria such as S. putrefaciens and Pseudomonas is often much lower, i.e., 106-107 cfu/g than on the aerobically stored fish. However, the level of bacteria of psychrophilic character such as P. phosphoreum reaches a level of 107-108 cfu/g when the fish spoil (Dalgaard et al., 1993).
The composition of the microflora also changes quite dramatically during storage. Thus, under aerobic iced storage, the flora is composed almost exclusively of Pseudomonas spp. and S. putrefaciens after 1-2 weeks. This is believed to be due to their relatively short generation time at chill temperatures (Morita, 1975; Devaraju and Setty, 1985) and is true for all studies carried out whether on tropical or temperate-water fish. At ambient temperature (25°C), the microflora at the point of spoilage is dominated by mesophilic Vibrionaceae and, particularly if the fish are caught in polluted waters, Enterobacteriaceae.
A clear distinction should be made between the terms spoilage flora and spoilage bacteria since the first describes merely the bacteria present on the fish when it spoils whereas the latter is the specific group that produce the off-odours and off-flavours associated with spoilage. A large part of the bacteria present on the spoiled fish have played no role whatever in the spoilage (Figure 5.9). Each fish product will have its own specific spoilage bacteria and the number of these will, as opposed to the total number, be related to the shelf life. In Figure 5.10, it is shown that the remaining shelf life of iced cod can be predicted from the conductometric detection time (in TMAO broth), which is inversely correlated with the number of hydrogen sulphide-producing bacteria.
It is not an easy task to determine which of the bacteria isolated from the spoiled fish are those causing spoilage, and it requires extensive sensory, microbiological and chemical studies. First, the sensory, microbiological and chemical changes during storage must be studied and quantified, including a determination of the level of a given chemical compound that correlates with spoilage (the chemical spoilage indicator). Second, bacteria are isolated at the point of sensory rejection. Pure and mixed cultures of bacteria are screened in sterile fish substrates for their spoilage potential, i.e., their ability to produce sensory (off-odours) and chemical changes typical of the spoiling product. Finally, the selected strains are tested to evaluate their spoilage activity, i.e., if their growth rate and their qualitative and quantitative production of off-odours are similar to the measurements in the spoiled product (Dalgaard, 1993).
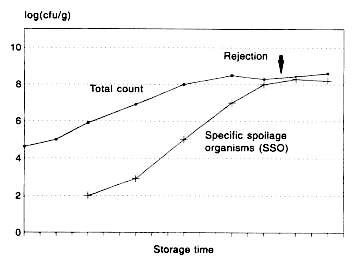
Figure 5.9 Changes in total counts and specific spoilage bacteria during storage (modified after Dalgaard (1993)

Figure 5.10 Comparison of remaining shelf life of iced cod and detection time in a TMAO broth (Jorgensen et al., 1988)
The latter step is particularly important, as some bacteria may produce the chemical compounds associated with spoilage but are unable to do so in significant amounts, and they are thus not the specific spoilage bacteria. When stored aerobically, levels of 108-109 cfu/g of specific spoilage bacteria are required to cause spoilage. The spoilage of packed fish is seen at a much lower level of 107 cfu P. phosphoreum per gramme. This relatively low level is probably due to the very large size (5 µm) of the bacterium resulting in a much higher yield of for example, TMA per cell (Dalgaard, 1993).
Spoilage potential and activity can be assessed in several fish substrates as sterile, raw fish juice (Lerke et al., 1963), heat-sterilized fish juice (Castell and Greenough, 1957; Gram et al. , 1987; Dalgaard, 1993) or on sterile muscle blocks (Herbert et al., 1971). The latter is the most complicated but is also that yielding results comparable to the product. If any of the fish juices are chosen, it is important that the growth rate of the spoilage bacteria in the model system is equal to the growth rate in the product.
A qualitative test for the ability of the bacteria to produce H2S and/or reduce TMAO may also be used when the spoilage flora is screened for potential spoilage bacteria. A medium where the reduction of TMAO to TMA is seen as a redox indicator changes colour, and the formation of H2S is evident from a black precipitation of FeS which has been developed for this purpose (Gram et al.,1987).
Shewanella putrefaciens has been identified as the specific spoilage bacteria of marine temperate- water fish stored aerobically in ice. If the product is vacuum-packed, P. phosphoreum participates in the spoilage and it becomes the specific spoilage bacteria of CO2 packed fish (see section 6.3). The spoilage flora on iced tropical fish from marine waters is composed almost exclusively of Pseudomonas spp. and S. putrefaciens. Some Pseudomonas spp. are the specific spoilers of iced stored tropical freshwater fish (Lima dos Santos, 1978; Gram et al., 1990) and are also, together with S. putrefaciens, spoilers of marine tropical fish stored in ice (Gillespie and MacRae, 1975; Gram, 1990).
At ambient temperature, motile aeromonads are the specific spoilers of aerobically stored freshwater fish (Gorzyka and Pek Poh Len, 1985; Gram et al., 1990). Barile et al. (1985) showed that a large proportion of the flora on ambient-stored mackerel consisted of S. putrefaciens, indicating that this bacterium may also take part in the spoilage.
Table 5.5 gives an overview of the specific spoilage bacteria of fresh fish products stored in ice and at ambient temperature.
Table 5.5 Dominating microflora and specific spoilage bacteria at spoilage of fresh, white fish (cod) (from Huss, 1994)
| Storage temperature | Packaging atmosphere | Dominating microflora | Specific spoilage organisms (SSO) | References |
| 0°C | Aerobic | Gram-negative psychrotrophic, non-fermentative rods (Pseudomonas spp., S. putrefaciens, Moraxella, Acinetobacter) | S. putrefaciens Pseudomonas 3 | 2,3,4,9 |
| 0°C | Vacuum | Gram-negative rods; psychrotrophic or with psychrophilic character (S. putrefaciens, Photobacterium) | S. putrefaciens P. phosphoreum | 1,9 |
| 0°C | MAP1 | Gram-negative fermentative rods with psychrophilic
character (Photobacterium) Gram-negative non-fermentative psychrotrophic rods (1-10% of flora; Pseudomonas, S. putrefaciens) Gram-positive rods (LAB 2) |
P. phosphoreum | 1,7 |
| 5°C | Aerobic | Gram-negative psychrotrophic rods (Vibrionaceae, S. putrefaciens) | Aeromonas spp. S. putrefaciens |
10 |
| 5°C | Vacuum | Gram-negative psychrotrophic rods (Vibrionaceae, S. putrefaciens) | Aeromonas spp. S. putrefaciens |
10 |
| 5°C | MAP | Gram-negative psychrotrophic rods (Vibrionaceae) | Aeromonas spp. | 6 |
| 20-30°C | Aerobic | Gram-negative mesophilic fermentative rods (Vibrionaceae, Enterobacteriaceae) | Motile Aeromonas spp. (A. hydrophila) |
2,4,5,8 |
1) Modified Atmosphere Packaging (CO2 containing)
2) LAB: Lactic Acid Bacteria
3) Fish caught in tropical waters or freshwaters tend to have a
spoilage dominated by Pseudomonas spp.
References: 1) Battle et al. (1985); 2) Dalgaard et al. (1993); 3) Donald and Gibson (1992); 4) Gorczyca and Pek Poh Len (1985); 5) Gram et al. (1987); 6) Gram et al. (1990); 7) Gram and Dalgaard (pers. comm.); 8) Jorgensen and Huss (1989); 9) Lima dos Santos (1978); 10) van Spreekens (1977)
Biochemical changes induced by bacterial growth during storage and spoilage
Comparison of the chemical compounds developing in naturally spoiling fish and sterile fish has shown that most of the volatile compounds are produced by bacteria (Shewan, 1962) as shown in Figure 5.11. These include trimethylamine, volatile sulphur compounds, aldehydes, ketones, esters, hypoxanthine as well as other low molecularweight compounds.
The substrates for the production of volatiles
are the carbohydrates (e.g., lactate and ribose), nucleotides (e.g.,
inosine mono-phosphate and inosine) and other NPN molecules. The
amino-acids are particularly important substrates for formation
of sulphides and ammonia. 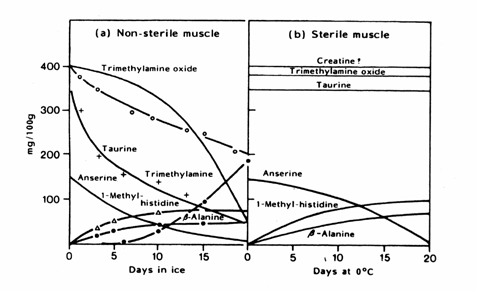
Figure 5.11 Changes in the nitrogenous extractives in a) spoiling and b) autolysing cod muscle (Shewan, 1962)
Microorganisms obtain far more energy from aerobic oxidation than from an anaerobic fermentation; thus the complete oxidation of 1 mole glucose (or other hexose) via Kreb's cycles yields 6 moles of CO2 and 36 moles of ATP. On the contrary, the fermentation of 1 mole glucose gives only 2 moles of ATP and two moles of lactic acid. The initial aerobic growth on fish is dominated by bacteria using carbohydrates as substrate and oxygen as terminal electron-acceptor with the concurrent production of CO2 and H2O.
Reduction of Trimethylarnine Oxide (TMAO)
The growth of oxygen-consuming bacteria results in the formation of anaerobic or microaerophilic niches on the fish. This does, however, not necessarily favour the growth of anaerobic bacteria. Some of the bacteria present on fish are able to carry out a respiration (with the ATP advantage) by using other molecules as electron acceptor. It is typical of many of the specific spoilage bacteria on fish that they can use TMAO as electron acceptorin an anaerobic respiration. The reduced component, TMA, which is one of the dominant components of spoiling fish, has a typical fishy odour. The level of TMA found in fresh fish rejected by sensory panels varies between fish species, but is typically around 10-15 mg TMA-N/100 g in aerobically stored fish and at a level of 30 mg TMA-N/100 g in packed cod (Dalgaard et al., 1993).
The TMAO reduction is mainly associated with the genera of bacteria typical of the marine environment (Alteromonas, Photobactetium, Vibrio and S. putrefaciens), but is also carried out by Aeromonas and intestinal bacteria of the Enterobacteriaceae. TMAO reduction has been studied in fermentative, facultative anaerobic bacteria like E. coli (Sakaguchi et al., 1980) and Proteus spp. (Stenberg et al., 1982) as well as in the non-fermentative S. putrefaciens (Easter et al, 1983; Ringo et al, 1984). During aerobic growth, S. putrefaciens uses the Kreb's cycle to produce the electrons that are later channelled through the respiratory chain. Ringo et al. (1984) suggested that during anaerobic respiration S. putrefaciens also uses the complete Kreb's cycle (Figure 5.12), whereas it has recently been shown that in the anaerobic respiration in S. putrefaciens, only part of the Kreb's cycle is used (Figure 5.13) and electrons are also generated by another metabolic pathway, namely the serine pathway (Scott and Nealson, 1994). S. putrefaciens can use a variety of carbon sources as substrate in its TMAO-dependent anaerobic respiration, including formate and lactate. Compounds like acetate and succinate that are used in the oxygen respiration cannot be used when TMAO is terminal electron acceptor (DiChristina and DeLong, 1994) and on the contrary, acetate is a product of the anaerobic TMAO reduction (Ringo et al., 1984; Scott and Nealson, 1994).
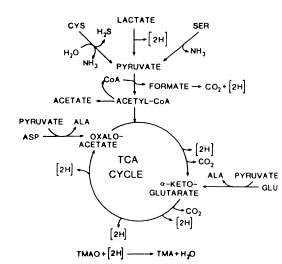
Figure 5.12 Anaerobic reduction of TMAO by Shewanella putrefaciens (formerly Alteromonas) as suggested by Ringo et al. (1984)
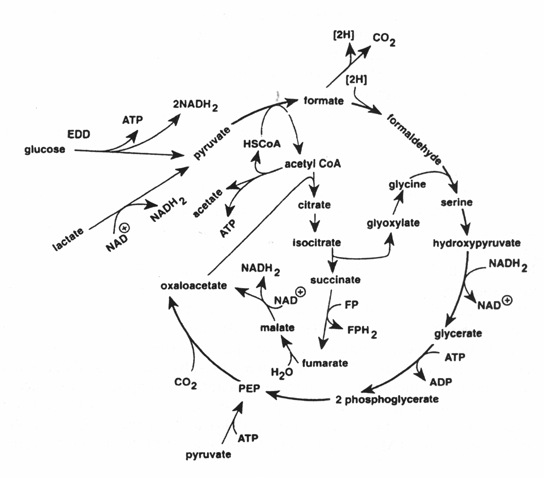
Figure 5.13 Proposed route of carbon during anaerobiosis for S. putrefaciens (Scott and Nealson, 1994)
Contrary to this, sugars and lactate are the main substrates generating electrons when Proteus spp. reduces TMAO. The reduction is accompanied by a production of acetate as the main product (Kjosbakken and Larsen, 1974).
TMAO is, as mentioned in section 4.4, a typical component of marine fish, and it has recently been reported that also some tropical freshwater fish contain high amounts of TMAO (Anthoni et al., 1990). However, TMA is not necessarily a characteristic component during spoilage of such fish because spoilage is due to Pseudomonas spp. (Gram et al., 1990).
The development of TMA is in many fish species paralleled by a production of hypoxanthine. Hypoxanthine can, as described in section 5.2. be formed by the autolytic decomposition of nucleotides, but it can also be formed by bacteria; and the rate of bacterial formation is higher than the autolytic. Both Jorgensen et al. (1988) and Dalgaard (1993) showed a linear correlation between the contents of TMA and hypoxanthine during iced storage of packed cod (Figure 5.14). Several of the spoilage bacteria produce hypoxanthine from inosine or inosine mono-phosphate, including Pseudomonas spp. (Surette et al., 1988) S. putrefaciens (van Spreekens, 1977; Jorgensen and Huss, 1989; Gram, 1989) and P. phosphoreum (van Spreekens, 1977).
In cod and other gadoid fishes, TMA constitutes most of the so-called total volatile bases, TVB (also called total volatile nitrogen, TVN) until spoilage. However, in the spoiled fish where the TMAO supplies are depleted and TMA has reached its maximum level, TVB levels still rise due to formation of NH3 and other volatile amines. A little ammonia is also formed in the first weeks of iced storage due to autolysis. In some fish that do not contain TMAO or where spoilage is due to a non-TMAO reducing flora, a slow rise in TVB is seen during storage, probably resulting from the deamination of amino-acids.
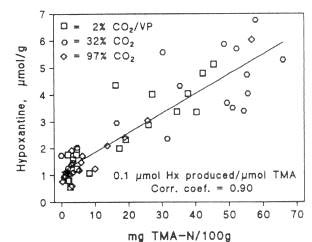
Figure 5.14 Relationship between contents of TMA and Hx during storage of packed cod in ice (Dalgaard et al., 1993)
Volatile sulphur-compounds are typical components of spoiling fish and most bacteria identified as specific spoilage bacteria produce one or several volatile sulphides. S. putrefaciens and some Vibrionaceae produce H2S from the sulphur containing amino-acid 1-cysteine (Stenstroem and Molin, 1990; Gram et al., 1987). On the contrary, neither Pseudomonas nor P. phosphoreum produce significant amounts of H2S. Thus, hydrogen sulphide, which is typical of spoiling iced cod stored aerobically, is not produced in spoiling CO2 packed fish (Dalgaard et al., 1993). Methylmercaptan (CH3SH) and dimethylsulphide ((CH3)2S) are both formed from the other sulphur-containing amino-acid, methionine. Taurine, which is also sulphur-containing, occurs as free amino-acid in very high concentrations in fish muscle. It disappears from the fish flesh during storage (Figure 5.11) but this is because of leakage rather than because of bacterial attack (Herbert and Shewan, 1975). The formation of compounds in naturally-spoiling cod as compared to sterile muscle is shown in Figure 5.15.
The volatile sulphur-compounds are very foul-smelling and can be detected even at ppb levels, so even minimal quantities have a considerable effect on quality. Ringo et al. (1984) have shown that cysteine is used as substrate in the Kreb's cycle when electrons are transferred to TMAO, and the formation of H2S and TMA is thus to some extent a linked reaction (Figure 5.12).
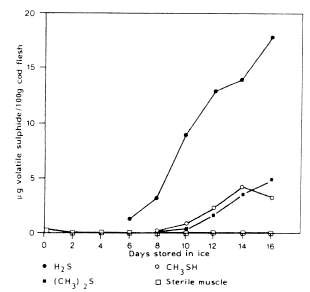
Figure 5.15 Production of HA CH3SH and (CH3)2S in naturally spoiling cod fillets and sterile muscle blocks (Herbert and Shewan, 1976)
Contrary to the iced spoilage by S. putrefaciens and the ambient spoilage by Vibrionaceae which is dominated by H2S and TMA, the spoilage caused by Pseudomonas spp. is characterized by absence of these compounds (Gram et al., 1989, Gram et al., 1990). Fruity, rotten, sulphydryl odours and flavours are typical of the Pseudomonas spoilage of iced fish. Pseudomonas spp. produce a number of volatile aldehydes, ketones, esters and sulphides (Edwards et al., 1987; Miller et al., 1973 a, 1973 b). However, it is not known which specific compounds are responsible for the typical off-odours (Table 5.6). The fruity off-odours produced by Pseudomonas fragi originate from monoaminomonocarboxylic amino-acids.
Table 5.6 Typical spoilage compounds during spoilage of fresh fish stored aerobically or packed in ice or at ambient temperature
| Specific spoilage organism | Typical spoilage compounds |
| Shewanella putrefaciens | TMA, H2S, CH3SH, (CH3)2S, Hx |
| Photobacterium phosphoreum | TMA, Hx |
| Pseudomonas spp. | ketones, aldehydes, esters, non-H2S sulphides |
| Vibrionaceae | TMA, H2S |
| anaerobic spoilers | NH3, acetic, butyric and propionic acid |
As mentioned above, TVB will continue to rise even after TMA has reached its maximum. This latter rise is due to proteolysis commencing when several of the free amino-acids have been used. Lerke et al. (1967) separated fish juice into a protein and a non-protein fraction and inoculated spoilage bacteria in each fraction and in the whole juice. The non-protein fraction of a fish juice spoiled as the whole juice whereas only faint off-odours were detected in the protein fraction of the juice. Although some authors have used the number of proteolytic bacteria as indicators of spoilage, it must be concluded that the turnover of the protein fraction is not of major importance in spoilage of fresh fish.
Some of the compounds typically formed by bacteria during spoilage of fish are shown in Table 5.7 together with the substrate used for the formation.
Table 5.7 Substrate and off-odour/off-flavour compounds produced by bacteria during spoilage of fish
| Substrate | Compounds produced by bacterial action |
| TMAO | TMA |
| cysteine | H2S |
| methionine | CH3SH, (CH3)2S |
| carbohydrates and lactate | acetate, CO2, H2O |
| inosine, IMP | hypoxanthine |
| amino-acid s (glycine, serine, leucine) | esters, ketones, aldehydes |
| amino-acids, urea | NH3 |
The formation of TMA is accompanied by a formation of ammonia during anaerobic storage of herring and mackerel (Haaland and Njaa, 1988). Prolonged anaerobic storage of fish results in vigorous production of NH3 owing to further degradation of the amino-acids, and in the accumulation of lower fatty acids as acetic, butyric and propionic acid. The very strong NH3-producers were found to be obligate anaerobes belonging to the family Bacteroidaceae genus Fusobacterium (Kjosbakken and Larsen, 1974; Storroe et al., 1975, 1977). These organisms grow only in the spoiled fish extract and have little or no proteolytic activity relying on already hydrolysed proteins.
During iced storage of fresh fatty fish, changes in the lipid fraction is caused almost exclusively by chemical action, e.g., oxidation, whereas bacterial attack on the lipid fraction contributes little to the spoilage profile. During storage of lightly preserved fish, lipid hydrolysis caused by bacteria may be part of the spoilage profile.
The two distinct reactions in fish lipids of importance for quality deterioration are:
They result in production of a range of substances among which some have unpleasant (rancid) taste and smell. Some may also contribute to texture changes by binding covalently to fish muscle proteins. The various reactions are either nonenzymatic or catalyzed by microbial enzymes or by intracellular or digestive enzymes from the fish themselves. The relative significance of these reactions, therefore, mainly depends on fish species and storage temperature.
Fatty fish are, of course, particularly susceptible to lipid degradation which can create severe quality problems even on storage at subzero temperatures.
Oxidation
The large amount of polyunsaturated fatty acid moieties found in fish lipids (see section 4.2) makes them highly susceptible to oxidation by an autocatalytic mechanism (Figure 5.16). The process is initiated as described below by abstraction of a hydrogen atom from the central carbon of the pentadiene structure found in most fatty acid acyl chains containing more than one double bond:
-CH = CH-CH2-CH = CH-
Contrary to the native molecule, the lipid radical (L) reacts very quickly with atmospheric oxygen making a peroxy-radical (LOO) which again may abstract a hydrogen from another acyl chain resulting in a lipid hydroperoxide (LOOH) and a new radical L. This propagation continues until one of the radicals is removed by reaction with another radical or with an antioxidant (AH) whose resulting radical (A) is much less reactive. The hydroperoxides produced in relatively large amounts during propagation are tasteless, and it is therefore perhaps not surprising that the widely used "peroxide value" (section 8.2) usually correlates rather poorly to sensorial properties.
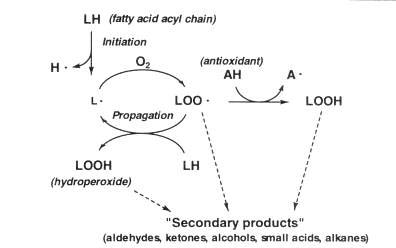
Figure 5.16 Autoxidation of polyunsaturated lipid
The hydroperoxides are readily broken down, catalyzed by heavy metal ions, to secondary autoxidation products of shorter carbon chain-length. These secondary products - mostly aldehydes, ketones, alcohols, small carboxylic acids and alkanes - give rise to a very broad odour spectrum and in some cases to a yellowish discoloration. Several of the aldehydes can be determined as "thiobarbituric acid-reactive substances" (section 8.2).
Metal ions are very important in the first step of lipid autoxidation - the initiation process - in catalyzing the formation of reactive oxygen species as for example the hydroxyl radical (OH). This radical immediately reacts with lipids or other molecules at the site where it is generated. The high reactivity may explain that free fatty acids have been found to be more susceptible to oxidation than the corresponding bound ones, because the amount of iron in the aqueous phase is probably greater than the amount bound to the surface of cellular membranes and lipid droplets.
Fatty acid hydroperoxides may also be formed enzymatically, catalyzed by lipoxygenase which is present in variable amounts in different fish tissues. A relatively high activity has been found in the gills and under the skin of many species. The enzyme is unstable and is probably important for lipid oxidation only in fresh fish. Cooking or freezing/thawing rather effectively destroys the enzyme activity.
The living cells possess several protection mechanisms directed against lipid oxidation products. An enzyme, glutathione peroxidase, exists which reduces hydroperoxides in the cellular membranes to the corresponding hydroxy-compounds. This reaction demands supply of reduced glutathione and will therefore cease post mortem when the cell is depleted of that substance. The membranes also contain the phenolic compound a-tocopherol (Vitamin E) which is considered the most important natural antioxidant. Tocopherol can donate a hydrogen atom to the radicals L- or LOO- functioning as the molecule AH in Figure 5.16. It is generally assumed, that the resulting tocopheryl radical reacts with ascorbic acid (Vitamin C) at the lipid/water interface regenerating the tocopherol molecule. Other compounds, for example the carotenoids, may also function as antioxidants. Wood smoke contains phenols which may penetrate the fish surface during smoking and thereby provide some protection against lipid oxidation.
Hydrolysis
During storage, a considerable amount of free fatty acids (FFA) appears (Figure 5.17). The phenomenon is more profound in ungutted than in gutted fish probably because of the involvement of digestive enzymes. Triglyceride in the depot fat is cleaved by triglyceride lipase (TL in Figure 5.18) originating from the digestive tract or excreted by certain microorganisms. Cellular lipases may also play a minor role.
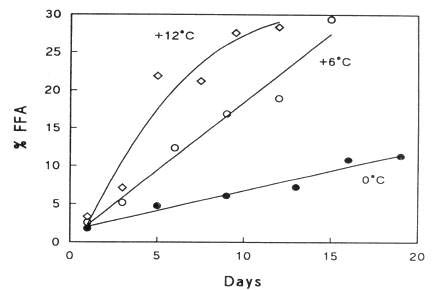
Figure 5.17 The development of free fatty acids in herring stored at different temperatures (Technological Laboratory, Danish Ministry of Fisheries, Annual Report, 1971)
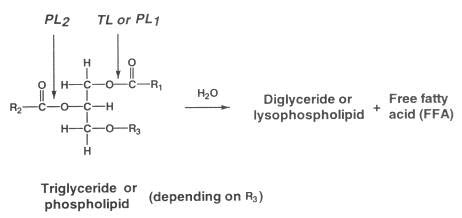
Figure 5.18 Primary hydrolytic reactions of triglycerides and phospholipids. Enzymes: PL1 & PL2 phospholipases;TL, triglyceride lipase
In lean fish, for example Atlantic cod, production of free fatty acids also occurs, even at low temperatures. The enzymes responsible are believed to be cellular phospholipases - in particular phospholipase A2 (PL2 in Figure 5.18) - although a correlation between activity of these enzymes and the rate of appearance of FFA has as yet not been firmly established. The fatty acids bound to phospholipids at glycerol-carbon atom 2 are largely of the polyunsaturated type, and hydrolysis therefore often leads to increased oxidation as well. Furthermore, the fatty acids themselves may cause a "soapy" off-flavour.
![]()
![]()
![]()