![]()
![]()
![]()
Plates 27 – 29 (pp. 199 – 201) and Figs. 8 – 10 (pp. 202 – 204)
Species affected and geographical range
Freshwater and euryhaline fish of many families. Common in fish of all major African
water systems (Fryer, 1968a). Several species occur in cichlids, in siluriforms and also
in brackish waters (E. latus in Sarotherodon melanotheron heudelotii in Ghana). Lates
spp. (Nile perches) in both West African water bodies and in some of the East African
Great Lakes are infected by Ergasilus kandti (Fryer, 1968a; Paperna, 1969; Thurston,
1970). All freshwater species are endemic to Africa (Fryer, 1968a), the euryhaline E.
lizae is widespread in the Atlantic and Mediterranean coastal regions (Roberts, 1970)
and has also become established in freshwater ponds (in grey mullets, tilapia and carp)
in Israel (Paperna & Lahav, 1971).
Description, taxonomy and diagnosis
Only subadult and adult females occur of fish, mostly on the gills, a few of the genus
Paraergasilus, may attach to sites other than gills. The cephalothorax constitutes half or
more of body length, the first of four thoracopodes occurs at about mid-length.
Segmentation of the thorax (except the first segment, fused with the head) and of the
abdomen is distinct. The second antenna terminal segment is hook-like in Ergasilus and
three clawed in Paraergasilus. Eggs are clustered in a bunch rather than arranged in a
single line (in caligoids). For detailed morphological examination, parasites fixed in
formalin or alcohol, should be treated by a clearing agent (- by graded transfer into lactic
acid or glycerin, or digested in sodium hydroxide).
Life cycle and biology
In Ergasilidae only the female is parasitic, and is found on the gills of fish. Males are
free-living and there is a prolonged, free-living larval development which includes three
to six stages of nauplii and four to six stages of copepodites (lasting from 10 days to over
a month). These free-living stages feed on nanoplankton (Ben Hassine & Raibaut, 1981;
Raibaut, 1985). Females attached to gills produce eggs in two sacs which are attached
to the genital segment. The number of eggs (20–100) is variable with species and
apparently also with age and metabolic health as influenced by the location of attachment
on the gills. The time required for hatching is temperature dependent, 3–6 days in optimal
ambient temperature. High subtropical summer temperatures, coupled with depletion of
dissolved oxygen and elevated salinities in lagoon habitats, was shown to be detrimental
to both developing eggs and the free living stages of E. lizae (syn. E. nanus) (Raibaut
et al., 1975). It has been established that elevated salinity delayed larval development
from 10 days at 15 ppt to 27 days at 26 ppt (Ben Hassine et al., 1983). Biological data
on ergasilids infecting fish of African tropical freshwaters are lacking.
Pathology
Ergasilids attached to gill filaments produce small foci of erosion; apparently feeding
involves excretion of proteolytic enzymes for external digestion. Such erosion processes
occur in E. megacheir infections in cichlids (Fryer, 1968a) and E. lizae infections in grey
mullets. Erosion and degradation processes may extend beyond the epithelial lining,
resulting in obstructed branchial blood vessels. Irritation often results in responsive
hyperplasia of the epithelium, which, as infection is prolonged, may extend over large
areas of the gills, causing fusion and embedding of lamellae, with a resulting decrease
in the respiratory function of the gills (Kabata, 1970; Paperna & Zwerner, 1981).
The nature and magnitude of the pathological changes in the gills varies with host and ergasilid species. Infection by the more opportunistic species, and under stressful conditions (adverse water conditions, deficient nutrition and overcrowding) is likely to induce severe clinical and pathological effects. Marked epithelial hyperplasia is stimulated in the gills of L. albertianus infected by E. kandti, in a variety of fish infected with E. cunningtoni (Fryer, 1968a) and in tilapia infected with E. lizae. Pond reared (70–80 g) M. cephalus infested with 100–200 copepods and those of 250 g infested by 1500–2000 copepods, were severely emaciated. Losses at harvest reached 50% compared with 10% in uninfested ponds (Yashuv, 1972; Paperna & Overstreet, 1981). Emaciation was also observed in wild caught E. lizae infected L. ramada.
Epizootiology
Ergasilids vary in their level of host specificity, some are specific at least to their host
genus (notably ergasilids infecting Cichlidae). Others, are less specific in their choice of
hosts or are even opportunistic (E. cunningtoni has been recorded from fish belonging
to seven families, Fryer, 1968a). However, the least specific and even the opportunistic
species (E. lizae, E. kandti, E. cunningtoni, E. sarsi, Paraergasilus lagoonaris - Fryer,
1968a; Paperna, 1969; Lahav & Sarig, 1968) still demonstrate, some predilection for a
particular host, while the occurrence in other (not phylogenetically related) fish is either
sporadic or linked to particular environmental or stressing circumstances.
In African lakes and rivers, infections with ergasilids are usually moderate. In habitats such as regressing pools in river beds at the dry seasons, confined fish may become heavily infected, for example, juvenile Clarias lazera found infected with about 100 E. sarsi in a pool in the upper reaches of the Volta river (Paperna, 1969). Heavy infestation with E. lizae (reported as E. nanus) has been reported to occur annually in Mugil cephalus congregating in the euryhaline L. Ischkeul in northern Tunisia, but other species of grey mullets in this lake were considerably less infected. Peak infestation in M. cephalus, in 100% of fish with up to 2000 copepods per fish, occurred in spring when water temperatures were moderately elevated (to 21°C) and while the salinity was declining (to 4 ppt). By mid-summer, when water temperatures are reaching 27°C and salinities are raised to 14 ppt, infestation sharply declines (Raibaut et al., 1975). Individual L. albertianus in Lake Albert were found to be infected with 352–1926 E. kandti. It has been suggested that these heavy infections serve as a contributing factor in mass mortalities of this fish occurring at times in the lake. It is also noteworthy that this Ergasilus was absent from L. albertianus introduced from L. Albert into the Victoria Nile, L. Kyoga, and L. Victoria (Thurston, 1970). E. kandti infects Lates spp. of L. Tanganyika and has been found to infect L. niloticus in the man-made Lake Volta (Paperna, 1968). The fastidious, nutritive and environmental requirements of the free-living stages seem to limit distribution of ergasilids when their hosts are translocated. This is further emphasised by the fact that of the numerous species of Ergasilus (over 80 - Kabata, 1985) and related copepod genera, very few ever become established in pond systems.
One of the few ergasilids which has adjusted well to fish ponds is Ergasilus lizae (syn. E. fryeri, Paperna, 1964b, “E. seiboldi” of Lahav & Sarig, 1968). It is also one of the few species whose free-living developmental stages could be demonstrated experimentally (Ben Hassine & Raibaut, 1981). Stocking fish ponds in Israel with Liza ramada caught in drainage canals resulted in the infestation of grey mullets (Mugil cephalus and L. ramada) farmed in the ponds. Heavy infections and mortalities occurred predominantly among M. cephalus. Thereafter, infection spread to further farms where tilapia hybrids also became infected and suffered heavy mortalities. Carp became moderately infected but with no clinical implications (Lahav & Sarig, 1968; Sarig, 1971; Paperna & Lahav, 1971; Paperna & Overstreet, 1981).
Control
In Israeli fish farms, ergasilid infections were effectively treated through repeated
bi-weekly applications of 0.125–0.150 ppm Bromex; the efficacy of each application is
about 92%. Treatment of grey mullets is feasible only in ponds with water chlorinities
above 400 ppm. At lower salinities recommended doses become toxic to the fish. This,
however, does not apply to tilapia or carp which tolerate treatment in all ranges of
salinities (with a safety index of 30). Other organophosphates - Neguvon (Dipterex,
Masoten) or Malathion are effective at similar concentrations and may be used to treat
mullets in freshwater too (Lahav & Sarig, 1968; Sarig, 1971).
Species affected and geographical range
Lernaea cyprinacea, an opportunistic species infecting fish of many families as well as
amphibians (Yashuv, 1959), is widely distributed in Eurasia, North America and southern
Africa mainly via translocations of edible and ornamental cyprinids (Hoffman, 1970). A
lernaeid population found only on jaws of Tilapia (Oreochromis) spp. in L. Victoria (Fryer,
1961a) has been identified as L. cyprinacea. Other lernaeids of African fish are endemic,
L. barnimiana may be found on both cyprinid and cichlid hosts, as well as on Lates
albertianus, while all the rest restrict their host preferences to fish of the same families
or genera. Fish of many diverse taxa are associated with either Lernaea spp. or
representatives of allied genera; Opistholernaea (each species on a particular host
genus - O. longa on Lates, Afrolernaea on Mormyridae, Lernaeagiraffa and Dysphorus
on Heterotis - Fryer, 1968a).
Host preferences may differ in populations from different geographical regions; L. hardingi throughout Africa is associated with Cichlidae, in L. Bangweulu and L. Meru they principally infect Chrisichthys mahusi and also rarely Synodontis nigromaculatus. Alongside widely distributed species, there are species which are not only host specific but apparently also restricted to one lake; L. bagri, L. tuberosa, and L. inflata (Fryer, 1959, 1968a).
The gill adapted lernaeids, Lamproglena and Lamproglenoides species from African fish are all endemic, each species occurring in representatives of different families: L. monodi (in cichlids), L. hemprichii (in the genus Hydrocynus, Characidae) and L. clariae (in Clarias spp.) are the most widely distributed (Fryer, 1968a).
Description, taxonomy and diagnosis
The rod-shaped, unsegmented, or partly segmented parasitic stage lernaeid female is
anchored, with the aid of a specialised holdfast organ, to the host skin or buccal mucosa.
Larval stages, copepodites and copepod-shaped males are attached to the gills.
Differentiation to lernaeid genera and to species in the genus Lernaea is based mainly on the morphology of the holdfast organ (anchors) of the parasitic females. The growth and branching of the latter, however, is greatly affected by the consistency of the tissue into which the holdfast organ is anchored. Infrageneric taxonomy may often be controversial, as intraspecific variations often overwhelm interspecific differences in the holdfast organ structure (Fryer, 1961a).
Differentiation to genera is as follows:
| Lernaea: | head with 2–3 (rarely 4) pointed or swollen horns, the trunk may thicken gradually towards its posterior end. |
| Lernaeagiraffa: | head horns are soft, short and rounded; “neck” elongated; posterior part of the body (trunk) abruptly thickened and flattened dorso-ventrally. |
| Opistholernaea: | head with 4 horns, with 2 posterior horns directed symmetrically backward to form a 90 degree angle. A lateral outgrowth extends from the “neck” region; it may be, however, located in very close proximity to the head to become functionally part of the anchor complex. |
| Afrolernaea: | head with anterior short protruberences; the neck is very long and the trunk abruptly thickening. |
| Dysphorus: | head horns are dendritic; the neck has branched appendages. |
| Lamproglena and Lamproglenoides: | attach to gills; parasitic females retain copepodid features; segmentation, and large mouth parts, with cephalothorax always less than half total length, second antenna small and weak, maxilipeds with terminal claws, thoracopods present, but sometimes fewer than 4 pairs. Copepodites and males are reminiscent to those of Lernaea. |
Copepodites of lernaeaids attached to gills are readily differentiated from ergasilid parasitic females in lacking the hook (spine) type terminal segment of the second antenna.
Life cycle and biology
Parasitic females of L. barnimiana of L. George produce egg sacs containing 75–205
eggs. At 21–25°C, hatching occurs after 2 days. Hatched nauplii moult into second stage
nauplii within 19 hours, and further moult to the third stage nauplius (metanauplius) within
42 hours after hatching. Free, infective copepodite stages appear 97 hours after
hatching. The latter survive up to 4 days in the absence of host fish.
Copepodites preferentially attach to fish gills and, less readily, to the walls of the buccal cavity or the skin. Copepodites undergo four successive moults, while increasing in size from 0.3–0.4 mm to 0.6–0.8 mm. In fifth stage copepodites sexual dimorphism becomes apparent, with females being larger than males (0.8–0.9 vs 0.7–0.85 mm).
At this stage, 13–16 days after hatching, copepodites abandon their attachment to their host, and either leave in search of a new host or relocate themselves on the same host. In their new attachment sites in the skin, fins or buccal cavity, of new or the same hosts, copepodites moult into their last copepodite stage (male and female cyclopoid stages). Fertilisation occurs in the fifth copepodite or the cyclopoid stage. By the termination of larval development, 15–18 days after hatching, males gradually disappear and females continue their transformation into adults through elongation of the body, loss of external segmentation, degeneration of legs and the development of a holdfast organ. Early juvenile females first appear 19 days after hatching; the first egg sac in the mature female was observed 23 days after hatching. Egg sacs were produced at three day intervals. Females survived up to 9 days after first oviposition and 32 days after hatching yielded about 3 pairs of eggs sacs with a total progeny of 800–1200 eggs (Thurston, 1969).
The time required for larval development of L. cyprinacea, infecting fish outside the tropical regions, is temperature dependent. At ambient temperatures of 20–25°C, the rate of development of L. cyprinacea was slightly faster than that of L. barnimiana. At 25°C, hatching occurred after 36 hours, the first copepodites appeared 3–5 days later, early females appeared 13–17 days after hatching and the first egg sacs 18–22 days after hatching. Egg sacs contained 60–200 eggs, and females survived 7–10 days after the appearance of the first egg sac. At 27–30°C the first females appeared 9–12 days after hatching and adult females survived 8 days. Temperatures of 30°C seem to be the upper limit for optimal development, egg development seems to be affected as levels of dissolved oxygen decline. Lower temperatures did not affect egg production, but prolonged larval development; early adult females at 12–16°C appeared 60 days after hatching. The life span of females at these temperatures was, however, extended to 2–3 months (Yashuv, 1959; Lahav & Sarig, 1964). At ambient temperatures of 12–19°C, during the winter season outside the tropics, only three or less generations are produced, as compared with seven or eight generations in similar lengths of time during the Mediterranean summer or in tropical waters.
Yashuv (1959) noted considerable variability in the development time of the various stages as well as in the life span of the anchored female. In addition to aquatic conditions, variations were related to the nature of the interaction between parasite and host. Repeated or prolonged infestation stimulated an accelerated host inflammatory response and ulceration around anchored females, which hastened their evacuation in being denied firm attachment and nutrition. Such processes have been suggested to be linked with activation of non-specific or specific defence mechanisms (Shields & Goode, 1978).
In the East African lakes, different fish species serve as preferential hosts for copepodites and adult Lernaea. Copepodites of L. cyprinacea, infecting cichlid fish in L. Victoria at the adult female-stage, develop in Bagrus docmac. B. docmac of L. George and B. bayad of L. Albert host copepodites of L. barnimiana, whose adult female stages attach to Barbus altianalis, cichlids and Lates albertianus (Thurston, 1969, 1970).
Copepodites, however, are opportunistic in their host preferences and in the absence of preferential hosts (in ponds and experimentally), both larval development and female stage infection will take place in the same individual or conspecific hosts.
Other species and populations of Lernaea also seem to be less fastidious in their choice of hosts for larval development than for their adult stages. Adult females of L. cyprinacea in L. Kinneret preferentially infect cyprinid fish, while copepodites may also be found on cichlid fish (Paperna, 1964b).
The life history of Lamproglena and Lamproglenoides follows the same pattern as other lernaeids, with at least two stages of copepodites attached to gills. Males are lost after fertilisation. Lamproglenids appear to develop and mature on the same host species.
Pathology
Both copepodites infecting gills, and females infecting any site on the fish integument,
may harm the fish. Heavy infection of gills by copepodites of L. barnimiana or L.
cyprinacea causes damage to the tissue which is in immediate contact with the parasite
and stimulates extensive hyperplasia of the entire gill epithelium. Copepodite infections
are potentially lethal to small fish, while large fish (such as Bagrus) will tolerate extremely
heavy gill infections (over 1000 per fish - Paperna, unpublished). The attachment of the
adult females to the integument stimulates an intense inflammatory response. The tissue
around the anchor turns into a granuloma or necrotic lesion, and is later transformed into
a fibrotic encapsulation (Kabata, 1970, 1985). A large proportion of the body of
Opistholernaea laterobranchialis is embedded in the host tissue; in L. Bangweulu it is
always found in the mouth of its host, while it is anchored within fibrous growth in or near
the eye socket (Fryer, 1968a). Economic losses due to infection not only results from
direct harm to the fish, but also from disfigurement which renders both fish grown for
food and ornamental fish unsuitable for sale.
Pathogenicity of lernaeids largely depends on their host size and their attachment site preferences. Infection by a single or 2–3 females is very damaging or even deadly to young or small fish (<40 mm long). Parasites may anchor in the liver, the intestine or the brain (Paperna & Thurston, 1968). In the sardine-like Engraulicypris (Rastrineobola), even one specimen of L. tuberosa or L. inflata was a serious encumbrance (Fryer, 1968a). The ubiquitous skin lernaeids, such as L. cyprinacea and L. hardingi and some populations of L. barnimiana, cause severe damage only in heavy infections.
Lernaeid species with narrower host ranges seem also to be more selective in their attachment site preferences. Species preferentially aggregating in particular sites induce severe local damage, often with adverse consequences to the host's function and survival. Aggregated infections of the mouth and lips occurs in Barbus altianalis with L. barnimiana in the Victoria Nile, and in cichlids by L. cf. cyprinacea in Lake Victoria. Strong currents in the Victoria Nile enable copepodites to settle only in the buccal cavity in B. altianalis. Lip and mouth infection in other East African lakes occurs in B. meridionalis with L. bagri and in cichlids with L. tilapiae, L. palati and Opistholernaea laterobranchialis. In mouth breeding cichlids, mouth and gill infection impairs breeding (Fryer, 1968; Fryer & Iles, 1972). Fin infections are caused by L. lophiara in cichlids, and Afrolernaea longicollis infects gills of mormyrid fish (Fryer, 1968). In Southeast Asia L. polymorpha aggregating around the eye in bighead (Aristichthys nobilis) cause destruction of the lens and blindness (Shariff, 1981). Gill infections of Lamproglena monodi cause some localised hyperplasia of the epithelial tissue, while other gill infections by L. clariae and L. intercedens cause intense epithelial proliferation which may seriously interfere with respiration.
Epizootiology
The prevalence of natural infections even in large water bodies such as lakes reached
94% in Mormyrus longirostris infected by Afrolernaea longicollis and 56% in B.
meridionalis infected with L. bagri. L. albertianus was frequently (50%) infected with L.
barnimiana in L. Albert but free of such infection in lakes Kyoga and Victoria. Heavy
infections of L. barnimiana were also frequent in B. altianalis in the Victoria Nile (up to
106 per fish). Nevertheless, in most fish populations infection was overdispersed, with
few individuals attracting larger numbers of parasites (66 L. cyprinacea in the buccal
cavity of O. variabilis, from L. Victoria; 55 L.bagri in B. meridionalis in L. Malawi) (Fryer,
1968a).
Secluded habitats such as small dams and fish ponds accelerate risks of infestation. In Ghana, L. hardingi occurs frequently in O. niloticus, Sarotherodon galilaeus, Sarotherodon melanotheron heudelotii and T. zillii stocked in dam reservoirs and ponds. Infection is usually low to moderate (up to 28% in some habitats). At times, however, in some dams infestation becomes extremely severe (Paperna, 1969). L. cyprinacea introduced into southern Africa (reported as Lernaea sp.) is sometimes found in large numbers on carp in dams and rivers and and on native Barbus and Labeo co-existing with native lernaeid species (L. barbicola and L. barnimiana -- Van As & Basson, 1984).
Heavy infections are not infrequent in Israeli fish farms, mainly in carp, they are less frequent in Asian carps and occur rarely in cichlids. There are also some records of epizootic infection in grey mullet grown in freshwater ponds (Yashuv, 1959; Sarig, 1971). Infection with adult L. cyprinacea is also common in native cyprinid fishes and Gambussia affinis from natural habitats, while being absent from cichlids and Clarias lazera; copepodites occur also in cichlids (Paperna, 1964a).
Infections of L. cyprinacea were reported from Oreochromis mossambicus and O. niloticus introduced to culture in Southeast Asia (Kabata, 1985). L. lophiara, parasitic on Central African cichlids has been identified in Thailand from O. mossambicus and various other indigenous and exotic fishes (Kabata, 1985).
Control
Parasitic females of lernaeids are tolerant to organophosphate and organohalogen
insecticides. A dip in potassium permanganate (KMnO4) - 20–25 ppm for 2–3 hours or
application of 8 ppm concentration to ponds effectively kills attached female lernaeids.
Treatment should be cautiously applied, preferably by experienced personnel, as
effective concentrations are too close to toxic levels (safety index 1.7–2.0). Suitable only
for fish over 25 g and tolerance will vary with species (for a protocol for application
methodology see Sarig, 1971; Kabata, 1985).
Recently the insecticide Dimilin (R) (Philips-Dupar, Netherlands product) was found effective against adult females at concentrations of 0.03–0.05 ppm. This insecticide has not been cleared for use with food fish and its degradation in the environment is slow so that contaminated waters should not be released until at least 30 days after treatment.
An alternative strategy is treatment directed to suppress lernaeid infection through eradication of copepodite stages by repeated treatments of ponds with Dipterex (0.25 ppm) and Bromex (0.12 ppm). Treatments are repeated to coincide with the duration of larval metamorphosis from early copepodite to adult stage, which is temperature dependent. Recommended time intervals for treatment of L. cyprinacea copepodites are: 12 days at 20°C, 9 days at 25°C, 7 days at 30°C, 5 days at 35°C. Below 20°C one monthly treatments will be sufficient.
Parasites, host species affected and geographical range
Paeonodes lagunaris is known only from the buccal cavity of Sarotherodon
melanotheron, from Sakumo lagoon in southern Ghana (Van Banning, 1974; Pauley,
1974).
Mugilicola smithae occurs on Anguilla mossambica, in South Africa (Jones & Hine, 1978).
Achtheres micropteri has been introduced with its host Micropterus salmoides into South Africa (Fryer, 1968a).
Sciaenophilus pharaonis (syn. S. inopius) was reported from Labeo niloticus and L. forskalii from the Nile in Egypt (Humes, 1957).
Description, taxonomy and diagnosis
The parasitic females (the only known stage) of the genera Mugilicola and Paeonodes
are Therodamasidae (Poecilostomatoida, the same suborder as Ergasilidae), and
characterised by a small head, embedded in the host integument (both from the roof of
the mouth), an unsegmented long neck and trunk bearing paired legs, and a minute
abdomen. Eggs are arranged in bunches. Paeonodes has four pairs of thoracic legs,
while Mugilicola has three. The head in Mugilicola is large, while in Paeonodes only the
clawed second antenna is readily distinguishable.
Achtheres micropteri is a lernaepodid copepod. Females are permanently attached to the inner surface of their host gill arches by a specially modified leg, the first maxilliped.
Segmentation and thoracic legs disappear in the mature stages. Egg sacs are claviform, in a bunch. Males are dwarfed and attached to females.
Sciaenophilus pharaonis is a gill adapted caligiid, with the flattened cup shaped head reduced and enlarged and elongated thoracic and abdominal segments.
Life cycle and biology
The life histories of Paeonodes lagunaris and Mugilicola smithae, are unknown but are
apparently reminiscent to that of ergasilids (18.1).
In Achtheres, larvae hatch into a free swimming first copepodite stage, at the second copepodite stage they attach to fish by an attachment filament and develop into adults. After mating, males cling to females (Hoffman, 1967).
The life history of Sciaenophilus pharaonis is unknown, but should be the same as other caligiids, e.g. copepodites hatch from eggs, attach to gills and develop into chalimi stages attached to the gill tissue by a specially formed filament. The chalimi undergo several moults until they transform into adult males and females (Raibaut, 1985).
Pathology and epizootiology
All these copepods seem to have little damaging effect on their host fish at the infection
levels observed (all were natural infections).
Paeonodes, Mugilicola and Sciaenophilus are marine copepods adapted to brackish water habitats, or carried with their host into inland waters (Sciaenophilus). P. lagunaris infection on S. melanotheron is highest (100%, mean 5.3 per fish) during the heavy rainy season when salinity (and temperature) of the lagoon water is low (<10 to 20 ppt; <26°C), and declines (up to 17%, with mean of 1 per fish) as salinities and temperatures are elevated (30 to 45 ppt, 27–8°C) (Pauley, 1974). Infestation of M. smithae occurs in elvers (in 0.6–3%) of the southeastern Cape rivers, and only very rarely in glass eels (0–0.3%) (Jones & Hine, 1978).
Achtheres micropteri, introduced with the black bass (Micropterus salmoides) remained confined to its exotic host (Fryer, 1968a).
Control
Not tried, apparently the same as for other crustaceans.
Species affected and geographical range
Twenty nine endemic species occur in Africa, in fish of diverse families. Argulus africanus
and Dolops ranarum are opportunists and occur in diverse fish in all major systems of
Africa. Allied species, A. rhiphidiophorus and A. cunningtoni, replace A. africanus in
some East African lakes connected to the upper Nile, and co-exist in others, in part at
least, due to later artificial introductions of fish. The remaining, including members of the
genus Chonopeltis, are host specific with a restricted distribution to a single water system
or to a lake. The largest number of species was reported from the Congo basin followed
by that of L. Tanganyika. In West Africa few species, mainly the ubiquitous, occur (Fryer,
1968a). In South African fish, in addition to a few locally endemic species, D. ranarum
is widespread as is the ubiquitous Eurasian species A. japonicus introduced apparently
with carp (Kruger et al., 1983; Van As & Basson, 1984; Avenant & Van As, 1985; Van
As, 1986; Avenant-Oldewage, 1991). In Lake Kinneret, an endemic species A.
tristramellae was found infecting the endemic cichlid genus Tristramella (Paperna,
1964b).
Description, taxonomy and diagnosis
Argulids (“fish lice”) are dorso-ventrally flattened and covered dorsally by a rounded or
horseshoe shaped carapace. Ventrally positioned head appendages are developed for
attachment, four thoracic segments each bear a pair of bifid swimming legs. The
abdomen is comprised of a single bi-lobed unit which contains testes or a round seminal
receptacle.
Only one species of Dolops is present in Africa, it differs from Argulus in having the second maxilla armed with a hook rather than a sucker, characteristic of the latter.
Chonopeltis are usually the smallest; head appendages are feeble and rudimentary, the mouth tube found in the other argulids is absent, whereas the cup-like sucker of the second maxilla is distinctly developed. The carapace is reduced in size and width. Unlike the above argulids, which are active swimmers, Chonopeltis is not capable of swimming if removed from the host.
For species diagnosis of Argulus and Chonopeltis readers should refer to relevant publications (Fryer, 1956, 1959, 1961b, 1968a, 1977; Avenant-Oldewage, 1991).
Life cycle and biology
Species of Argulus and Dolops ranarum are parasitic throughout life, but leave the host
to moult or to lay eggs, and during this process will also change hosts. Both males and
females may survive free living for as long as 15 days (Hoffman, 1977). A. africanus, A.
cunningtoni, A. rhiphidiophorus and D. ranarum have a preference for smooth-skinned
fish (Siluriforms and lung fish), the same species may, however, infect buccal and
opercular mucosal integuments of scaly fish, notably cichlids (Fryer, 1968a). These
opportunistic argulids spread all over the body or the skin in smooth skinned hosts,
(although in Bagrus docmac they preferentially settle on the head and near the mouth)
while in scaled fish they occur only on the buccal and branchial cavity mucosa (Avenant
& Van As, 1985; Paperna, 1991). Chonopeltis are site specific and remain constantly on
the host until departure for oviposition.
Eggs are deposited in masses on substrates in the water and may become covered by a gelatinous substance. Argulid species vary in the number of eggs they produce, in total (20–600) and per each batch (Fryer, 1968a; Hoffman, 1977). Egg strings laid by A. japonicus contained 5–226 eggs, and each female laid 1–9 strings (Shafir & Van As, 1986). Newly hatched argulids either resemble modified nauplii (in A. japonicus), or more often, are already transformed to “juvenile adults” (in A. africanus and other spp. and in D. ranarum). In Chonopeltis the larvae are intermediate between nauplii-type and “juvenile adult” (Fryer, 1968a).
Dolops ranarum females deposit up to 566 eggs (Fryer, 1959); eggs are heavily yolked. Eggs develop to hatching 25–35 days at 24°C (Fryer, 1964). At 20°C, hatching occurs after 57 days (Avenant et al., 1989). Eggs do not hatch simultaneously; at 24°C, the time between hatching of the first and last egg is about 10 days.
The annual cycle of D. ranarum in the tropical region is unknown, while outside the tropics, under a seasonal climatic regime (with winter temperatures declining below 14°C), D. ranarum has a one year life span which terminates in reproduction. Observations of seasonal occurrence in an impoundment near Pretoria in Transvaal (Avenant & Van As, 1986) showed that the highest infections occurred in its main hosts, Oreochromis mossambicus and Clarias gariepinus, by late summer and autumn (March to May) and the lowest by spring (October). In these natural infections, individuals showed a progressive increase in total length from November (early summer) till August–September (late winter, early spring), when they reached maximum sizes, and 50% of the females, all exceeding 9 mm in length, were gravid. Departures from hosts for spawning is evident from the extreme decline of infection observed by October. After spawning females are apparently unable to relocate to new hosts. The smallest individuals found were 2 mm in length for males and 3.5 mm for females and while newly hatched juveniles were never found on these host fish, this could suggest that young stages utilise different hosts. Very small (1.45 mm long) individuals were revealed from the mouth folds of one Oreochromis andersoni (Avenant et al., 1989). Only brief data are available on other African argulids: duration of development of A. africanus eggs is longer than in Dolops ranarum (30–36 days at 22–24°C - Fryer, 1956) suggesting a longer generation time.
Egg development of the eurasian exotic A. japonicus is asynchronous as well as temperature dependent. Hatching started after 61–10 days within a temperature span of 15–35°C. There was some impairment of hatching success in temperatures above 30°C (Shafir & Van As, 1986). When maintained at 25–27°C, eggs hatch after 10–20 days, parasites reach maturity 21 days later and the life cycle was completed in four to five weeks (Sarig, 1971). Adults can survive ambient water temperatures below 10°C, while oviposition ceases at temperatures below 16°C. Embryonic development is interrupted at 10°C and larval development is completely interrupted below 12°C (Bauer, 1959; Shafir & Van As, 1986). Finding gravid females at any time of the year suggests that, unlike Dolops ranarum, reproduction in A. japonicus in southern Africa, and evidently also in lsrael, is continuous and not restricted to a particular season. Growth of the parasite population is, however, delayed in winter, when low temperatures slow down or even halt egg development to hatching.
Another important factor in distribution of argulids is their tolerance to water quality, in particular dissolved oxygen. D. ranarum, the only branchiuran possessing haemoglobin, seems to be capable of withstanding low levels of dissolved oxygen and to survive on siluriforms and lungfish which enter eutrophic, deoxygenated habitats. D. ranarum seems to be capable of surviving on mud aestivating fish (Fryer, 1968a).
Pathology
Argulids infecting the skin or mouth of fish induce severe local damage to the integument.
Damage is caused by the piercing proboscis stylet which enters deep into the skin's
dermal layer and secretes lytic and toxic substances resulting in acute haemorrhagic,
inflamed wounds. Attachment to and crawling on the skin also causes irritation and
abrasions leading first to proliferation and later to desquamation and erosion of the
epithelium. Damage is quickly alleviated where parasites can actively change
attachment sites. When a shift of attachment site is slowed or prevented by
overcrowding, in heavy infections, particularly with site specific parasite species, chronic
inflammation develops, the infected surface of the integument ulcerates, the epithelium
is destroyed and the dermis becomes exposed (Kabata, 1970, 1985; Paperna &
Zwerner, 1976).
Histopathological changes include both proliferative changes in the epithelium and inflammatory changes; haemorrhages and infiltration, of either predominantly lymphocytes (in A. japonicus infections), or (in D. ranarum and A. africanus infections of Bagrus docmac) of eosinophiles. Persistent damage to the skin further leads to the depletion of mucous cells, to degeneration and disintegration of the epithelial layer, disruption of the basal membrane and distortion of the underlying sheets of collagen of the dermis (Paperna & Zwerner, 1976; Paperna, unpublished). In D. ranarum infections there was a marked epithelial oedema (spongiosis), acantholysis (loss of adhesion between adjacent cells) and patchy cellular degeneration evident by karyolysis (Paperna, unpublished).
Persistent irritation caused in heavy infections may affect fish appetite with resulting anorexia, and cessation of growth (Kabata, 1985, 1978; Paperna, 1991). Induced wounds become secondarily contaminated by bacteria and fungi (Saprolegnia). A. coregoni infection of salmonids was followed by furunculosis caused by Aeromonas salmonicida (Shimura et al., 1983). Another aspect of Argulus infection is their possible role as vectors of viral infections; spring viraemia of carp (SVC) has been transmitted via A. foliaceus (Ahne, 1985). In Israel, a decline in A. japonicus populations in fish farms during the past decade coincided with complete disappearance of papiloma (carp pox), once highly prevalent in farmed carp (Landsberg, 1989).
Epizootiology
Dolops ranarum is the only ubiquitous argulid potentially pathogenic to farmed fish. Lates
albertianus of lakes Albert and Kyoga and of the Victoria Nile are infected with D. ranarum
(Thurston, 1969). Particularly heavy natural infection (to over 30 per fish) has been noted
in most fished Bagrus docmac (78%) landed in northern L. Victoria. Parasites were also
often numerous on Clarias gariepinus and lungfish. The same fish were infected, though
with smaller numbers, with A. africanus (41% of B. docmac) (Paperna, unpublished).
Mbahinzireki (1980) reports an average prevalence of 37% D. ranarum infection in B.
docmac from the same waters, with a mean of six per fish. Infections were, as observed
earlier, heavier (60–90% with up to 37 D. ranarum) in the larger market sized fish, and
less abundant in the younger (less than 40 mm long) fish. In this survey A. africanus was
reported to be rarely found.
A. africanus were as numerous as, or even outnumbered, D. ranarum in scaled fish, notably cichlids (in 55% of T. esculenta and in 29% of T. variabilis). In Lake Albert, A. africanus is absent and replaced with A. cunningtoni and A. rhiphidiophorus which heavily infect both smooth skinned siluriforms, scaled cichlids and L. albertianus. In other lakes, other species may produce massive infestations. In L. Tanganyika dead specimens of Auchenoglanis occidentalis were completely covered by A. striatus and A. inciscus (Fryer, 1968a).
In southern Africa D. ranarum occurs predominantly on two hosts, C. gariepinus and O. mossambicus (overall prevalence in surveyed impoundments in Transvaal, 24% of 793 and 26% of 2356 examined fish of the two species respectively). Infection is less frequent in Eutropius depressirostris and species of Labeo and Barbus. Natural infections are usually low (about six per fish) and only exceptionally high (up to 35, in C. gariepinus, in a dam where prevalence was as high as 88%).
A. japonicus introduced to southern Africa on carp and/or goldfish, is today an omnipresent parasite of both carp and native fish; species of Barbus and Labeo, Clarias gariepinus and occasionally also cichlid fish. In some dams and lakes, prevalence of infection in cyprinids and in catfish can reach 93–100% and 78% respectively, with 10 to over 250 parasites per fish (Kruger et al., 1983). Extremely heavy infections of A. japonicus occurred in winter on C. gariepinus trapped in dams, restricted (and stressed?) by the low ambient temperatures (8–13°C) (Kruger et al., 1983; Shafir & Van As, 1985).
In Israel A. japonicus which in the past have been a serious pest of farmed carp, for several years has rarely been found on pond reared carp (Landsberg, 1989). However, it became a frequent parasite of fish in natural habitats, particularly of the Jordan system, and the population is also on the increase among pond reared ornamental carp and goldfish (Paperna, unpublished). In the ponds it occurred throughout the year, while its reproduction and abundance peaked in the summer months at ambient water temperatures of 20–28°C. It has been suggested that the decline in the population of this argulid in ponds has been due to successful eradication efforts with insecticides, taking into consideration the residual nature of Lindane (Paperna, 1991) (and in spite of the observed induced drug resistance, Lahav et al., 1962; see below). The reduction could also have been an outcome of intensification of stocking densities in ponds with a resulting acceleration of eutrophication and increase in nitrite waste levels (Landsberg, 1989).
Control
Both organohalogen and organophosphate insecticides effectively kill agulids with
reliable safety margins to fish (safety indices of 12–20) (Sarig, 1971). Use of Lindane
was abandoned because of its residual effects, risking humans and environmental safety
as well as the resistance acquired by argulids following repeated treatments (Lahav et
al., 1962). Insecticides in current use are Malathion and Dipterex (Dylox, Masoten,
Neguvon) applied to ponds at a dose of 0.25 ppm (safety index of 12 for carp and tilapia),
or Bromex (Dibrom, Naled) at a dose of 0.12 ppm (safety index of 32) (Sarig, 1971).
Pyrethrum extracts were tested with success, but were never introduced into routine use
(Yashuv, pers. comm.).
Species affected and geographical range
Isopoda are parasites of marine fish and infections have been reported in euryhaline fish
in estuaries. Nerocila orbignyi, a common parasite of marine and estuarine fish, including
grey mullets (Trilles, 1965) was reported to infect Sarotherodon galilaeus in a Nile Delta
lake (Wunder, 1962).
A few species of cymothoid isopods (Lironeca spp.) occur in African freshwater fish (apparently as marine relics) in the Congo basin and in L. Tanganyika (in a cichlid, a clupeid and a citharinid) (Fryer, 1965a, b, 1968a,b).
Opportunistic euryhaline cymothoids of several families and genera infect farmed fish in Southeast Asia, including cultured tilapia (by Aliotropus typus, A egidae) (Kabata, 1985).
Description, taxonomy and diagnosis
Isopoda are the largest crustaceans found on fish (20–50 mm long). The body consists
of three regions, the head is unsegmented and bears two pairs of antennae, a pair of
variable sized eyes and a mouth. The maxilliped covers all other mouth parts, a second
region, the peraeon, of 7 segments, each with a pair of legs (peraeopods) and a six
segmented pleon, with pairs of pleopods, except the last which together with biramous
uropods form a swimming tail pleotelson.
The rather uniform morphology makes identification, even to family and genera, difficult. A key and description of individual genera is provided by Kabata (1985).
Life cycle and biology
The ‘cymothoid’ isopods attach to fish early in life and pass through a male stage before
becoming female. The presence of a mature female prevents male stage specimens
from further development. Both males and females remain permanently attached to the
fish. Egg and larval development takes place in a special brood-pouch on the females
ventrum. Another group, the ‘gnathiid’ isopods, are marine and only parasitic during the
larval stage.
Pathology and epizootiology
Isopoda may be either highly host specific, opportunistic in their choice of hosts or
facultative parasites, attaching to fish if they are weak or trapped in nets or cages. The
host specific ones are the least harmful to their hosts (e.g. Lironeca spp. of African
freshwater fish - Fryer, 1965a,b, 1968b). Even host specific species attached and
causing degenerative changes in the tongue cartilage do not cause clinical harm to the
fish (Kabata, 1970). With the opportunistic and facultative parasites, the degree of
damage to fish varies with the site of attachment and ratio between the isopod and the
host. Their a priori large size is likely to cause pressure damage and abrasions, if
attached to fish skin, and even more so inside the gill chamber. Lesion to the integument
causes hyperplasia, or desquamation and later on dermal inflammation and necrosis.
Infection in the gill chamber often leads to mechanical damage to the lamellar structure
(Paperna & Zwerner, 1976). The parasite can also impede opercular respiratory
movements (Kabata, 1985). The opportunistic Nerocila orbignyi is haematophage, and massive infection led to death of cage cultured Dicentrarchus labrax (Euzet & Raibaut, 1985). There is a record of mortality of farmed milkfish (Chanos chanos) in the Philippines infested with Aliotropus typus (Kabata, 1985).
Members of the genus lchthyoxenus become enclosed in a pouch-like cavity in its host's (various Southeast Asian cyprinids) body wall. The epithelial lining of the pouch wall undergoes degradation and large pouches could exert pressure on the viscera (Kabata, 1985).
Control
Not practised. Organophosphate insecticides used for other piscine crustaceans are
likely to be effective.
REFERENCES
Ahne, W., 1985. Argulus foliaceus L. and Piscicola geometra L. as mechanical vectors of spring viraemia of carp virus (SVCV). J. Fish Dis., 8: 241–242.
Avenant-Oldewage, A., 1991. A new species of Chonopeltis (Crustacea: Branchiura) from the Kruger National Park, southern Africa. J. Afr. Zool., 105: 313–321.
Avenant, A., Loots, G.C. & Van As, J.G., 1989. A redescription of Dolops ranarum (Stuhlmann, 1981) (Crustacea: Branchiura). Sys. Parasitol., 13: 141–151.
Avenant, A.M. & Van As, J.G., 1985. Occurrence and distribution of Dolops ranarum (Stuhlmann, 1891) (Crustacea: Branchiura), ectoparasites of freshwater fish in Transvaal, South Africa. J. Fish Biol., 27: 403–416.
Avenant, A.M. & Van As, J.G., 1986. Observation on the seasonal occurrence of the fish ectoparasite Dolops ranarum (Stuhlmann 1891) (Crustacea: Branchiura) in the Transvaal. S. Afr. J. Wildlife Res., 16: 62–64.
Avenant, A., Van As, J.G. & Loots, G.C., 1989. On the hatching and morphology of Dolops ranarum larvae (Crustacea: Branchiura). J. Zool. Lond., 217: 511–519.
Bauer, O.N., 1959. The ecology of freshwater fish. Inves. Gosud. Nauch. -Issled. Inst. Ozer. Rech. Ryb. Khoz. 49: 5–206 (In Russian, English transl. Israel Prog. Sci. Trans. cat. No. 622, 1962, 3–215).
Ben Hassine, O.-K., 1983. Les copepodes parasites de poisson Mugilide en Mediterranee occidentale (Cote française et tunisiennes). Ph.D. Thesis Université des Sciences et Techniques du Languedoc.
Ben Hassine, O.K. & Raibaut, A., 1981. Realisation experimentale du cycle evolutif de Ergasilus lizae Kroyer, 1863, Copepodite parasite de Poisson Mugilides. Premiers resultat de l'infestation. Archiv de l'Institute Pasteur de Tunis, 58: 423–430.
Ben Hassine, O.K., Braun, M. & Raibaut, A., 1983. Etude comperative de l'infestation de Mugil cephalus Linne, 1758 par le copepode Ergasilus lizae Kroyer, 1863 dans deux lagunes du littoral Mediterraneen Français. Rapp. Comm. Int. Mer Medit., 28: 379–384.
Capart, A., 1944. Note sur les copepodes parasites. 3. Copepodes des poisson d'eau douce du Congo Belge. Bull. Mus. R. Hist. Nat. Belg., 20: 1–24.
Euzet, L. & Raibaut, A., 1985. Les maladies parasitaires en pisciculture marine. Symbioses, 17: 51–68.
Fryer, G., 1956. A report on the parasitic Copepoda and Branchiura of the fish of Lake Nyasa. Proc. Zool. Soc. Lond., 127: 239–344
Fryer, G., 1959. A report on the parasitic Copepoda and Branchiura of the fish of Lake Bangweulu (Northern Rhodesia). Proc. Zool. Soc. Lond., 132: 517–550.
Fryer, G., 1960. Studies on some parasitic crustaceans on African freshwater fish, with a description of a new copepod of the genus Ergasilus and a new branchiuran of the genus Chonopeltis. Proc. Zool. Soc. Lond. 133: 629–647.
Fryer, G., 1961a. Variation and systematic problems in a group of lernaeid copepods. Crustaceana, (2): 275–285.
Fryer, G., 1961b. The parasitic Copepoda and Branchiura of Lake Victoria and the Victoria Nile. Proc. Zool. Soc. Lond., 137: 41–60.
Fryer, G., 1964. Further studies on the parasitic Crustacea of African freshwater fish. Proc. Zool. Soc. Lond., 143: 79–102.
Fryer, G., 1965a. Crustacean parasites of African freshwater fish mostly collected during the expeditions to Lake Tanganyika, and to Lakes Kivu, Edward and Albert by the Institute Royal des Science Naturelle Belgique. Bull. Inst. R. Sci. Nat. Belg., 41: 1–22.
Fryer, G., 1965b. A new isopod of the genus Lironeca parasitic on a cichlid fish of Lake Tanganyika. Rev. Zool. Bot. Afr., 81: 376–384.
Fryer, G., 1965c. Parasitic crustacea of African freshwater fish from the Nile and the Niger systems. Proc. Zool. Soc. Lond. 145: 285–303.
Fryer, G., 1966. Habitat selection and gregarious behaviour in parasitic crustaceans. Crustaceana, (10): 199–209.
Fryer, G., 1968a. The parasitic Crustacea of African freshwater fish: their biology and distribution. J. Zoology London,, 156: 35–43.
Fryer, G., 1968b. A new parasitic isopod of the family Cymothoidae from the clupeid fish of Lake Tanganyika - a further Lake Tanganyika enigma. J. Zool Lond., 156: 35–43.
Fryer, G., 1977. On some species of Chonopeltis (Crustacea, Branchiura) from the rivers of the extreme South-West Cape region of Africa. J. Zool., 182: 441–445.
Fryer, G. & lles, T.D., 1972. The cichlid fishes of the great lakes of Africa. T.F.H. Publications, Neptune City, N.J.
Harding, J.P., 1950. Some species of Lernaea (Crustacea, Copepods: Parasites of freshwater fish). Bull. Brit. Mus. (Nat. Hist.) Zool. 1, No. 1.
Hoffman, G.L., 1967. Parasites of North American Freshwater Fishes. University of California Press, Berkeley & Los Angeles.
Hoffman, G.L., 1970. Intercontinental and transcontinental dissemination and transfaunation of fish parasites with emphasis on whirling disease (Myxosoma cerebralis). In: Snieszko, S.F. (ed.) A Symposium on Diseases of Fish and Shellfish, Amer. Fish. Soc. spec. publ. 5, pp. 69–81.
Hoffman, G.L., 1976. The anchor parasite (Lernaea elegans) and related species (Parasites of freshwater fish. VI. Miscellaneous.). US Department of the Interior, Fish and Wildlife Service, Fish Disease Leaflet 46 (November).
Hoffman, G.L., 1977. Argulus, a branchiuran parasite of freshwater fish. United States Department of the Interior, Fish Disease Leaflet 49.
Humes, A.G., 1957. Two new caligoid copepods from Egyptian fish. J. Parasitol., 43: 201–208.
Jones, J.B. & Hine, P.M., 1978. A new species of Mugicola parasitic on South African elvers (Copepoda, Therodamasidae). Zool. Afr., 13: 213–219.
Kabata, Z., 1970. Crustacea as enemies of fish. Book 1, Snieszko, S., Axelrod H.R. (ed.), Diseases of Fish, T.F.H. Publications, Jersey City, N.J.
Kabata, Z., 1985. Parasites and Diseases of Fish Cultured in the Tropics. Taylor & Francis.
Kruger, I., Van As, J.G. & Saayman, J.E., 1983. Observation on the occurrence of the fish louse Argulus japonicus Thiele, 1900 in western Transvaal. S. Afr. J. Zool., 18: 408–410.
Lahav, M. & Sarig, S., 1964. Observation on the biology of Lernaea cyprinacea L. in fish ponds in Israel. Bull. Fish Cult. Israel (Bamidgeh), 16: 77–86.
Lahav, M. & Sarig, S., 1968. Ergasilus sieboldi Nordmann infestation of grey mullet in Israel's fish ponds. Bull. Fish Cult. Israel (Bamidgeh), 19: 69–80.
Lahav, M., Sarig, S. & Shilo, M., 1964. The eradication of Lernaea in storage ponds of carp through destruction of the copepodid stage by Diptertex. Bull. Fish Cult. Israel (Bamidgeh), 16: 87–94.
Lahav, M., Shilo, M. & Sarig, S., 1962. Development of resistance to lindane in Argulus population of fish ponds. Bull. Fish Cult. Israel (Bamidgeh), 14: 67–76.
Landsberg, J.H., 1989. Parasites and associated diseases of fish in warm water culture with special emphasis on intensification. In: Shilo, M. & Sarig, S. (ed.) Fish Culture in Warm Water Systems: Problems and Trends. CRC Press Inc. Boca Raton, Flo. pp. 195–252.
Mbahinzireki, G.B., 1980. Observation on some common parasites of Bagrus docmac Forskahl (Pisces: Siluroidea) of Lake Victoria. Hydrobiol., 75: 273–280.
Paperna, I., 1964a. The metazoan parasite fauna of Israel inland water fishes. Bamidgeh (Bull. Fish Cult. Israel) 16: 3–66.
Paperna, I., 1964b. Parasitic Crustacea (Copepoda and Branchiura) from inland water fish of Israel. Israel J. Zool., 13: 58–68.
Paperna, I., 1968. Ectoparasitic infections of fish of Volta lake, Ghana. Bull. Wildl. Dis. Ass., 4: 135–137.
Paperna, I., 1969. Parasitic Crustacea from fish of the Volta Basin and South Ghana. Rev. Zool. Bot. Afr., 80: 208–216.
Paperna, I., 1991. Diseases caused by parasites in the aquaculture of warm water fish. Annual Rev. Fish Dis., 1: 155–194.
Paperna, I. & Lahav, M., 1971. New records and further data on fish parasites in Israel. Bull. Fish Cult. Israel (Bamidgeh), 16: 77–86.
Paperna, I. & Overstreet, R.M. 1981. Parasites and diseases of Mullets (Mugilidae). In: Oren, O.H. (ed.) Aquaculture of Grey Mullets. IBP 26, Cambridge University Press, U.K.
Paperna, I. & Thurston, J.P., 1968. Report on ectoparasitic infections of freshwater fish in Africa. Bull. Off. int. Epizoot., 69: 1192–1206.
Paperna, I. & Zwerner, D.E., 1976. Parasites and diseases of striped bass, Morone saxatilis (Walbaum) from the lower Chesapeake bay. J. Fish Biol. 9: 267–287.
Paperna, I. & Zwerner, D.E., 1981. Host-parasite relationship of Ergasilus labrachis Kroyer (Cyclopidea, Ergasilidae) and the striped bass, Morone saxatilis (Walbaum) from the lower Chesapeake bay. Ann. Parasitol. Hum. Comp., 57: 393–405.
Pauly, D., 1974. On some features of the infestation of the mouth-breeding fish Tilapia melanotheron Ruppel, 1852 by the parasitic copepod Paeonodes lagunaris van Banning. Beaufortia (Zool. Mus. Uni. Amsterdam), 22 (No. 287): 9–15.
Raibaut, A., 1985. Les cycle evolutifs des copepodes parasites et les modalites de l'infestation. Ann Biol., 25: 233–274.
Raibaut, A., Ben Hassine, O.K. & Prunus, G., 1975. Etude de l'infestation de Mugil (Mugil) cephalus Linne 1758 (Poisson, Teleosteens, Mugilidae) par le copepode Ergasilus nanus Van Beneden, 1870 dans le Lac Ischkeul (Tunisie). Bull. Soc. Zool. France, 100: 427–437.
Roberts, L., 1970. Ergasilus (Copepoda: Cyclopoidea): revision and key to species of North America. Trans. Am. Microsc. Soc., 89: 134–161.
Sarig, S., 1971. The prevention and treatment of diseases of warmwater fish under subtropical conditions, with special emphasis on intensive fish farming. T.F.H. Publications Inc., Jersey City, N.J. 127 p.
Shafir, A. & Van As, J.G., 1986. Laying, development and hatching of eggs of the fish ectoparasite Argulus japonicus (Crustacea: Branchiura). J. Zool. Lond., 210: 401–414.
Shariff, M., 1981. The histopathology of the eye of bighead carp, Aristichthys nobilis (Richardson), infested with Lernaea piscinae Harding, 1950. J. Fish Dis., 4: 161–168.
Shields, R.J. & Goode, R.P., 1978. Host rejection of Lernaea cyprinacea L. (Copepoda). Crustaceana, 35: 301–307.
Shimura, S., Inoue, K., Kudo, M. & Eugusa, S., 1983. Studies on effects of parasitism of Argulus coregoni (Crustacea: Branchiura) on furunculosis of Onchorhynchus masou (Salmonidae). Fish Pathol., 18: 37–40.
Thurston, J.P., 1969. The biology of Lernaea barnimiana (Crustacea, Copepoda) from Lake George, Uganda. Rev. Zool. Bot. Afr., 80: 15–33.
Thurston, J.P., 1970. The incidence of Monogenea and parasitic Crustacea on the gills of fish in Uganda. Rev. Zool. Bot. Afr., 82: 113–130.
Trilles, J.P., 1965. Specificite parasitaire chez les isopodes Cymothoidae mediteraneens. Note preliminaire. Vie et Milieu, 15: 105–116.
Van As, J.G., 1986. A new species of Chonopeltis (Crustacea: Branchiura) from the Limpopo system, southern Africa. S. Afr. J. Zool., 21: 348–351.
Van As, J.G. & Basson, L., 1984. Checklist of freshwater fish parasites from southern Africa. S. Afr. J. Wildl., 14: 49–61.
van Banning, P., 1974. A new species of Paeonodes (Therodamasidae, Cyclopoida) a parasitic copepod of the fish Tilapia melanotheron from the Sakumo lagoon, Ghana, Africa. Beaufortia (Zool. Mus. Univ. Amsterdam), 22 (No. 286): 1–7.
Wunder, W., 1962. Nerocila orbignyi ein proterandrischer hermaphrodit und parasitischer isopode auf Tilapia galilaea im see Borullus in Aegypten. Zool Anz., (Suppl.1) 25: 140–151.
Yashuv, V.A., 1959. On the biology of Lernaea cyprinacea in fish ponds. Bull. Fish Cult. Israel (Bamidgeh), 11: 80–89.
Yashuv, V.A., 1972. Efficacy of mullet growth in fish ponds. Bull. Fish Cult. Israel (Bamidgeh), 24: 12–25.
ILLUSTRATIONS
Plate 27. Parasitic Crustacea: (p. 199) a. Heavy Ergasilus lizae infection of gills of pond reared Mugil cephalus, Israel (courtesy S. Sarig). b. Carp tail with heavy Lernaea cyprinacea infection, Israel. c. L. hardingi attached to tail of Oreochromis niloticus, Uganda. d. Released adult female L. cyprinacea (courtesy L. Basson). e. Haemmorhagic lesions around L. cyprinacea attached to carp skin. f. Heavy invasion of Barbus holubi by Argulus japonicus, Transvaal, South Africa (courtesy L. Basson).
Plate 28. Parasitic Crustacea continued: (p. 200) g. Argulus africanus from Bagrus docmac, northern L. Victoria (SEM × 64). h. Dolops ranarum, of same host and locality as a (SEM × 47). i. Eggs of Argulus japonicus (live, by courtesy of S. Sarig). j. Eggs of Dolops ranarum (live, by courtesy of A. Avenant-Oldewage). k. A. japonicus from farmed carp, Israel (live, by courtesy of S. Sarig). I. D. ranarum inside the mouth of Clarias gariepinus, from Transvaal, South Africa (by courtesy of L. Basson). m. Histological section of Bagrus docmac skin at D. ranarum attachment area showing proliferation and displacement of the epithelium and haemmorhagic inflammation in the dermal layer.
Plate 29. Parasitic Crustacea continued: page 201 with legend.
Fig. 8. Parasitic Copepoda: (p. 202) A. Ergasilus lizae (1.1 mm. long; after Ben Hassine, 1983). B. antennae of Ergasilus cunningtoni (0.97 mm, after Capart, 1944). C. Ergasilus kandti (0.7 mm long; after Capart, 1944). D. Ergasilus latus (from cichlid fishes) (0.9 mm long, after Fryer, 1960). E. Paraergasilus minutus (from cichlid fishes) (0.35–0.45 mm long, after Fryer, 1956). F. Afrolernaea longicollis, whole (15 mm long) and head (after Fryer, 1964). G. Lernaeagiraffa heterodicola (19 mm long, after Fryer 1965a). H. Dysphorus torquatus (15 mm long, after Fryer, 1965c) I. Lamproglena intercedens (5 mm long, after Fryer, 1965c). J. Lamproglena monodi (3.4 mm long, after Capart, 1944). K. Paeonodes lagunaris (4.6–6.8 mm long, after Van Banning, 1974). L. Achtheres micropteri (2–7 mm long, after Hoffman, 1967).
Fig. 9. Parasitic Copepoda continued and Branchiura: (p. 203) A. Lernaea barnimiana (9 mm long). B. same, with a-symmetric anchor (after Fryer, 1961a). C,D. Anchors of Lernaea cyprinacea from jaws of Oreochromis spp. from Lake Victoria (after Fryer, 1961). E. Lernaea hardingi (10 mm long). F. Opistholernaea laterobranchialis (14 mm long, after Fryer, 1959). G. Lernaea palati (8 mm long, after Fryer, 1956). H. Lernaea lophiara anchors (after Harding, 1950). I. Lernaea barili, anchors (after Harding, 1950). J. Development of Lernaea cyprinacea (from common carp, Israel): 1–2. Nauplii; 3. free copepodite; 4–6. Copepodites attached to gills. 8. Premature males and females; 9. Mature male; 10. Early metamorphosing female. K. Sciaenophilus pharaonis (12 mm long, after Humes, 1957). L,M. Chonopeltis brevis (8.5 mm long, after Fryer, 1960). N,O. Argulus brachypeltis, 9.9 mm long, after Fryer, 1965c).
Fig. 10. Parasitic Isopoda: page 204 with legend.
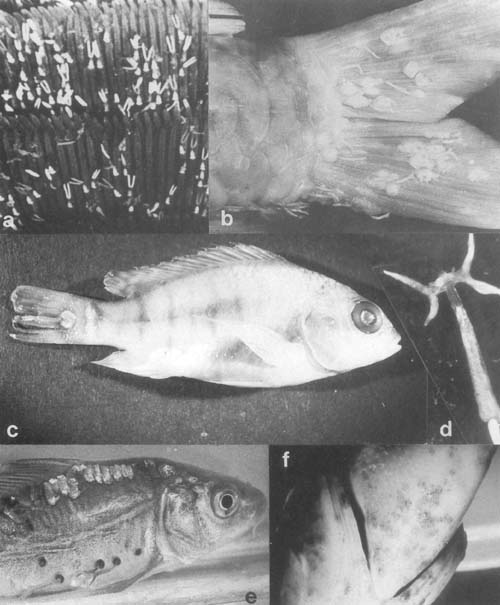
Plate 27. Parasitic Crustacea (legend p. 198).
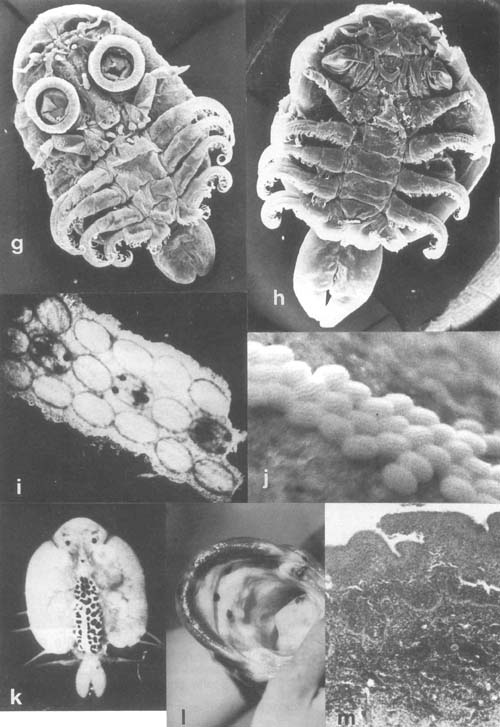
Plate 28. Parasitic Crustacea continued (legend p. 198).
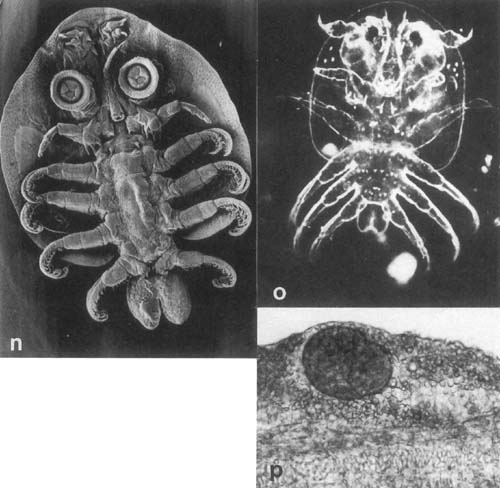
Plate 29. Parasitic Crustacea continued n. Argulus japonicus from C. gariepinus (SEM × 50). o. Newly hatched A. japonicus (live, by courtesy of S. Sarig). p. Unionid glochidium (Mollusca) in gills of O. aureus, L. Kinneret.
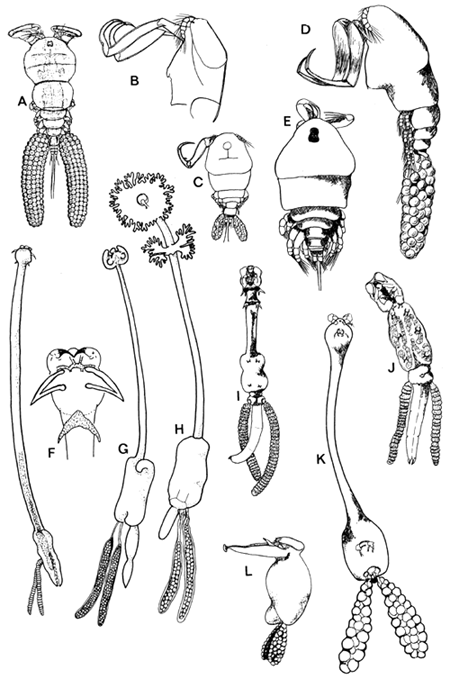
Fig. 8. Parasitic Copepoda (legend p. 198).

Fig. 9. Parasitic Copepoda continued and Branchiura (legend p. 198).
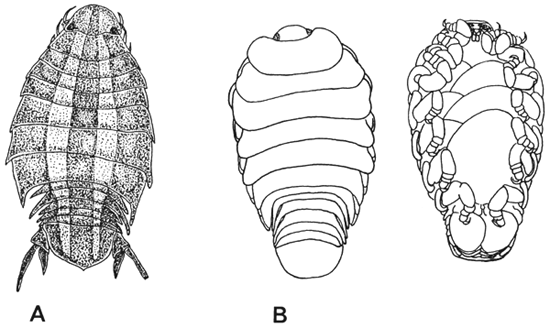
Fig. 10. Parasitic Isopoda: A. Nerocila orbignyi (8.5 mm long, by courtesy of P. Oreccia). B. Lironeca tanganyikae (12mm long, after Fryer, 1965b).
![]()
![]()
![]()