![]()
![]()
![]()
Como ha sido expuesto en el primer Capítulo, en condiciones de acceso abierto fracasa la óptima asignación de recursos pesqueros, debido entre otros factores, a la ocurrencia de altos costos de exclusión. El esfuerzo pesquero ejercido por una embarcación se ve influenciado por la forma en que otras hagan uso del recurso, de tal manera que la entrada de más embarcaciones incrementará el esfuerzo de pesca sobre éste, afectando los rendimientos de los diferentes usuarios de la pesquería. Para realizar estimaciones y predicciones de los posibles impactos bioeconómicos en pesquerías sujetas a diferentes regímenes de explotación, se hace necesario modelar matemáticamente estos sistemas dinámicos. Para ello se requiere conocer los fundamentos de la teoría bioeconómica de pesquerías, los cuales permitirán explorar e introducir las complejidades inherentes al manejo de recursos pesqueros en los siguientes Capítulos.
El presente Capítulo se centrará en el desarrollo del modelo bioeconómico clásico de Gordon (1954), también llamado Gordon-Schaefer, porque al igual que el de este último, parte del modelo logístico de crecimiento poblacional de Verhulst (1838). En la primera sección se presenta el modelo logístico, exponiendo algunas de sus limitaciones en especies marinas. En la segunda sección se detalla el modelo de rendimiento excedente propuesto por Schaefer (1954) en términos de biomasa y de captura por unidad de esfuerzo; se discuten sus supuestos y se presentan algunas variantes propuestas por otros autores. En la tercera sección se presenta el modelo bioeconómico de Gordon (1954) en términos de captura y esfuerzo, y en función de oferta y demanda, tanto en su versión estática como dinámica. Se plantean sus supuestos y limitaciones, así como modificaciones desarrolladas por otros autores. En la cuarta sección se desarrolla el modelo dinámico propuesto por Smith (1969), el cual condiciona el comportamiento de largo plazo de una flota pesquera a variaciones en los ingresos netos. En la quinta sección se presentan los modelos de producción excedente basados en mortalidad, en sus versiones biológica y bioeconómica. Por último, en la sexta sección se presenta el análisis bioeconómico de modelos que consideran la estructura por edades de una población, tanto en la versión clásica estática (Beverton & Holt, 1957) como dinámica (Seijo & Defeo, 1994b).
La ecuación logistica propuesta por Verhulst (1838) describe el crecimiento de una población en base a la siguiente expresión matemática (Graham, 1938):

donde:
r = tasa intrínseca de crecimiento poblacional. B(t) = biomasa de la población en el tiempo t. K = capacidad de carga del ambiente.
El comportamiento de una población es descrito por una curva sigmoide, donde la biomasa no explotada se incrementará hasta un nivel máximo equivalente a la población virgen B∞, determinado por K (Pitcher & Hart, 1982). Este nivel de equilibrio puede ser definido solamente en el contexto de la producción básica del medio, de la eficiencia relativa del crecimiento de la población, de su conversión de alimento y por tanto de la abundancia de los componentes que incluye el ecosistema (Caddy & Sharp, 1986). La población se mantendrá en equilibrio en la medida que los factores que hacen decrecer la biomasa tales como la depredación, enfermedades, etc., sean balanceados por aquellos que la aumentan, es decir el crecimiento individual y el reclutamiento. La tasa de crecimiento poblacional (dB/dt) correspondiente a cada nivel de biomasa B(t) del recurso, es presentada en la Figura 2.1a para una población hipotética cuya dinámica es descrita por una capacidad de carga de 3.5 millones de toneladas y una tasa intrinseca de crecimiento de 0.36. En las Figuras 2.1b y 2.1c se puede observar la curva sigmoide y el crecimiento en biomasa por unidad de tiempo, respectivamente.

Figura 2.1 Función de crecimiento logístico de la biomasa de un recurso para una capacidad de carga de 3.5 millones de toneladas y una tasa intrínseca de crecimiento de 0.36.
El crecimiento de una población determinará una progresiva incidencia de fenómenos de denso-dependencia a niveles cercanos a la capacidad de carga del sistema. Esto variará de acuerdo a la estrategia de vida de cada especie y al tipo de ecosistema considerado. Ciertas especies tienden a ocupar rápidamente nichos vacantes en un ecosistema, mientras que otras están mejor adaptadas a un crecimiento poblacional lento bajo condiciones de una alta competencia por recursos limitantes (Pianka, 1972). Las primeras poseen una mayor susceptibilidad a factores extrinsecos o ambientales, debido a que tienden a habitar ambientes usualmente variables e impredecibles, y en consecuencia rara vez alcanzan tamaños poblacionales cercanos a la capacidad de carga del ambiente. Las segundas habitan sistemas estables o predecibles, el tamaño poblacional es poco variable en el tiempo y está limitado por procesos de competencia intra e interespecífica en comunidades usualmente saturadas (Pianka, 1972). En el caso de especies marinas, su ubicación dentro de un grupo u otro resulta difícil, dado que la complejidad de sus ciclos de vida parece ubicarlos en ocasiones dentro de un grupo y en otro período dentro de otro (Rothschild, 1986). Tal es el caso de especies con ciclos de vida complejos, que incluyen una fase dispersiva planctónica y una adulta bentónica; durante el estadío larval se encuentran expuestas de manera más marcada a efectos ambientales que en su etapa adulta, en la cual los efectos denso-dependientes pueden ocurrir en forma más intensa.
Bajo condiciones de explotación, Schaefer (1954) introduce el término de tasa de captura C(t), la cual se define como:
C(t) = qf (t)B(t) (2.2)
donde la captura depende del esfuerzo de pesca f(t) y de la biomasa poblacional B(t) en el tiempo t; y q representa el coeficiente de capturabilidad, definido como la fracción de la población que es extraída por una unidad de esfuerzo (Gulland, 1983).
La ecuación 2.2 implica que la misma captura puede obtenerse a diferentes niveles de esfuerzo y biomasa. La limitante en este sentido es que q, considerado constante para un período de tiempo determinado, no puede obtenerse sin una estimación independiente de la biomasa.
En el caso de pesquerías uniespecificas donde se emplea un mismo tipo de arte, el concepto de q ha sido aplicado de manera más común, debido a que el esfuerzo puede ser estandarizado. En regiones tropicales, donde existe una alta diversidad de artes y especies, es dificil englobar en este coeficiente las complejidades que se presentan en el esfuerzo (Arceo & Seijo, 1991a). En estos casos se ha propuesto el uso de funciones de producción que den mejores aproximaciones (e.g. Prochaska & Cato, 1980; Taylor & Prochaska, 1985), dado que existen una serie de variables asociadas al esfuerzo que pueden intervenir en la producción (Staniford, 1987; Arceo, 1991; Arceo & Seijo, 1991b). Agnello & Anderson (1983) abordan la relación captura - esfuerzo en función de la eficiencia de artes de pesca, utilizando una función tipo Cobb-Douglas. A efectos del desarrollo del modelo de Schaefer, se seguirá manejando el concepto de q, no obstante las limitaciones antes mencionadas. Un análisis detallado de las variaciones de esta variable en función de las características de los recursos y de los artes de pesca empleados, es detallado en Caddy (1979a), Arreguín-Sánchez (1992) y Hilborn & Walters (1992).
Considerando una población explotada, los cambios en biomasa se representarán de acuerdo a la siguiente expresión:

Cuando la población se encuentra en condiciones de equilibrio, la tasa de incremento poblacional repone las pérdidas debidas a la mortalidad, y por tanto no hay cambios de la biomasa en el tiempo (e.g. dB/dt=0); el rendimiento en equilibrio puede definirse como:

Entonces, la biomasa de la población en equilibrio (Be) en función del esfuerzo de pesca se define como:

Para un nivel dado de esfuerzo, la población en equilibrio (Be) alcanzará un nivel donde la tasa de captura es igual a la de crecimiento. Por tanto, para cada nivel de esfuerzo existirá un nivel de población en equilibrio (Anderson, 1986). Como resultado se tiene una relación inversa entre la población en equilibrio y el esfuerzo pesquero (Fig.2.2a). Este comportamiento se puede observar a través de la captura por unidad de esfuerzo (CPUE), considerándose en este caso como un indice relativo de la abundancia de la biomasa de la población en el tiempo (Fig.2.2c).
Para obtener la función que relaciona la captura con el esfuerzo bajo condiciones de equilibrio, se sustituye la ecuación (2.7) en (2.2):

La función (2.8) se representa mediante una parábola, donde la captura (C) para un esfuerzo (f) dado en una población en equilibrio es denominado rendimiento sostenible, dado que las pérdidas por mortalidad natural y por pesca son compensadas por un incremento poblacional debido a los procesos de crecimiento individual y reclutamiento; en consecuencia el rendimiento de la pesquería se puede mantener en el tiempo. Esta curva puede ser considerada como la función de producción de la pesqueria en el largo plazo (Fig. 2.2b), donde para un nivel dado de esfuerzo f1, se obtiene un rendimiento C1, el cual será mayor a medida que el primero incremente hasta un nivel de máximo rendimiento (MRS) correspondiente a un nivel de esfuerzo f2, después del cual decrece si el esfuerzo aumenta. En este punto, la población ya no podrá reponer las pérdidas por mortalidad, y a medida que la presión de pesca aumente, la captura tenderá a disminuir debido al decremento en la biomasa de la población (C3) a un nivel de esfuerzo f3.
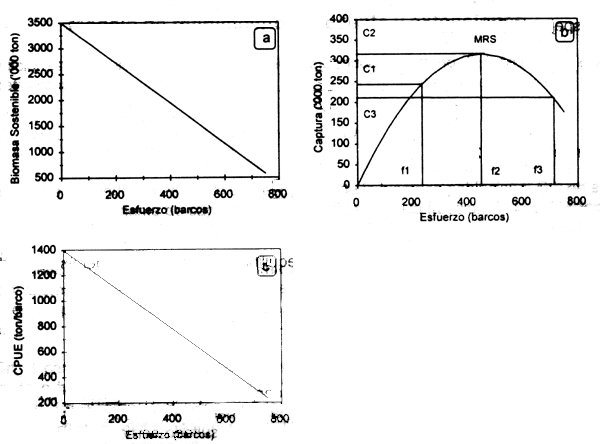
Figura 2.2 Curvas de biomasa en equilibrio (a) rendimiento sostenible (b) y CPUE (c) basados en el modelo de Schaefer.
a) Existen condiciones de equilibrio poblacional, mediante el cual se compensan los procesos de mortalidad con los de crecimiento y reclutamiento.
b) Para obtener el rendimiento de la población sujeta a explotación asumiendo condiciones de equilibrio, se parte de que la mortalidad por pesca F es directamente proporcional al esfuerzo f, siendo q la constante de proporcionalidad, tal que:
F=qf (2.9)
c) La CPUE es directamente proporcional a la abundancia de la población, o sea:

d) El recurso se mantiene en un ambiente fisico estable determinado por una capacidad de carga constante.
e) El recurso lo compone una sola especie, y por tanto el esfuerzo es aplicado exclusivamente a ésta.
f) La tecnología utilizada en la extracción se mantiene constante.
(i) El inciso (a) asume equilibrio, lo cual constituye un serio inconveniente a la hora de administrar los recursos (Caddy, 1984). Ello implica que se dejan de lado todos los procesos que gobiernan la productividad de un stock (e.g.reclutamiento), muchos de los cuales poseen un alto grado de estocasticidad inclusive en una escala de tiempo reducida.
(ii) El coeficiente de capturabilidad q no siempre puede ser considerado constante. Diferentes comportamientos de agregación en cardúmenes de recursos pelágicos o en recursos bentónicos. así como factores relacionados con la diferente selectividad y accesibilidad por clases de talla al arte de pesca, violan dicho supuesto.
(iii) La CPUE no siempre es un índice insesgado de abundancia relativa de una población como se asume en el inciso (c), en especial en recursos sedentarios donde la distribución espacial no es homogénea. La limitada movilidad de estos organismos implica que no puedan redistribuirse en el área de pesca una vez aplicado el esfuerzo pesquero. La estrategia dirigida de exterminación de parches determina un comportamiento agregado de los usuarios del recurso, lo cual limita la aplicabilidad del modelo (ver Caddy, 1975, 1989a, b; Conan, 1984; Orensanz et al., 1991 para revisiones detalladas acerca del tópico).
(iv) El asumir que se conoce el tamaño de la población resulta riesgoso, debido a la dificultad en la obtención de estimadores insesgados de tamaño poblacional, así como a las fuentes de variabilidad que pudieran producir intensas fluctuaciones en su magnitud.
(v) En el inciso (d) se ignora la dinámica de la distribución espacial del recurso, los procesos biológicos que generan la biomasa, así como las interacciones intra e interespecíficas y los efectos de las variaciones estocásticas en el medio ambiente y en la abundancia de la población. Al suponer que el recurso se mantiene en un ambiente fisico estable que determina una capacidad de carga constante, se limita la aplicabilidad de este modelo determinístico (Pitcher & Hart, 1982; Sissenwine, 1984a, b).
(vi) El supuesto (e) ignora la ocurrencia de interdependencias biológicas (e.g. competencia, depredación) y tecnológicas: ver Capitulo 3. Además, no considera la asignación diferencial del esfuerzo en el corto plazo, esto es, cómo el pescador decide entre la captura de un recurso u otro, cuando el método y el arte de pesca le permiten tener más de una especie objetivo, así como definir cuándo y dónde salir a pescar (ver Hilborn, 1985; Eales & Wilen, 1986; Sampson, 1993).
(vii) Por otro lado, incrementos en el poder de pesca (tecnificación de la captura) por parte de la flota determinan variaciones en q que restringen la aplicabilidad del modelo y limitan el supuesto (f).
El modelo de Schaefer requiere información sobre la biomasa, tasa de crecimiento poblacional y capacidad de carga del ambiente, cuya obtención en la práctica es sumamente dificil. A efectos de mitigar este problema, una estimación alternativa del rendimiento sostenible es efectuada en términos de CPUE. Aún cuando esta información es más fácil de adquirir, en el caso de pesquerías multiespecificas (sobre todo en áreas tropicales), se hace dificil el discernimiento del esfuerzo efectivo aplicado a cada especie. Por otra parte, las estadísticas de esfuerzo en la mayoría de los casos corresponden más al esfuerzo nominal que al efectivo. Sin embargo, esta información es más accesible que aquella específica de la población (e.g. composición por edades), razón por la cual los modelos globales se han utilizado ampliamente.
La ecuación (2.10) asume proporcionalidad entre la CPUE y la biomasa en condiciones de equilibrio. Si se define CPUE como U (notación generalizada en los modelos globales) se tiene que:

Este nuevo valor de la biomasa es sustituido en el modelo para obtener el rendimiento en equilibrio (Re):

o sea:
Re=r(U/p2)(U∞-U)
Reordenando para resolver en términos de CPUE en equilibrio (Ue):
Ue=Uinfin-(U∞ q/r)f (2.13)
donde (Uxq/r) es la pendiente (b) de la recta y Ux es la ordenada al origen.
En términos de esfuerzo, la función puede ser definida como sigue:
Ue=U∞-f-bf' (2.14)
y derivando la ecuación anterior con respecto al esfuerzo e igualándola a cero, se obtiene el esfuerzo para el MRS:

Como el nivel dado para el rendimiento sostenible ocurre a la mitad de la máxima biomasa (UMRS=Ux/2), el MRS se obtiene de la siguiente forma:

Se han desarrollado diversas alternativas al modelo de rendimiento de Schaefer. Una de las variaciones comúnmente usadas es el modelo exponencial propuesto por Fox (1970), quien usa la curva de crecimiento de Gompertz (Pitcher & Hart, 1982), la cual muestra una asimetría que es más característica de crecimiento en peso. En este caso la población en equilibrio disminuye más lentamente con respecto al esfuerzo aplicado, obteniéndose una función exponencial a diferencia de la lineal propuesta por Schaefer en términos de Ue. Los cambios en la biomasa en función del peso bajo condiciones de equilibrio, serán definidos por la ecuación (2.17).
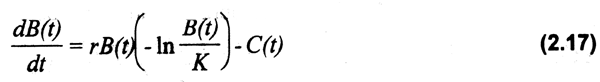
Mientras que en la curva logistica del modelo de Schaefer el MRS se alcanza a un nivel de K/2, en el de Fox corresponde a rK/q.
En este caso sólo se presenta la derivación del modelo de Schaefer (1954), el cual es utilizado como base para el modelo bioeconómico de Gordon. Para las variantes del modelo ver Fox (1970), Pella & Tomlinson (1969), Pope (1979) y Pitcher & Hart (1982).
La curva de rendimiento sostenible en función del esfuerzo es la base del modelo económico de Gordon (1954), quien deriva la función de rendimiento incorporando el componente económico. Aborda el problema de los recursos de propiedad común y desarrolla la teoria de acceso abierto. Su principal mérito ha sido clarificar en términos económicos el concepto de “sobrepesca” en pesquerias no reguladas.
El modelo establece que los ingresos netos π derivados de la actividad pesquera están en función de los ingresos totales sostenibles (ITS) y los costos totales (CT):
π=ITS − CT (2.18)
los cuales pueden expresarse de la siguiente forma:
π= pC &minus - cf (2.19)
siendo p el precio (constante) de la especie capturada y c los costos (constantes) por unidad de esfuerzo. Estos últimos incluyen costos fijos, variables y de oportunidad, tanto de capital como de mano de obra. Los costos fijos son independientes de que la flota opere o no (depreciación, intereses, gastos administrativos, etc.), mientras que los variables sólo se producen cuando el pescador sale a pescar (combustible, carnada, alimentos etc). Los costos de oprtunidad se definen en base a la siguiente mejor alternativa de inversión del capital o bien de ocupación de la mano de obra. Esto implica que los ingresos que hubieran podido ser generados en la siguiente mejor actividad alternativa deben ser incluidos dentro de los costos. El dueño de una embarcación desea obtener el mayor beneficio como resultado de cualquier tipo de inversión, y por tanto, puede decidir entre invertir en la pesca, en el banco o en otro proyecto que sea la siguiente mejor alternativa. Lo mismo sucede con respecto a su mano de obra, ya que podría dedicarla a otra actividad productiva. Asi, esperará que al invertir en la pesca ésta le reditúe al menos lo que pudo haber ganado en su siguiente mejor opción de inversión de capital o de mano de obra.
Ahora bien, si se sustituye la función correspondiente a la tasa de captura (2.2) en (2.19), los ingresos netos en función del esfuerzo serán definidos como:
π= (pqB &minus - c)f (2.20)
Al igual que en el modelo biológico, Gordon (1954) supone condiciones de equilibrio en la población para obtener un rendimiento sostenible en el largo plazo. Considera asimismo que en equilibrio económico los ITS igualan a los CT y por tanto π(t) es igual a cero, no existiendo estimulo para entrar o salir de la pesqueria. Si adicionalmente la biomasa de la expresión (2.20) se asume en equilibrio biológico para el nivel de esfuerzo correspondiente (la derivada de la biomasa con respecto al tiempo es igual a cero), entonces se presentan condiciones de equilibrio biológico y económico simultáneamente, razón por la cual se le denomina equilibrio bioeconómico. La biomasa en equilibrio bioeconómico (BEBE) se define igualando a cero la ecuación (2.20) y despejando B:

Bajo estas condiciones la biomasa será siempre mayor a cero dado que el esfuerzo se reducirá o cesará cuando los costos totales superen a los ingresos totales. En consecuencia, el modelo pronostica: (1) sobreexplotación si la curva de costos totales intersecta a la de ingresos totales sostenibles con niveles de esfuerzo pesquero superiores a los requeridos para operar en MRS; y (2) no extinción del recurso, ya que antes que éste se agote deja de ser rentable ejercer esfuerzo sobre la especie en cuestión. La condición de no extinción dependerá del tipo de crecimiento asumido para el recurso y la forma que adquiera la función de captura definida en 2.2 (ver Clark, 1985; Anderson, 1986). Asimismo, debe precisarse que la predicción de no extinción será correcta si y sólo si la biomasa resultante en equilibrio bioeconómico excede el nivel de biomasa umbral requerido para darle viabilidad de existencia a la población.
Los ingresos totales sostenibles (bajo condiciones de equilibrio) se obtienen al multiplicar la función de biomasa (2.4) por el precio p:

La curva de ingresos totales sostenibles en función del esfuerzo tendrá la misma forma que la de rendimiento sostenible de Schaefer (Fig.2.1) pero en términos monetarios. Los costos totales se obtienen partiendo de la misma función que en el caso anterior, pero en términos del esfuerzo pesquero (f):

Despejando f y multiplicando por el costo unitario (c) se definen los costos totales en función del esfuerzo en el largo plazo:

En las Figuras 2.3a y b se presentan los niveles de biomasa en equilibrio y rendimiento sostenible, que resultarian de los niveles de esfuerzo en máximo rendimiento económico (fMRE), en máximo rendimiento sostenible (fMRS) y equilibrio bioeconómico (fMRE).
La expresión gráfica del modelo de Gordon en términos de ITS y CT se presenta en la Figura 2.3c. En ausencia de regulación, el aumento del esfuerzo ocasiona un incremento en los ingresos hasta alcanzar el nivel de equilibrio bioeconómico, que corresponde a un nivel de esfuerzo (fEBE) donde los costos totales igualan a los ingresos totales sostenibles, en este caso es igual el esfuerzo requerido para alcanzar el MRS. El equilibrio económico indica que no existe incentivo para entrar o salir de la pesqueria, y el biológico, que la población se mantiene en el tiempo. El máximo rendimiento económico (MRE) se alcanza en fMRE donde las diferencias entre los CT y ITS se maximizan. El área bajo la curva de ITS por encima de CT corresponde a los beneficios netos o renta económica, los cuales siempre serán mayores hacia la izquierda del MRS. En este sentido, la posición de la curva de costos es muy importante, ya que si éstos se reducen existirán ganancias (aunque con menores rendimientos) y por tanto habrá estimulos para incrementar el esfuerzo hasta alcanzar un nuevo nivel de equilibrio, ateniéndose también un nuevo nivel de esfuerzo para obtener el máximo rendimiento económico. Se asume que el esfuerzo extra es producido por la operación de botes nuevos que operan de una manera eficiente, más que por la expansión del esfuerzo de los ya existentes (Anderson, 1986). Gordon (1954) pronostica como se demostrará posteriormente, que el esfuerzo en el equilibrio bioeconómico será el doble del ejercido al nivel de máximo rendimiento económico.
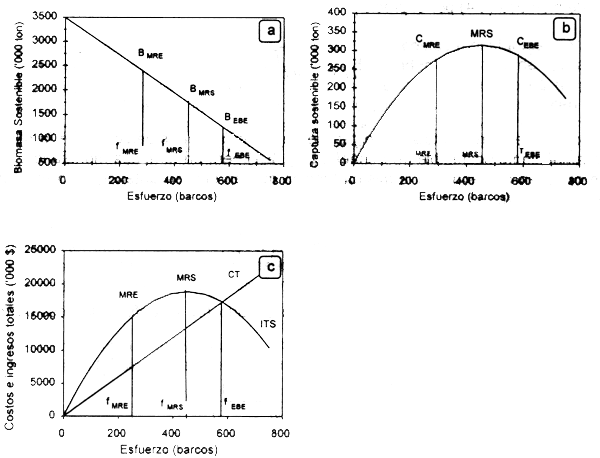
Figura 2.3 Curvas de biomasa sostenible (a), captura sostenible (b), e ingresos totales sostenibles (ITS) y costos totales (CT) (c) del modelo bioeconómico estático de Gordon-Schaefer.
El análisis presentado anteriormente muestra a la pesqueria como un todo. Tanto los rendimientos promedio (Rp) como los rendimientos marginales (Rm), que corresponden a la aplicación de una unidad adicional de esfuerzo, serán sostenibles por unidad de tiempo. Los primeros caerán continuamente conforme aumenta el esfuerzo hasta llegar a cero (Fig.2.4a), es decir, cuando se agota el recurso. El Rm decrece iguálmente hasta llegar a cero en el nivel de MRS, después del cual se hace negativo (Fig.2.4a). Esto es, el rendimiento obtenido por cada unidad adicional de esfuerzo será progresivamente inferior al anterior. Esto resulta como consecuencia de que el pescador individual considera exclusivamente sus rendimientos, sin tomar en cuenta el efecto que el aumento en su esfuerzo pesquero tiene sobre otros usuarios, y por tanto opera siempre en términos de sus rendimientos promedio.
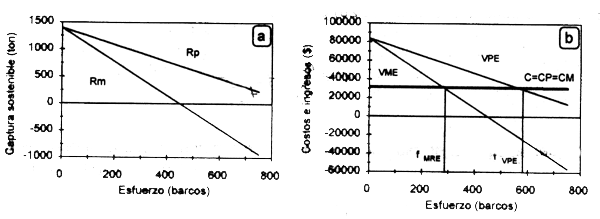
Figura 2.4 Capturas promedio y marginales sostenibles (a), e ingresos, costos promedio y marginales (b) en función del esfuerzo bajo condiciones de acceso abierto.
Cuando se piensa en la implementación de medidas de manejo, una de las metas en términos biológicos consiste en mantener en el tiempo los niveles de esfuerzo y capturas en el MRS. Para ello, se parte del modelo estático de Schaefer, hallando la primera derivada de la función de caputa (2.8) e igualándola a cero:

Despejando f y reordenando términos se obtiene el nivel de esfuerzo para MRS:

El MRS se obtiene al sustituir (2.28) en (2.8):

Para maximizar la renta económica que el recurso le podria generar a la sociedad, es necesario que la pesqueria opere con un nivel de esfuerzo pesquero que maximice el rendimiento económico (e.g. esfuerzo en MRE). Este criterio se obtendrá al maximizarse las diferencias entre los ingresos totales sostenibles y los costos totales del esfuerzo pesquero. En este caso, se obtiene cuando el valor del producto marginal del esfuerzo pesquero (VME) es igual a los costos por unidad de esfuerzo, e.g. VME=c. Considerando la tasa de captura expresada en la ecuación (2.2), se sustituye la expresión que determina la biomasa en función del esfuerzo pesquero:
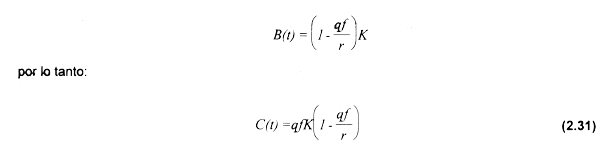
Multiplicando (2.31) por el precio promedio de la especie objetivo y dividiendo entre el esfuerzo, se obtiene el valor promedio del esfuerzo pesquero (VPE):

Derivando la ecuación de captura de Schaefer (2.8) con respecto al esfuerzo pesquero se tiene que:

Multiplicando la ecuación anterior por el precio promedio de la especie (p), se obtiene el valor del producto marginal del esfuerzo pesquero (VME)::

lgualando la expresión (2.33) al costo unitario del esfuerzo pesquero, c, y despejando f, se obtiene el esfuerzo en máximo rendimiento económico (fMRE):
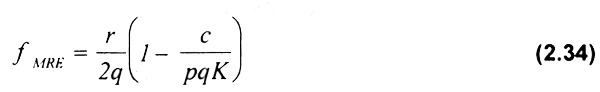
El equilibrio bioeconómico se alcanzará cuando el valor promedio del esfuerzo pesquero sea igual a los costos por unidad de esfuerzo (VPE = c). Este último se estima igualando la función (2.32) a los costos (c) y despejando f. Al hacer lo anterior se podrá observar que fEBE = 2fMRE, es decir que el esfuerzo en equilibrio bioeconómico es dos veces mayor que el esfuerzo en máximo rendimiento económico (Fig. 2.4b; ver también Fig. 2.3.c).
El modelo tiene implicitos varios supuestos, tales como:
a) El precio es considerado constante. Asimismo, los costos marginales y promedio se asumen constantes para cualquier nivel de esfuerzo pesquero.
b) Los cambios en el esfuerzo se darán de manera inmediata como respuesta a las variaciones en los ingresos netos.
c) El recurso responderá inmediatamente a la reducción en el esfuerzo.
d) Los costos totales crecen de manera proporcional al esfuerzo de tal manera que un cambio de pendiente en la curva de costos determinara un desplazamiento en el nuvel en que se alcanza el equilibio bioeconomico y el MRE.
e) Como resultado de su derivación con respecto al modelo de Schaefer, no considera cambios en la población dependientes de las caracteristicas intrinsecas a su ciclo de vida, tales como longevidad, temporalidad del ciclo reproductivo y variabilidad en el reclutamiento (Lawson, 1984).
Entre las limitaciones de tipo logistico cabe mencionar que puede llevar años recopilar la información de datos biológicos requeridos y la colecta de las estadisticas de esfuerzo y rendimiento, y más aún los económicos. Además, en la mayoría de los casos la precisión de las estadísticas utilizadas es poco confiable, sobre todo en paises en desarrollo donde los costos de información son muy altos.
Clark (1985) menciona que es dificil distinguir si el efecto de disminución de una población se debe a presiones de pesca o a procesos naturales. En la práctica parece ser que la capacidad del esfuerzo para ciertas pesquerías es mucho más que dos veces el nivel óptimo; Clark (1985) concluye que el modelo sirve para explicar el problema de sobrepesca, pero parece inadecuado para proponer medidas de manejo.
Smith (1969) propone un modelo donde asume que el esfuerzo pesquero cambia proporcionalmente a los ingresos netos:

donde φ es una constante positiva que describe la dinámica de la flota en el largo plazo. Smith emplea esta ecuación para modelar los cambios en la flota de una pesquería, los cuales se obtienen de sustituir (2.20) en (2.35):
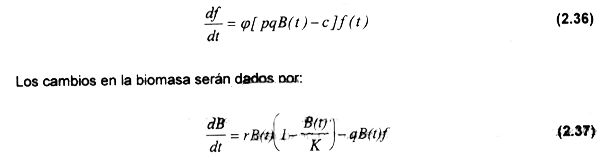
Si π(t) es mayor a cero, habrá un aumento en el esfuerzo resultante de la entrada de barcos a la pesquería, mientras que se producirá una disminución de éste en caso contrario. El parámetro φ puede estimarse empiricamente de acuerdo al comportamiento de la flota (entradas y salidas) en función de variaciones en los ingresos netos, que tendrán relación con los costos ejercidos para los correspondientes niveles de esfuerzo.
El modelo de Smith aunque robusto, puede complementarse para incorporar un retraso entre la toma de decisión y la entrada o salida de embarcaciones. El capital tiene larga vida de trabajo y los factores institucionales, tales como sistema de pago compartido de grupos de pescadores, tienden a amortiguar el retorno del capital, de tal forma que las consecuencias del período de ajuste del esfuerzo no se reflejan inmediatamente. El ingreso de nuevas embarcaciones a la pesquería tendrá un retraso si se considera el tiempo de obtención de permisos de pesca y de créditos y el tiempo requerido para la construcción de la embarcación. Los beneficios pueden obtenerse por varios períodos sin tener un cambio significativo sobre el esfuerzo.
Seijo (1987) propone la adición de un término de retraso distribuido representado por la función de densidad probabilistica Erlang (Manetsch, 1976), que incorpora el tiempo requerido para la salida o entrada de barcos a la pesquería, una vez que se manifiesta el efecto de cambios en el esfuerzo dados los ingresos netos anuales. El modelo incluye un parámetro que sirve para retrasar el tiempo de ocurrencia de un fenómeno (DEL), en este caso entrada y salida de barcos, el cual se define como tiempo de tránsito promedio y variará de acuerdo a la categoria de las embarcaciones. En el Capitulo 5 se describe la aplicación del modelo de retraso distribuido a pesquerías marinas.
Las decisiones de dinámica del esfuerzo pesquero en el corto plazo no son expresadas en el modelo de Smith, por lo que serán introducidas, discutidas y modeladas en capítulos posteriores.
Ejemplo 2.1 Modelo bioeconómico dinámico
Dada una pesquería de una especie pelágica cuyos parámetros aparecen en la Tabla 2.1, se analiza el comportamiento dinámico de la biomasa de la población, los rendimientos, la flota, los ingresos y costos.
| Parámetro/Variable | Valor | Unidad de medida |
|---|---|---|
| Tasa intrinseca de crecimiento | 0.36 | -- |
| Coeficiente de capturabilidad | 0.0004 | -- |
| Capacidad de carga del ecosistema | 3500000 | ton |
| Precio de la especie | 60 | US$/año |
| Costo unitario del esfuerzo | 30000 | US$/año |
| Biomasa inicial de la población | 3500000 | ton |
| Parámetro de dinámica de la flota | 0.000005 | -- |
Aplicando el modelo dinámico Gordon-Schaefer con los parámetros bioeconómicos de la Tabla 2.1 se puede observar la trayectoria dinámica de la biomasa, el rendimiento y los ingresos y costos totales (Fig. 2.5).
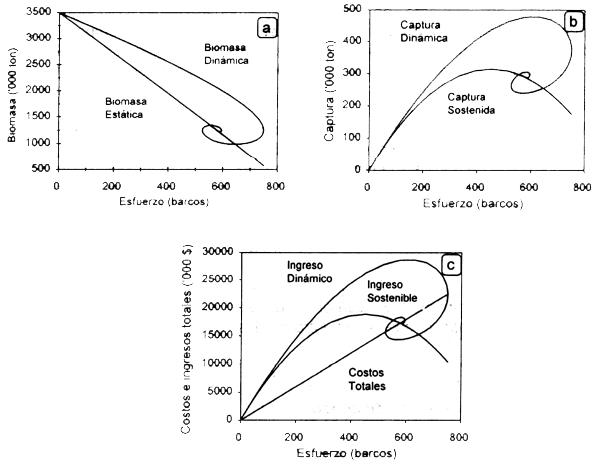
Figura 2.5 Trayectoria estática (equilibrio) y dinámica de la biomasa (a), las capturas (b) y los costos e ingresos (c) resultantes de la aplicación de diferentes niveles de esfuerzo.
En esta figura se incluyen también las curvas en equilibrio para las variables de desempeño antes mencionadas para observar la diferencia cuantitativa de considerar el efecto de diferentes niveles de esfuerzo utilizando el modelo estático y el dinámico. El modelo muestra el comportamiento dinámico de las diferentes variables donde se tiende al equilibrio en todos los casos, el cual se alcanza con 578 barcos. El esfuerzo en máximo rendimiento económico (fMRE) es de 289 embarcaciones.
En la Figura 2.6 se presenta la evolución en el tiempo de las variables de desempeño bioeconómico en el modelo dinámico. En la Figura 2.6a se puede observar la dinámica de la biomasa como respuesta a la dinámica del esfuerzo. En la Figura 2.6b se presenta el rendimiento en el tiempo que obtiene una flota que va modificando su esfuerzo pesquero en la medida que la renta obtenida en el tiempo varia. En la Figura 2.6c pueden observarse las tendencias crecientes, decrecientes y estabilizantes del esfuerzo de pesca en la medida que la renta crece, decrece, se hace negativa y posteriormente tiende a cero, convergiendo a nivel de esfuerzo en el equilibrio bioeconómico.
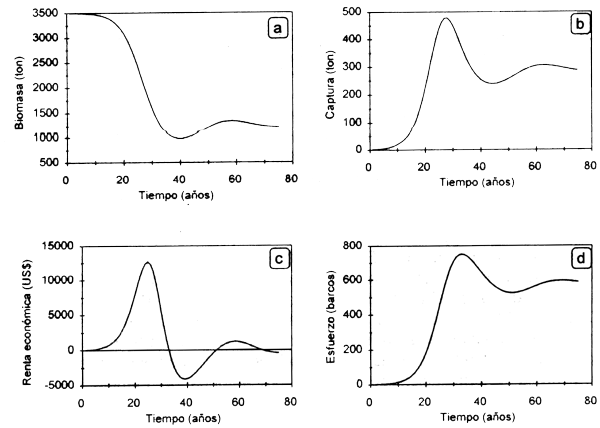
Figura 2.6 Dinámica de la biomasa (a), la captura (b), la renta económica (c) y el esfuerzo pesquero (d).
Como se puede observar, los incrementos en el esfuerzo dan lugar a aumentos en las capturas y por tanto en los ingresos generados por la pesquería. Como consecuencia, la CPUE decrece de manera paulatina si el esfuerzo continúa creciendo, lo que indicaria disminución de la biomasa de la población. Si el esfuerzo pesquero sigue aumentando a un nivel mayor de 630 embarcaciones, se observa que las capturas y los ingresos netos disminuyen. Posteriormente se entra a una dinámica de entrada y salida de embarcaciones a la pesquería en la medida que la renta se hace positiva o negativa, hasta un nivel de esfuerzo en el que la renta tiende a cero, en condiciones de equilibrio bioeconómico.
Analizando la información en el tiempo se observa que a los 33 años se alcanzan los mayores niveles de esfuerzo pesquero, lo que da lugar a una caida brusca de la biomasa en los años siguientes, repercutiendo en los ingresos netos. Al no obtener ganancias, no hay ingreso de embarcaciones, lo que permite a la población recuperarse y tiende a estabilizarse cerca de las 1200 ton después de 50 años de operación de la pesquería, al igual que el esfuerzo pesquero. En este punto los ingresos netos son iguales a cero, ya que los costos totales son iguales a los ingresos totales.
Los modelos no lineales “captura-mortalidad” se basan en el análisis conjunto de series de datos correspondientes a la captura commercial (variable dependiente) en función de la tasa instantánea de mortalidad total Z (variable independiente) (Csirke & Caddy, 1983).
La creciente adquisición de series de tiempo prolongadas, tanto de capturas como de valores de Z, provee un atrayente campo para la aplicación de estos modelos “output-output” (tanto la captura como la mortalidad total son salidas de los subsistemas biológico y económico), lo cual ha suscitado recientes publicaciones destinadas a realizar una revisión de la veracidad de diferentes tipos de ajuste a los modelos captura-mortalidad (Caddy & Defeo, 1996). Otro motivo fundamental es que dichas técnicas son adecuadas para formular estrategias de manejo aversas al riesgo y en consecuencia para adoptar medidas precautorias de administración de recursos pesqueros (ver Capitulo 8 y también FAO, 1993). En esta sección se describen los modelos “captura-mortalidad”, tanto en base a las aproximaciones clásicas de Schaefer (1954) como de Fox (1970). La mayor parte está tomada de los clásicos articulos de Caddy & Csirke (1983) y Csirke & Caddy (1983), asi como del trabajo realizado por Caddy & Defeo (1996), quienes extendieron la teoria desarrollada en los anteriores trabajos al uso del modelo exponencial, adicionando asimismo un análisis de la variabilidad, riesgo e incertidumbre en la estimación de parámetros y en la determinación de medidas precautorias de manejo (ver Capitulo 8 al respecto).
Obtención de valores de mortalidad. Los valores anuales de Zt (mortalidad total en el año t) pueden ser obtenidos de la fracción descendente de una curva de captura basada en edades (Robson & Chapman, 1961) o longitudes (Sparre et al., 1989). También puede ser obtenida de tallas medias (e.g. Beverton & Holt, 1956) o bien usando métodos tales como el de Heinke ó Robson & Chapman (ver Ricker, 1975).
Los análisis convencionales de Análisis de la Población Virtual/Análisis de Cohortes (Pope, 1972; Jones, 1981) también pueden ser utilizados para estimar valores de mortalidad por pesca F por edad, los cuales podrian ser transformados en Z al adicionar una estimación de mortalidad natural M. Es importante considerar que el análisis de cohortes opera en base a las “diagonales” que siguen la historia de una cohorte dada a través del tiempo; es posible sin embargo considerar las series horizontales de capturas (e.g. la composición por edades de las capturas en un año dado) en forma independiente a fin de generar curvas de captura que provean de valores de Z a ser usados como entradas a los modelos “captura-mortalidad”.
Un simple promedio de estimaciones independientes de Zt basadas en el cociente de los logaritmos de las abundancias de sucesivas clases de edad que forman una importante proporción de las captura anual, puede ser obtenido desde la edad de primera captura en adelante. Una discusión de las diferentes aproximaciones para estimar mortalidad puede ser encontrada en los textos clásicos de Ricker (1958), Pauly (1980), Jones (1981), Caddy (1986) y Sparre et al. (1989).
a) Ajuste directo a través de regresión múltiple. Tal como fue expresado en la ecuación (2.4), el modelo logistico de crecimiento poblacional propuesto por Graham (1935) en condiciones de explotación pesquera (ver Ricker, 1975) está dado por:

Csirke & Caddy (1983) expresaron este rendimiento en equilibrio en términos del valor en equilibrio de la tasa de mortalidad anual (ver p. 45 y también Caddy, 1986), y redujeron la ecuación (2.38) a una forma cuadrática:
Ct=aZ2t+bZt+c (2.39)
donde: Ct es la captura correspondiente al año t.
Zt es el coeficiente promedio de mortalidad total para el año t.
Bajo los supuestos del modelo logistico, la ecuación (2.39) da una parábola que intercepta la abscisa a la derecha del origen (Fig.2.7). De esta forma, usando regresión múltiple, donde Zt y Z2t son las dos variables independientes de este polinomio de segundo orden, podemos estimar la curva que relaciona los valores anuales de captura con la mortalidad total. Dicho ajuste provee una estimación del coeficiente de mortalidad natural M a través de la solución de esta ecuación para Z = M (Ct y F = 0) de la siguiente manera (Csirke & Caddy, 1983):

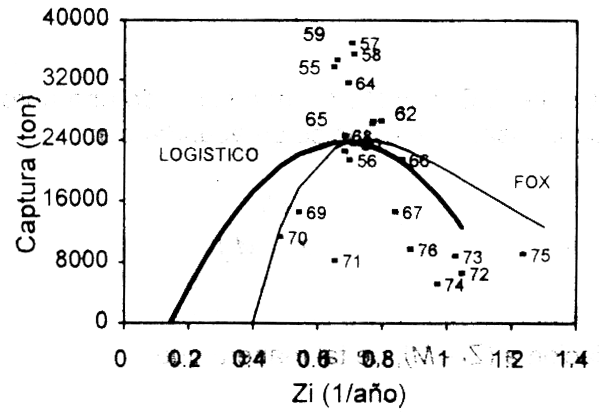
Figura 2.7 Ejemplo ilustrativo del modelo captura-mortalidad de Csirke & Caddy (1983).
Asimismo, los parámetros del modelo logístico pueden obtenerse para aquellos valores de Z correspondientes a MRS, el cual ocurre cuando (Csirke & Caddy, 1983):
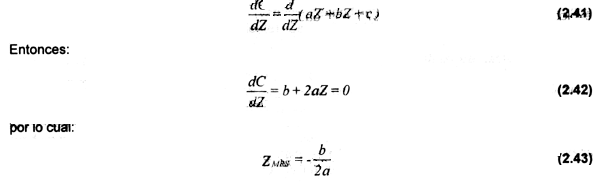
FMRS, definido como el nivel de mortalidad por pesca en MRS, está dado por la substracción de
M a ZMRS:
FMRS = ZMRS- M (2.44)
Los otros parámetros del modelo logístico están dados por:
r=2 FMRS (2.45)

b) Aproximación lineal. Csirke & Caddy (1983) han sugerido una aproximación alternativa para el ajuste del modelo logístico, basada en el indice de abundancia:

el cual es analizado en relación a (Zt - M), de tal manera que:

La ecuación (2.49) ha sido preferida al ajuste anterior debido a objeciones teóricas al ajuste directo del modelo (Hoenig & Hoenig, 1986), el cual en algunos casos tiende a subestimar M. Este modelo requiere de una aproximación especial, ya que se emplean en forma secuencial diferentes valores semilla de M en la ecuación (2.49), de los cuales se selecciona aquél que provea el valor más alto de bondad de ajuste (e.g. coeficiente de determinación R2). Dicho valor puede ser usado para transformar las tasas anuales de mortalidad total Zt en tasas de mortalidad por pesca Ft. Es muy importante hacer notar que en este caso no se requieren de valores de esfuerzo o CPUE.
Los parámetros de esta versión lineal del modelo logistico pueden ser obtenidos como sigue:
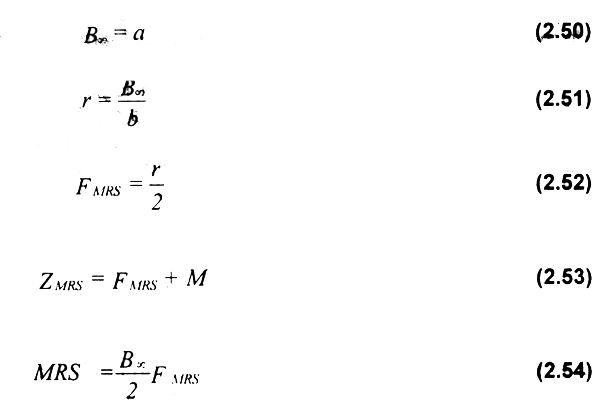
Caddy & Defeo (1996) extendieron la teoria de los modelos de producción excedente basados en mortalidad para dar cabida a la versión exponencial del modelo logístico.
a) Aproximación no lineal. La versión exponencial del modelo logistico para datos de captura y esfuerzo (Fox, 1970) puede ser expresada como:
Ct=FtB∞e-eft (2.55)
donde el rendimiento anual Ct y el esfuerzo ft están relacionados en una forma exponencial, y Ux es la tasa de captura correspondiente al stock virgen. A efectos de transformar el esfuerzo en tasas de mortalidad analiticas, si consideramos que q y M son constantes, se tiene que:

Definiendo b'=b/q y considerando que U/q=B, entonces:
Ct=(ZtM) B∞ e-b'(Zt-M) (2.57)
Esto puede ser expresado como:

donde Bx y b' pueden ser estimados por técnicas de regresión no lineal, usando procedimientos de minimización de errores (e.g. quasi-Newton, Marquardt) para la estimación de parámetros.
Como en el caso de la aproximación logística, alternativa, el modelo es ajustado para diferentes valores semilla de M, seleccionando aquél que maximice cierto criterio de bondad de ajuste; una vez obtenido el mejor valor de M, los parámetros de manejo restantes pueden ser calculados diferenciando la ecuación (2.60) con respecto a F y llevándola a cero al nivel de MRS:

y ZMRS se logra en función de la suma de FMRS a M.
b) Aproximación lineal. Una aproximación alternativa a la anterior puede lograrse al tomar los logaritmos naturales de ambos miembros de la igualdad y ajustando una regresión lineal de la forma Yi = a - b Xi:
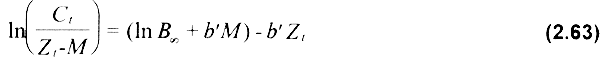
usando a In(Ct/Zt - M) y Zt como variables dependiente e independiente, respectivamente. La estimación de parámetros sigue el mismo razonamiento que en la aproximación previa, usando diferentes valores semilla de M, y difiriendo en la estimación de la biomasa virgen, la cual puede ser deducida de la ecuación (2.63) como:
B∞ = e(a -b'M) (2.64)
Ventajas. Algunas de las conclusiones emanadas de la aplicación de estos modelos de “captura-mortalidad” sugieren lo siguiente (ver también Caddy & Defeo, 1996):
1) La pobre calibración de los errores en la estimación del esfuerzo y los sesgos que esto conlleva en el uso de esta variable en los modelos captura-esfuerzo tradicionales, proporciona ventajas comparativas a los modelos captura-mortalidad.
2) Los cambios en el coeficiente de capturabilidad q con la intensidad de pesca y biomasa pueden ser reducidos empleando esta aproximación.
3) Caddy & Defeo (1996) demostraron la utilidad de extender la teoría desarrollada por Csirke & Caddy (1983) al uso del modelo exponencial. Una de las principales criticas de los modelos logísticos basados en el uso del esfuerzo como variable de entrada ha sido que, particularmente luego de alguna aproximación al equilibrio de algún tipo, la relación tasa de captura vs esfuerzo no es siempre lineal, sino que tiende a caer a una tasa progresivamente decreciente, haciendo la aplicación de los modelos exponenciales más adecuada que aquella basada en el logístico. Esta crítica es menos válida cuando se usan tasas de mortalidad para ajustar modelos de producción (Csirke & Caddy, 1983), debido a que la experiencia sugiere que se obtienen curvas de rendimiento más simétricas al graficar las capturas contra la tasa de mortalidad total. La conclusión a la cual arriban Caddy & Defeo (1996) es que ambos tipos de modelo sean considerados con precaución, y a efectos de manejo, sea utilizado en mayor medida aquél que provea la aproximación más cauta, ésta evaluada en función del simple análisis de riesgo propuesto por dichos autores. En el Capítulo 8 se analizará con mayor detalle aspectos relacionados con la estimación en la variabilidad de los parámetros y del análisis de riesgo a efectos de manejo.
4) Estos modelos, en su versión estática, asumen equilibrio. Los valores medios de tasas de mortalidad Zt obtenidos de métodos basados en curvas de captura u otros (ver Caddy & Defeo, 1996) representan el impacto de la pesca en el presente y pasado reciente en forma más cercana que las aproximaciones que emplean valores anuales de esfuerzo de pesca (o bien de Zt basados en el cociente de la abundancia de una clase de edad simple en dos sucesivos períodos de tiempo). Por tanto, es posible compensar desviaciones de mortalidad anual con respecto al equilibrio, tal como el análogo a los promedios móviles (“past averaging”) de esfuerzo de pesca para dar cuenta de los efectos de mortalidad en años previos (Gulland, 1961; Fox, 1975). Esta aproximación puede resultar bastante conservadora en caso de usar una prolongada serie de datos en el número de datos consecutivos a promediar. El problema se agrava en caso de usar modelos basados en esfuerzo (no en el caso de mortalidad), en los cuales es dificil decidir cuantos años deberian ser incluidos; en este caso variaria dependiendo de los cambios en la intensidad de pesca con el tiempo. En caso de considerar que el promedio móvil de estimaciones de mortalidad es apropiado, entonces los modelos “captura-mortalidad” tienen una ventaja definida (Caddy, 1986): el recíproco de la mortalidad promedio para las clases de edad plenamente reclutadas es en si un estimador simple del número de clases de edad anuales que constituyen la captura. Entonces, es una medida del tiempo en el cual la aproximación de promedios móviles hacia atrás (“past mortality averaging”) deberia ser aplicado. Por tanto, 1/Zt podría ser considerado como un indice del número de grupos de edad plenamente reclutados que sobreviven de la explotación en años anteriores y que de una otra forma constituyen la captura actual. Lo importante de este razonamiento radica en que cuando existen estimaciones de mortalidad para varios grupos de edad, hay un cierto grado de ajuste semiautomático de la mortalidad pasada en cada estimación individual anual de mortalidad (Caddy, 1986).
5) Las estimaciones de parámetros sin dar cuenta de ajustes al equilibrio son de todas maneras valiosas. Idealmente, a menos que existan variaciones anuales considerables en el esfuerzo de pesca, las series de captura-mortalidad tienden a mostrar cierto grado de autocorrelación serial sin las fuertes oscilaciones desde la izquierda a la derecha de las curvas de rendimiento características de muchos modelos de producción captura-esfuerzo, y por tanto con amplias desviaciones del equilibrio.
Desventajas. Algunas de las principales limitaciones a la aplicación de este tipo de modelos consisten en:
1) La estructura de la base de datos debe abarcar un amplio rango de valores de tasas de mortalidad. Las estimaciones de los parámetros son en fuerte grado dependientes de los valores extremos, en particular de dicha variable independiente. La interpretación y evaluación de los efectos de estos puntos con criterios de manejo debe ser cautelosamente analizada, ya que los valores de Z situados en los extremos de las curvas de producción influyen en el comportamiento de los modelos y pueden ser responsables de amplios intervalos de confianza (Caddy & Defeo, 1996). Paradójicamente, dichos puntos proveen la información más importante desde un punto de vista biológico-pesquero, y contribuyen significativamente a la explicación de la variabilidad observada en las bases de datos. Caddy & Defeo (1996) mostraron que, al omitirse estos datos por un procedimiento de remuestreo, se obtuvieron valores de los parámetros que influenciaron fuertemente la estimación de sus medias e intervalos de confianza (ver también Capítulo 8).
2) Tal como se mencionó, anteriormente, la versión analizada de estos modelos es estática y asume equilibrio. Uno de los tópicos que ha sido objecto de reciente debate (Hilborn & Walters, 1992, Polacheck et al., 1993, Caddy & Defeo, 1996) es la dicotomía entre modelos dinámicos vs estáticos, los cuales asumen equilibrio para la estimación de parámetros. Si bien la teoría de modelos de producción con tasas de mortalidad está mucho menos desarrollada que aquella basada en el esfuerzo de pesca, existe la posibilidad de obtener una versión dinámica, en la medida en que permita disipar dudas sobre los desajustes producidos por la ausencia de equilibrio.
La aproximación anterior constituye un enfoque netamente biológico. Con el propósito de obtener un punto de referencia bioeconómico en el proceso de manejo precautorio de pesquerias, a continuación se presenta una expresión que estima la renta generada por el recurso (π) a partir de la ecuación captura-mortalidad. Se toma como ejemplo de desarrollo el modelo linearizado de producción excedente de Fox (1970) adaptado a criterios captura-mortalidad:

donde: p es el precio promedio de la especie capturada. c es el costo unitario del esfuerzo pesquero.
Diferenciando la ecuación anterior, se obtiene una expresión de la renta marginal (πM) con respecto a cambios en el coeficiente de mortalidad por pesca:

Ahora bien, igualando a cero la renta marginal y resolviendo la ecuación con respecto a F, se obtiene una expresión que representa el valor de mortalidad por pesca que genera el máximo rendimiento económico (FMRE):
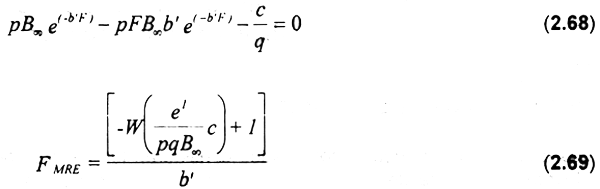
haciendo a = 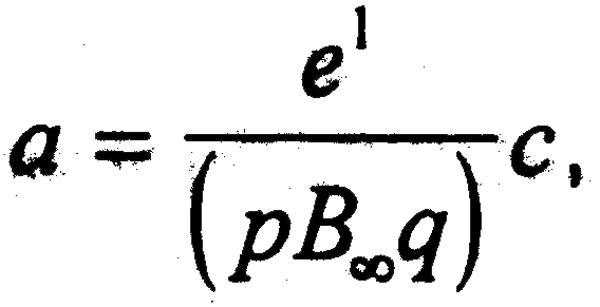 se tiene que W [ a ] es una función especial de MathCad ®1 que puede
se tiene que W [ a ] es una función especial de MathCad ®1 que puede
1 Mathcad 5.0 for Windows. 1994. Mathsoft, Inc.
expresarse de la forma siguiente:
a = Wexp(W) (2.70)
En base a lo anterior se obtiene la expresión para FMRE:

Ejemplo ilustrativo Dados los parámetros del modelo linearizado exponencial captura-mortalidad: p = $60; q = 0.001; b' = .85; c = $ 200000; y Bx = 15.0×106 ton y después de estimar numéricamente el valor de W = 0.404, la mortalidad por pesca que genera un máximo rendimiento económico FMRE se obtendrá al sustituir estos valores en la ecuación 2.53 teniéndose que FMRE = 0.701/año y una captura en máximo rendimiento económico CMRE = 5.795×106 ton correspondiente a un esfuerzo en fMRE igual a 701 embarcaciones.
Los modelos de estructura por edades consideran los factores de crecimiento y mortalidad que afectan la biomasa de una población distribuída de manera homogénea en el espacio y en el tiempo. A diferencia de los modelos globales, los cuales consideran la biomasa total de la población, las aproximaciones derivadas de estos modelos se caracterizan por incluir de manera explícita la estructura por edades de la población y los procesos que la alteran, donde la edad está asociada a la talla, crecimiento individual, reclutamiento, fecundidad y vulnerabilidad a la pesca (Hilborn & Walters, 1992).
Estos modelos parten de los trabajos iniciales de Beverton & Holt (1957) quienes definen a la captura (C) como el producto de la mortalidad por pesca (F) por la biomasa de la población, la cual resulta del producto del peso individual (W) por el número de individuos presentes en ese momento de tiempo (N).
La captura en número y peso resulta de integrar la abundancia de las clases de edad de aquellos individuos reclutados a la pesquería (tr) hasta la de máxima longevidad (t1):
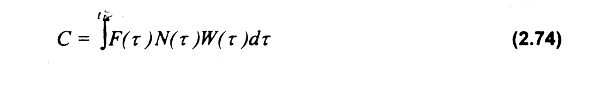
Considerando la extrema variabilidad en los parámetros poblacionales y en la mortalidad por pesca (ésta ocasionada por una distribución agregada del esfuerzo de pesca) en especies sedentarias, el clásico análisis de rendimiento por recluta (Beverton & Holt, 1957) debería ser realizado en forma separada para cada edad identificada (j edades), cada una sujeta a diferente intensidad de pesca y talla de media captura (Caddy, 1975):
Ct= ∑ (C/R)j(R)j (2.75)
donde Cj es el rendimiento por edad, C/Rj es el rendimiento por recluta γ Rj es el reclutamiento estimado por edad j. De esta manera los supuestos clásicos de los modelos de agrupamiento dinámico se harían efectivos. La extensión del modelo clásico a efectos de un análisis multiespecífico es igualmente viable (Ursin, 1982), aunque en caso de especies sedentarias debería considerarse en una primera instancia la heterogeneidad espacial mencionada y luego integrar los resultados. Un análisis bioeconómico multiespecífico es posible con la inclusión de los costos unitarios del esfuerzo de pesca (c) y el precio de la especie capturada (p) en la ecuación tradicional (Arreguín-Sánchez, 1989).
El modelo supone las siguientes condiciones:
a) Reclutamiento a la pesquería en forma de “filo de navaja”, al igual que la selección de edades al arte de pesca. Se supone la incorporación de los reclutas a la pesquería en un mismo momento.
b) Todos los peces de una cohorte son capturados al mismo tiempo.
c) Distribución homogénea de la población, y por tanto los individuos que la componen tienen la misma probabilidad de ser capturados.
d) La tasa de exploitación se asume constante durante toda la vida de los organismos. Los rendimientos son estables en el tiempo si el patrón de exploitación se mantiene, lo que implica reclutamiento constante.
e) La mortalidad natural es constante independientemente de la edad, inclusive a partir de aquella que es reclutada a la pesquería; la mortalidad por pesca es igual a cero para edades menores a la de primera captura, por lo que las pérdidas en N(t) antes de la edad de primera captura (tc) serán:
N(t)=R[-M(t-tr)] (2.76)
donde R es el número de peces vivos al tiempo t=tr (edad de reclutamiento a la población). Ahora bien. N(t) para todas las edades capturadas se define como:
N(t) = R'e[-(F-M)(t-t)] (2.77)
donde R' indica el número de individuos reclutados a la pesquería.
f) Otra suposición implícita es que el crecimiento individual se ajusta a la función propuesta por von Bertalanffy (1938):
W(t)=W∞[1-e-kp(t-t0)]b (2.78)
donde W∞ es el peso asintótico que alcanzan los individuos, kp es el parámetro de curvatura, t0 es el parámetro de ajuste de la ecuación γ b es el exponente de la relación peso-longitud.
g) El modelo tradicional asume que el reclutamiento es independiente del tamaño del stock, γ que tampoco se ve afectado por variaciones en la intensidad de pesca, supuesto que ha sido considerado como el menos satisfactorio e irreal. Sin embargo, se ha tratado de incorporar al modelo las clásicas curvas de reclutamiento de Beverton & Holt (1957) y la de Ricker (1975). Considerando el modelo de reclutamiento de los primeros e incorporando los elementos anteriores en el modelo, la ecuación que define el modelo estático de rendimiento por recluta queda expresada de la siguiente forma:
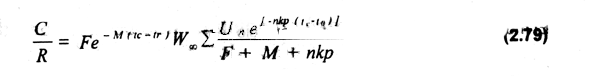
Muchas de las limitaciones del modelo han sido mencionadas en los propios supuestos que lo sustentan (e.g. mortalidad natural y por pesca constantes, independencia del reclutamiento con respecto al tamaño del stock parental). El considerar la mortalidad por pesca constante puede tener un serio impacto en el manejo de pesquerías, debido a que en general hay diferencias marcadas con la edad producidas por ejemplo por diferencias en su accesibilidad al arte de pesca. En tal sentido, se omiten los diferentes tipos de tecnologia empleados por diferentes grupos de pescadores en casos de esfuerzo pesquero heterogéneo. La no incorporación de elementos económicos dentro del modelo constituye una limitante adicional a la hora de evaluar diferentes estrategias de manejo.
A diferencia del modelo estático propuesto por Beverton & Holt (1957), el modelo dinámico toma en cuenta los influjos y eflujos para cada cohorte de la estructura de la población, aplicando los principales conceptos de tablas de vida (Begon et al., 1990). La incorporación de los reclutas es dinámica, permitiendo un manejo estacional del reclutamiento γ su distribución (Seijo, 1986; Seijo & Defeo, 1994b).
Los cambios en el número de individuos en la población de acuerdo al modelo dinámico se definen como:
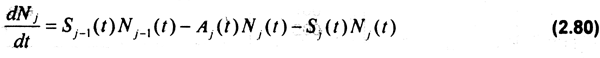
donde Sj es la sobrevivencia de los organismos de clase de edad j γ Aj corresponde a la tasa finita de mortalidad total (Gulland, 1983). Dicha tasa de sobrevivencia es expresada como sigue:
Sj-1(t)1- [MN j-1(t)+FRj-1(t)] (2.81)
donde MN(t) es la tasa de mortalidad natural γ FR(t) es la tasa de explotación, ambos derivados de los correspondientes coeficientes de mortalidad natural y por pesca.
Factorizando y ordenando términos:
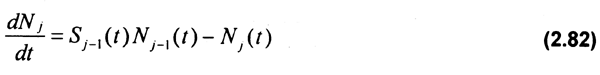
Así, el número de individuos de cada cohorte (Nj), se obtiene integrando en el intervalo de t+DT los individuos de la edad j-1 que sobreviven γ crecen dentro de la cohorte en el tiempo t, considerando las pérdidas dadas por la tasa de la mortalidad total de los individuos de la cohorte (Aj), asi como la tasa a la cual los organismos de la cohorte que sobreviven (Sj) son incorporados en la cohorte j+1 en el tiempo t (Seijo & Defeo, 1994b).
Integrando numéricamente la ecuación (2.84) con el método de Euler, se obtiene la dinámica de la estructura de la población:
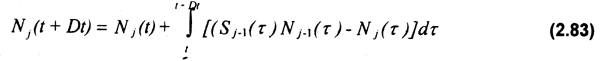
Nj(t-DT)=Nj(t)+DT[Sj-1Nj-1(t)-Nj(t)] (2.84)
En este caso el modelo también emplea la función de crecimiento de von Bertalanffy para la estimación del peso promedio de los individuos, por lo que la biomasa para cada clase de edad será:
Bj ( t + Dt ) = Nj ( t + Dt ) t Wj (2.85)
de donde se deriva la biomasa total B(t) como resultado de la sumatoria de la biomasa de todas las clases de edad:

La captura Cj(t) y la mortalidad por pesca (Fj) por clase de edad se definen en base a las ecuaciones 2.2 y 2.9 respectivamente, pero en este caso tanto la biomasa como q son discriminados por clase de edad.

Cj (t) = qj Bj (t) f (t) (2.88)
Los ingresos totales IT se obtendrán al multiplicar el precio por edad de la especie (pj) por la captura derivada de cada clase de edad:

El precio puede variar por tallas, γ por tanto puede ser introducido en la ecuación.
Los costos totales CT(t) e ingresos netos π(t) se obtienen de acuerdo a las ecuaciones desarrolladas en ocasión del modelo bioeconómico Gordon-Schaefer.
Ejemplo 2.2 Modelo bioeconómico de estructura por edades dinámico
Dada una pesquería de arrastre hipotética cuyos parámetros aparecen en la Tabla 2.2, se analiza el comportamiento dinámico de la biomasa de la población, los rendimientos, la flota y los ingresos, considerando variaciones en la edad de primera captura y el esfuerzo de pesca.
| Parámetro/Variable | Valor | Unidad de medida |
|---|---|---|
| Edad máxima de la especie | 10 | años |
| Edad de primera madurez sexual | 2 | años |
| Fecundidad promedio | 5000 | huevos |
| Edad de primera captura | 2 | años |
| Proporción por sexos | 0.5 | -- |
| Coeficiente de mortalidad natural | 0.2 | 1/año |
| Parámetro de curvatura ec. von Bertalanffy | 0.5 | 1/año |
| Parámetro de ajuste ec. von Bertalanffy (t0) | 0.0 | -- |
| Longitud asintótica | 100 | mm |
| Peso asintótico | 200 | g |
| Parámetros para la ecuación de selectividad | L50%=20 | mm |
| L75%=30 | mm | |
| Area barrida por día | 0.1 | Km2 |
| Area total de distribución del recurso | 10 | Km2 |
| Máximo reclutamiento observado (Rmax) | 20000000 | organismos |
| Precio de la especie | 10000 | $/ton |
| Costo unitario del esfuerzo | 75000 | US$/barco/año |
| Parámetro de dinámica de la flota | 0.00005 | -- |
Como se puede observar en la Figura 2.8, a medida que se reduce la edad de primera captura (tc)la pendiente inicial de la curva de biomasa es mayor y la captura se incrementa, cayendo posteriormente a niveles menores a los de la edad immediata superior, estabilizándose en ambos casos después de los 45 años (Fig. 2.8a, b y c). Sin embargo, un aumento indiscriminado en la edad de primera captura pudiera no justificar la salida de embarcaciones a la pesquería, ya que el componente de la estructura de la población autorizada para ser capturada posiblemente no cubra los costos del esfuerzo de pesca. En este ejemplo, con edades mínimas de captura mayores a 4 años este comportamiento podría esperarse. Debe observarse que, con los parámetros bioeconómicos proporcionados en este ejemplo ilustrativo, las tasas máximas de captura γ de renta se obtienen al incrementar la edad mínima de captura de 1 a 2 años. Desde luego, la biomasa resultante de una mayor edad mínima de captura tiene un comportamiento dinámico que indica mayores niveles de biomasa y menor fluctuación de la misma. Respecto a la dinámica del esfuerzo de pesca, éste tiende a incrementarse de manera proporcional a las rentas generadas por la pesquería ante los diferentes escenarios de edades mínimas de primera captura (Fig. 2.8d). En el largo plazo, y bajo condiciones de acceso abierto, con una edad mínima de captura de 4 años la pesquería es capaz de soportar más del doble del número de embarcaciones que con la edad mínima de captura de 1 año.

Figura 2.8 Modelo bioeconómico de estructura por edades: efecto dinámico de diferentes edades mínimas de captura en la biomasa (a), la captura (b), la renta (c) y el esfuerzo de pesca (d).
Ahora bien, graficando las variables de desempeño bioeconómico de la pesquería (biomasa, rendimiento γ renta) con respecto a cambios dinámicos en el esfuerzo de pesca, se pueden observar las trayectorias dinámicas ante diferentes edades mínimas de captura. Con una edad mínima de 4 años, la biomasa decrece ante diferentes niveles de esfuerzo a una tasa menor que las obtenidas con edades mínimas de captura de 1, 2 y 3 años (Fig. 2.8a). Asimismo puede observarse que, dados los parámetros bioeconómicos especificados, el esfuerzo de pesca máximo que llega a tener la pesquería es de cerca de 200 embarcaciones, cantidad que tiende a disminuir ya que a partir de ese nivel de esfuerzo la renta se hace negativa (Fig. 2.8c). El número de embarcaciones resultantes con las diferentes edades mínimas de captura, en condiciones de equilibrio bioeconómico, es de 67, 90, 115 y 142 (Fig. 2.8d). Es decir, el operar con una edad mínima de captura relativamente grande (4 años) permite soportar un mayor número de embarcaciones γ por consiguiente generar un mayor número de empleos. La captura obtenible con el nivel de esfuerzo en equilibrio bioeconómico se incrementa de 158 ton con una edad mínima de 1 año a 272 ton con una edad mínima de 4 años (Fig. 2.8c).
Como puede observarse de la discusión previa, el modelo bioeconómico de estructura por edades permite explorar el impacto de diferentes edades mínimas de captura. Esta importante variable de control en pesquerías donde la selectividad por tallas es viable, es un instrumento de ordenación que los modelos globales del tipo Gordon-Schaefer no tienen la capacidad matemática de analizar.
![]()
![]()
![]()