![]()
![]()
![]()
Population “senegalo-mauritanienne” de chinchard Trachurus trecae
par
N. Barkova (AtlantNIRO, URSS)
En 1988 les individus de 26–31 cm predominaient dans les captures de T. Trecae.
L'echantillonnage pour determiner la composition par taille et l'âge a été effectué généralement à bord des navires de recherches scientifiques et communiquées par les observateurs à bord de bateaux de pèche (tab. 1–3)
Pour les calculs on a utilisé le poids moyen par classe d'âge en tenant compte de l'absence de variations interannuelles importantes de ce paramètre.
| Age (ans) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| Poids (kg) | 0.060 | 0.138 | 0.250 | 0.340 | 0.400 | 0.450 | 0.550 | 0.680 |
Estimation du stock
En vue d'estimer la biomasse de Trachurus trecae (de 1981 à 1988) on a utilisé l'analyse de la population virtuelle (APV) et l'analyse par cohorte, proposée par Jones (1974). Les paramètres de l'équation de von Bertalanffy, utilisés dans cette méthode, sont: K=0.22;t0=-0.59;L=50.20;W=1.123 kg.
Dans les calculs on a employé la prise sommaire de Trachurus trecae au large du Sahara et de la Mauritanie et les données sur la composition taille-âge de 1981 à 1988. Puisque les statistiques concernant le chinchard n'étaient passéparéespar espèces, on les a différenciées à partir des proportions dans les prises.
Seules les captures des carangidés ont été prises pour 100%. Au cours de l'estimation on a utilisé les données fournies par les observateurs embarqués à bord des navires de pêche, de détection et de recherches scientifiques. Les migrations saisonnières ont été prises en compte.
Les données obtenues on été traitées mensuellement.
La standardisation de l'effort de pêche a été effectuée à l'aide de la méthode de Shimada-Shaffer. On a pris le BMRT comme bateau-standard. Le coefficient de mortalité naturelle momentannée avait été déterminé d'avance (CECAF) et était égal à 0.6.
La détermination des coefficients terminaux de mortalité par pêche a été réalisée à l'aide des différentes méthodes de l'APV proposées par Pope et Shepher. (1983), Babajan (1984). Tout comme pour le sautres espèces l'erreur moyenne quadratique d'appro ximation de prise par effort à partr de l'équation linéaire de regression dont l'argument était la biomasse, determinée par l'APV a été utilisée en tant que critère d'adequation des méthodes. L'indice de force de liaison (présenté par le coefficient de corrélation), prévu par la méthode, a été également utilise en tant que critère complémentaire. Dans le cas de gamma-méthode c'est la valeur du coefficient de corrélation entre les logarithmes de l'abondance d'une classe d'âge et de l'indice relatif de prise par effort. Les erreurs d'approximation pour les différentes méthodes sont les suivantes:
| Méthode | Erreur |
|---|---|
| Equation de regression | 2.732 |
| Méthode de plusieurs années | 2.745 |
| Gamma-méthode | 2.715 |
| Méthode d'Armstrong | 2.749 |
| Méthode de Saville | 2.739 |
| Méthode de Highdol-Jones | 2.739 |
| Méthode de Laurec- Shepher | 2.780 |
| Ro-méthode | 2.900 |
| Méthode des valeurs moyennes pondérées | 2.708 |
L'analyse des résultats obtenus a montré que les moindres erreurs d'approximation étaient obtenues pour la méthode des valeurs moyennes pondérées (2.708). Les coefficients terminaux de mortalité par pêche sont présentés au tab.6.
La répartition des prises industrielles par classe d'âge (exprimées en nombre) aussi bien que les valeurs de biomasse et d'abondance, obtenues par l'APV, sont présentées aux tab. 5, 7, 8.
Un des problèmes les plus importants
consiste en obtention de l'estimation correcte du recrutement. Tout de même actuellement on ne réalise pas de travaux spéciaux sur l'évaluation des jeunes de cette espèce. L'absence de ohalutage expérimental au large du Sahara et de la Mauritanie en 1988 a rendu difficile l'estimation du recrutement. C'est pourquoi on a réalisé l'analyse de relation sentre le stock paternel et le recrutement en appliquant les modèles linéaires de Beverton et Holt,de Ricker, de Cushing, de Chapman qui utilisent la méthode des moindres carrés avec la linéarisation préliminaire pour la détermination des paramètres. La fidélité de l'évaluation des paramètres a été déterminée d'après le test de Student, la probabilité de confiance étant de 0.95. La moinire erreur d'approximation de recrutement calculé par l'APV, par rapport au recrutement, précalculé d'après telle ou telle relation, sert de critère de l'utilisation de telle ou telle méthode. Cette erreur a été calculée d'après la formule suivante:
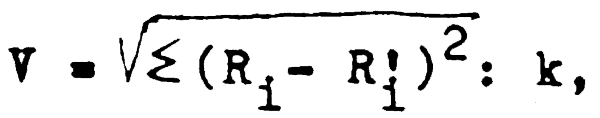
avec Ri - recrutement (abondance de la classe d'âge 1, déterminée par l'APV) Ri - recrutement, précalculé d'après telle ou telle relation; k - sous-échantillon.
Les erreurs d'approximation, obtenues à l'aide des différentes méthodes, sont présentées dans le Tab. 10
Les erreurs, obtenues d'après le modèle de Beverton et Holt et celle du Cushing sont très voisines, ce qui rend difficile le choix du modèle. Dans les buts du choix plus correct du rapport nous avons obtenu et analysé les écarts de la valeur du recrutement, déterminé par l'APV (dans notre cas-classe d'àge 1) et du recrutement, précalculé d'après les modèles
D'après le modèle de Cushing, la relation se détermine par l'équation suivante:
R' = A. PB,
où r - recrutement; P - stock paternel; A - const.
A et B - paramètres de relation. On a obtenu les valeurs suivantes des paramètres: A = 0.71.107; B = - 0.8469.
D'après le modèle de Beverton et Holt la relation se détermine par l'équation:

où E - stock paternel, A et B - paramètres du modèle.
Les calculs ont donné les valeurs suivantes de recrutement: - modèle de Cushing - 5183 millions d'ind; modéle de Beverton et Holt - 4733 millions d'ind.
En tant que critère d'adéquation de sméthodes on a employé les coefficients de correlation entre le recrutement, obtenu par l'APV et celui précalculé d'après le modèle. Pour le modèle de Cushing ce coefficient a représenté 0.881 et pour le modèle de Beverton et Holt- 0.876. Les valeurs élevées des coefficients de correlation obtenus permettent d'utiliser tous les deux modèles également. Mais, puisque la série des observations, utilisées pour la détermination des paramètres du modèle est courte, il faut les appliquer avec précautions. Nous avons préféré la valeur du recrutement, précalculé d'après le modèle de Cushing, car il se trouve pratiquement au niveau du recrutement moyen de plusieures années, qui etait égal à 5226 millions d'ind.
La répartition des prises par classe d'âge est représentée au tab. 5.
L'analyse des estimations rétrospectives de la biomasse permet de constater que les variations importantes de la biomasse de l'espèce considérée n'ont pas été observées au cours de 9 dernières années (1981–1988). Après une légère augmentation de la biomasse pendant la période de 1983 à 1985 (jusqu'à 25% par rapport à la période précédente) on peut noter une certaine diminution et la stabilisation du stock au niveau de 1400000 t. Compte tenu du fait que la qualité des statistiques de pêche est mauvaise ces dernières années et la différenciation des prises de chinchard et des résultats de l'analyse de la structure de taille nar espèce est insufissamment correcte, nous avons utilisé l'analyse des cohortes de Jones L'information essentielle utilisée dans cette méthode, est celle concernant la composition des prises par taille aussi bien que les paramètres de l'équation de von Bertalanffy. On a effectué les calculs à partir de données de la composition par taille pour la période de 1986 à 1988 (tab.11) On a utilisé les paramètres de l'equation de von Bertalanffy suivants: K = 0.22; to = -0.59; L = 50.20 et le rapport de l'équation de croissance W = 0.01 L2.98. L'evaluation du stock selon cette méthode a été effectuée pour chaque année avec des intervalles différents centrage (1986–1987, 1987–1988, 1986–1988) et pour les différentes valeurs de départ de la mortalité par pêche, se trouvant dans une gamme de 0.1 à 1.0. Il est à noter que la valeur du stock ne change pratiquement pas selon l'intervalle du temps, mais varie de 1285000 à 1335000 t, ce qui permet de constater l'état stable du stock de Trachurus trecae ces dernières années (tab. 12)
| Taille, cm | Zone du Sahara | Zone de Mauritanie | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Mois | Mois | ||||||||
| yII | yIII | IX | X | I | II | III | Iy | y | |
| I5 | 0,04 | ||||||||
| I6 | 0,I | ||||||||
| I7 | 0,0I | ||||||||
| I8 | 0,I | ||||||||
| I9 | 0,2 | 0,I | I,I | 0,03 | |||||
| 20 | I,2 | 0,04 | 3,3 | 3,6 | 0,03 | ||||
| 2I | 4,6 | 0,2 | 0,I | 0,7 | I2,9 | 3,6 | 0,4 | I,I | |
| 22 | II,5 | I,2 | 0,3 | 4,3 | 25,0 | 2,6 | 3,6 | 4,5 | |
| 23 | I,0 | I4,6 | 2,8 | 0,5 | I2,2 | I9,5 | 7,2 | I5,0 | 7,3 |
| 24 | 2,6 | I3,2 | 4,5 | 4,0 | I8,6 | I2,6 | I6,9 | 20,9 | 7,7 |
| 25 | I,I | I9,8 | 5,5 | 9,3 | I0,8 | 5,0 | 2I,5 | I5,9 | 6,3 |
| 26 | I,7 | 7,4 | 6,4 | I3,6 | I0,4 | I,9 | II,I | 8,6 | 7,0 |
| 27 | 7,I | 6,4 | 6,4 | I0,5 | 7,I | 4,I | 4,7 | 7,2 | 8,0 |
| 28 | I4,5 | 9,4 | II,5 | 9,2 | 7,4 | 6,7 | 3,4 | 6,3 | II,4 |
| 29 | 33,I | 8,9 | I9,4 | I5,9 | 3,9 | 2,7 | 4,8 | 6,9 | I7,I |
| 30 | 23, | 7,4 | I8,6 | I6,5 | 4,4 | 2,0 | 6,5 | 5,3 | I3,2 |
| 3I | 8,2 | 2,9 | I3,0 | 9,0 | 4,0 | I,I | 3,9 | 4,I | 6,9 |
| 32 | 4,4 | I,6 | 6,3 | 7,4 | 6,5 | I,6 | 3,0 | 2,2 | 4,I |
| 33 | 2,0 | 0,7 | 2,8 | 2,8 | 3,7 | 0,6 | I,9 | I,8 | 2,6 |
| 34 | 0,3 | 0,3 | 0,7 | 0,5 | 2,9 | 0,5 | I,5 | 0,9 | I,8 |
| 35 | 0,2 | 0,I | 0,2 | - | I,3 | 0,5 | 0,8 | 0,4 | 0,7 |
| 36 | + | 0,0I | 0,3 | 0,9 | 0,04 | 0,6 | 0,2 | 0,2 | |
| 37 | + | 0,6 | + | 0,3 | 0,3 | 0,I | |||
| 38 | 0,I | 0,2 | 0,05 | 0,02 | |||||
| 39 | 0,3 | 0,0I | |||||||
| 40 | 0,I | - | |||||||
| 4I | 0,07 | 0,0I | |||||||
| Nbre d'ex | 4II | 79II | 2900 | 9I7 | 2I29 | I497 | 40I4 | 7657 | 5054 |
| Longueur, cm | ZEE mauritanienne | Sahara Occidental | rang de taille total pour 1988 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| mois | rang de taille annue | mois | rang de taille annuel | ||||||||
| IY | Y | YI | III | IY | YIII | IX | X | ||||
| I6 | 0,02 | 0,0I | 3,0 | I,0 | 0,03 | 0,0I | |||||
| I7 | 0,I | 0,02 | 0,05 | 28,0 | I,7 | 0,06 | 0,05 | ||||
| I8 | 0,3 | 0,2 | 0,2 | 23,0 | 27,5 | I,4 | I,3 | 0,5 | |||
| I9 | 0,2 | 0,8 | 0,7 | 0,7 | 7,0 | 44,3 | 3,3 | 2,4 | I,I | ||
| 20 | I,4 | I,5 | 2,I | I,8 | I2,0 | 20,4 | I5,2 | I,3 | 6,6 | 2,9 | |
| 2I | 3,3 | I,0 | I,4 | I,5 | 4,0 | 3,7 | I8,I | 3,0 | 8,I | 3,I | |
| 22 | 3,I | 0,5 | I,4 | I,3 | 9,0 | 0,4 | I2,8 | 7,6 | 8,4 | 3,0 | |
| 23 | 6,6 | 0,4 | I,9 | 2,0 | 8,0 | 0,7 | 5,2 | I0,8 | 7,4 | 3,3 | |
| 24 | 9,4 | 0,6 | 2,9 | 2,9 | 6,0 | 0,3 | 5,7 | I0,2 | 7,2 | 3,9 | |
| 25 | I3,0 | I,3 | 3,6 | 4,0 | 0,I | I0,0 | I0,0 | 0,02 | 8,6 | 5,I | |
| 26 | I2,7 | I,6 | 8,6 | 6,5 | I0,0 | I0,8 | 3,3 | 9,4 | 7,2 | ||
| 27 | II,I | 6,3 | 20,7 | I4,0 | 8,6 | I6,0 | I0,8 | I2,3 | I3,6 | ||
| 28 | I2,6 | I6,4 | 28,0 | 2I,6 | 9,5 | I0,9 | 24,0 | II,5 | I9,2 | ||
| 29 | II,2 | 20,0 | I7,8 | I7,7 | 9,3 | 2I,8 | 7,I | I5,2 | |||
| 30 | 6,I | I4,2 | 6,5 | 9,3 | 6,4 | 23,4 | 5,8 | 8,5 | |||
| 3I | 3,4 | II,9 | 2,2 | 6,0 | 2,4 | II,3 | 2,5 | 5,2 | |||
| 32 | 2,3 | 9,9 | I,0 | 4,5 | I,I | 2,8 | I,0 | 3,7 | |||
| 33 | 2,2 | 6,9 | 0,5 | 3,I | + | I,7 | 0,2 | 2,4 | |||
| 34 | 0,8 | 3,8 | 0,2 | I,6 | + | 0,9 | 0,0I | I,2 | |||
| 35 | 0,7 | I,7 | 0,2 | 0,8 | 0,07 | 0,0I | 0,6 | ||||
| 36 | 0,6 | 0,I | 0,3 | 0,2 | |||||||
| 37 | 0,2 | 0,02 | 0,I | 0,08 | |||||||
| 38 | 0,02 | 0,0I | 0,0I | ||||||||
| Nbre d'ex. | 623 | 7322 | 7243 | I5I88 | I00 | 680 | 200 | I200 | I478 | 3658 | I8846 |
| Lcm | 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | 1986 | 1987 | 1988* |
|---|---|---|---|---|---|---|---|---|---|---|
| I0 | ||||||||||
| I1 | 0,2 | 0,4 | 0,0I | |||||||
| I2 | 0,4 | 0,1 | 1,2 | 0,06 | 0,4 | 0,2 | ||||
| I3 | 0,2 | 0,3 | 0,5 | 3,8 | 0,03 | 0,I | 0,8 | 0,5 | ||
| I4 | 2,0 | 0,2 | 0,I | 5,9 | 0,2 | 0,I | 2,3 | I,3 | ||
| I5 | 2,4 | 0,3 | 0,I | 4,0 | I,0 | 0,I | 2,0 | I,I | 0,0I | |
| I6 | 0,6 | I,0 | 0,8 | 2,4 | I,7 | 0,2 | I,5 | I,3 | 0,02 | 0,0I |
| I7 | 0,5 | I,3 | 3,9 | I,5 | 2,6 | 0,4 | I,8 | 2,5 | 0,0I | 0,05 |
| I8 | I,5 | 0,3 | 3,5 | I,0 | 3,8 | I,5 | 3,6 | 3,2 | 0,02 | 0,5 |
| I9 | 3,3 | 0,2 | 4,3 | I,I | 3,4 | 5,4 | 6,3 | I,5 | 0,2 | I,I |
| 20 | 6,9 | I,I | 4,7 | 2,9 | 2,5 | 6,2 | 7,7 | I,I | I,0 | 2,9 |
| 2I | 5,3 | 6,I | 5,4 | 2,8 | 2,6 | I7,8 | 6,9 | I,I | 2,9 | 3,I |
| 22 | 2,4 | 7,8 | 6,7 | 0,7 | 5,I | I6,8 | 5,4 | I,5 | 6,9 | 3,0 |
| 23 | I,7 | 3,I | 9,8 | 2,I | 7,2 | I5,3 | 5,4 | 3,7 | II,3 | 3,3 |
| 24 | 2,8 | 2,0 | 8,9 | 3,2 | 4,8 | I2,I | 6,5 | 5,7 | I4,I | 3,9 |
| 25 | 5,9 | 3,3 | 5,7 | 4,6 | 3,I | 6,9 | 8,6 | 7,2 | I0,9 | 5,I |
| 26 | I4,I | 4,I | 3,9 | 5,I | I,8 | 4,I | 7,2 | 7,I | 7,6 | 7,2 |
| 27 | I2,I | 5,2 | 3,I | 2,7 | 2,0 | 3,6 | 5,I | 9,5 | 6,6 | I3,6 |
| 28 | I0,3 | I5,2 | 3,0 | 4,2 | 4,I | 4,8 | 5,9 | 9,6 | 8,I | I9,2 |
| 29 | 9,7 | 22,7 | 4,9 | 5,6 | 8,5 | I,4 | 5,6 | 6,0 | 9,6 | I5,2 |
| 30 | I2,0 | I4,9 | 8,0 | II,4 | 9,8 | 2,I | 6,8 | 8,5 | 8,I | 8,5 |
| 3I | 4,4 | 7,7 | 8,5 | I4,5 | 9,7 | 0,5 | 3,9 | 8,0 | 4,6 | 5,2 |
| 32 | I,4 | I,7 | 6,4 | I2,I | 9,3 | 0,I | 3,3 | I0,2 | 3,5 | 3,7 |
| 33 | 0,3 | 0,4 | 3,I | 4,5 | 6,9 | 0,I | I,6 | 5,9 | I,9 | 2,4 |
| 34 | 0,I | 0,2 | 2,I | I,5 | 4,5 | 0,07 | 0,8 | 2,3 | I,4 | I,2 |
| 35 | 0,I | 0,I | I,0 | 0,6 | 2,4 | 0,I | 0,4 | 0,7 | 0,6 | 0,6 |
| 36 | 0,I | 0,5 | 0,2 | I,2 | 0,03 | 0,I | 0,2 | 0,3 | 0,2 | |
| 37 | - | 0,I | 0,7 | 0,I | 0,08 | 0,I | 0,2 | 0,08 | ||
| 38 | - | 0,I | 0,4 | 0,03 | 0,0I | 0,06 | 0,0I | |||
| 39 | - | 0,I | 0,2 | 0,0I | 0,04 | |||||
| 40 | 0,I | 0,04 | 0,02 | |||||||
| 4I | 0,2 | 0,0I | ||||||||
| 42 | 0,I | |||||||||
| 43 | 0,06 | |||||||||
| 44 | 0,02 | |||||||||
| 45 | 0,04 | |||||||||
| 46 | 0,0I | |||||||||
| Nombre d.indi | 6345 | 5865 | 23667 | I3899 | 23899 | 20254 | 39022 | 43304 | 32490 | I8846 |
| Lmoy, cm | 25,50 | 26,90 | 25,50 | 25,80 | 27,84 | 23,47 | 25,46 | 27,50 | ||
| Prmoy, kg | 0,I55 | 0,I82 | 0,I55 | 0,I6I | 0,2I3 | 0,I23 | 0,I5I | 0,209 |
| Age, ans | I979 | I980 | I98I | I982 | I983 | I984 | I985 | I986 | I987 | I988* |
|---|---|---|---|---|---|---|---|---|---|---|
| I | 5,3 | 2,7 | 9,0 | 20,2 | I0,7 | 5,8 | 24,7 | I8,I | 2I,9 | 5.8 |
| 2 | 25,9 | 22,3 | 30,9 | 9,6 | I4,5 | 79,I | I4,7 | I4,4 | 28,0 | 8.0 |
| 3 | 34,9 | I8,4 | I8,5 | I2,9 | 9,2 | 9,5 | 20,8 | I2,5 | I0,6 | 37.2 |
| 4 | I4,6 | 26,0 | II,0 | I2,5 | I8,7 | 4,9 | 27,8 | I3,3 | I0,2 | 21.8 |
| 5 | I7,3 | 27,9 | I2,2 | I8,6 | I6,I | 0,5 | 9,0 | 29,5 | I9,I | 12.1 |
| 6 | I,7 | 2,I | I0,7 | I9,4 | I3,6 | 0,2 | I,9 | I0,7 | 8,0 | 9.5 |
| 7 | 0,3 | 0,5 | 6,0 | 6,0 | II,6 | 0,I | 0,8 | I,3 | I,6 | 5.2 |
| 8 | - | 0,I | I,7 | 0,8 | 5,6 | 0,I | 0,3 | 0,2 | 0,6 | 0.4 |
| 9 | ||||||||||
| 10 |
| Classe d'âge | I981 | I982 | I983 | I984 | I985 | I986 | I987 | I988 |
|---|---|---|---|---|---|---|---|---|
| I | 39,06 | 54,97 | 42,05 | I9,95 | I36,9I | II3,84 | I43,80 | 33,60 |
| 2 | 305,77 | 26,I2 | 56,98 | 271,34 | 8I,48 | 90,57 | I83,30 | 46,30 |
| 3 | I83,06 | 35,I0 | 36,I5 | 32,67 | II5,29 | 78,62 | 69,60 | 2I5,30 |
| 4 | I08,85 | 34,0I | 73,40 | I6,85 | I54,II | 83,65 | 67,00 | I25,I0 |
| 5 | I20,72 | 50,6I | 63,27 | I,72 | 49,89 | I85,53 | I25,40 | 70,00 |
| 6 | I06,88 | 52,79 | 53,44 | 0,69 | I0,53 | 67,30 | 52,50 | 55,00 |
| 7 | 59,37 | I6,33 | 45,58 | 0,34 | 4,43 | 8,I8 | I0,50 | 30,I0 |
| 8 | I6,82 | 2,I8 | 22,0I | 0,34 | I,66 | I,26 | 4,00 | 2,30 |
| Total | 989,5 | 272,I | 393,0 | 343,3 | 551,3 | 628,9 | 656,I | 578,7 |
| Classe d'âge | I98I | I982 | I983 | I984 | I985 | I986 | I987 | I988* |
|---|---|---|---|---|---|---|---|---|
| I | 0,0300 | 0,0096 | 0,0086 | 0,0063 | 0,0379 | 0,0380 | 0,0373 | 0,0087 |
| 2 | 0,2984 | 0,0I64 | 0,0I83 | 0,I062 | 0,0477 | 0,0476 | 0,I202 | 0,0226 |
| 3 | 0,4080 | 0,0764 | 0,0426 | 0,0I95 | 0,0008 | 0,0896 | 0,0706 | 0,3I56 |
| 4 | 0,30I4 | 0,I892 | 0,3547 | 0,0377 | 0,I840 | 0,I340 | 0,I568 | 0,2734 |
| 5 | 0,349I | 0,3527 | I,I249 | 0,0I86 | 0,2296 | 0,5705 | 0,4860 | 0,3849 |
| 6 | 0,6I98 | 0,4034 | I,480I | 0,0445 | 0,2327 | 0,9487 | 0,507I | 0,6840 |
| 7 | I,62I2 | 0,28I4 | I,3754 | 0,0430 | 0,7236 | 0,457I | 0,6I07 | I,III0 |
| 8 | I,II77 | 0,3397 | I,4I49 | 0,0437 | 0,482I | 0,795I | 0,7I33 | 0,4I43 |
| Classe d'âge | I98I | I982 | I983 | I984 | I985 | I986 | I987 | I988x |
|---|---|---|---|---|---|---|---|---|
| I | 400I,36 | 7666,82 | 6547,98 | 4255,23 | 489I,I7 | 4050,33 | 5207,4I | 5I36,I |
| 2 | I552,92 | 2I3I,09 | 4I67,54 | 3562,94 | 2320,76 | 2584,60 | 2I39,94 | 2753,I |
| 3 | 7I2,I8 | 632,37 | II50,52 | 2245,65 | I758,39 | I2I4,34 | I352,5I | I04I,4 |
| 4 | 548,07 | 259,9I | 32I,53 | 605,09 | I208,62 | 88I,26 | 609,32 | 69I,66 |
| 5 | 535,54 | 222,52 | II8,05 | I23,76 | 3I9,8I | 55I,85 | 423,00 | 285,88 |
| 6 | 295,69 | 207,3I | 85,82 | 2I,04 | 66,67 | I39,5I | I7I,I9 | I42,79 |
| 7 | 9I,24 | 87,3I | 76,0I | I0,72 | II,04 | 28,99 | 29,65 | 56,58 |
| 8 | 3I,50 | 9,90 | 36,I7 | I0,54 | 5,64 | 2,94 | I0,07 | 8,84 |
| Total | 7768,5 | II2I7,2 | I2503,6 | I0835,0 | I0582,I | 9453,8 | 9943,I | I0II6, |
| Classe d'âge | I98I | I982 | I983 | I984 | I985 | I986 | I987 | I988x |
|---|---|---|---|---|---|---|---|---|
| I | 240,08 | 460,0I | 39I,88 | 255,3I | 293,47 | 243,02 | 3I2,44 | 308,I7 |
| 2 | 2I4,30 | 294,09 | 575,I2 | 49I,69 | 320,27 | 356,67 | 295,3I | 379,93 |
| 3 | I78,05 | I58,09 | 287,63 | 56I,4I | 439,60 | 303,58 | 338,I3 | 260,36 |
| 4 | I86,35 | 88,37 | I09,32 | 205,73 | 4I0,93 | 299,63 | 207,I7 | 235,I6 |
| 5 | 2I4,22 | 89,0I | 47,22 | 49,5I | I27,92 | 220,74 | I69,20 | II4,35 |
| 6 | I33,06 | 93,29 | 38,62 | 9,47 | 30,00 | 62,78 | 77,04 | 64,26 |
| 7 | 50,I8 | 48,02 | 4I,80 | 5,90 | 6,07 | I5,95 | I6,3I | 3I,I2 |
| 8 | 2I,42 | 6,73 | 24,59 | 7,I7 | 3,83 | 2,00 | 6,85 | 6,0I |
| Total | I237,7 | I237,2 | I5I6,2 | I586,2 | I632,I | I504,4 | I422,4 | I399,4 |
| Modèle | Erreur |
|---|---|
| Rapport linéaire | 713 |
| Modèle de Beverton et Holt | 662 |
| Modèle de Ricker | 67161 |
| Modèle du Cushing | 660 |
| Modèle de Chapman | 1139 |
| Années | Données initiales | D'après Beverton et Helt | D'après Cushing | |||
|---|---|---|---|---|---|---|
| Recrutement Ri (abondan- ce des indi- vidus à l'age d'un an, APV) | Stock pater nel E (abon dance des classes d' age 2–8, APV) | recrutement précalcule | Ecart Ri et R'i % % | recrute- ment précalcu- lé | Ecart Ri:R'i, Ri:R'i, % | |
| 1982 | 7666.82 | 3735.65 | 6601.53 | 14.9 | 6724.08 | 12.3 |
| 1983 | 6547.98 | 3540.52 | 7105.46 | 8.5 | 7036.63 | 7.5 |
| 1984 | 4255.23 | 5919.46 | 4476.39 | 5.2 | 4553.30 | 7.0 |
| 1985 | 4891.17 | 6569.18 | 4245.18 | 13.2 | 4168.91 | 14.8 |
| 1986 | 4050.33 | 5685.28 | 4580.29 | 13.1 | 4711.15 | 16.3 |
| 1987 | 5207.41 | 5400.54 | 4726.48 | 9.2 | 4921.20 | 5.5 |
| Longueur (cm) | 1986 | 1987 | 1988 |
|---|---|---|---|
| I2 | I,3 | 0,66 | |
| I3 | 3,I | 3,28 | |
| I4 | 8,2 | I,3I | |
| I5 | 6,9 | 0,07 | |
| I6 | 8,2 | 0,66 | 0,I7 |
| I7 | I5,7 | 0,66 | 0,52 |
| I8 | 20,I | 3,28 | I,I6 |
| I9 | 9,4 | 3,28 | 5,2I |
| 20 | 6,9 | II,82 | 8,68 |
| 2I | 6,9 | 24,29 | 9,84 |
| 22 | 9,4 | 46,62 | 8,I0 |
| 23 | 23,3 | 67,63 | I2,I5 |
| 24 | 35,9 | 70,9I | I7,36 |
| 25 | 45,3 | 57,78 | 23,I5 |
| 26 | 44,7 | 36,II | 38,I9 |
| 27 | 59,8 | 34,I4 | 83,33 |
| 28 | 60,4 | 38,74 | I25,02 |
| 29 | 37,8 | 46,62 | I0I,27 |
| 30 | 53,5 | 45,3I | 53,24 |
| 3I | 50,3 | 39,40 | 34,72 |
| 32 | 64,2 | 44,65 | 24,88 |
| 33 | 37,I | 40,7I | I6,78 |
| 34 | I4,5 | 22,32 | 8,68 |
| 35 | 4,4 | I0,5I | 4,05 |
| 36 | I,3 | 3,94 | I,74 |
| 37 | 0,6 | I,3I | 0,46 |
| 38 | 0,33 | ||
| 39 | 0,26 | ||
| Total | 629,2 | 656,6 | 578,7 |
| Intervalle de classe de taille | Poids de poisson | Coefficient de mortalité naturelle | Coefficient de mortalité par pêche | Estimation du stock | ||
|---|---|---|---|---|---|---|
| Biomasse (millions d'ind.) | Abondance (milliers de tonne) | |||||
| I | 2 | 3 | 4 | 5 | 6 | 7 |
| I986–I987 rr. | ||||||
| I2–I3 | 4,I70 | 20,87 | 0,60 | 0,0056 | 3300,68 | 68,889 |
| I4–I5 | 8,250 | 3I,97 | 0,60 | 0,0I2I | 2846,54 | 9I,006 |
| I6–I7 | I2,6I0 | 46,42 | 0,60 | 0,0206 | 2430,I9 | II2,8I7 |
| I8–I9 | I8,480 | 64,67 | 0,60 | 0,0339 | 2050,29 | I32,586 |
| 20–2I | 24,955 | 87,I4 | 0,60 | 0,05I9 | I704,39 | I48,5I9 |
| 22–23 | 73,475 | II4,27 | 0,60 | 0,I793 | I39I,I5 | I58,973 |
| 24–25 | I04,945 | I46,5I | 0,60 | 0,3I8I | I07I,8I | I57,028 |
| 26–27 | 87,375 | I84,27 | 0,60 | 0,346I | 768,93 | I4I,693 |
| 28–29 | 9I,800 | 228,0I | O,60 | 0,5042 | 530,06 | I20,857 |
| 30–3I | 94,255 | 278,I4 | 0,60 | 0,8203 | 329,03 | 9I,5I4 |
| 32–33 | 93,330 | 335,I0 | 0,60 | I,7828 | I65,83 | 55,569 |
| 34–35 | 25,885 | 399,32 | 0,60 | I,9040 | 4I,09 | I6,408 |
| 36–37 | 3,575 | 47I,24 | 0,60 | I,2399 | 7,08 | 3,334 |
| 38–39 | 0,590 | 5I0,22 | 0,60 | 0,3000 | I,77 | 0,903 |
| Total | 643,73 | I66 37,84 | I300,I | |||
| I | 2 | 3 | 4 | 5 | 6 | 7 |
| I987–I988 | ||||||
| I2–I3 | 3,940 | 20,87 | 0,60 | 0,0055 | 3I74,9I | 66,264 |
| I4–I5 | I,400 | 3I,97 | 0,60 | 0,002I | 2738,I4 | 87,540 |
| I6–I7 | I,005 | 46,42 | 0,60 | 0,00I7 | 2343,69 | I08,802 |
| I8–I9 | 6,485 | 64,67 | 0,60 | 0,0I22 | I987,59 | I28,532 |
| 20–2I | 27,3I5 | 87,I4 | 0,60 | 0,0583 | I662,76 | I44,892 |
| 22–23 | 67,250 | II4,27 | 0,60 | 0,I683 | I354,47 | I54,78I |
| 24–25 | 84,600 | I46,5I | 0,60 | 0,2599 | I047,42 | I53,455 |
| 26–27 | 95,885 | I84,27 | 0,60 | 0,3829 | 767,55 | I4I,439 |
| 28–29 | I55,845 | 228,0I | 0,60 | 0,9394 | 52I,4I | II8,883 |
| 30–3I | 86,335 | 278,I4 | 0,60 | 0,9523 | 266,03 | 73,99I |
| 32–33 | 63,5I0 | 335,I0 | 0,60 | I,5390 | I25,29 | 4I,985 |
| 34–35 | 22,780 | 399,32 | 0,60 | I,8395 | 37,02 | I4,784 |
| 36–37 | 3,725 | 47I,24 | 0,60 | I,3854 | 6,8I | 3,2II |
| 38–39 | 0,590 | 5I0,22 | 0,60 | 0,4000 | I,47 | 0,753 |
| 620,754 | I6034,56 | I239,359 | ||||
| I | 2 | 3 | 4 | 5 | 6 | 7 |
| I986-I988 r.r. | ||||||
| I2–I3 | 4,I70 | 20,87 | 0,60 | 0,0057 | 32II,06 | 67,0I9 |
| I4–I5 | 8,250 | 3I,97 | 0,60 | 0,0I25 | 2769,I5 | 88,53I |
| I6–I7 | 8,637 | 46,42 | 0,60 | 0,0I45 | 2363,9I | I09,740 |
| I8–I9 | I4,443 | 64,67 | 0,60 | 0,027I | I997,7I | I29,I86 |
| 20–2I | 22,8I0 | 87,I4 | 0,60 | 0,0486 | I663,95 | I44,995 |
| 22–23 | 55,733 | II4,27 | 0,60 | 0,I383 | I359,56 | I55,362 |
| 24–25 | 83,467 | I46,5I | 0,60 | 0,2526 | I062,00 | I55,59I |
| 26–27 | 98,757 | I84,27 | 0,60 | 0,3883 | 780,3I | I43,790 |
| 28–29 | I36,630 | 228,0I | 0,60 | 0,79I6 | 528,94 | I20,600 |
| 30–3I | 92,I57 | 278,I4 | 0,60 | 0,9333 | 288,74 | 80,309 |
| 32–33 | 76,I07 | 335,I0 | 0,60 | I,7438 | I37,34 | 46,02I |
| 34–35 | 2I,487 | 399,32 | 0,60 | I,8299 | 35,04 | I3,993 |
| 36–37 | 3,II7 | 47I,24 | 0,60 | I,I5I0 | 6,5I | 3,068 |
| 38–39 | 0,590 | 5I0,22 | 0,60 | 0,3000 | I,77 | 0,903 |
| 626,37 | I6205,09 | I258,207 | ||||
![]()
![]()
![]()