![]()
![]()
![]()
Low temperature (e.g. chilling and freezing) injury can occur in all plants, but the mechanisms and types of damage vary considerably. Many fruit, vegetable and ornamental crops of tropical origin experience physiological damage when subjected to temperatures below about +12.5 °C, hence well above freezing temperatures. However, damage above 0 °C is chilling injury rather than freeze injury. Freeze injury occurs in all plants due to ice formation. Crop plants that develop in tropical climates, often experience serious frost damage when exposed to temperature slightly below zero, whereas most crops that develop in colder climates often survive with little damage if the freeze event is not too severe. Some exceptions are lettuce, which originated in a temperate climate, but can be damaged at temperatures near 0 °C and some subtropical fruits trees that can withstand temperatures to -5 to -8 °C. Species or varieties exhibit different frost damage at the same temperature and phenological stage, depending on antecedent weather conditions, and their adaptation to cold temperatures prior to a frost night is called "hardening". During cold periods, plants tend to harden against freeze injury, and they lose the hardening after a warm spell. Hardening is most probably related to an increase in solute content of the plant tissue or decreases in ice-nucleation active (INA) bacteria concentrations during cold periods, or a combination. During warm periods, plants exhibit growth, which reduces solute concentration, and INA bacteria concentration increases, which makes the plants less hardy.
Frost damage occurs when ice forms inside the plant tissue and injures the plant cells. It can occur in annuals (grasses and legumes of forage and silage crops; cereals; oil and root crops; horticultural; and ornamental crops) multi-annuals and perennials (deciduous and evergreen fruit trees). Frost damage may have a drastic effect upon the entire plant or affect only a small part of the plant tissue, which reduces yield, or merely product quality.
In this chapter, a short discussion of the mechanisms, types and symptoms of freeze injury is presented. For interested readers, Levitt (1980), Sakai and Larcher (1987) and Li (1989) provide extensive reviews of both freezing and chilling injury. Later in this chapter, a short discussion of hardening, sensitivity, kind-of-damage and critical damage temperatures of important crops are presented.
Direct frost damage occurs when ice crystals form inside the protoplasm of cells (intracellular freezing), whereas indirect damage can occur when ice forms inside the plants but outside of the cells (i.e. extracellular freezing). It is not cold temperature but ice formation that actually injures the plants (Westwood, 1978). It is believed that intracellular ice formation causes a "mechanical disruption of the protoplasmic structure" (Levitt, 1980). The extent of damage due to intracellular freezing depends mainly on how fast the temperature drops and to what level it supercools before freezing. There is little or no evidence that the duration of the freezing affects injury. In fact, Levitt (1980) states that freeze injury seems to be independent of time for short periods (e.g. 2-24 hours).
Direct intracellular freeze injury is associated with rapid cooling. For example, Siminovitch, Singh and de la Roche (1978) observed intracellular freezing and cell death when winter rye plants were cooled at 8 °C per minute to -12 °C when the supercooled water froze inside the cells. When plants were cooled to -12 °C over 23 minutes, ice formation was extracellular and the plants fully recovered after thawing. In climate chamber studies to determine critical temperatures, plant cuttings are typically cooled at a rate of between 1.0 and 2.0 °C h-1. This is a slower rate than in the rye plant experiment and a slower rate than some of the rates that often occur in nature. Indeed, Levitt (1980) reports that, in nature, freeze injury results from extracellular ice crystal formation and there is no evidence of intracellular freezing.
Although the evidence is not strong, it seems that the rate of thawing after a freeze is also partially related to the amount of damage. Citrus growers in southern California commonly believe that slowing the warming process after a freeze night can reduce frost damage. In fact, growers justify operating wind machines longer into the morning following a freeze night in order to slow the thawing process. Yoshida and Sakai (1968) suggested that thawing rate will slow the rehydration of cells in plants that experience extracellular freezing and that might reduce the damage due to fast thawing.
Levitt (1980) proposed that cells were gradually killed as a result of growth of the extracellular ice mass. Recall that the saturation vapour pressure is lower over ice than over liquid water. As a result of extracellular ice formation, water will evaporate from the liquid water inside the cells and will pass through the semipermeable cell membranes and deposit on the ice crystals outside of the cells. As water is removed from the cells, the solute concentration increases and reduces the chances of freezing. However, as ice continues to grow, the cells become more desiccated. Typically, in injured plants, the extracellular ice crystals are much larger than the surrounding dead cells, which have collapsed because of desiccation. Therefore, the main cause of frost damage to plants in nature is extracellular ice crystal formation that causes secondary water stress to the surrounding cells. In fact, there is a close relationship between drought-tolerant plants and freeze-tolerant plants.
Note that antitranspirants are often promoted as a method of freeze protection. It is argued that the frost damage occurs because of cell dehydration and the antitranspirants are purported to reduce water loss from the plants and provide freeze protection. However, the cell desiccation results from evaporation of cellular water in response to a vapour pressure gradient caused by extracellular ice formation and not because of transpiration. There is no evidence that antitranspirants reduce desiccation due to extracellular ice crystal formation.
Plants resist low temperatures by avoidance or tolerance. Strategies to avoid low temperatures include:
snow retention throughout the winter, which protects both the aerial and subterranean parts of the plants (Ventskevich, 1958);
the biophysical effect of dense canopies, which shield part of the plant from the cold sky;
bulky organs (e.g. trunks or big fruits) with high heat capacity that lag their temperature behind air temperature, which may save them from damaging temperatures (Turrell and Austin, 1969); and
artificial frost protection methods, which modify the microclimate of the plants (e.g. foams, covers and fogging).
Tolerance of low temperature can be achieved by:
avoiding freezing through a decrease of the freezing point or an increase in the degree of supercooling (Burke et al., 1976);
tolerance of extracellular freezing by reducing the amount of ice formed due to an increase of the concentration of solutes in the protoplasm (Li and Palta, 1978);
tolerance of a higher degree of desiccation due to the plasmolysis of the protoplasm (Gusta, Burke and Kapoor, 1975); or
increasing the permeability of the plasma membrane to avoid intracellular freezing (Alden and Hermann, 1971; Levitt, 1980).
The temperature at which freezing occurs can fluctuate considerably depending on to what extent the plants have hardened. However there are plants (e.g. many C4 plants, palm tree leaves and tomato plants) that have very little or no hardening capacity (Larcher, 1982; Olien, 1967). Hardening involves both mechanisms of avoidance and tolerance of freezing. The accumulation of sugars or sugar alcohols lower the freezing temperature of tissues (e.g. in olive and citrus tree leaves) and supercooling increases in many deciduous and evergreen fruit trees in response to low air temperature. Some cells may harden by increasing the proportion of unsaturated fatty acids of plasma membrane lipids, which would increases membrane stability during desiccation. Since hardening is an active process that depends on assimilate level in the tissues, all conditions that deplete the pool of assimilates in the tissues reduce hardening.
Although cold temperatures cause fruit plants to harden against frost damage, hardiness is quickly lost during a few warm days. The fruit buds will regain hardiness but at a much slower rate than they lose it. This is the basis for the practice of cooling crops with sprinklers during daytime warm periods to reduce temperature and avoid the loss of hardening.
In the past, researchers have attributed fluctuations in freeze sensitivity to physiological changes, but the contribution of INA bacteria to the sensitivity, which might also be a factor to consider, has generally been ignored. For example, a rapid increase in ice-nucleating bacterial concentration might also occur during warm periods. As cold temperatures return, the concentration of bacteria might decline slowly.
Plants fall into four freeze-sensitivity categories: (1) tender; (2) slightly hardy; (3) moderately hardy; and (4) very hardy (Levitt, 1980). Tender plants are those that have not developed avoidance of intracellular freezing (e.g. mostly tropical plants). Slightly hardy plants include most of the subtropical fruit trees, deciduous trees during certain periods, and fruit and vegetable horticultural [truck] crops that are sensitive to freezing down to about -5 °C. Moderately hardy plants include those that can accumulate sufficient solutes to resist freeze injury to temperatures as low as -10 °C mainly by avoiding dehydration damage, but they are less able to tolerate lower temperatures. Very hardy plants are able to avoid intracellular freezing as well as avoid damage due to cell desiccation.
FIGURE 4.1
Typical 10 percent and 90 percent bud kill
temperatures for cherry trees corresponding to average dates observed at the
Washington State University, Prosser Research and Extension Centre (Proebsting
and Mills, 1978)
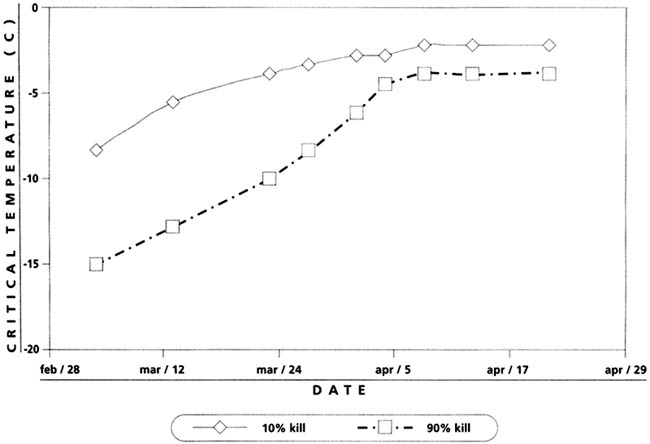
Although freeze sensitivity categories give general information about the cold that a plant organ can endure before frost damage occurs, hardening and phenological stage are almost as important. For example, temperature that produces both 10 percent (T10) and 90 percent (T90) bud kill increases as the season progresses from first swelling to post bloom (Figure 4.1). In addition, the temperatures that produce T90 bud kill in deciduous trees increases more rapidly and approach the temperatures that produce T10 kill.
Wang and Wallace (2003) presented a list of fresh fruits and vegetables by freeze susceptibility categories (Table 4.1.) showing relative sensitivities when exposed to freezing temperatures. Caplan (1988) gave a list of freeze-tolerance groupings for annual flowers (Table 4.2). Table 4.4 provides an extensive list of some of these and other crops.
TABLE 4. 1
Susceptibility of fresh fruits and vegetables
to freezing injury
|
MOST SUSCEPTIBLE |
MODERATELY SUSCEPTIBLE |
LEAST SUSCEPTIBLE |
|
Apricots |
Apples |
Beets |
|
Asparagus |
Broccoli |
Brussels sprouts |
|
Avocados |
Carrots |
Cabbage, mature and savoury |
|
Bananas |
Cauliflower |
Dates |
|
Beans, snap |
Celery |
Kale |
|
Berries (except cranberries) |
Cranberries |
Kohlrabi |
|
Cucumbers |
Grapefruit |
Parsnips |
|
Eggplant |
Grapes |
Rutabagas |
|
Lemons |
Onion (dry) |
Salsify |
|
Lettuce |
Oranges |
Turnips |
|
Limes |
Parsley |
|
|
Okra |
Pears |
|
|
Peaches |
Peas |
|
|
Peppers, sweet |
Radishes |
|
|
Plums |
Spinach |
|
|
Potatoes |
Squash, Winter |
|
|
Squash, Summer |
|
|
|
Sweet potatoes |
|
|
|
Tomatoes |
|
|
SOURCE: Wang and Wallace, 2003.
TABLE 4.2
Categories for freeze hardiness of various
annual flowers
|
HARDY |
TOLERANT |
TENDER |
SENSITIVE |
|
Cornflower |
Bells of Ireland (Moluccella) |
Aster |
Ageratum |
|
Ornamental cabbage |
|
Nicotiana |
Balsam |
|
Pansy |
Coreopsis |
Petunia |
Begonia |
|
Primrose |
Pinks (Dianthus) |
Scabiosa |
Cockscomb |
|
Violet |
Pot Marigold (Calendula) |
Statice |
Impatiens |
|
|
Snapdragon |
Sweet alyssum |
Lobelia |
|
|
Stock (Matthiola incana) |
Verbena |
Marigold |
|
|
Sweet pea |
|
Moss rose (Portulaca) |
|
|
Torenia |
|
Periwinkle (Vinca) |
|
|
|
|
Phlox, annual |
|
|
|
|
Salpiglossis |
|
|
|
|
Salvia |
|
|
|
|
Zinnia |
SOURCE: Based on Purdue University publication HO-14, as cited by Caplan, 1988.
There are numerous studies on critical damage (Tc) temperatures for a variety of crops. These numbers were obtained using a range of methods and one should use caution when attempting to use published critical temperatures to manage starting and stopping temperatures for active protection methods. For example, some researchers have compared long-term commercial damage records with temperature measurements from standard shelters. In some instances, the temperature sensor, shielding, mounting height, etc., are not reported. These factors can affect the results and it is difficult to apply information from one location to another because insufficient information was supplied. Also, there are always microclimate differences, even within a research plot, that can affect results. For example, the authors have observed spatial differences of 1.0 °C or more within a couple of hundred metres in an orchard during a freeze night, measured at the same height above the ground on flat terrain. Therefore, it is somewhat questionable that Tc values from shelter temperatures are universally applicable.
Many researchers have cut small branches from trees and placed them in climate control chambers where the excised branches were cooled to a range of subzero temperatures and the damage was observed. While this process is more standardized than field measurements, the microclimate inside of a climate control chamber is not the same as branches exposed to the sky. For example, one could determine the amount of damage for branches exposed to 30 minutes at a range of temperatures, but within a tree the uncut branches will have a broad range of temperatures. Branches in the upper tree canopy will be exposed to the sky and therefore will probably be colder than air temperature. Conversely, branches embedded in the canopy are likely to be warmer and thus less prone to damage. In deciduous trees before the leaves develop fully, there is usually an inversion from the ground upward, so the coldest air temperatures are near the bottom of the trees. When trees have most of their leaves expanded, however, the minimum temperature, on radiation frost nights, rises to the height where most leaves are. In any case, using temperatures from a weather shelter only provides a rough guideline for expected damage.
In addition to variations of plant part temperatures within a tree and spatially throughout an orchard, vineyard or field, there are also variations in INA bacteria, which are now known to be a factor determining how low plants will supercool. To our knowledge, no researchers have taken into account differences in ice-nucleating bacteria concentrations when evaluating critical temperatures. For example, almond trees are known to have large concentrations of INA bacteria. If one block of an orchard was sprayed with bactericides that reduced the INA bacterial population and another was not sprayed, then the critical temperature for the block with fewer bacteria should have lower critical temperatures. This is another factor that complicates the decision about starting active protection methods. In general, the best approach is to use the published values as a guideline and start and stop protection based on an additional safety factor correction to published Tc values. It is better to err on the high side.
It is important to note that critical temperatures determined in a laboratory are obtained in carefully controlled freezers with slow air movement. The air temperature in the freezer is lowered in small, predetermined steps and held for 20 to 30 minutes or more after each step to allow the buds to come into equilibrium. This practice has given rise to the common misconception that buds have to be at a temperature for 20 to 30 minutes or so before damage will occur. The truth is that for short periods (2 to 24 h) the duration plant tissue is below a particular temperature is less important than how low the temperature goes (Levitt, 1980). Plant tissues cool at a rate dependent on the radiation balance and the temperature difference between the tissue and its environment. Therefore, if the air suddenly drops several degrees the tissue can rapidly cool below critical levels and result in freeze injury. If the plant tissue contains supercooled water, mechanical shock or agitation of the leaves and buds by wind machines or helicopters could initiate ice crystal formation, resulting in damage even if the tissues are above the chamber-determined critical temperature values. However, the chamber values provide guidelines as to when freeze protection measures need to be implemented.
Vegetable crop damage symptoms vary widely and can sometimes be confused with biotic damage. Table 4.3 shows a list of frost damage symptoms of some vegetable crops. Species differ greatly in their resistance to frost, but the maximum level of resistance is only attained when environmental conditions allow hardening to take place. Variety is often as important as species in defining resistance to frost, specially when there are winter and spring types. In general, also, there is an inverse relation between earliness of a variety and frost resistance.
Field experiments on critical damage temperatures for fresh fruits and vegetable crops are somewhat limited, but the highest freezing temperatures from studies on fruit and vegetable storage are provided in Table 4.4. Although the critical damage temperatures might be slightly higher than the air temperatures at which damage is expected under field conditions, the information in Table 4.4 can be useful as a guide.
During severe frost events with no snow, the young leaves of grasses and winter cereals seedlings may be damaged, but recovery is possible if the tillering node is not affected. However, if this meristem is damaged, winterkill will occur.
TABLE 4.3
Frost damage symptoms for vegetable crops
(Caplan, 1988)
|
CROP |
SYMPTOMS |
|
Artichoke |
Epidermis becomes detached and forms whitish to light tan blisters. When blisters are broken, underlying tissue turns brown. |
|
Asparagus |
Tip becomes limp and dark and the rest of the spear is water soaked. Thawed spears become mushy. |
|
Beet |
External and internal water soaking and sometimes blackening of conductive tissue. |
|
Broccoli |
The youngest florets in the centre of the curd are most sensitive to freezing injury. They turn brown and give off strong odour. |
|
Cabbage |
Leaves become water soaked, translucent and limp. Upon thawing the epidermis separates. |
|
Carrot |
Blistered appearance, jagged length-wise cracks. Interior becomes water soaked and darkens upon thawing. |
|
Cauliflower |
Curds turn brown and have a strong off-odour when cooked. |
|
Celery |
Leaves and petioles appear wilted and water soaked upon thawing. Petioles freeze more readily than leaves. |
|
Garlic |
Thawed cloves appear greyish yellow and water soaked. |
|
Lettuce |
Blistering of dead cells of the separated epidermis on outer leaves, and become tan with increased susceptibility to physical damage and decay. |
|
Onion |
Thawed bulbs are soft, greyish yellow and water soaked in cross- section. Damage is often limited to individual scales. |
|
Pepper, bell |
Dead, water-soaked tissue in part or all of pericarp surface with pitting, shrivelling and decay follow thawing. |
|
Potato |
Freezing injury may not be externally evident, but shows as grey or bluish-grey patches beneath the skin. Thawed tubers become soft. |
|
Radish |
Thawed tissues appear translucent and the roots soften and shrivel. |
|
Sweet potato |
A yellowish-brown discoloration of the vascular ring and a yellowish green, water-soaked appearance of other tissues. Roots soften and become susceptible to decay. |
|
Tomato |
Water soaked and soft upon thawing. In partially frozen fruits, the margin between healthy and dead tissue is distinct, especially in green fruits. |
|
Turnip |
Small water-soaked spots or pitting on the surface. Injured tissues appear tan or grey and give off an objectionable odour. |
TABLE 4.4
The highest freezing temperature for fresh
fruits and vegetables
|
COMMON NAME |
SCIENTIFIC NAME |
TEMPERATURE |
||||
|
Acerola; Barbados cherry |
Malpighia glabra |
-1.4 |
||||
|
Apple |
Malus pumila |
-1.5 |
||||
|
Apricot |
Prunus armeniaca |
-1.1 |
||||
|
Artichoke - globe |
Cynara scolymus |
-1.2 |
||||
|
Artichoke - Jerusalem |
Helianthus tuberosus |
-2.5 |
||||
|
Asian pear, Nashi |
Pyrus serotina; P. pyrifolia |
-1.6 |
||||
|
Asparagus, green, white |
Asparagus officinalis |
-0.6 |
||||
|
Avocado |
Persea americana |
|
||||
| |
cv. Fuerte, Hass |
|
-1.6 |
|||
|
cv. Fuchs, Pollock |
|
-0.9 |
||||
|
cv. Lula, Booth |
|
-0.9 |
||||
|
Banana |
Musa paradisiaca var. sapientum |
-0.8 |
||||
|
Barbados cherry |
Malpighia glabra |
-1.4 |
||||
|
Beans |
|
|
||||
| |
Snap; Wax; Green |
Phaseolus vulgaris |
-0.7 |
|||
|
Lima beans |
Phaseolus lunatus |
-0.6 |
||||
|
Beet, bunched |
Beta vulgaris |
-0.4 |
||||
|
Beet, topped |
|
-0.9 |
||||
|
Berries |
|
|
||||
| |
Blackberries |
Rubus spp. |
-0.8 |
|||
|
Blueberries |
Vaccinium corymbosum |
-1.3 |
||||
|
Cranberry |
Vaccinium macrocarpon |
-0.9 |
||||
|
Dewberry |
Rubus spp. |
-1.3 |
||||
|
Elderberry |
Sambucus spp. |
-1.1 |
||||
|
Loganberry |
Rubus spp. |
-1.7 |
||||
|
Raspberries |
Rubus idaeus |
-0.9 |
||||
|
Strawberry |
Fragaria spp. |
-0.8 |
||||
|
Broccoli |
Brassica oleracea var. italica |
-0.6 |
||||
|
Brussels-sprouts |
Brassica oleracea var. gemmifera |
-0.8 |
||||
|
Cabbage |
|
|
||||
| |
Chinese; Napa |
Brassica campestris var. pekinensis |
-0.9 |
|||
|
Common, early crop |
Brassica oleracea var. capitata |
-0.9 |
||||
|
Late crop |
- " - |
-0.9 |
||||
|
Cactus pear, prickly pear fruit |
Opuntia spp. |
-1.8 |
||||
|
Carambola, Starfruit |
Averrhoa carambola |
-1.2 |
||||
|
Carrots, topped |
Daucus carota |
-1.4 |
||||
|
Cauliflower |
Brassica oleracea var. botrytis |
-0.8 |
||||
|
Celeriac |
Apium graveolens var. rapaceum |
-0.9 |
||||
|
Celery |
Apium graveolens var. dulce |
-0.5 |
||||
|
Cherimoya; custard apple |
Annona cherimola |
-2.2 |
||||
|
Cherry, sour |
Prunus cerasus |
-1.7 |
||||
|
Cherry, sweet |
Prunus avium |
-2.1 |
||||
|
Chicory |
see Endive |
|
||||
|
Chilies |
see Pepper |
|
||||
|
Citrus |
|
|
||||
| |
Calamondin orange |
Citrus reticulata x Fortunella spp. |
-2.0 |
|||
|
California & Arizona, (USA) dry areas |
|
-1.1 |
||||
|
Florida (USA), humid areas |
|
-1.1 |
||||
|
Lemon |
Citrus limon |
-1.4 |
||||
|
Lime, Mexican, |
Citrus aurantifolia; |
-1.6 |
||||
|
Orange |
Citrus sinensis |
|
||||
|
California & Arizona (USA), dry areas |
|
-0.8 |
||||
|
Florida (USA), humid areas |
|
-0.8 |
||||
|
Blood orange |
|
-0.8 |
||||
|
Seville; sour |
Citrus aurantium |
-0.8 |
||||
|
Pummelo |
Citrus grandis |
-1.6 |
||||
|
Tangelo, Minneola |
Citrus reticulata x paradisi |
-0.9 |
||||
|
Tangerine |
Citrus reticulata |
-1.1 |
||||
|
Chives |
Allium schoenoprasum |
-0.9 |
||||
|
Coconut |
Cocos nucifera |
-0.9 |
||||
|
Collards, kale |
Brassica oleracea var. acephala |
-0.5 |
||||
|
Corn, sweet and baby (maize) |
Zea mays |
-0.6 |
||||
|
Cucumber, slicing |
Cucumis sativus |
-0.5 |
||||
|
Currants |
Ribes spp. |
-1.0 |
||||
|
Custard apple |
see Cherimoya |
|
||||
|
Dasheen |
see Taro |
|
||||
|
Date |
Phoenix dactylifera |
-15.7 |
||||
|
Dill |
Anethum graveolens |
-0.7 |
||||
|
Eggplant |
Solanum melongena |
-0.8 |
||||
|
Endive, Escarole |
Cichorium endivia |
-0.1 |
||||
|
Fennel |
Foeniculum vulgare |
-1.1 |
||||
|
Fig |
Ficus carica |
-2.4 |
||||
|
Garlic bulb |
Allium sativum |
-2.0 |
||||
|
Gooseberry |
Ribes grossularia |
-1.1 |
||||
|
Grape |
Vitis vinifera |
fruit |
-2.7 |
|||
| |
|
stem |
-2.0 |
|||
|
Grape, American |
Vitis labrusca |
-1.4 |
||||
|
Horseradish |
Armoracia rusticana |
-1.8 |
||||
|
Jujube; Chinese date |
Ziziphus jujuba |
-1.6 |
||||
|
Kale |
Brassica oleracea var. acephala |
-0.5 |
||||
|
Kiwano |
see African horned melon |
|
||||
|
Kiwifruit; |
Actinidia chinensis |
-0.9 |
||||
|
Kohlrabi |
Brassica oleracea var. gongylodes |
-1.0 |
||||
|
Leafy greens |
|
|
||||
| |
Cool season |
various genera |
-0.6 |
|||
|
Warm season |
various genera |
-0.6 |
||||
|
Leek |
Allium porrum |
-0.7 |
||||
|
Lettuce |
Lactuca sativa |
-0.2 |
||||
|
Longan |
Dimocarpus longan |
-2.4 |
||||
|
Loquat |
Eriobotrya japonica |
-1.9 |
||||
|
Mango |
Mangifera indica |
-1.4 |
||||
|
Melons |
|
|
||||
| |
Cantaloupes, netted melons |
Cucurbita melo var. reticulatus |
-1.2 |
|||
|
Casaba |
Cucurbita melo |
-1.0 |
||||
|
Crenshaw |
Cucurbita melo |
-1.1 |
||||
|
Honeydew, orange-flesh |
Cucurbita melo |
-1.1 |
||||
|
Persian |
Cucurbita melo |
-0.8 |
||||
|
Mombin |
see Spondias |
|
||||
|
Mushrooms |
Agaricus, other genera |
-0.9 |
||||
|
Nashi |
see Asian pear |
|
||||
|
Nectarine |
Prunus persica |
-0.9 |
||||
|
Okra |
Abelmoschus esculentus |
-1.8 |
||||
|
Olives, fresh |
Olea europea |
-1.4 |
||||
|
Onions |
Allium cepa |
|
||||
| |
Mature bulbs, dry |
|
-0.8 |
|||
|
Green onions |
|
-0.9 |
||||
|
Papaya |
Carica papaya |
-0.9 |
||||
|
Parsley |
Petroselinum crispum |
-1.1 |
||||
|
Parsnip |
Pastinaca sativa |
-0.9 |
||||
|
Peach |
Prunus persica |
-0.9 |
||||
|
Pear, European |
Pyrus communis |
-1.7 |
||||
|
Peas (pod, snow, snap, sugar) |
Pisum sativum |
-0.6 |
||||
|
Peppers |
|
|
||||
| |
Bell Pepper, Paprika |
Capsicum annuum |
-0.7 |
|||
|
Hot peppers, Chiles |
Capsicum annuum and C. frutescens |
-0.7 |
||||
|
Persimmon, kaki |
Diospyros kaki |
|
||||
| |
Fuyu |
|
-2.2 |
|||
|
Hachiya |
|
-2.2 |
||||
|
Pineapple |
Ananas comosus |
-1.1 |
||||
|
Plantain |
Musa paradisiaca var. paradisiaca |
-0.8 |
||||
|
Plums and Prunes |
Prunus domestica |
-0.8 |
||||
|
Pomegranate |
Punica granatum |
-3.0 |
||||
|
Potato, early crop |
Solanum tuberosum |
-0.8 |
||||
| |
late crop |
|
-0.8 |
|||
|
Pumpkin |
Cucurbita maxima |
-0.8 |
||||
|
Quince |
Cydonia oblonga |
-2.0 |
||||
|
Radish |
Raphanus sativus |
-0.7 |
||||
|
Rhubarb |
Rheum rhaponticum |
-0.9 |
||||
|
Rutabaga |
Brassica napus var. napobrassica |
-1.1 |
||||
|
Salsify, vegetable oyster |
Trapopogon porrifolius |
-1.1 |
||||
|
Sapotes |
|
|
||||
| |
Caimito, star apple |
Chrysophyllum cainito |
-1.2 |
|||
|
Canistel, eggfruit |
Pouteria campechiana |
-1.8 |
||||
|
Black sapote |
Diospyros ebenaster |
-2.3 |
||||
|
White sapote |
Casimiroa edulis |
-2.0 |
||||
|
Shallot |
Allium cepa var. ascalonicum |
-0.7 |
||||
|
Spinach |
Spinacia oleracea |
-0.3 |
||||
|
Squash |
|
|
||||
| |
Summer (soft rind); courgette |
Cucurbita pepo |
-0.5 |
|||
|
Winter (hard rind); calabash |
Cucurbita moschata; C. maxima |
-0.8 |
||||
|
Star-apple |
see Sapotes |
|
||||
|
Starfruit |
see Carambola |
|
||||
|
Strawberry |
see Berries |
|
||||
|
Sweet potato, yam [in USA] |
Ipomoea batatas |
-1.3 |
||||
|
Tamarind |
Tamarindus indica |
-3.7 |
||||
|
Taro, cocoyam, |
Colocasia esculenta |
-0.9 |
||||
|
Tomato |
Lycopersicon esculentum |
|
||||
| mature green | |
-0.5 |
||||
| firm ripe | |
-0.5 |
||||
|
Turnip root |
Brassica campestris var. rapifera |
-1.0 |
||||
|
Watercress; |
Lepidium sativum |
-0.3 |
||||
|
Watermelon |
Citrullus vulgaris |
-0.4 |
||||
|
Witloof chicory (endive) |
|
-0.1 |
||||
|
Yam |
Dioscorea spp. |
-1.1 |
||||
SOURCE: From Whiteman, 1957, as reported in the University of California, Davis, Postharvest web page: http://postharvest.ucdavis.edu/Produce/Storage/propa.shtml.
NOTE: Some taxonomic names may have changed since 1957.
In early and late winter and in early spring, plants may be less hardy, which enhances damage. Snow retention reduces this type of damage (Ventskevich, 1958). Later in the season, during flowering and initial grain growth of cereals, frost damage reduces the number of kernels per spike. The visual result is that a bleached and thinner band forms on the spikes for each frost event, awns become curly, and because the weight of grain is less, spikes are upright near maturity (Figure 4.2).
For cereal crops, the relative resistance to freezing of cereals is (from most resistant): Rye > Bread wheat > Triticale > Barley > Oats and Durum wheat. During the winter, the critical temperatures change in relation to the degree of hardening. However, when hardening is complete, no plant destruction occurs with temperatures that range between -40 to -45 °C for rye, up to above -10 °C for durum wheat (Lecomte, 1989).
Freezing can damage many field crops including annual forage and silage crops, which lose leaf area and hence decrease dry matter production. Table 4.5 shows the critical temperatures for many field crops relative to phenological stages.
FIGURE 4.2
Frost damage to wheat crop

The terminal third of the spike is thinned and the awns are curled (above); and later the spikes remain upright since the grain weight is small (right).

Photos: J P de Melo-Abreu (ISA)
TABLE 4. 5
A range of critical damage temperatures
(°C) for grain, forage and silage crops
|
CROP GER |
GERMINATION |
FLOWERING |
FRUITING |
|
Spring wheat |
-9, -10 |
-1, -2 |
-2, -4 |
|
Oats |
-8, -9 |
-1, -2 |
-2, -4 |
|
Barley |
-7, -8 |
-1, -2 |
-2, -4 |
|
Peas |
-7, -8 |
-2, -3 |
-3, -4 |
|
Lentils |
-7, -8 |
-2, -3 |
-2, -4 |
|
Vetchling |
-7, -8 |
-2, -3 |
-2, -4 |
|
Coriander |
-8, -10 |
-2, -3 |
-3, -4 |
|
Poppies |
-7, -10 |
-2, -3 |
-2, -3 |
|
Kok-saghyz |
-8, -10 |
-3, -4 |
-3, -4 |
|
Lupin |
-6, -8 |
-3, -4 |
-3, -4 |
|
Spring vetch |
-6, -7 |
-3, -4 |
-2, -4 |
|
Beans |
-5, -6 |
-2, -3 |
-3, -4 |
|
Sunflower |
-5, -6 |
-2, -3 |
-2, -3 |
|
Saff lower |
-4, -6 |
-2, -3 |
-3, -4 |
|
White mustard |
-4, -6 |
-2, -3 |
-3, -4 |
|
Flax |
-5, -7 |
-2, -3 |
-2, -4 |
|
Hemp |
-5, -7 |
-2, -3 |
-2, -4 |
|
Sugar-beet |
-6, -7 |
-2, -3 |
- |
|
Fodder-beet |
-6, -7 |
- |
- |
|
Carrot |
-6, -7 |
- |
- |
|
Turnip |
-6, -7 |
- |
- |
|
Cabbage |
-5, -7 |
-2, -3 |
-6, -9 |
|
Soybeans |
-3, -4 |
-2, -3 |
-2, -3 |
|
Italian millet |
-3, -4 |
-1, -2 |
-2, -3 |
|
European yellow lupine |
-4, -5 |
-2, -3 |
- |
|
Corn [maize] |
-2, -3 |
-1, -2 |
-2, -3 |
|
Millet |
-2, -3 |
-1, -2 |
-2, -3 |
|
Sudan grass |
-2, -3 |
-1, -2 |
-2, -3 |
|
Sorghum |
-2, -3 |
-1, -2 |
-2, -3 |
|
Potato |
-2, -3 |
-1, -2 |
-1, -2 |
|
Rustic tobacco |
-2, -3 |
- |
-2, -3 |
|
Buckwheat |
-1, -2 |
-1, -2 |
-0.5, -2 |
|
Castor plant |
-1, -1.5 |
-0.5, -1 |
-2 |
|
Cotton |
-1, -2 |
-1, -2 |
-2, -3 |
|
Melons |
-0.5, -1 |
-0.5, -1 |
-1 |
|
Rice |
-0.5, -1 |
-0.5, -1 |
-0.5, -1 |
|
Sesame |
-0.5, -1 |
-0.5, -1 |
- |
|
Hemp mallow |
-0.5, -1 |
- |
- |
|
Peanut |
-0.5, -1 |
- |
- |
|
Cucumber |
-0.5, -1 |
- |
- |
|
Tomato |
0, -1 |
0, -1 |
0, -1 |
|
Tobacco |
0, -1 |
0, -1 |
0, -1 |
SOURCE: After Ventskevich, 1958.
The limits of the natural distribution of many plants including some deciduous fruit trees are related to the minimum temperature at which supercooling can occur (i.e. homogeneous nucleation point), which is near -40 °C. Below the homogeneous nucleation point, freezing is intracellular and lethal (Burke et al., 1976; Weiser et al., 1979; Ikeda, 1982).
Generally, deciduous crop sensitivity to freezing temperature increases from first bloom to small-nut or -fruit stages, and this is when a crop is most likely to be damaged. Sensitivity is also higher when warm weather has preceded a freeze night than if the cold temperatures preceded the freeze. Plants are known to harden against freezing when exposed to cold temperatures over long periods and this hardening is lessened if exposed to warm temperatures. Considerable information on sensitivity of deciduous fruit trees relative to phenological stages are provided on the Washington State University - Prosser Research and Extension Centre WEB site (http://fruit.prosser.wsu.edu/frsttables.htm) and on the Michigan State University - Van Buren County Cooperative Extension Web site: (http://www.msue.msu.edu/vanburen/crittemp.htm). On both Web sites, photographs are provided to display the phenological stages for a variety of crops. Another review on spring frost injury and hardiness is presented in Rodrigo (2000).
Although less common than spring injury, winter frost injury typically affects deciduous fruit trees. In northern production areas, when winters are very severe, bark, woody tissue or buds can freeze. Bark injuries include:
crotch area injuries, which occur in trees with narrow crotch angles that harden late or sometimes incompletely;
sunscald injuries on sunny, cold winter days, when clouds then block the sun and cause a rapid cooling towards air temperature that may produce freezing;
bark splitting, which may occur under very cold conditions; and
trunk, collar and root injuries that occur when the soil protective effect is insufficient to avoid freezing of those plant parts (Myers, 1988).
Under extreme winter temperatures, or when trees fail to harden-off, woody tissues of branches are damaged (tip dieback) or trunks freeze (blackheart). In blackheart, xylem cells are killed, the wood oxidises, becomes dark and discoloured and the vessels fill with gummy occlusions. Blackheart usually doesn't kill the trees immediately, but opportunistic wood rotting organisms invade the injured trees and reduce productivity and longevity. Dormant winter buds often supercool to very low temperatures (e.g. -25 °C in peach winter buds and -41 °C for buds of azaleas). Winterkill of buds and bark tissues commonly occurs in plants that partially lose hardiness due to relatively warm periods. During spring-time, supercooling capacity gradually reduces as the buds expand and form flowers. Fully open flowers typically have critical temperatures between -1 °C and -3 °C (Burke et al., 1976).
Flowers are often damaged by spring frosts and the symptoms are darkened petals. Usually the flower style is more sensitive than the ovary to frost damage. After fertilization, the seeds are the most sensitive organs. A few days after a freeze event, the proportion of damaged flowers is obvious. When cut with a knife, healthy flowers have light green interior while damaged flowers are brownish (Figure 4.3A).
Seeds are essential for the normal development of most fruits, but some varieties of damaged apples and pears are able to sustain parthenocarpic development to yield misshapen fruits. Stone fruits are more susceptible to seed loss because they have only one or two seeds, while apples and pears are less susceptible, having more seeds.
When fruit experiences freeze injury, a coarse russet tissue grows and covers a portion or even the entire outside of the fruit. Although the damage can originate much earlier, russet rings show after full bloom (Figure 4.3B).
FIGURE 4.3
Frost damage to an apple flower (left); and
on small fruits (right)
[russet patches near the eyes and
rings].


Photos: A. Castro Ribeiro (ESAB, IPB, Portugal)
Table 4.6 lists critical temperatures for almond tree varieties, where some of the data came from field observations using temperatures from standard shelters and some came from excised-branch climate chamber studies. In the table, the full-bloom data for cv. Peerless are somewhat different for the field and chamber data, which illustrates the problem when comparing chamber studies with field observations. According to Hansen's full-bloom data, 25 percent damage would be expected at -2.2 °C, whereas only 1 percent damage was observed at -2.2 °C in the chamber studies. In general, for the same damage level, temperatures from chamber studies tend to be lower than from field studies. Therefore, the critical damage temperatures in the field are likely to be higher and damage could result when critical temperatures from chamber studies are used. If the bud, blossom or small-nut temperatures were measured directly in the field rather than using shelter temperatures, then the critical temperatures should be closer to those observed in the chamber studies. However, measuring bud, blossom or small-nut temperatures is not simple. The main point is that published critical temperatures should not be viewed as absolutely correct, but only as a guideline for making decisions about when to start and stop active protection methods.
TABLE 4.6
Damage expected (%) to some almond varieties
at various development stages after 30 minutes below the indicated
temperature
|
VARIETY |
|
STAGE |
TEMPERATURE |
|||||||
| |
|
|
-5.6 |
-5.0 |
-4.4 |
-3.9 |
-3.3 |
-2.8 |
-2.2 |
-1.7 |
|
Peerless |
[F] |
Full bloom |
|
|
|
100 |
75 |
45 |
25 |
|
| |
showing pink |
|
100 |
75 |
50 |
25 |
|
|
|
|
|
Peerless |
[C] |
full bloom |
|
|
|
79 |
50 |
14 |
1 |
|
| |
petal fall |
|
|
|
|
|
63 |
14 |
3 |
|
| |
nut stage |
|
|
|
|
|
46 |
45 |
9 |
|
|
NePlus Ultra |
[F] |
full bloom |
|
|
100 |
75 |
50 |
25 |
|
|
|
Mission |
[F] |
showing pink |
100 |
80 |
60 |
|
|
|
|
|
|
Drake |
[F] |
full bloom |
|
100 |
75 |
50 |
25 |
|
|
|
| |
showing pink |
75 |
50 |
25 |
|
|
|
|
|
|
|
Nonpareil |
[F] |
full bloom |
75 |
60 |
40 |
20 |
|
|
|
|
| |
showing pink |
20 |
10 |
|
|
|
|
|
|
|
|
Nonpareil |
[C] |
nut stage |
|
|
|
|
|
19 |
14 |
3 |
|
Butte |
[C] |
nut stage |
|
|
|
|
90 |
45 |
27 |
10 |
NOTES: [C] indicates tests with excised branches in a freezing chamber (Connell and Snyder, 1988).
[F] indicates results of several years of unpublished field observations by Harry Hansen (retired USA National
Weather Service) using a Stevenson screen and fruit frost shelter temperatures.
Table 4.7 contains a listing of the widely used deciduous tree crop critical temperatures corresponding to the main phenological stages (Proebsting and Mills, 1978). While these critical temperatures were developed in chamber studies, they do provide some guidance as to critical temperatures for use in the field. To account for the difference between field and chamber measured critical temperatures, the Tc values to use for management in the field should be slightly higher than those listed in the table.
TABLE 4.7
Critical temperature (Tc;
°C) values for several deciduous fruit tree crops
|
CROP |
STAGE |
10% KILL |
90% KILL |
|
Apples |
Silver tip |
-11.9 |
-17.6 |
|
Green tip |
-7.5 |
-15.7 |
|
|
1/2" green |
-5.6 |
-11.7 |
|
|
Tight cluster |
-3.9 |
-7.9 |
|
|
First pink |
-2.8 |
-5.9 |
|
|
Full pink |
-2.7 |
-4.6 |
|
|
First bloom |
-2.3 |
-3.9 |
|
|
Full bloom |
-2.9 |
-4.7 |
|
|
Post bloom |
-1.9 |
-3.0 |
|
|
Apricots |
Tip separates |
-4.3 |
-14.1 |
|
Red calyx |
-6.2 |
-13.8 |
|
|
First white |
-4.9 |
-10.3 |
|
|
First bloom |
-4.3 |
-10.1 |
|
|
Full bloom |
-2.9 |
-6.4 |
|
|
In shuck |
-2.6 |
-4.7 |
|
|
Green fruit |
-2.3 |
-3.3 |
|
|
Cherries (Bing) |
First swell |
-11.1 |
-17.2 |
|
Side green |
-5.8 |
-13.4 |
|
|
Green tip |
-3.7 |
-10.3 |
|
|
Tight cluster |
-3.1 |
-7.9 |
|
|
Open cluster |
-2.7 |
-6.2 |
|
|
First white |
-2.7 |
-4.9 |
|
|
First bloom |
-2.8 |
-4.1 |
|
|
Full bloom |
-2.4 |
-3.9 |
|
|
Post bloom |
-2.2 |
-3.6 |
|
|
Peaches (Elberta) |
First swell |
-7.4 |
-17.9 |
|
Caylx green |
-6.1 |
-15.7 |
|
|
Caylx red |
-4.8 |
-14.2 |
|
|
First pink |
-4.1 |
-9.2 |
|
|
First bloom |
-3.3 |
-5.9 |
|
|
Late bloom |
-2.7 |
-4.9 |
|
|
Post bloom |
-2.5 |
-3.9 |
|
|
Pears (Bartlett) |
Scales separate |
-8.6 |
-17.7 |
|
Blossom buds exposed |
-7.3 |
-15.4 |
|
|
Tight cluster |
-5.1 |
-12.6 |
|
|
First white |
-4.3 |
-9.4 |
|
|
Full white |
-3.1 |
-6.4 |
|
|
First bloom |
-3.2 |
-6.9 |
|
|
Full bloom |
-2.7 |
-4.9 |
|
|
Post bloom |
-2.7 |
-4.0 |
|
|
Prunes (Italian) |
First swell |
-11.1 |
-17.2 |
|
Side white |
-8.9 |
-16.9 |
|
|
Tip green |
-8.1 |
-14.8 |
|
|
Tight cluster |
-5.4 |
-11.7 |
|
|
First white |
-4.0 |
-7.9 |
|
|
First bloom |
-4.3 |
-8.2 |
|
|
Full bloom |
-3.1 |
-6.0 |
|
|
Post bloom |
-2.6 |
-4.3 |
The 10 percent kill and 90 percent kill imply that 30 minutes at the indicated temperature is expected to cause 10 percent and 90 percent kill of the plant part affected during the indicated phenological stage.
SOURCE: Proebsting and Mills, 1978
Grapes and wine grapes are often damaged by spring-time frosts. Since leaves form first, they are more prone to damage, but flowers and small berries are also sometimes damaged. Full recovery is common for leaf damage, but fruit damage can reduce production. The occurrence of early autumn frosts increases susceptibility to fungi attacks (e.g. botrytis). During winter, dormant buds are very rarely damaged, since they can resist temperatures below -10 °C, down to -20 or even -30 °C (Leddet and Dereuddre, 1989). Table 4.8 shows critical temperatures for grapes in relation to developmental stage.
TABLE 4.8
Critical temperature (Tc)
values (°C) for grapevines
|
Grape (1) |
New growth: |
|
-1.1 |
|
|
Woody vine: |
-20.6 |
- |
||
| |
French hybrids |
-22.2 |
-23.3 |
|
| |
American |
|
-27.8 |
|
| |
10% kill |
90% kill |
||
|
Grapes (cv. Concord) (2) |
First swell |
-10.6 |
-19.4 |
|
|
Late swell |
-6.1 |
-12.2 |
||
|
Bud burst |
-3.9 |
-8.9 |
||
|
First leaf |
-2.8 |
-6.1 |
||
|
Second leaf |
-2.2 |
-5.6 |
||
|
Third leaf |
-2.2 |
-3.3 |
||
|
Fourth leaf |
-2.2 |
-2.8 |
||
The 10 percent kill and 90 percent kill imply that 30 minutes at the indicated temperature is expected to cause 10 percent and 90 percent kill of the plant part affected during the indicated phenological stage
NOTES: (1) Krewer, 1988. The critical temperature was reported without giving the percentage kill. (2) www.msue.msu.edu/vanburen/crtmptxt.htm.
Blackberries and blueberries are hardy in winter, so frost damage occurs almost exclusively to the flowers and small fruits during spring-time. In contrast, if protective measures are not implemented, strawberries and kiwifruit are damaged in cold winters. First bloom is critically important for strawberry production, so frost damage during that phase is serious. When young, the cambium of young kiwifruit is often damaged by relatively high temperatures in autumn and spring, as well as by frost during cold winters. The first expanded leaves are tender and hence sensitive to damage. Critical temperatures of several small-fruit crops are shown in Table 4.9.
TABLE 4.9
Critical temperature (Tc)
values (°C) for several small fruits
|
CROP |
PHENOLOGICAL STAGE |
|
|
| |
|
? |
? |
|
Blackberry(1) |
Dormant flower buds |
|
-73.0 |
|
Open flower buds |
|
-2.2 |
|
| |
? |
? |
|
|
Blackberry(1) |
Dormant flower buds |
-27.2 |
-28.9 |
|
Open flower buds |
|
-2.2 |
|
| |
|
90% kill |
|
|
Blueberry(2) |
Swelled flower buds |
|
-6.1 |
|
Individual flowers distinguishable |
|
-3.9 |
|
|
Flowers distinctly separated, corollas expanded but closed |
|
-2.2 |
|
|
Fully opened flowers |
|
-0.6 |
|
| |
|
? |
|
|
Kiwifruit(3) |
Dormant flower buds |
|
-18.0 |
|
Green tip |
|
-3.0 |
|
|
Leaf veins visible |
|
-2.0 |
|
|
Expanded leaf |
|
-1.5 |
|
|
Individual flowers distinguishable |
|
-1.0 |
|
| |
|
90% kill |
|
|
Strawberry(2) |
Tight bud |
|
-5.6 |
|
Tight with white petals |
|
-2.2 |
|
|
Full bloom |
|
-0.6 |
|
|
Immature fruit |
|
-2.2 |
The 10 percent kill and 90 percent kill imply that 30 minutes at the indicated temperature is expected to cause 10 percent and 90 percent kill of the plant part affected during the indicated phenological stage.
SOURCES: (1) Krewer, 1988. The critical temperature was reported without giving the percentage kill. (2) Powel and Himelrick, 2000. (3) Vaysse and Jourdain, 1992.
Most citrus do not have a pronounced and stable dormancy. Growth is only reduced in winter and a spread of 1 to 2 °C in the freezing point of fruits is common between orchards and varieties, and even between trees. As the air temperature drops during the night, the fruit temperature typically lags behind and it is often a few degrees Celsius higher than the air temperature, especially during the evening. The bigger the fruit, the greater the lag between fruit and air temperature. Supercooling plays also a role in the freezing temperature and explains the importance of freezing nuclei concentration and white frost or dew formation on the fruit surface. Also, it is known that the peel has a lower freezing temperature than the flesh inside. Therefore, frost damage can occur inside the fruit without any obvious damage on the outside. Despite all these confounding factors, some critical fruit temperatures for the major citrus crops are presented in Table 4.10.
TABLE 4.10
Critical fruit temperatures
(Tc) when citrus fruits, buds or blossoms begin to
freeze
|
CITRUS SPECIES |
CRITICAL TEMPERATURE (°C) |
|
Green oranges |
-1.9 to -1.4 |
|
Half ripe oranges, grapefruit and mandarins |
-2.2 to -1.7 |
|
Ripe oranges, grapefruit and mandarins |
-2.8 to -2.2 |
|
Button lemons |
-1.4 to -0.8 |
|
Tree ripe lemons |
-1.4 to -0.8 |
|
Green lemons (diameter >12 mm) |
-1.9 to -1.4 |
|
Lemon buds and blossoms |
-2.8 |
SOURCE: After Puffer and Turrell, 1967.
When air temperature (Ta) drops rapidly following a warm day, citrus fruit temperatures (Tcf) lag behind the air temperature drop and the temperature difference (Tcf - Ta) is bigger for larger fruit. When protecting small fruit and there is a rapid air temperature drop after sunset, wind machines and heaters should be working when Ta reaches Tc (Table 4.10). For larger fruit, on nights with rapid air temperature drop during the evening, start the wind machines or heaters when Ta is slightly lower than Tc (e.g. when Ta = Tc - 0.5 °C). During mild advection frosts or on nights with higher humidity and slower temperature drop, Tcf is closer to Ta, so the wind machines or heaters should be working when the Ta » Tc (Table 4.10). If the fruit and leaves are wet from rainfall, fog or dew and the wet-bulb temperature (Tw) is expected to fall below Tc during the night, wind machines and heaters should be started as soon as possible in the evening to dry the fruit surfaces before the wet fruit temperature falls below Tc. Otherwise, damage to the peel is likely.
During weather conditions when the air temperature is expected to reach the dew-point (Td) temperature, which is higher than Tc and the predicted minimum temperature is below Tc, then it is wise to start the wind machines or heaters before Ta falls to Td and dew or frost begins to condense on the fruit.
On nights following light rainfall or snow or when dew or frost forms on the fruit, damage can occur to the fruit rind even when the shelter temperature is above the critical damage fruit temperature. This occurs because the temperature of the wet part of the fruit can fall to the wet-bulb temperature, due to the removal of sensible heat as the water evaporates. This is the cause of rind spot damage that occurs in some years. This is also true for spot damage on deciduous fruit damage during autumn freezes. The wet-bulb temperature is always between the air and dew-point temperatures and the wet-bulb temperature is lower when the dew-point is low. If the fruit is wet going into a freeze night, protection should be started as early as possible. In these conditions, the objective is to evaporate the water off the fruit before the wet-bulb temperature reaches 0 °C. Using heaters or wind machines before nightfall will help to evaporate water off the plants. However, using wind machines when the fruit is wet after the wet-bulb temperature falls below the critical fruit damage could cause rind injury or worse. Table 4.11 gives the air temperature corresponding to a wet-bulb temperature Tw = 0 °C for a range of dew-point temperatures and elevations.
TABLE 4.11
Air temperatures (°C) corresponding to a
wet-bulb temperature Tw = 0 °C for a range of dew-point
temperatures and elevations.
|
DEW-POINT TEMPERATURE |
ELEVATION (METRES ABOVE MEAN SEA LEVEL) |
|||
|
(°C) |
0 m |
500 m |
1000 m |
1500 m |
|
0 |
0.0 |
0.0 |
0.0 |
0.0 |
|
-2 |
1.2 |
1.3 |
1.4 |
1.5 |
|
-4 |
2.3 |
2.5 |
2.6 |
2.8 |
|
-6 |
3.3 |
3.5 |
3.7 |
3.9 |
|
-8 |
4.1 |
4.4 |
4.6 |
4.9 |
|
-10 |
4.8 |
5.1 |
5.4 |
5.8 |
|
-12 |
5.4 |
5.8 |
6.1 |
6.5 |
|
-14 |
6.0 |
6.3 |
6.7 |
7.1 |
|
-16 |
6.4 |
6.8 |
7.2 |
7.7 |
|
-18 |
6.8 |
7.2 |
7.7 |
8.1 |
![]()
![]()
![]()