![]()
![]()
![]()
Rex D. Pieper
SUMMARY
At the time of colonial settlement there was extensive grassland from the prairies of Canada to the Gulf of Mexico on mainly level topography. Great Plains grassland is in three strips running north-south: tall-grass, mixed -grass and short-grass, with the tall-grass in the better watered west. Precipitation increase from west to east (320 to 900 mm) is the main factor governing primary productivity; periodic droughts occur. Bison, the dominant large herbivore until the mid-nineteenth century, have largely been replaced by cattle. About half of the beef cattle in the United States of America are in the Great Plains. Woody vegetation types are embedded, with the trees varying according to latitude. C4 species comprise more than 80 percent from 30° to 42°N, while C3 species increase dramatically north of 42°N. Only 1 percent of the tall-grass remains; half of the short-grass is uncultivated; unproductive cropland is being put back to grass. Cattle predominate; sheep are far fewer and declining. Most land is privately owned, much in small farms. There is extensive ranching in dry areas. Grazing is seasonal, especially in the north with supplemental feed in winter. In favourable areas, sown pasture are used sometimes alongside range grazing. Rotational grazing is common, although research results on its advantage are mixed. Fire is used to suppress undesirable plants and increase fodder production. Grassland monitoring includes long-term ecological research sites. Introduced plants can cause problems: Euphorbia esula is an aggressive weed and Bromus japonicus often competes with native grasses. Many small operations are no longer economically viable, so many are being abandoned. Livestock enterprises should remain viable, although they have to compete with systems based on forage grown under irrigation.
When European settlers first moved into the central portion of what is now the United States of America (USA), they encountered an extensive, unbroken grassland extending from the prairies of Canada to the Gulf of Mexico and Mexico. While this grassland was generally free from woody plants, apparently there was a dynamic ecotone between the mountains and deserts to the west and the eastern deciduous forests in central and northern portions of the grassland (Bazzaz and Parrish, 1982; Gleason, 1913; Transeau, 1935). The general impression held by many observers was that conditions in this vast grassland prior to European settlement were pristine and little modified by man (Deneven, 1996; Leopold et al., 1963). However, Flores (1999) presents persuasive arguments that native American populations were relatively high in the Great Plains and probably exerted considerable influence on other components of these grassland ecosystems.
Figure 6.1 - The extent of grasslands in North America.
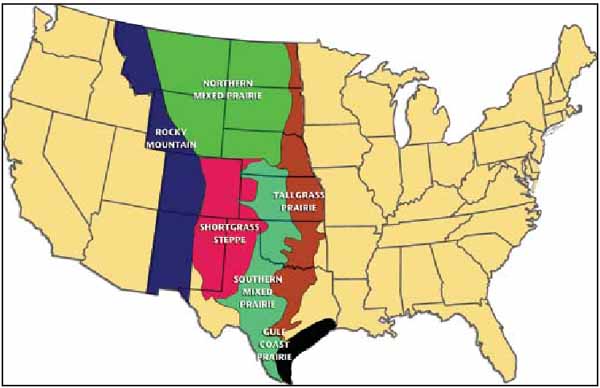
Figure 6.2
Map showing grassland types in central
USA.
SOURCE: Lauenroth et al.,1994.
The extent of grasslands in North America is shown in general terms in Figure 6.1. At coarse scales the Great Plains grassland is commonly divided into tall-grass (true prairie), mixed - (or mid-)grass, and short-grass (Bazzaz and Parrish, 1982; Gleason and Cronquist, 1964; Lauenroth, et al., 1994; Laycock, 1979; Sieg, Flather and McCanny, 1999; Sims, 1988) (Figure 6.2). Plates 6.1a-c show examples of the three major grassland types from North Dakota to New Mexico. The map in Lauenroth et al. (1994) (Figure 6.2) shows that the tall-grass prairie formed a narrow band from Canada south to the Gulf Coast Prairie in Texas. The diagram in Barbour, Burk and Pitts (1987) shows a transect (Figure 6.3) from the Pacific Ocean to the Atlantic at latitude 37°N (the northern boundary of Arizona and New Mexico). This diagram shows short-grass vegetation occupying the area between 105° and 101°W, the mixed prairie to 98°W and the tall-grass prairie to 93°W. The map in Lauenroth et al. (1994) shows that the northern mixed prairie forms a broad band in Canada, eastern Montana, the Dakotas and eastern Wyoming (Figure 6.2). The southern mixed-grass prairie is constricted between the tall-grass prairie to the east and the short-grass prairie to the west, and found in southern Nebraska, central Kansas and Oklahoma, and Texas (Lauenroth et al., 1994; Lauenroth, Burke and Gutmann, 1999).

Plate 6.1a
Examples of the three major grassland
types, from North Dakota in the north to New Mexico in the south - (a)
Short-grass prairie (central New Mexico).
REX PIEPER

Plate 6.1b
Examples of the three major grassland
types, from North Dakota in the north to New Mexico in the south - (b) Northern
mixed -grass prairie (central North Dakota).
REX PIEPER

Plate 6.1c
Examples of the three major grassland
types, from North Dakota in the north to New Mexico in the south - (c)
Tall-grass prairie (eastern Kansas).
REX PIEPER
Figure 6.3 - Transect showing topographical changes across central USA.
SOURCE: Barbour, Burk and Pitts, 1987.
Figure 6.4 shows the precipitation gradient east of the Rocky Mountains in the central part of the Great Plains resulting from the rain-shadow effect of the Rocky Mountains. Annual precipitation increases from about 320 mm at Greeley, Colorado, to nearly 900 mm at Kansas City, Missouri. Seasonal precipitation patterns also vary from south to north in the central plains (Trewartha, 1961). In southern portions of the short-grass prairie, summer maxima are the rule, while further east there is one peak in the late spring -early summer and another in late summer-early autumn (type 3b in Trewartha, 1961). In the northern Great Plains, spring peaks are common (type 3c in Trewartha, 1961) while in the upper Mississippi Valley-Great Lakes region, summer and autumn peaks occur (Trewartha, 1961).
The major temperature gradient is from warm temperatures in southern grassland to cooler temperatures in the north (Figure 6.5). The gradient is steeper for January temperatures than for July temperatures. Colder winter temperatures in the north have many implications for both plants and animals. However, snow cover in the north may ameliorate extremely cold air temperatures at the soil surface. Winter temperatures in southern locations allow cool-season plant growth almost any time that there is adequate soil moisture (Smeins, 1994; Holechek, Pieper and Herbel, 2001).
Before European settlement, the three grassland types in the central portion of the continent were fairly comparable in area: short-grass was 615 000 km2, mixed prairie was 565 000 km2 and tall-grass prairie was 570 000 km2 (Van Dyne and Dyer, 1973). Today, the tall-grass prairie is much constricted because of conversion to intensive agriculture; originally, it extended eastward into southern Minnesota, most of Iowa, northern Missouri and northern Illinois and western Indiana (Lauenroth, Burke and Gutmann, 1999). Currently, the tall-grass prairie occurs mostly as isolated tracts, such as the Osage Hills in Oklahoma and the Flint Hills in Kansas.
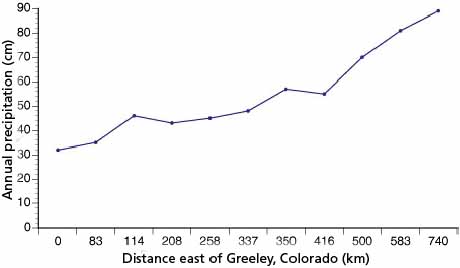
Figure 6.4
Mean annual precipitation gradient for
various sites on the Great Plains east of the Rocky Mountains.
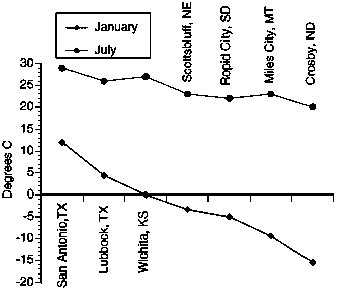
Figure 6.5
Average January and July temperatures
for various sites on a south-north transect in the Great Plains.
Topographically, the Great Plains are relatively level, but minor topographic variations are important in influencing plant species distributions and other inclusions within the grassland. Often, poorly drained sites in depressions, either with or without standing water, offer habitats for plants and animals not found in adjacent grassland.
Grassland soils have developed from a variety of parent materials: limestone, sandstone, shale, metamorphic and igneous outwash, and loess (Buol, Hole and McCracken, 1980; Dodd and Lauenroth, 1979; Miller and Donohue, 1990; Sims, Singh and Lauenroth, 1978). The major soils are mollisols, deep soils with dark A horizons and high (>50 percent) base saturation (especially calcium) (Miller and Donohue, 1990). Surprisingly, the A horizon has a clay content nearly equal to that of the B horizon (Baxter and Hole, 1967). Several soil organisms including a common prairie ant (Formica cinerea) are apparently involved in translocation of clays from the B horizon to the A (Buol, Hole and McCracken, 1980). Buol, Hole and McCracken (1980) describe the soil forming process for mollisols as “melanization”. This process consists of five specific processes (Hole and Nielsen, 1968):
1. Growth of plant roots into the soil profile.
2. Partial decay of organic material in the soil.
3. Mixing of the soil by soil micro-organisms.
4. Eluviation and illuviation of organic colloids and some inorganic colloids.
5. Formation of resistant “ligno-protein” residues producing the dark colour in the soil.
Soils developed in semi -arid portions of the grassland, classified as aridisols formed by calcification, often develop calcium carbonate (caliche) layers at varying depths below the surface (Breymeyer and Van Dyne, 1979). Other soils found in the grassland include alfisols, found extensively in Kansas, Oklahoma and Texas (Sims, 1988).
At the time of early exploration of the American grassland, bison (Bison bison) were the dominant large herbivore in the Great Plains (Plate 6.2), although pronghorn antelope (Plate 6.3) were also abundant (Shaw, 1996; Yoakum, O'Gara and Howard, 1996). Seton (1927), Garretson (1938) and Danz (1997) estimated that as many as 40-60 million head of bison were present in the North American grassland before settlement. Numbers of pronghorn were probably comparable to those for bison (Nelson, 1925; Yoakum, O'Gara and Howard, 1996).
Grasslands also provided habitat for a wide variety of small mammals, including prairie dogs (Cynomys spp.), jackrabbits (Lepus spp.), ground squirrels (Spermophilus spp.), gophers (Geomys spp. and Thomomys spp.) and voles (Microtus spp.). Originally, several species of prairie dogs occupied over 800 000 ha of grasslands in central USA (Kreitzer and Cully, 2001; Summer and Linder, 1978), but by the early 1990s their distribution had been reduced by 98 percent (Vanderhoff, Robel and Kemp, 1994).
A wide number of invertebrates such as grasshoppers, beetles, ants, sap feeders and members of other orders are important components of grassland ecosystems (Blocker, 1970; McDaniel, 1971; Risser et al., 1981). Some, such as grasshoppers, have been studied because of their economic importance (Hewitt, 1977; Hewitt and Onsager, 1983) while others, such as nematodes, have only recently been properly assessed as to abundance and importance (Freckman, Duncan and Larson, 1979; Smolik, 1974).

Plate 6.2
Remnant herd of bison (Bison
bison).
S.G. REYNOLDS

Plate 6.3
Pronghorn antelope on mixed grass
prairie - North Dakota.
DUANE McCARTNEY
Central plains grasslands also support diverse populations of birds (Bolen and Crawford, 1996; Guthery, 1996; Knopf, 1996; Wiens, 1973). However, within the grassland geographical region, habitats other than grasslands have higher numbers of avian species. Only 11 percent of the bird species within the grassland geographic area were actually inhabitants of grassland per se: 51 percent were associated with woodland and forest habitats and 22 percent with wetland habitats (Bolen and Crawford, 1996). Nevertheless, birds are abundant in grasslands. Glover (1969) listed over 150 species found on the Central Plains Experimental Range in short-grass habitat in north-central Colorado. These included both primary consumers and secondary consumers. Common grassland birds include western meadowlark (Sturnella neglecta), grasshopper sparrow (Ammodramus savannarum), horned lark (Eremophila aplestris) and chestnut-collared longspur (Calcarius ornatus) (Wiens, 1973, 1974).
Cattle (Plates 6.4 and 6.5) have largely replaced bison as the dominant large herbivore on the Great Plains. The importance of Great Plains cattle to the beef industry in the USA is reflected in data quoted by Holechek, Pieper and Herbel (2001) indicating that 50 percent of all USA beef cattle are found in the northern and southern Great Plains. Although Lauenroth et al. (1994) concluded that vegetation changes resulting from the shift from bison to cattle have been minimal, there are differences in grazing patterns and behaviour: bison select a diet higher in grasses than cattle; bison select a diet higher in digestibility than cattle; bison spend less time grazing than cattle; and cattle are restricted in grazing by fences while bison were free to move over the landscape (Danz, 1997; Donohue, 1999; Stueter and Hidinger, 1999; Pieper, 1994).

Plate 6.4
Herding cattle in a mixed -grass zone,
Saskatchewan, Canada.
MAE ELSINGER

Plate 6.5
Cattle grazing on Deseret Ranch near
Salt Lake City, Utah, USA.
DUANE McCARTNEY
While the Central Plains grassland of North America is often divided into three subdivisions, as noted earlier, the patterns are more complex at finer scales. For example, Sieg, Flather and McCanny (1999) show 24 major vegetation types extending from Canada to the coastal prairies in Texas. Similar rangeland also occurs in northern Mexico. Woody vegetation types embedded in the grassland includes aspen parkland in the north, mesquite -acacia savannah, shinnery oak savannah, cross timbers, mesquite-buffalo grass, juniper-oak savannah, oak-hickory forest and oak-hickory-pine forest in the south (Sieg, Flather and McCanny, 1999). These woodland types have been described in more detail by Dahl (1994), McClendon (1994), Pettit (1994), Engle (1994) and Smeins (1994). Several species of juniper have expanded into grasslands in recent times. In Oklahoma some estimates of expansion of eastern red cedar (Juniperus virginiana) are as high as 113 000 ha annually (Engle, 2000; Gehring and Bragg, 1992). In Texas, ash (Juniperus ashei) and redberry (Juniperus pinchoti) junipers have also expanded into grasslands (Smeins, 2000). Reduced fire intensity and frequency is considered one of the primary factors in this expansion (McPherson, 1997).
Major grass dominants in the tall-grass prairie (Plate 6.6) are big bluestem (Andropogon gerardii) little bluestem (Schizachyrium scoparium), Indian grass (Sorghastrum nutans) and switchgrass (Panicum virgatum). In the mixed -grass prairie (see Plates 6.7, 6.8 and 6.9), needle-and-thread grass (Stipa comata) and western wheatgrass (Pascopyrum smithii) are common grasses, but many other species are abundant on specific sites. Sims (1988) states that the vegetational diversity of the mixed prairie is highest of all grassland types in the USA (not unusual for vegetation often considered an ecotone). Two major grass dominants of short-grass (Plate 6.10) vegetation are blue grama (Bouteloua gracilis) (Plate 6.11) and buffalo grass (Buchloe dactyloides). Many forb species are also common in the grassland. Thus, the term “grassland” may be somewhat misleading if it implies only grasses are present in high abundance. Typical grassland scenes near Mandam in North Dakota and near Salt Lake City, Utah, are shown in Plates 6.12 and 6.13, respectively.

Plate 6.6
Tall-grass prairie - Sheyenne National
Grassland area, North Dakota, USA.
JEFF PRINTZ, USDA-NRCS

Plate 6.7
Mixed-grass prairie - west side of the
Missouri river, just south of Mandan, North Dakota.
JEFF PRINTZ, USDA-NRCS

Plate 6.8
Mixed-grass prairie - north-central
North Dakota, USA.
JEFF PRINTZ, USDA-NRCS

Plate 6.9
Mixed-grass prairie on the Monet
Prairie Farm Rehabilitation Community Pasture, Saskatchewan,
Canada.
MAE ELSINGER

Plate 6.10
Short-grass prairie - from a badlands
area.
JEFF PRINTZ, USDA-NRCS
Within the central North America n grassland, there are also minor variations in species composition related to micro-relief patterns. Ayyad and Dix (1964) reported that three species (Festuca scabrella, Carex obtusa ta and Galium boreale) were most abundant on moist and cool north-facing slopes, while other species (Phlox hoodii, Carex filifolia, Stipa comata, Artemisia frigida) reached highest abundance on relatively warm and dry south-facing slopes. Species occupying intermediate habitats were Koeleria cristata, Carex eleocharis, Stipa spartea and Agropyron dasystachum. Redmann (1975) and Sims (1988) reported on micro-topographical and soils variations in mixed prairie vegetation in North Dakota. Agropyron smithii, Carex pennsylvanica and Stipa comata were dominant species on rolling upland with fine-textures soils, while sites at lower elevations with medium-textured soils supported stands of Sporobolus heterolepis. In short-grass vegetation in New Mexico, blue grama was present in all topographic positions, while Lycurus phleoides, Aristida wrightii, Stipa neomexicana and Bouteloua curtipendula occurred on upper slopes. Sporobolus cryptandrus and Muhlenbergia torrey i along with blue grama were dominant on lower slopes, with blue grama and buffalo grass, Muhlenbergia repens and Hilaria jamesii in moister depressions (Beavis et al., 1981).

Plate 6.11
Bouteloua gracilis - Blue
grama.
S.G. REYNOLDS

Plate 6.12
Mixed grass prairie near Mandan, North
Dakota.
DUANE McCARTNEY

Plate 6.13
Grassland with sagebrush near Salt
Lake City, Utah.
DUANE McCARTNEY
There is also a north-south gradient in the relative proportion of C3 and 4 species (Sims, Singh and Lauenroth, 1978). C4 species comprise more than 80 percent of the flora from 30° to 42°N, while C3 species increase dramatically north of 42°N (Sims, 1988).
Several environmental variables act to control primary production in the Central Plains grasslands in North America. Precipitation, as it is translated into soil water content through infiltration, is often considered the main control for primary production (Laurenroth, 1979; Sims, Singh and Lauenroth, 1978; Sims and Singh, 1978a, b). Several studies have provided regression analyses showing the relationship between precipitation and above ground net primary productivity (ANPP). Lauenroth (1979) showed a linear relationship between ANPP and mean annual precipitation for 52 grassland sites around the world, with r2 of 0.51 under mean annual precipitation ranging from about 100 mm to about 1500 mm. Later, Lauenroth, Burke and Gutmann (1999) presented similar analyses for a much larger data set from central USA grasslands for normal rainfall, favourable and unfavourable rainfall patterns. The r2 values were 0.56 for normal years, 0.66 for favourable years and 0.43 for unfavourable years (favourable years represent the wettest 10 percent of the years; unfavourable are the driest 10 percent of the years; and normal the middle 80 percent (Soil Conservation Service, 1973)). Of course, the ANPP-precipitation relationship is not linear over the complete range of precipitation values, and often annual precipitation is a relatively poor predictor of ANPP or end-of-season standing crop (Pieper, 1988). For example, Smoliak (1956) found that May-June precipitation was highly related to end-of-season standing crop in northern Great Plains short-grass prairie (r2= 0.86), while Hart and Samuel (1985) found a high correlation between spring -summer precipitation and herbage yield in short-grass vegetation in eastern Wyoming (r2 = 0.95). Since primary production is so closely related to precipitation, the general pattern of primary production (Plate 6.14) follows those gradients of precipitation, increasing from west to east. Lauenroth (1979) reported that general average annual production was about 200 g/m2 for short-grass, 300 g/m2 for mixed -grass prairie and 500 g/ m2 for USA International Biological Programme locations. These general averages mask the tremendous variation across these grasslands. For example, Risser et al. (1981) reported that above ground peak live standing crop for 23 tall-grass locations varied from 180 g/m2 at Junction, Kansas, to nearly 600 g/m2 in Oklahoma County, Oklahoma.

Plate 6.14
Estimating annual biomass productivity
on the Monet Prairie Farm Rehabilitation Area, Saskatchewan,
Canada.
MAE ELSINGER
Considerable portions of the central grasslands of the USA have been converted to intensive agriculture (Gunderson, 1981). Thomas, Herbel and Miller (1990) estimated that only about 1 percent of the original tall-grass prairie is still in native vegetation, while Lauenroth, Burke and Gutmann (1999) estimated that approximately 50 percent of the short-grass prairie is still uncultivated.
Wheat is the major crop grown on the western edge of the Great Plains, although larger percentages of the land in wheat occurred in central and eastern Kansas and Oklahoma and northeastern North Dakota (Lauenroth, Burke and Gutmann, 1999). The pattern of acreage of wheat grown in the Great Plains was one of breaking native sod during times of plentiful precipitation and high wheat prices, and abandonment of these lands during drought and periods of low wheat prices (Holechek, Pieper and Herbel, 2001; Sims, 1988; Stoddart, Smith and Box, 1975). The dust bowl of the 1930s occurred in southeastern Colorado, southeastern Kansas and the Texas and Oklahoma panhandles, largely on land unsuited for cultivation without irrigation (Costello, 1944; Holechek, Pieper and Herbel, 2001; Jordan, 1995; Sims, 1988). Considerable effort has been expended in developing seeding techniques to “recover” these abandoned fields (Bement et al., 1965).
In 1985, the Food Security Act provided the opportunity for land owners to retire cropland, and provided cost-share funding to establish grass cover, wildlife habitat or trees (Joyce, 1989). Under the Conservation Reserve Programme (CRP) of this USA Act, many land owners converted cropland to grassland (Mitchell, 2000). The CRP is a voluntary cropland retirement programme under which the Federal Government pays an annual rental fee and a cost share for conversion from cropland to a permanent cover of grass, wildlife or trees. The basic goals for creation of the CPR were to: (1) take highly erosive cropland out of production and to establish a permanent perennial vegetation cover; (2) to decrease farm commodity surpluses; (3) to generate stable incomes for participants; and (4) to enhance natural resource values, including soil, water, air quality and wildlife (Goetz, 1989; Heimlich and Kula, 1989). The map presented by Mitchell (2000) shows that CRP lands are concentrated in the plains states, with high densities in the northern Great Plains (Montana and North Dakota), the corn belt (southern Iowa and northern Missouri) and the southern Great Plains (eastern Colorado, western Kansas and the panhandles of Oklahoma and Texas). Originally, the CRP programme was designed for a 10-15-year period, but in separate Acts in 1990 and 1996, the CRP was extended and broadened (Mitchell, 2000).
Most land in the Great Plains is under private ownership (Holechek, Pieper and Herbel, 2001). For example, Neubauer (1963) reported that there was nearly 34 million hectares of private range and pasture land in the Dakotas, Nebraska and Kansas, but only 1 million hectares of state and Indian land and 1.4 million hectares of federal land. There are National Forest lands in the Dakotas, Nebraska, Arkansas and Missouri, but most of these are forest or woodlands (Mitchell, 2000). Some Forest Service Land is in National Grasslands, but these are relatively small compared with non-federal land. Licht (1997) lists 15 separate National Grasslands in the Plains states. These vary in size from less than 600 ha for McClelland Creek National Grassland, to over 400 000 ha in the Little Missouri National Grassland in North Dakota. The Bureau of Land Management (BLM) has mineral rights to considerable areas outside the West, but little surface rights in the Great Plains (Holechek, Pieper and Herbel, 2001). Most of the BLM holdings in the Plains grassland is in Montana and Wyoming, but there are also a few allotments in South Dakota (Licht, 1997; Wester and Bakken, 1992).
Major crops in tall-grass regions are maize and soybeans (Lauenroth, Burke and Gutmann, 1999), with maize more important in the north and soybean in the central and southern portions of the grasslands. Wheat is the other major cereal crop, with higher yields in eastern portions, but greater areas in the west and north in locations with less than 500 mm/yr precipitation (Lauenroth, Burke and Gutmann, 1999). Cotton is important in eastern New Mexico, western Oklahoma and northern Texas.
Livestock production
Several types of enterprises constitute the livestock production systems of Central North America. In the drier portions of the region (short-grass and mixed -grass), extensive range grazing operations are the norm. These operations are typically cow-calf operations with the young animal sold for finishing in feedlots (Neumann and Lusby, 1986). In other cases, stocker programmes - whereby weaned animals are retained and maintained by feeding roughages to ensure growth but not improvement in condition - are practised (Neumann and Lusby, 1986; Wagnon, Albaugh and Hart, 1960). Stocking rates on native short-grass prairie vary considerably depending on precipitation, range condition and other environmental factors. Klipple and Costello (1960) reported that moderate stocking in eastern Colorado was about 21 ha per animal unit year (AUY), while Bement (1969) recommended 19.4 ha to support 1 AUY. In the panhandle of Nebraska, Burzlaff and Harris (1969) reported that moderate stocking was 14 ha per AUY. These stocking rates were based on summer grazing from May through October. In mixed-grass regions, stocking rates are normally higher, e.g. 9.3 ha AUY in North Dakota (Rogler, 1951). In tall-grass prairie, stocking rates are much higher: 4 ha AUY in Oklahoma (Harlan, 1960). In the Prairie Provinces of Canada (see Plates 6.15, 6.16 and 6.17), Smoliak et al. (1976) reported that carrying capacities ranged from 21.6 to 10.9 ha AUY depending on range type and condition. Holechek, Pieper and Herbel (2001) reviewed several grazing studies in the Great Plains, and recommended 35-40 percent removal (utilization) of current annual production to maintain vigorous plants and grassland ranges in a highly productive condition. Bement (1969) recommended leaving 300-350 kg/ha at the end of the summer grazing period to support both livestock performance and desirable vegetation condition for short-grass rangeland in eastern Colorado.

Plate 6.15
Mixed-grass zone: roped steer on the
Tecumseh Prairie Farm Rehabilitation Area, Saskatchewan, Canada.
MAE ELSINGER

Plate 6.16
Mixed-grass zone: cattle on the
Caledonia Prairie Farm Rehabilitation Area, Saskatchewan, Canada.
MAE ELSINGER

Plate 6.17
Winter grazing on the Canadian
prairie.
MAE ELSINGER
Most livestock operations in the Great Plains are relatively small. Over 85 percent of the farms and ranches in the Great Plains (including North and South Dakota, Nebraska, Montana, Wyoming, Kansas and Oklahoma) had less than 100 head of cattle, and only 5 percent had more than 500 head (Mitchell, 2000). Over 46 percent had less than 50 head. These were probably small operations where cattle were produced in conjunction with cropping operations. There were over 180 000 individual units in these seven states in 1993. Numerically, cattle are much more important than sheep in the Central Plains grasslands (Mitchell, 2000). Total cattle numbers for most of the states making up the Great Plains was about 25 million head, compared with about 5 million sheep (Ensminger and Parker, 1986; Mitchell, 2000). Sheep numbers have declined during the last 50 years because of predator problems, economic conditions, lack of herders in some western states, lack of demand for mutton and lamb, and other factors. These data on livestock numbers do not distinguish among different types of operations.
The northern and southern Great Plains (including Texas) support about half of the total beef cattle in the USA (Holechek, Pieper and Herbel, 2001), while the rest of the West supports less that 10 percent of the total. There was no distinction between those in feedlots and those on farms and ranches, but these numbers illustrate the importance of the Great Plains as a livestock-pro-ducing area.
In more mesic situations, mixed operations are common, whereby animals are raised under pasture or confined conditions in combination with cultivated agriculture practices (Neumann and Lusby, 1986). In many cases, the number of animals in these situations is relatively small.
Although grasslands have the potential to be grazed year long, they are grazed mostly seasonally by livestock. In northern areas, inclement weather largely precludes grazing during the winter (Holechek, Pieper and Herbel, 2001; Neumann and Lusby, 1986; Stoddart and Smith, 1955). Native grass hay and alfalfa have been used extensively as winter feed in northern areas and as supplemental feed in southern areas (Neumann and Lusby, 1986; Newell, 1948; Rogler and Hurtt, 1948). Keller (1960) showed that wild hay (native grasses) occupied over 3.6 million hectares in the six plains states, followed by alfalfa on about 2.8 million hectares and cereal hay on about 0.4 million hectares.
Another factor involved in decisions for seasonal grazing is that of declining nutritive quality of forage as the growing season progresses (Adams et al., 1996; Rao, Harbers and Smith, 1973; Scales, Streeter and Denham, 1971). Protein content of Great Plains forage generally reaches a peak during the early summer period and declines sharply as the forage matures into winter (Adams et al., 1996). During the late summer, forage quality was rated as moderate, while in the winter it was rated as low quality. Other nutritional variables often change in a similar pattern, such as phosphorus content, digestibility and intake. Changes in nutritive quality of forage depend to some degree on species composition: cool-season grasses (C3) have higher nutritive quality early in the season than warm-season species (C4) that grow later, during the heat of the summer. Presence of forbs may also increase mineral content of herbivore diets (Pieper and Beck, 1980; Holechek, 1984). In New Mexico, protein and phosphorus content of side-oats grama (Bouteloua curtipendula) was significantly lower than that of five other grass species (Pieper et al., 1978). Tall-grass prairie plants often become coarse and relatively unpalatable late in the growing season.
One approach to mitigation of the problems of low nutritive quality and palatability of forage late in the growing season is intensive early stocking (Bernardo and McCollum, 1987; Lacey, Studiner and Hacker, 1994; Smith and Owensby, 1978; McCollum et al., 1990; Olson, Brethour and Launchbaugh, 1993). In Montana, early spring grazing was beneficial for most vegetational characteristics compared with summer grazing, but livestock performance was not reported (Lacey, Studiner and Hacker, 1994). In tall-grass prairie in Kansas, intensive early grazing improved steer gains per unit area compared with season-long grazing, resulted in more even utilization of the pastures and increased desirable perennial grass production, but reduced gain per steer (Smith and Owensby, 1978). Other studies in Kansas indicated that stocking density could be increased 2 to 3 times by early-season grazing compared with summer-season grazing (Launchbaugh and Owensby, 1978). However, Olson, Brethour and Launchbaugh (1993) cautioned against using early-season grazing at high stocking rates when vigour of cool-season plant species is a concern. McCollum et al. (1990) reported that total beef production was increased 19 percent under increased early-season stocking compared with traditional season-long stocking.
Other stocking plans for the Great Plains include several types of rotational grazing systems (Holechek, Pieper and Herbel, 2001). The objective of many of these plans is to increase individual plant vigour and overall plant productivity. The basic design for rotation systems is to defer grazing on different pastures or portions of the entire area at different seasons in different years (Vallentine, 1990). This objective is met by adjusting the number of pastures and herds to ensure that the same area is not grazed at the same period each year. Research results comparing specialized grazing systems to continuous grazing have been mixed (Herbel, 1974; Herbel and Pieper, 1991; Hickey, 1969; Van Poolen and Lacey, 1979). In some cases there has been little difference in either cattle production or vegetation status between continuous grazing and some form of rotation grazing in the central Great Plains (Hart et al., 1988; Lodge, 1970; Rogler, 1951; McIlvain and Shoop, 1969; McIlvain and Savage, 1951; McIlvain et al., 1955). Generally, rotational grazing systems, whereby livestock are concentrated in one pasture for short periods, decreased livestock performance (gain per head), presumably because of lower selection, lower nutritive quality of forage selected and lower digestibility (Malechek, 1984; Pieper, 1980). Field studies in some cases confirmed this (Fisher and Marion, 1951; Heitschmidt, Kothmann and Rawlins, 1982; Pieper et al., 1991; Smith et al., 1967). Other studies indicated that there was some vegetation improvement, including higher production, increases in abundance of plant species desirable for grazing, or increases in plant cover (Herbel and Anderson, 1959; Smith and Owensby, 1978; Pieper et al., 1991) under specialized grazing systems. Van Poollen and Lacey (1979) reported that six studies in the northern Great Pains showed virtually no difference in herbage yields under continuous grazing and rotation systems, while in tall-grass prairie (Flint Hills in Kansas) herbage yields were 17 percent higher under specialized grazing systems than under continuous grazing. In some cases, improved management and more uniform grazing distribution under specialized grazing systems are confounded with the grazing system.
During the 1970s and 1980s, interest in short-duration or time-controlled grazing, as advocated by Savory (1999), peaked. This grazing approach is based on having a large number of paddocks and moving livestock rapidly through the paddocks, especially during periods of rapid plant growth (Savory, 1983, 1999; Savory and Parsons, 1980). The grazing period is often only a matter of a few days or even, in extreme cases, hours, since all the livestock normally allocated to the entire area are concentrated into one paddock at a time. Savory has stated that stocking could be doubled over that recommended by standard Soil Conservation Service procedures (Bryant et al., 1989). Results of experiments involving short-duration grazing in the Great Plains have been mixed. Holechek, Pieper and Herbel (2001) evaluated nine studies conducted on USA and Canadian grasslands. Many of these studies showed little difference in herbage yield between short-duration grazing and continuous grazing (Manley et al., 1997; Pitts and Bryant, 1987; Thurow, Blackburn and Taylor, 1988; White et al., 1991). In some cases there was some advantage for short-duration grazing, depending on stocking rate (Heitschmidt, Downhower and Walker, 1987). In New Mexico, on blue grama rangeland, short-duration grazing apparently benefited blue grama compared with continuous grazing (White et al., 1991). Stocking rate was more influential than grazing system in most of these studies (Bryant et al., 1989; Holechek, Pieper and Herbel, 2001; Pieper and Heitschmidt, 1988).
Short-duration grazing apparently reduced infiltration and increased runoff compared with non-grazed or continuous grazing conditions in grasslands (McCalla, Blackburn and Merrill, 1984; Pluhar, Knight and Heitschmidt, 1987; Weltz and Wood, 1986). Concentrating livestock, even for short periods, tends to compact the soil and negates any possible benefit from hoof action. However, in New Mexico, infiltration and runoff had returned nearly to normal in short-duration pastures following the rest period (Weltz and Wood, 1986).
Since grasslands are generally grazed seasonally, provision for feed for the rest of the year is necessary. The use of complementary pastures along with native range is one approach for meeting the nutritional needs of livestock during periods when grazing of native rangeland is not practical (Gillen and Berg, 2001; Hart et al., 1988; Hoveland, McCann and Hill, 1997; Keller, 1960; Lodge, 1970; Nichols, Sanson and Myran, 1993; Smoliak, 1968). Such complementary pastures may involve old world bluestems (Gillen and Berg, 2001), introduced grasses such as crested (Plate 6.18) and other wheatgrasses (Holechek, 1981; Rogler, 1960), other cool-season grasses (Nichols, Sanson and Myran, 1993) and legumes.
Fertilization is another practice used to enhance livestock performance in the Great Plains (Nyren, 1979; Wight, 1976). Nitrogen is most often the limiting nutrient, but in some cases phosphorus and potassium may also be limiting (Nyren, 1979; Vallentine, 1989; Wight, 1976). An extensive literature on range fertilization has developed that shows, in general, greater vegetational response in northern mixed prairie rangelands than in southern areas (Vallentine, 1989). Nitrogen fertilization may change species composition by favouring cool-season species if applications are made early in the growing season (Nyren, 1979; Vallentine, 1989). In northern mixed prairie, early application of nitrogen may stimulate the aggressive cool-season species western wheatgrass (originally Agropyron smithii, now Pascopyrum smithii) at the expense of warm-season species (Nyren, 1979). High nitrogen fertilizer rates late in the growing season to benefit warm-season species such as blue grama may stimulate cool-season species the next spring (Wight, 1976). In southern areas, cool-season introduced species such as Kentucky bluegrass (Poa pratensis) may be stimulated (Owensby, 1970; Rehm, Sorensen and Moline, 1976; Vallentine, 1989).

Plate 6.18
Crested wheatgrass (Agropyron
cristatum).
S.G. REYNOLDS
Power (1972) argued that on many northern Great Plains rangelands, inorganic nitrogen is immobilized when nitrogen is added as fertilizer and sufficient nitrogen must be added to overcome that immobilized. He stated that the system could be maintained if annual fertilizer additions plus mineralization equals immobilization plus irreversible losses.
Even though substantial responses in herbage yield can be accomplished with range fertilization in grasslands, the practice may be only marginally feasible economically. In blue grama rangeland in south-central New Mexico, despite doubling of herbage and cattle production from annual additions of 40 kg/ha, economic returns were marginal (Chili et al., 1998). The economics of range fertilization depend mostly on cost of fertilizer and livestock prices.

Plate 6.19
Prescribed burn to control aspen
growth on the mixed -grass of the Wolverine Prairie Farm Rehabilitation Area,
Saskatchewan, Canada.
MAE ELSINGER
Fire is another useful tool in managing Great Plains grassland (Wright, 1974, 1978; Wright and Bailey, 1982). Vallentine (1989) lists 18 separate objectives in rangeland burning but suggests that there are three main reasons to burn: 1. To kill or suppress undesirable brush plants (Plate 6.19). 2. To prevent invasion of inferior species in the understorey. 3. To increase forage production and thus grazing capacity.
Especially in tall-grass prairie, prescribed burning is often used to reduce old growth and stimulate new, more palatable growth (Anderson, Smith and Owensby, 1970; McMurphy and Anderson, 1965; Smith and Owensby, 1972). Wright (1978) suggested that burning in tall-grass vegetation increased palatability, suppressed encroachment of trees and shrubs and reduced competition from cool-season plants. However, timing of the burning is very important. Cool-season grasses are detrimentally affected by spring burning (Hensel, 1923; Wright, 1978). Spring burning tends to increase summer gains of cattle, but gains may not hold up into the autumn (Anderson, Smith and Owensby, 1970; Vallentine, 1989). Late winter burning may initiate spring growth two to three weeks earlier than in the absence of burning (Ehrenreich and Aikman, 1963).
Grasslands are very dynamic, both spatially and temporally (Dix, 1964; Sims, 1988). Consequently, different authors have considered different factors as influential in the development of grassland vegetation and animal communities. Since grasslands operate as ecosystems, it may be futile to try to isolate causal factors, since some internal components may interact with others to change the nature of the system. For example, Larson (1940) considered that heavy bison grazing prior to settlement helped maintain the short-grass prairie. However, bison evolved under grassland conditions - vegetation, climate, other herbivores and predators - and attaching causality to one component may involve circular reasoning.
Sauer (1950), in contrast, argued that grasslands were maintained by periodic fires. Without fires, grasslands would progress to woodlands or forest. Indeed, grasslands of central North America have developed under frequent fire regimes, caused both naturally and by man (Flores, 1999; Sauer, 1950). Flores (1999) suggested that natural lightning fires had results that differed from those set by native Americans. He argued that maintenance of southern Great Plains grasslands populated by large grazing animals depended on fire management by native Americans.
One prominent feature of grasslands is a variable climate, with periodic droughts a common feature (Dix, 1964). The drought of the 1930s that resulted in the dust bowl in the Great Plains has been well documented (Albertson and Weaver, 1942; Robertson, 1939; Savage, 1937; Weaver and Albertson, 1936, 1939, 1940; Whitman, Hanson and Peterson, 1943). During the drought of the 1930s, blue grama abundance was reduced dramatically (by as much as 70 to 80 percent) in Kansas (Weaver and Albertson, 1956). In North Dakota mixed prairie, blue grama was reduced to about 40 percent of pre-drought (1933) levels in 1936-37 (Whitman, Hanson and Peterson, 1943). Other species negatively affected by the drought were western wheatgrass, needle-and-thread and prairie junegrass (Koeleria macrantha). The one species not affected by the drought was threadleaf sedge (Carex filifolia) (Whitman, Hanson and Peterson, 1943).
In western Nebraska, tree ring analyses indicated that over the last 400 years nearly 160 were "drought " years and 237 "wet" years (Weakly, 1943). Borchert (1950) reported that drought years tended to be clumped, with an average duration of nearly 13 years, compounding the effects of the drought.
Geologically, the Great Plains consists of a valley between the relatively young Rocky Mountains to the west and the older Appalachian chain to the east. The valley is drained by the Missouri and Mississippi River systems, which have carried sediment from both the Rocky and Appalachian Mountains and sorted and deposited these sediments (Dix, 1964).
In geological time, the central North America n grasslands have undergone many transformations since the late Cretaceous period (Axelrod, 1958; Dix, 1964; Donart, 1984). At this time the deciduous Pan Tropical Forest covered most of the present USA (Axelrod, 1979). From these forests, two distinct forests developed during Eocene times: the Arcto-tertiary and Neotropical-tertiary forests (Dix, 1964; Axelrod, 1958). Both of these forests contributed to the central grasslands that probably developed in more recent times (Pliocene and Pleistocene series) as the forests retreated (Dix, 1964). These vegetational shifts were also accompanied by climatic shifts, geologic events and changing patterns of herbivore utilization of these grasslands. Glaciation during the Pleistocene had dramatic influences on landscapes and vegetation, but the influence on the grasslands is not completely understood (Dix, 1964; Flint, 1957; Love, 1959).
Traditionally, research on grasslands of the Great Plains has been conducted by the Land Grant Universities. These were established by the Morrill Act of 1862, whereby one university in each state was designated the Land Grant University (Holechek, Pieper and Herbel, 2001). The Hatch Act of 1887 and the Smith-Lever Act of 1914 completed the tripartite focus of the Land Grant Universities in teaching, research and service (National Research Council, 1996). Each of these universities in the grassland states has been conducting research into agricultural practices and they have received research support from federal, state and other grant sources. Departments involved in these efforts have been those of Agronomy, Horticulture, Agricultural Economics, Animal Science and allied disciplines. The Cooperative Extension Service has served as the link between the research and application by farmers and ranchers (see Plates 6.20 and 6.21 with examples from Canada). Range research in the Land Grant Universities has been conducted by researchers attached to other departments in the main grassland states: with Animal Science in Montana, New Mexico, North and South Dakota; and with Agronomy in Nebraska, Kansas and Oklahoma. The School of Forestry at the University of Montana also has a range faculty offering degrees in Range Science (Bedell, 1999). Colorado State University has a separate Range Science Department, as does Texas Tech (with Wildlife Science) and Texas A & M University.

Plate 6.20
Plant identification by range
management officers on native prairie in the mixed grass zone at Big Muddy,
Saskatchewan, Canada.
MAE ELSINGER

Plate 6.21
Pasture sampling in the mixed -grass
zone, Saskatchewan, Canada.
MAE ELSINGER
Research in grassland agriculture has also been conducted by the Agriculture Research Service. Experimental stations have concentrated on range and livestock problems in the Northern Great Plains in North Dakota; Sydney, Montana (now closed); Central Plains in Colorado; and the Southern Great Plains in Oklahoma. Research conducted at the universities and Agricultural Research Service (ARS) experimental stations has been broad based, featuring both basic and applied research.
Grassland research has also been conducted by ecologists in Biology or Botany Departments in Great Plains states. The University of Nebraska was in the forefront of ecological research under the direction of Dr John E. Weaver during the 1930s and 1940s, although earlier scientists, including Dr Frederic E. Clements, were instrumental in establishing ecology at the University of Nebraska (Tobey, 1981). Drs Clair Kucera at the University of Missouri, Warren Whitman at North Dakota State University, Lloyd C. Hulbert at Kansas State University, William Penfound, Elroy Rice and Paul G. Risser at Oklahoma State University and John Aikman at Iowa State University also had strong grassland research programmes (Kucera, 1973; Tobey, 1981).
In the late 1960s and early 1970s, another major research effort in grassland ecology was launched with the International Biological Programme (IBP) (Golley, 1993). The Grassland Biome was established with its focus at Colorado State University, with Dr George M. Van Dyne as Director. Satellite research areas were established at the Cottonwood site in mixed prairie vegetation in South Dakota (operated by South Dakota State University); tall-grass vegetation in Oklahoma at the Osage site (operated by Oklahoma State University); and short-grass vegetation at the Pantex Site in Texas (operated by Texas Tech University) and the Pawnee Site in Colorado (operated by Colorado State University) (Van Dyne, Jameson and French, 1970). Other grassland sites, such as desert grassland, mountain grassland and California annual grassland, were also included in the project. Although the project generated much ecological information and extensive literature on aspects of grassland ecology, the goal of publishing a synthesis volume for each of the grassland types was not realized (Golley, 1993). Only the tall-grass volume was actually published (Risser et al., 1981).
The other major outcome of the IBP programme was establishment of Long-Term Ecological Research (LTER) sites. The argument was made that ecological problems could only be approached by looking at dynamics of ecosystems in a long time frame - 10 years or more. The Conyza Prairie in Kansas (Knapp et al., 1998) and the Central Plains Experimental Range (the old Pawnee Site of the IBP) are currently LTER sites.
Great Plains agriculture is now facing many challenges from various sources. This analysis will focus on only a few of these. In the Great Plains, as well as most of the West, many small towns and communities are facing extreme economic conditions and many are being abandoned (Flores, 1999; Licht, 1997). Licht (1997) reported that 81 percent of the Great Plains counties lost population between 1980 and 1990. There are probably multiple reasons for this decline in rural communities such as reliance on railroads; influence of technology and government; better transportation to larger urban areas; location of agricultural agents in county seats; and governmental policies (Burns, 1982). However, Licht (1997) argues that:
"...the main reason for the collapse of rural communities in the Great Plains is indisputable; the region’s inhospitable climate, lack of economically valuable natural resources, high transportation costs and other factors meant that it was never capable of supporting numerous vibrant economies with high human densities."
Along with the constriction of rural communities, many agricultural enterprises have been caught in an economic squeeze, with, on the one hand, high production costs and, on the other, low prices for their products. This is true in both intensive agriculture and the livestock sector. Consequently, many small-scale operators (such as family farms) are no longer able to operate economically (Licht, 1997).
Other biological problems relate to environmental concerns. Even though Lauenroth et al. (1994) considered that grazing lands in the Great Plains had changed little since European settlement, farming and other disturbances have had a great effect. For example, Klopatek et al. (1979) showed that most counties in the Great Plains had lost some of the potential natural vegetation, with the greatest impact in the mesic eastern edge and the least disturbed being the more xeric western short-grass plains. From 85 to 95 percent of the bluestem prairie vegetation types had been converted to cropland (Sieg, Flather and McCanny, 1999). These types of disturbance and vegetation shifts represent habitat fragmentation for many wildlife species that developed in unbroken tracts of grassland. Conversion to cropland created habitats for other species, but loss of both plant and animal diversity in the grassland is a concern (Sieg, Flather and McCanny, 1999; Leach and Givnish, 1996; Licht, 1997). For example Sieg, Flather and McCanny (1999) reported that in the Canadian province of Saskatchewan and six Great Plains states in the USA, 19 percent of the breeding bird species declined in numbers from 1966 to 1996. However, the number of listed, threatened and endangered (LT&E) plant and animal species is relatively low compared with other regions of the country (Ostlie et al., 1997; Sieg, 1999). No pattern of LT&E species could be discerned for the Great Plains. Large blocks of counties showed no known LT&E species, up to a maximum of 9-12 species. These counties were scattered throughout the Great Plains (Sieg, 1999). Examples of LT&E species are the black -tailed prairie dog (Cynomys ludovicianus), the black-footed ferret (Mustela nigrepes) and the western prairie fringed orchid (Platanthera praeclara) (Sieg, Flather and McCanny, 1999).
Other environmental concerns regard use of pesticides and herbicides, commercial fertilizers, grazing by introduced domestic livestock, status of riparian areas and introduction of invasive plants and animals. Crested wheatgrass was introduced into the Great Plains in the early 1900s (Holechek, 1981; Rogler, 1960). The species played a major role in restoration of abandoned wheat fields in the northern Great Plains in the 1930s (Rogler, 1960), but some workers regard crested wheatgrass as an invader species leading to near monocultures.
One of the most troublesome introduced plant species is leafy spurge (Euphorbia esula) (Plate 6.22), a perennial forb accidentally introduced into the USA from eastern Europe or western Asia (Biesboer and Koukkari, 1992). It is an aggressive weed that currently infests over one million hectares in the USA (DiTomaso, 2000) and over 650 000 ha in the northern Great Plains (Leistritz, Leitch and Bangsund, 1995). Estimates of loss of livestock grazing because of leafy spurge encroachment onto northern Great Plains rangelands were 736 000 Animal Unit Months (AUMs), or US$ 37 million annually (Leistritz, Leitch and Bangsund, 1995).
Introduction of annual grasses such as Japanese brome (Bromus japonicus) has also altered grassland vegetation (Haferkamp et al., 1993; Haferkamp, Heitschmidt and Karl, 1997). Japanese brome occurs throughout the Great Plains (Hitchcock, 1950) and often competes with native perennial grasses.

Plate 6.22
Leafy spurge (Euphorbia esula)
infestation on sandy soils in the mixed -grass zone, Saskatchewan,
Canada.
MAE ELSINGER
Haferkamp, Heitschmidt and Karl (1997) reported that presence of Japanese brome reduced yield of western wheatgrass in eastern Montana, but removal of Japanese brome reduced total standing crop since other species did not completely replace the brome. In addition to economic consequences, Huenneke (1995) and Hobbs and Huenneke (1992) listed the following ecological impacts of plant invasions (for lack of a more appropriate word): spread of toxic substances, replacement of native species, alteration of hydrological characteristics, alteration of soil properties, and changed nutrient cycling.
Other suggestions for managing the Great Plains include increasing the number of national grasslands and developing a "buffalo commons" for certain portions of the Great Plains (Licht, 1997; Popper and Popper, 1994). Although such proposals have appeal to some environmental groups, Licht (1997) discusses several limitations of such proposals.
Riparian areas in the Great Plains are also of concern, as they are in many other areas of the country (Johnson, 1999). These systems occupy less than one percent of the land surface, yet are vitally important for catchment processes; plant and animal diversity; and uses by man, including both industrial and agricultural purposes (Johnson and McCormick, 1979; Swanson, 1988). Riparian systems in the Great Plains have been modified by human activities such as clearing for agriculture, grazing, canalization, damming and water diversion (Johnson, 1999). Johnson (1999) presents case studies on how these activities have altered the Missouri River in North Dakota, the Platte River in Nebraska and Foster Creek in South Dakota.
It is likely that agriculture will continue to dominate the Great Plains into the foreseeable future. While technology will continue to develop new approaches, such as no-till cultivation, more efficient use of water and fertilizer, and methods to survey and monitor landscapes, some of these technologies will be difficult to apply because of economic, sociological and biological constraints.
For example, we now have the technology to consider management at relatively large scales (Ludwig et al., 1997). Modern tools. including remote sensing (Tueller, 1989), geographical information systems (GIS) and global positioning systems (GPS), provide the opportunity to consider land management and ecological situations across landscapes and habitats. However, these approaches need to be applied with consideration of some of the limitations, such as lack of adequate ground truth data for remote sensing and GIS applications. Development of landscape -scale planning for the Great Plains might entail consideration of crop agriculture areas, riparian habitats and "natural " vegetation types. The extent and arrangement of agricultural areas and grassland vegetation types would be challenging, even if all those concerned could agree on percentages of land areas devoted to each and the spatial distribution of land devoted to different or several uses because of differences in land ownership patterns and extent of agricultural development. Developments such as the buffalo commons would be difficult to effect because of the large extent of private land in the region. Eventually, lack of water resources will have heavy impacts on both agricultural and industrial development.
Major cities in the Great Plains may not expand into adjacent farmland and wild land to the same extent as those in other western cities, but there will probably be some expansion. Cities along the western edge of the Great Plains - Fort Collins, Denver, Colorado Springs, Pueblo, etc. - will continue to grow because of favourable perceptions of location near the mountains.
Livestock enterprises, although stressed economically, will probably remain relatively stable. Walker (1995) stated that grazing systems have largely not changed the selective nature of livestock grazing. Stocking rate is the primary factor determining livestock and vegetational responses (Holechek, 1988; Walker, 1995). Competition with forages produced under irrigation will probably continue to erode livestock production from rangelands in the Great Plains (Glimp, 1991).
Genetic modification of both plants and animals has the potential to change plant and animal agriculture in the Great Plains (Walker, 1997). However, public acceptance of genetically modified plants and animals will influence how fast these technologies are used.
Adams, D.C., Clark, R.T., Klopfenstein, T.J. & Volesky, J.D. 1996. Matching the cow with the forage resources. Rangelands, 18: 57-62.
Albertson, F.W. & Weaver, J.E. 1942. History of the native vegetation of western Kansas during seven years of continuous drought. Ecological Monographs, 12: 23-51.
Anderson, K.L, Smith, E.F. & Owensby, C. 1970. Burning bluestem range. Journal of Range Management, 23: 81-92.
Axelrod, D.E. 1958. Evolution of the Madro-tertiary Geoflora. Botanical Review, 24: 434-509.
Axelrod, D.E. 1979. Desert vegetation, its age and origin. pp. 75-82, in: J.R. Goodin and D.K. Northington (eds). Aridland plant resources. Lubbock, Texas, USA: ICASALS, Texas Tech University Press.
Ayyad, M.A.G. & Dix, R.L. 1964. An analysis of a vegetation -microenvironemtal complex on prairie slopes in Saskatchewan. Ecological Monographs, 34: 421-442.
Barbour, M.G., Burk, J.H. & Pitts, W.D. 1987. Terrestrial plant ecology. 2nd ed. Menlo Park, California, USA: The Benjamin Cummings Pub. Co.
Baxter, F.P. & Hole, F.D. 1967. Ant (Formica cinerea) pedoturbation in a Prairie soil. Soil Science Society of America Proceedings, 31: 425-428.
Bazzaz, F.A. & Parrish, J.A.D. 1982. Organization of grassland communities. pp. 233-254, in: J.R. Estes, R.J. Tyrl and J.N. Brunken (eds). Grasses and Grasslands. Systematics and ecology. Norman, Oklahoma, USA: University of Oklahoma Press.
Beavis, W.D., Owens, J.C., Ortiz, M., Bellows, T.S. Jr., Ludwig, J.A. & Huddleston, E.W. 1981. Density and developmental stage of range caterpillar Hemilueca oliviae Cockerill, as affected by topographic position. Journal of Range Management, 34: 389-392.
Bedell, T.E. 1999. The educational history of range management in North America. Denver, Colorado, USA: Society for Range Management.
Bement, R.E. 1969. A stocking-rate guide for beef production on blue -grama range. Journal of Range Management, 22: 83-86.
Bement, R.E., Barmington, R.D., Everson, A.C., Hylton, L.O. Jr. & Remmenga, E.E. 1965. Seeding of abandoned cropland in the Central Great Plains. Journal of Range Management, 18: 53-65.
Bernardo, D.B. & McCullom, F.T. 1987. An economic analysis of intensive-early stocking. Oklahoma State University Agricultural Experiment Station Research Report, No. 887.
Biesboer, D.D. & Koukkari, W.L. 1992. The taxonomy and biology of leafy spurge. pp. 51-57, in: R.A. Masters, S.J. Nissen and G. Friisoe (eds). Leafy spurge symposium. Lincoln, Nebraska, USA, 1992. Lincoln, Nebraska, USA: Dow Elanco and Nebraska Leafy Spurge Working Task Force.
Blocker, H.D. 1970. The impact of insects in grassland ecosystems. pp. 290-299, in: R.L. Dix and R.G. Beidleman (eds). The grassland ecosystem. Range Science Department Series, No. 2. Colorado State University, Fort Collins, Colorado, USA.
Bolen, E.C. & Crawford, J.A. 1996. The birds of rangelands. pp. 15-27, in: P.R. Krausman (ed). Rangeland wildlife. Denver, Colorado, USA: Society for Range Management.
Borchert, L.R. 1950. The climate of the central North America n grassland. Annals of the Association of American Geographers, 40: 1-39.
Breymeyer, A.I. & Van Dyne, G.M. (eds). 1979. Grasslands, systems analysis and management. New York, New York, USA: Cambridge University Press.
Bryant, F.C., Dahl, B.E., Pettit, R.D. & Britton, C.M. 1989. Does short-duration grazing work in arid and semiarid regions? Journal of Soil and Water Conservation, 44: 290-296.
Burns, N. 1982. The collapse of small towns on the Great Plains: a bibliography. Emporia State Research Studies, Emporia, Kansas, USA.
Buol, S.W., Hole, F.D. & McCracken, R.J. 1980. Soil genesis and classification. 2nd ed. Ames, IA, USA: Iowa State University Press.
Burzlaff, D.F. & Harris, L. 1969. Yearling steer gains and vegetation changes of western Nebraska rangeland under three rates of stocking. University of Nebraska Agricultural Experiment Station Bulletin, No. SB.505.
Chili, P., Donart, G.B., Pieper, R.D., Parker, E.E., Murray, L.W. & Torell, L.A. 1998. Vegetational and livestock response to nitrogen fertilization in south-central New Mexico. New Mexico State University Agricultural Experiment Station Bulletin, No. 778.
Costello, D.F. 1944. Natural revegetation of abandoned plowed land in the mixed prairie association of northeastern Colorado. Ecology, 25: 312-326.
Deneven, W. 1996. Carl Sauer and native American population size. The Geographical Review, 86: 385-397.
Dahl, B. 1994. Mesquite-buffalograss. pp. 27-28, in: T.N. Shiflet (ed). Rangeland cover types of the United States. Denver, Colorado, USA: Society for Range Management.
Danz, D.P. 1997. Of bison and man: from the annals of a bison yesterday to a refreshing outcome from human involvement with America’s most valiant of beasts. Boulder, Colorado, USA: University of Colorado Press.
DiTomaso, J.M. 2000. Invasive weeds in rangelands: species, impacts and management. Weed Science, 48: 255-265.
Dix, R.L. 1964. A history of biotic and climatic changes within the North American Grassland. pp. 71-89, in: D.J. Crisp (ed). Grazing in terrestrial and marine environments. Oxford, UK: Blackwell Scientific.
Dodd, J.L. & Lauenroth, W.K. 1979. Analysis of the response of a grassland ecosystem to stress. pp. 43-58, in: N.R. French (ed). Perspectives in grassland ecology. New York, New York, USA: Springer-Verlag.
Donart, G.B. 1984. The history and evolution of western rangelands in relation to woody plant communities. pp. 1235-1258, in: Developing strategies for rangeland management. Boulder, Colorado, USA: Westview Press.
Donohue, D.L. 1999. The western range revisited. Removing livestock from public lands to conserve native biodiversity. Norman, Oklahoma, USA: University of Oklahoma Press.
Engle, D. 1994. Cross timbers - Oklahoma. p. 37, in: T.N. Shiflet (ed). Rangeland cover types of the United States. Denver, Colorado, USA: Society for Range Management.
Ehrenreich, J.H. & Aikman, J.M. 1963. An ecological study of certain management practices on native plants in Iowa. Ecological Monographs, 33: 113-130.
Engle, D.M. 2000. Eastern redcedar (Juniperus virginiana). Expanded abstract. 53rd Annual Meeting of the Society for Range Management, Boise, Idaho, USA. Society for Range Management, Denver, Colorado, USA.
Ensminger, M.E. & Parker, R.O. 1986. Sheep and goat science. 5th ed. Danville, Illinois, USA: Interstate Printers and Pub.
Fisher, C.E. & Marion, P.T. 1951. Continuous and rotation grazing on buffalo and tobosa grassland. Journal of Range Management, 4: 48-51.
Flint, R.F. 1957. Glacial and Pleistocene geology. New York, New York, USA: John Wiley and Sons.
Flores, D. 1999. Essay: the Great Plains "wilderness" as a human-shaped environment. Great Plains Research, 9: 343-355.
Freckman, D.W., Duncan, D.A. & Larson, J.R. 1979. Nematode density and biomass in an annual grassland ecosystem. Journal of Range Management, 32: 418-422.
Garretson, M.S. 1938. The American bison. New York, New York, USA: New York Zoological Society.
Gehring, J.L. & Bragg, T.B. 1992. Changes in prairie vegetation under eastern red cedar (Juniperus virginiana L.) in an eastern Nebraska bluestem prairie. American Midland Nauralist, 128: 209-217.
Gillen, R.L. & Berg, W.A. 2001. Complementary grazing of native pasture and old world bluestem. Journal of Range Management, 54: 348-355.
Gleason, H.A. 1913. The relation of forest distribution and prairie fires in the Middle West. Torreya, 13: 173-181.
Gleason, H.A. & Cronquist, A. 1964. The natural geography of plants. New York, New York, USA: Columbia University Press.
Glimp, H.A. 1991. Can we produce lambs for $0.40/lb? Symposium Proceedings - Sheep Forage Production Systems. Denver, Colorado, USA, 1991. Denver, Colorado, USA: American Sheep Industry.
Glover, F.A. 1969. Birds in grassland ecosystems. pp. 279-289, in: R.L. Dix and R.G. Beidleman (eds). The grassland ecosystem. A preliminary synthesis. Range Science Department Scientific Series, No. 21. Colorado State University, Fort Collins, Colorado, USA.
Goetz, H. 1989. The conservation reserve program - where are we heading? Rangelands, 11: 251-252.
Golley, F.B. 1993. A history of the ecosystem concept in ecology: more than the sum of the parts. New Haven, Conneticut, USA: Yale University Press.
Guthery, F.S. 1996. Upland gamebirds. pp. 59-69, in: P.R. Krausman (ed). Rangeland wildlife. Denver, Colorado, USA: Society for Range Management.
Gunderson, J. 1981. True prairie - past and present. Rangelands, 3: 162.
Haferkamp, M.R., Karl, M.G., MacNeil, M.D., Heitschmidt, R.K. & Young, J.A. 1993. Japanese brome in the northern Great Plains. pp. 112-118, in: Research & Rangeland Agriculture: past, present & future. USDA Ft. Keogh Livestock and Range Research Laboratory, Miles City, Montana, USA.
Haferkamp, M.R., Heitschmidt, R.K. & Karl, M.G. 1997. Influence of Japanese brome on western wheatgrass yield. Journal of Range Management, 50: 45-50.
Harlan, J.R. 1960. Production characteristics of Oklahoma forages, native range. Oklahoma State University Agricultural Experiment Station Bulletin, No. B547.
Hart, R.H. & Samuel, M.J. 1985. Precipitation, soils and herbage production on southeast Wyoming range sites. Journal of Range Management, 38: 522-525.
Hart, R.H., Samuel, M.J., Test, P.S. &Smith, M.A. 1988. Cattle, vegetation and economic responses to grazing systems and grazing pressure. Journal of Range Management, 41: 282-286.
Hart, R.H., Waggoner, J.W. Jr., Dunn, T.G., Kaltenbach, C.C. & Adams, L.D. 1988. Optional stocking rate for cow-calf enterprises on native range and complementary improved pastures. Journal of Range Management, 41: 435-440.
Heimlich, R.E. & Kula, O.E. 1989. Grazing lands: how much CRP land will remain in grass? Rangelands, 11: 253-257.
Heitschmidt, R.K., Downhower, S.L. & Walker, J.W. 1987. Some effects of a rotational grazing treatment on quantity and quality of available forage and amount of ground litter. Journal of Range Management, 40: 318-321.
Heitschmidt, R.K., Kothmann, M.M. & Rawlins, W.J. 1982. Cow-calf response to stocking rates, grazing systems, and winter supplementation at the Texas Experimental Ranch. Journal of Range Management, 35: 204-210.
Hensel, R.L. 1923. Recent studies on the effect of burning on grassland vegetation. Ecology, 4: 183-188.
Herbel, C.H. 1974. A review of research related to development of grazing systems on native ranges of the western United States. pp. 138-149, in: K.W. Kreitlow and R.H. Hart (Coordinators). Plant morphogenesis as the basis for scientific management of range resources. USDA Miscellaneous Publication, No. 1271.
Herbel. C.H. & Anderson, K.L. 1959. Response of true prairie vegetation on major Flint Hills range sites to grazing treatment. Ecological Monographs, 29: 171-186.
Herbel, C.H. & Pieper, R.D. 1991. Grazing management. pp. 361-385, in: J. Skujins (ed). Semiarid lands and deserts. Soil resource and reclamation. New York, New York, USA: Marcel Dekker.
Hewitt, G.B. 1977. Review of forage losses caused by rangeland grasshoppers. USDA Miscellaneous Publication, No. 1348. Washington, D.C., USA
Hewitt, G.B. & Onsager, J.A. 1983. Control of grasshoppers on rangeland of the United States - a perspective. Journal of Range Management, 36: 202-297.
Hickey, W.C. Jr. 1969. A discussion of grazing management systems and some pertinent literature (abstracts and excerpts) 1895-1966. USDA Forest Service Regional Office, Denver, Colorado, USA.
Hitchcock, A.S. 1950. Manual of the grasses of the United States. USDA Miscellaneous Publication, No. 200. U.S. Printing Office, Washington, D.C., USA
Hobbs, R.J. & Huenneke, L.F. 1992. Disturbance, diversity and invasion: implications for conservation. Conservation Biology, 6: 324-337.
Hole, F.D. & Nielsen, G.A. 1968. Some processes of soil genesis under prairie. Proceedings of a Symposium on Prairie and Prairie Restoration. Galesburg, Illinois, USA, 1968. Galesburg, Illinois, USA: Knox College.
Holechek, J.L. 1981. Crested wheatgrass. Rangelands, 3: 151-153.
Holechek, J.L. 1984. Comparative contribution of grasses, forbs and shrubs to the nutrition of range ungulates. Rangelands, 6: 245-248.
Holechek, J.L. 1988. An approach for setting the stocking rate. Rangelands, 10: 10-14.
Holechek, J.L., Pieper, R.D. & Herbel, C.H. 2001. Range management principles and practices. Upper Saddle River, New Jersey, USA: Prentice Hall.
Hoveland, C.S., McCann, M.A. & Hill, N.S. 1997. Rotational vs. continuous stocking of beef cows and calves on mixed endophyte-free tall fescue-bermudagrass pasture. Journal of Production Agriculture, 10: 193-194.
Huenneke, L.F. 1995. Ecological impacts of plant invasions in rangeland ecosystems. Abstracts of the Annual Meeting of the Society for Range Management. Phoenix, Arizona, USA, 1992. Society for Range Management, Denver, Colorado, USA.
Johnson, R.R. & McCormick, J.F. 1979. Strategies for protection and management of floodplain wetlands and other riparian ecosystems. USDA Forest Service General Technical Report, No. WO-12.
Johnson, W.C. 1999. Response of riparian vegetation to streamflow regulation and land use in the Great Plains. Great Plains Research, 9: 357-369.
Jordan, C.F. 1995. Conservation. Replacing quantity with quality as a goal for global management. New York, New York, USA: John Wiley & Sons.
Joyce, L.A. 1989. An analysis of the range forage situation in the United States, 1989- 2040. USDA Forest Service General Technical Report, No. RM-180.
Keller, W. 1960. Importance of irrigated grasslands in animal production. Journal of Range Management, 13: 22-28.
Klipple, G.E. & Costello, D.F. 1960. Vegetation and cattle response to different intensities of grazing on short-grass ranges of the central Great Plains. USDA Agricultural Technology Bulletin, No. 1216.
Klopatek, J.M., Olson, R.J., Emerson, C.J. & Jones, J.L. 1979. Land-use conflicts with natural vegetation in the United States. Environmental Sciences Division, Oak Ridge National Laboratory Publication, No. 1333. Oak ridge, Tennessee, USA.
Knapp, A.K, Briggs, J.M., Harnett, D.C. & Collins, S.L. 1998. Grassland dynamics: long-term ecological research in tallgrass prairie. New York, New York, USA: Oxford University Press.
Knopf, F.L. 1996. Perspectives on grazing nongame bird habitats. pp. 51-58, in: P.R. Krausmann (ed). Rangeland wildlife. Denver, Colorado, USA: Society for Range Management.
Kretzer, J.E. & Cully, J.F. Jr. 2001. Effects of black -tailed prairie dogs on reptiles and amphibians in Kansas shortgrass prairie. Southwestern Naturalist, 46: 171-177.
Kucera, C.L. 1973. The challenge of ecology. Saint Louis, Missouri, USA: C.V. Mosby.
Lacey, J., Studiner, S. & Hacker, R. 1994. Early spring grazing on native range. Rangelands, 16: 231-233.
Larson, F. 1940. The role of the bison in maintaining the short grass plains. Ecology, 21: 113-121.
Lauenroth, W.K. 1979. Grassland primary production: North American grasslands in perspective. pp. 3-24, in: N.R. French (ed). Perspectives in grassland ecology. New York, New York, USA: Springer-Verlag.
Lauenroth, W.K., Milchunas, D.G., Dodd, J.L., Hart, R.H., Heitschmidt, R.K. & Rittenhouse, L.R. 1994. Effects of grazing on ecosystems of the Great Plains. pp. 69-100, in: M. Vavra, W.A. Laycock and R.D. Pieper (eds). Ecological implications of livestock herbivory in the West. Denver, Colorado, USA: Society for Range Management.
Lauenroth, W.K., Burke, I.C. & Gutmann, M.P. 1999. The structure and function of ecosystems in the Central North American Grassland Region. Great Plains Research, 9: 223-259.
Launchbaugh, J.L. & Owensby, C.E. 1978. Kansas rangelands: their management based upon a half century of research. Kansas State University Agricultural Experiment Station Bulletin, No. 622.
Laycock, W.A. 1979. Introduction. pp. 1-2, in: N.R. French (ed). Perspectives in grassland ecology. New York, New York, USA: Springer-Verlag.
Leach, M.K. & Givnish, T.J. 1996. Ecological determinants of species loss in remnant prairies. Science, 273: 1555-1558.
Leistritz, F.L, Leitch, J.A. & Bangsund, D.A. 1995. Economic impact of leafy spurge on grazingland and wildland in the northern Great Plains. Abstracts of the Annual Meeting of the Society for Range Management. Phoenix, Arizona, USA, 1995. Society for Range Management, Denver, Colorado, USA.
Leopold, A.S., Cain, S.A., Cottan, C.M., Gabrielson, I.N. & Kinball, T.L. 1963. Wildlife management in the National Parks: the Leopold Report. Advisory Board on Wildlife Management, Washington, D.C., USA.
Licht, D.S. 1997. Ecology and economics of the Great Plains. Lincoln, Nebraska, USA: University of Nebraska Press.
Lodge, R.W. 1970. Complementary grazing systems for the northern Great Plains. Journal of Range Management, 23: 268-271.
Love, D. 1959. The post-glacial development of the flora of Manitoba: a discussion. Canadian Journal of Botany, 37: 547-585.
Ludwig, J., Tongway, D., Fruedenberger, D., Noble, J. & Hodgkinson, K. 1997. Landscape ecology, function and management: principles from Australia’s rangelands. Collingwood, Australia: CSIRO Publications.
Malechek, J.C. 1984. Impacts of grazing intensity and specialized grazing systems on livestock responses. pp. 1129-1165, in: Developing strategies for rangeland management. Boulder, Colorado, USA: Westview Press.
Manley, W.A., Hart, R.H., Samuel, M.J., Smith, M.A., Waggoner, J.W. & Manley, J.T. 1997. Vegetation, cattle and economic responses to grazing strategies and pressures. Journal of Range Management, 50: 638-646.
McClendon, T. 1994. Mesquite. pp. 46-47, in: T.N. Shiflet (ed). Rangeland cover types of the United States. Denver, Colorado, USA: Society for Range Management.
McCalla, G.R. II, Blackburn, W.H. & Merrill, L.B. 1984. Effects of livestock grazing on infiltration rates, Edwards Plateau of Texas. Journal of Range Management, 37: 265-269.
McCollum, F.T., Gillen, R.L., Engle, D.M. & Horn, G.W. 1990. Stocker cattle performance and vegetation response to intensive-early stocking of Cross Timbers rangeland. Journal of Range Management, 43: 99-103.
McDaniel, B. 1971. The role of invertebrates in the grassland biome. In: Preliminary analysis of structure and function in grasslands. Range Science Department Scientific Series, No. 10. Colorado State University, Fort Collins, Colorado, USA.
McIlvain, E.H. & Shoop, M.C. 1969. Grazing systems in the southern Great Plains. Abstracts of the Annual Meeting of the American Society for Range Management, 22: 21-22.
McIlvain, E.H. & Savage, D.A. 1951. Eight-year comparisons of continuous and rotational grazing on the Southern Great Plains Experimental Range. Journal of Range Management, 4: 42-47.
McIlvain, E.H., Baker, A.L. Kneebone, W.R. & Gates, D.H. 1955. Nineteen-year summary of range improvement studies at the U.S. Southern Great Plains Field Station, Woodward, Oklahoma. USDA Agricultural Research Service Progress Reports, No. 5506.
McMurphy, W.E.L. & Anderson, K.L. 1965. Burning Flint Hills range. Journal of Range Management, 18: 265-269.
McPherson, G.R. 1997. Ecology and management of North American savannas. Tucson, AZ, USA: University of Arizona Press.
Miller, R.W. & Donohue, R.L. 1990. Soils - an introduction to soils and plant growth. 6th ed. Englewood Cliffs, New Jersey, USA: Prentice Hall.
Mitchell, J.E. 2000. Rangeland resource trends in the United States: A technical document supporting the 2000 USDA Forest Service RPA assessment. USDA Forest Service General Techical Report, No. RMRS-GTR-68.
National Research Council. 1996. Colleges of Agriculture at the Land Grant Universities. Public service and public policy. Board on Agriculture, National Research Council. Washington, D.C., USA: National Academy Press.
Nelson, E.W. 1925. Status of pronghorn antelope, 1922-24. USDA Bulletin, No. 1346. Washington, DC., USA.
Neubauer, T.A. 1963. The grasslands of the West. Journal of Range Management, 16: 327-332.
Neumann, A.L. & Lusby, K.S. 1986. Beef cattle. 7th ed. New York, New York, USA: John Wiley and Sons.
Newell, L.C. 1948. Hay, fodder and silage crops. pp. 281-287, in: Grass, the [USDA] Yearbook of Agriculture. Washington, DC., USA: U.S. Govt. Printing Office.
Nichols, J.T., Sanson, D.W. & Myran, D.D. 1993. Effect of grazing strategies and pasture species on irrigated pasture beef production. Journal of Range Management, 46: 65-69.
Nyren, P.E. 1979. Fertilization of northern Great Plains rangelands: a review. Rangelands, 1: 154-156.
Olson, K.E., Brethour, J.R. & Launchbaugh, J.L. 1993. Shortgrass range vegetation and steer growth response to intensive-early stocking. Journal of Range Management, 46: 127-132.
Ostlie, W.R., Schneider, R.E., Aldrich, J.M., Faust, T.M., McKim, R.L.B. & Chaplin, S.J. 1997. The status of biodiversity in the Great Plains. Arlington, Virginia, USA: The Nature Conservancy.
Owensby, C.E. 1970. Effects of clipping and supplemental nitrogen and water on loamy upland bluestem range. Journal of Range Management, 23: 341-346.
Pettit, R. 1994. Sand shinnery oak. pp. 42-43, in: T.N. Shiflet (ed). Rangeland cover types of the United States. Denver, Colorado, USA: Society for Range Management.
Pieper, R.D. 1980. Impacts of grazing systems on livestock. pp. 131-151, in: K.C. McDaniel and C.D. Allison (eds). Proceedings grazing management systems for southwest rangelands symposium.. Albuquerque, New Mexico, USA, 1980. Range Improvement Task Force, New Mexico State University, Las Cruces, New Mexico, USA.
Pieper, R.D. 1988. Rangeland vegetation productivity and biomass. pp. 449-467, in: P.T. Tueller (ed). Vegetation science applications for rangeland analysis and management. Dordrecht, The Netherlands: Kluwer Academic Publishers.
Pieper, R.D. 1994. Ecological implications of livestock grazing. pp. 177-211, in: M. Vavra, W.A. Laycock and R.D. Pieper (eds). Ecological implications of livestock herbivory in the West. Denver, Colorado, USA: Society for Range Management.
Pieper, R.D. & Beck, R.F. 1980. Importance of forbs on southwestern rangelands. Rangelands, 2: 35-36.
Pieper, R.D. & Heitschmidt, R.K. 1988. Is short-duration grazing the answer? Journal of Soil and Water Conservation, 43: 133-137.
Pieper, R.D., Nelson, A.B., Smith, G.S., Parker, E.E., Boggino, E.J.A. & Hatch, C.F. 1978. Chemical composition and digestibility of important range grass species in south-central New Mexico. New Mexico State University Agricultural Experiment Station Bulletin, No. 662.
Pieper, R.D., Parker, E.E., Donart, G.B., Wallace, J.D. & Wright, J.D. 1991. Cattle and vegetation response of four-pasture rotation and continuous grazing systems. New Mexico State University Agricultural Experiment Station Bulletin, No. 756.
Pitts, J.C. & Bryant, F.C. 1987. Steer and vegetation response to short duration and continuous grazing. Journal of Range Management, 40: 386-390.
Pluhar, J.J., Knight, R.W. & Heitschmidt, R.K. 1987. Infiltration rates and sediment production as influenced by grazing systems in the Texas rolling plains. Journal of Range Management, 40: 240-244.
Popper, D.E. & Popper, F.J. 1994. The buffalo commons: a bioregional vision of the Great Plains. Landscape Architecture, 84: 144-174.
Power, J.F. 1972. Fate of fertilizer nitrogen applied to a northern Great Plains rangeland ecosystem. Journal of Range Management, 25: 367-371.
Rao, M.R., Harbers, L.H. & Smith, E.F. 1973. Seasonal change in nutritive value of bluestem pastures. Journal of Range Management, 26: 419-422.
Redmann, R.E. 1975. Production ecology of grassland plant communities in western North Dakota. Ecological Monographs, 45: 83-106.
Rehm, G.W., Sorensen, R.C. & Moline, W.J. 1976. Time and rate of fertilizer application for seeded warm-season and bluegrass pastures. I. Yield and botanical composition. Agronomy Journal, 68: 759-764.
Risser, P.G., Birney, E.C., Blocker, H.D., May, S.W., Parton, W.J. & Wiens, J.A. 1981. The true prairie ecosystem. Stroudsburg, PA, USA: Hutchinson Ross.
Robertson, J.H. 1939. A quantitative study of true-prairie vegetation after three years of extreme drought. Ecological Monographs, 9: 431-492.
Rogler, G.A. 1951. A twenty-five year comparison of continuous and rotation grazing in the Northern Plains. Journal of Range Management, 4: 35-41.
Rogler, G.A. 1960. Growing crested wheatgrass in the western states. USDA Leaflet, No. 469.
Rogler, G.A. & Hurtt, L.C. 1948. Where elbowroom is ample. pp. 447-482, in: Grass, the [USDA] Yearbook of Agriculture. U.S. Printing Office, Washington, DC, USA.
Sauer, C.O. 1950. Grassland, fire and man. Journal of Range Management, 3: 16-21.
Savage, D.A. 1937. Drought, survival of native grass species in the central and southern Great Plains. USDA Technical Bulletin, No. 549.
Savory, A. 1983. The Savory grazing method or holistic resource management. Rangelands, 5: 555-557.
Savory, A. 1999. Holistic management, a new framework for decision making. Washington, D.C., USA: Island Press.
Savory, A. & Parsons, S.D. 1980. The Savory grazing method. Rangelands, 2: 234-237.
Scales, G.H., Streeter, C.L. & Denham, A.H. 1971. Nutritive value and consumption of range forage. Journal of Animal Science, 33: 310-312.
Seton, E.T. 1927. Lives of game animals. Vol 3. New York, New York, USA: Literary Guild of America.
Shaw, J.H. 1996. Bison. pp. 227-236, in: P.R. Krausmann (ed). Rangeland wildlife. Denver, Colorado, USA: Society for Range Management.
Sieg, C.H., Flather, C.H. & McCanny, S. 1999. Recent biodiversity patterns in the Great Plains: implications for restoration and management. Great Plains Research, 9: 277-313.
Sims, P.L. 1988. Grasslands. pp. 324-356, in: M.G. Barbour and W.D. Billings (eds). North American terrestrial vegetation. New York, New York, USA: Cambridge University Press.
Sims, P.L. & Singh, J.S. 1978a. The structure and function of ten western North American grasslands. II. Intra-seasonal dynamics in primary producer compartments. Journal of Ecology, 66: 547-572.
Sims, P.L. & Singh, J.S. 1978b. Intra-seasonal dynamics in primary producer compartments in ten western North American grasslands, III. Net primary production, turnover and efficiency of energy capture and water use. Journal of Ecology, 66: 573-597.
Sims, P.L., Singh, J.S. & Lauenroth, W.K. 1978. The structure and function of ten western North American grasslands. I. Abiotic and vegetational characteristics. Journal of Ecology, 66: 251-285.
Smeins, F. 1994. Little bluestem-Indiangrass-Texas winter grass - SRM 717. pp. 65-66, in: T.N. Shiflet (ed). Rangeland cover types of the United States. Denver, Colorado, USA: Society for Range Management.
Smeins, F.E. 2000. Ecology and management of Ashe juniper and redberry juniper. Abstracts of the Annual Meeting of the Society for Range Management. Boise, Idaho, USA, 2000. Denver, Colorado, USA: Society for Range Management.
Smith. E.F., Anderson, K.L., Owensby, C.E. & Hall, M.C. 1967. Different methods of managing bluestem pasture. Kansas State University Agricultural Experiment Station Bulletin, No. 507.
Smith, E.F. & Owensby, C.E. 1972. Effects of fire on true prairie grasslands. Proceedings of the Tall Timbers Ecology Conference, 12: 9-22.
Smith, E.F. & Owensby, C.E. 1978. Intensive-early stocking and season-long stocking of Kansas Flint Hills range. Journal of Range Management, 31: 14-17.
Smoliak, S. 1956. Influence of climatic conditions on forage production of shortgrass rangeland. Journal of Range Management, 9: 89-91.
Smoliak, S. 1968. Grazing studies on native range, crested wheatgrass and Russian wildrye pastures. Journal of Range Management, 21: 47-50.
Smoliak, S., Johnston, A., Kilcher, M.R. & Lodge, R.W. 1976. Management of prairie rangeland. Canada Department of Agriculture, Ottawa, Canada.
Smolik, J.D. 1974. Nematode studies at the Cottonwood Site. US/IBP Grassland Biome Technical Report, No. 251. Colorado State University, Fort Collins, Colorado, USA.
Soil Conservation Service. 1973. National soils handbook. Washington, D.C., USA: USDA.
Steuter, A.A. & Hidinger, L. 1999. Comparative ecology of bison and cattle on mixed-grass prairie. Great Plains Research, 9: 329-342.
Stoddart, L.A. & Smith, A.D. 1955. Range management. 2nd ed. New York, New York, USA: McGraw-Hill Book Co.
Stoddart, L.A., Smith, A.D. & Box, T.W. 1975. Range Management. 3rd ed. New York, New York, USA: McGraw-Hill Book Co.
Summer, C.A. & Linder, R.L. 1978. Food habits of the black -tailed prairie dog. Journal of Range Management, 31: 134-136.
Swanson, S. 1988. Riparian values as a focus for range management and vegetation science. pp. 425-445, in: P.T. Tueller (ed). Vegetation science applications for rangeland anslysis and management. Dordrecht, The Netherlands: Kluwer Academic.
Thomas, G.W., Herbel, C.H. & Miller, G.T. Jr. 1990. Rangeland resources. pp. 284- 290, in: G.T. Miller Jr. (ed). Resource conservation and management. Belmont, California, USA: Wadsworth Pub. Co.
Thurow, T.L., Blackburn, W.H. & Taylor, C.A. 1988. Infiltration and interrill erosion responses to selected livestock grazing strategies, Edwards Plateau, Texas. Journal of Range Management, 41: 296-302.
Tobey, R.C. 1981. Saving the prairies. The life cycle of the founding school of American Plant Ecology, 1895-1955. Berkeley, California, USA: University of California Press.
Transeau, E.N. 1935. The prairie peninsula. Ecology, 16: 423-437.
Trewartha, G. 1961. The earth’s problem climates. Madison, Wisconsin, USA: University of Wisconsin Press.
Tueller, P.T. 1989. Remote sensing technology for rangeland management applications. Journal of Range Management, 42(6): 442-453.
Vallentine, F.F. 1989. Range development and improvements. 3rd ed. San Diego, California, USA: Academic Press.
Vallentine, F.F. 1990. Grazing management. San Diego, California, USA: Academic Press.
Vanderhoff, J.L, Robel, R.J. & Kemp, K.E. 1994. Numbers and extent of black tailed prairie dogs in Kansas. Transactions of the Kansas Academy of Science, 97: 36-43.
Van Dyne, G.M. & Dyer, M.I. 1973. A general view of grasslands - a human and ecological perspective. pp. 38-57, in: Analysis of structure, function and utilization of grassland ecosystems. Proposal submitted to the National Science Foundation. Colorado State University, Fort Collins, Colorado, USA.
Van Dyne, G.M., Jameson, D.A. & French, N.R. 1970. Analysis of structure and function of grassland ecosystems. A progress report and a continuation proposal. Colorado State University, Fort Collins, Colorado, USA.
Van Poollen, H.W. & Lacey, J.R. 1979. Herbage response to grazing systems and stocking intensities. Journal of Range Management, 32: 250-253.
Wagnon, K.A., Albaugh, R. & Hart, G.H. 1960. Beef cattle production. New York, New York, USA: MacMillan.
Walker, J.W. 1995. Viewpoint: grazing management and research now and in the next millennium. Journal of Range Management, 48: 350-357.
Weakly. H.E. 1943. A tree ring record of precipitation in western Nebraska. Journal of Forestry, 41: 816-819.
Weaver, J.E. & Albertson, F.W. 1956. Grasslands of the Great Plains: their nature and use. Lincoln, Nebraska, USA: Johnson Publishing.
Weaver, J.E. & Albertson, F.W. 1940. Deterioration of grassland from stability to denudation with decrease in soil moisture. Botanical Gazette, 101: 598-624.
Weaver, J.E. & Albertson, F.W. 1939. Major changes in grassland as a result of continued drought. Botanical Gazette, 100: 576-591.
Weaver, J.E. & Albertson, F.W. 1936. Effects of the great drought on the prairies of Iowa, Nebraska and Kansas. Ecology, 17: 567-639.
Weltz, M. & Wood, M.K. 1986. Short-duration grazing in central New Mexico: effects on infiltration rates. Journal of Range Management, 39: 365-368.
Wester, D. & Bakken, T. 1992. Early allotments in South Dakota revisited. Rangelands, 14: 236-238.
White, M.R., Pieper, R.D., Donart, G.B. & White-Trifaro, L. 1991. Vegetation response to short-duration and continuous grazing in southcentral New Mexico. Journal of Range Management, 44: 399-404.
Whitman, W., Hanson, H.C. & Peterson, R. 1943. Relation of drought and grazing to North Dakota range lands. North Dakota Agricultural College Agricultural Experiment Station Bulletin, No. 320.
Wiens, J.A. 1973. Pattern and process in grassland bird communities. Ecological Monographs, 43: 237-270.
Wiens, J.A. 1974. Climatic instability and the "ecological saturation" of bird communities in North American grasslands. The Condor, 76: 385-400.
Wight, J.R. 1976. Range fertilization in the northern Great Plains. Journal of Range Management, 29: 180-185.
Wright. H.A. 1974. Range burning. Journal of Range Management, 27: 5-11.
Wright, H.A. 1978. Use of fire to manage grasslands of the Great Plains: Central and Southern Great Plains. pp. 694-696, in: D.N. Hyder (ed). Proceedings of the First International Rangeland Congress. Denver, Colorado, USA, 1978. Society for Range Management, Denver, Colorado, USA.
Wright, H.A. & Bailey, A.W. 1982. Fire ecology: United States and southern Canada. New York, New York, USA: John Wiley and Sons.
Yoakum, J.D., O’Gara, B.W. & Howard, V.W. Jr. 1996. Pronghorn on western rangelands. pp. 211-226, in: P.R. Krausmann (ed). Rangeland wildlife. Denver, Colorado, USA: Society for Range Management.
![]()
![]()
![]()
){kind=link}
){kind=link}