![]()
![]()
![]()
Oscar H. Padin
Instituto de Limnología “Dr. Raúl A. Ringuelet”
Berisso, Provincia de Buenos Aires, Argentína
8.1 RESUMEN
El presente trabajo constituye un ensayo de evaluación del impacto de Phalacrocorax olivaceus, Podiceps major y Sterna Trudeaus sobre la mortalidad de las poblaciones de peces de la laguna Chascomús (35° 34' S, 58° 00' W. Provincia de Buenos Aires, Argentina), a partir de la numerosidad de dichas aves y mediante la aplicación de un modelo energético general.
El consumo de energía anual estimado para las tres especies consideradas suma 20.451.191 kcal, las que constituyen 21.873 kg de pescado. Esta última cifra representa 7,26 kg por hectárea extraídos anualmente de la laguna, correspondiendo a P. olivaceus el 87,15%, a P. major el 12,62% y a S. trudeaui el 0,23% del total. El consumo de pescado referido representa sólo el 0,97% de la única cifra publicada de producción de peces en lagunas pampásicas, y el 37,34% del promedio anual pesquero comercial correspondiente al período 1958–1972, en esta laguna.
ABSTRACT
The present paper represents an attempt to evaluate the impact of Phalacrocorax olivaceus, Podiceps major and Sterna trudeaui on the mortality of fish populations of Chascomús pond (35°34'S; 58°00'W., Buenos Aires Province, Argentina), performed on the basis of the densities of these birds and with the aid of a generalized energetic model. The estimated annual energy consumption of the three species considered amounts to 20,451,191 kcal, which represent 21,873 kg of fish. The latter figure implies the extraction from the pond of 7.26 kg of fish per ha annually, 87.15% of which corresponds to P. olivaceus, 12.62% to P. major, and only 0.23% to S. trudeaui. This biomass represents only 0.97% of the only available estimate of fish production in a Pampasic pond, and 37.34% of the average (1958–1972) annual commercial fishing yield for this system.
8.2 INTRODUCCION
El estudio de la ecología energética de las poblaciones de aves en la Argentina ha experimentado un notable incremento en la última década, particularmente en lo referente a la diversidad de la dieta y relaciones tróficas. Sin embargo, estos trabajos por sí sólos no permiten cuantificar el impacto de las aves sobre las poblaciones de sus presas. Para ello es necesario conocer la numerosidad de la especie dentro de su área de actividad y la tasa de renovación del alimento o dieta diaria individual.
El valor calórico de la dieta puede ser estimado indirectamente por medio de ecuaciones derivadas de estudios químicos, calorimétricos y respirométricos, siendo estos últimos los más difundidos por su sencillez y precisión.
El presente trabajo constituye un ensayo de evaluación del impacto de las aves ictiófagas sobre la mortalidad de las poblaciones de peces de una laguna pampásica, a partir de la numerosidad de dichas aves y mediante la aplicación de un modelo de consumo energético deducido experimentalmente por otros autores para aves no-paseriformes. Sus resultados permitirán discriminar uno de los componentes de la mortalidad natural de peces y complementar otras investigaciones sobre vehiculización de materia y energía en el ecosistema lagunar, en relación con el manejo del recurso pesquero.
8.3 MATERIALES Y METODOS
Para el presente trabajo se tomaron como base las estimaciones de numerosidad realizadas durante el año 1982 en la laguna Chascomús por Daciuk et al. (1985), para tres especies de régimen esencialmente ictiófago. Estas son: el biguá (Phalacrocorax olivaceus olivaceus), el macá grande (Podiceps major) y el viotín corona blanca (Sterna trudeaui), cuya alimentación ha sido estudiada en distintos limnotopos por Bo (1956), Mogilner y Aramburu (1969), Beltzer y Oliveros (1982), Beltzer (1983) y Oliveros y Beltzer (1983).
La laguna de Chascomús se encuentra ubicada al NE de la Provincia de Buenos Aires (35° 34' S., 58° 00' W.), formando parte de la cuenca del río Salado (Figura 1). Zoogeográficamente pertence al Dominio Pampásico, definido por Ringuelet (1961) y es considerada por su ubicación y características como un área piloto para el desarrollo de investigaciones limnológicas de interés regional.
Los cálculos de dispendio energético se realizaron utilizando las ecuaciones propuestas por Kendeigh et al. (1977) para el metabolismo total o presupuesto energético diario (Daily energy budget, DEB)
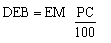
donde: EM es el metabolismo de existencia (existence metabolism), determinado experimentalmente, y PC (percentage) constituye un porcentaje del metabolismo de existencia relacionado con la actividad libre del ave. El EM es la tasa de consumo de energía correspondiente a aves enjauladas, manteniendo peso constante y fuera del período reproductivo o migratorio.
Si reemplazamos en (1) por las constantes respectivas tendremos:
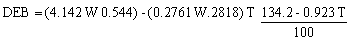
donde: T es la temperatura ambiente media mensual en °C y W es el peso fresco del ave en gramos; en este caso se emplearon los pesos promedios de aves capturadas durante los censos (Cuadro 1).
El valor de peso utilizado para P. olivaceus fue igual al obtenido por Marelli (1945) para machos, muy similar al representado por Oliveros y Beltzer (1983) y algo más alto que el citado por Marelli op. cit. para hembras y por Contreras y Fernández (1980). Los pesos correspondientes a P. major y S. trudeaui no pudieron ser comparados con informacion de otros autores.
Para estimar la biomasa de peces consumidos por las aves, a partir de los valores de DEB calculados, se utilizaron los índices discutidos por Morales et al. (1981). A partir de un análisis bibliográfico, estos autores proponen una eficiencia o digestibilidad del 85% en el caso de aves ictiófagas y un valor calórico de pescado referido a peso fresco de 1,1 kcal por gramo.
Las estimaciones precendentes se comparan con las obtenidas mediante la aplicación de las ecuaciones discutidas por Lasiewski y Dawson (1967). Las mismas corresponden a los requerimientos energéticos de aves en reposo; ayunando y dentro de la zona de termoneutralidad, definida como tasa metabólica estandar (SM) por Kendeight et al. (1977).
8.4 RESULTADOS
Los valores de consumo de pescado calculados a partir de las densidades de aves y la temperatura ambiente media mensual se presentan en el Cuadro 2 para las tres especies estudiadas. La población de P.olivaceus en la laguna Chascomús consume anualmente 19.062 kg de pescado, con un máximo en el mes de marzo (6.346 kg) y un mínimo en junio (23 kg). La dieta diaria individual oscila entre 201 g en diciembre y 258 g en junio.
P. major consume anualmente 2.671 kg de pescado, con un máximo en el mes de julio (1.112 kg) y un mínimo en agosto (28 kg). La dieta diaria individual oscila entre 178 g en diciembre y 230 g en junio. S. trudeaui consumió durante los meses en que fue detectado en la laguna, 50 kg de pescado, con un máximo en el mes de octubre (12 kg) y un mínimo en abril y diciembre (5 kg). La dieta diaria individual fue estimada entre 49 g en diciembre y 63 g en octubre.
Sumando los valores para las tres especies se obtiene un total de 20.451.191 kcal, las que constituyen 21.873 kg de pescado. Esta cifra representa 7,26 kg por hectárea extraídos anualmente de la laguna por estas aves, correspondiendo a P. olivaceus el 87,15%, a P. major el 12,62 y a S. trudeaui el 0,23% del total.
8.5 DISCUSION
Dado que las estimaciones se basan en información energética y de eficiencia alimentaria derivada del estudio de otras especies, debemos hacer algunas consideraciones al respecto. Varios autores (Zar, 1968, 1970; Lasiewski y Dawson, 1969; Lasiewski y Calder, 1971) han discutido las ecuaciones que relacionan el metabolismo estándar con el peso del cuerpo, analizando la conveniencia y posibles errores surgidos de su generalización. La natación, el buceo y el vuelo implican un aumento sustancial en el consumo de energía (Tucker, 1970, 1971; Prange y Schmidt Nielsen, 1970). La temperatura ambiente, el alejarse del rango de termoneutralidad, actúa con un efecto similar (Schmidt Nielsen, 1976).
Es por ello que la ecuación elegida para las estimaciones del dispendio energético debe considerar estos factores, como queda en evidencia si comparamos los valores del metabolismo estándar obtenidos mediante las ecuaciones discutidas por Lasiewski y Dawson (1969). Ellos oscilan entre el 50 y el 58% del valor obtenido por nosotros para el metabolismo total (DEB), introduciendo -de ser elegidos para los cálculos-una subestimación importante del impacto de las aves sobre las poblaciones de sus presas.
Los valores de la dieta diaria individual obtenidos por P. olivaceus y P. major están cerca del límite inferior del rango indicado por Margalef (1983) para aves ictiófagas medianas y grandes. El valor obtenido para S. trudeaui está bastante por debajo de este límite inferior. En el caso particular de P. olivaceus, la dieta estimada se sitúa cercana al valor de peso máximo del contenido estomacal encontrado por Oliveros y Beltzer (1983).
El consumo de pescado calculado (7,26 kg/ha/año), constituye sólo el 0,97% de la producción anual de pejerrey Basilichthys bonariensis bonariensis, calculada por Freyre (1976) para la laguna de Lobos, aunque este último valor de producción, el único publicado para lagunas pampásicas, parece excepcionalmente alto.
En relación con la pesca artesanal en la laguna Chascomús, considerando el promedio anual obtenido de las estadísticas pesqueras para el período 1958–1972, el consumo de peces por las aves ictiófagas estudiadas representa el 37,34% de este valor. Estos primeros resultados plantean la necesidad de extender las observaciones de la numerosidad de aves a otras lagunas, así como obtener información sobre la producción y sus fluctuaciones multianuales en dichos ambientes.
8.6 AGRADECIMIENTOS
El autor desea expresar su agradecimiento al Dr. D. Boltovsky por las valiosas sugerencias aportadas al primer manuscrito; al Prof. A. Beltzer, quien facilitó su bibliografía; y, finalmente, a L. Protogino, A. Miquelarena, J. Iwaszkiw y H. López, por su apoyo constante.
8.7 BIBLIOGRAFIA
BELTZER, A.H. y Oliveros, O. 1982. Alimentación del macá grande (Podiceps major) en el valle aluvial del río Paraná medio (Podicipediformes: Podicipedidae). EN: Rev. Asoc. Cienc. Nat. Litoral. 13:5–10.
BELTZER, A.H. 1983. Nota sobre fidelidad y participación trófica del “bigua común” (Phalacrocorax olivaceus) en ambientes del río Paraná medio (Pelecaniformes: Phalacrocoracidae). EN: Rev. Asoc. Cienc. Nat. Litoral. 14(2):111–114.
BO, N.A. 1956. Observaciones morfológicas y etológicas sobre el biguá. EN: El Hornero. 10(2):147–157.
CONTRERAS, J.R. y FERNANDEZ, A. 1980. Ecología de la avifauna de la laguna del Vivorón, Departamento de Maipú, Provincia de Mendoza. EN: Revista del Museo de San Rafael. 8(1):3–14.
DACIUK, J., PADIN, O.H. y PROTOGINO, L.C. 1985. Censos de la avifauna de la laguna Chascomús (Prov. de Buenos Aires, Argentina). EN: Physis. Buenos Aires, Argentina. B. 43(105:93–102).
FREYRE, L.R. 1976. La población del pejerrey de la laguna de Lobos. EN: Limnobios. 1(4):105–128.
KENDEIGH, S.; DOL NICK, J. y GAVRILOV, M. 1977. Avian energetics. EN: Granivorous birds in ecosystems. IN: Pinowski, J. y Kendeigh, Eds. Cambridge, England, Cambridge Univ. Press.
LASIEWSKI, R.C. y DAWSON, W.R. 1967. A re-examination of relation between standard metabolic rate and body weigh in birds. IN: Condor 69:13–23.
LASIEWSKI, R.C. 1969. Calculation and miscalculation of the ecuations relating avian standard metabolism to body weight in birds. IN: Condor. 69:335–336.
LASIEWSKI, R.C. y CALDER, W.A. 1971. A preliminary allometric analysis of respiratory variables in resting birds. IN: Respiration Phisiology. 11:152–166.
MARELLI, C.A. 1945. Investigación sobre el biguá y la destrucción del pejerrey en la laguna Alsina. EN: Suelo Argentino. 4(38):59–64.
MARGALEF, R. 1983. Limnología. Barcelona, España, Editorial Omega. 1010 p.
MOGILNER, R. y ARAMBURU, R.H. 1969. Relaciones alimentarias de las aves acuáticas en la laguna Chascomús. Buenos Aires, Dirección Rec. Pesq., Convenio Estudio Riqueza Ictícola. Tomo 4:17–52.
MORALES, G., PINOWSKI, J., MADRIZ, M. y GOMEZ, F. 1981. Densidades poblacionales, flujo de energía y hábitos alimentarios de las aves ictiófagas de los módulos de Apure. EN: Acta Biol. Venez. Venezuela. 11(2):1–45.
OLIVEROS, O. y BELTZER, A.H. 1983. Alimentación del biguá común (Phalacrocorax olivaceus) en el valle aluvial del río Paraná medio (Pelacaniformes: Phalacrocoracidae). EN: Neotropica. 29(82):225–230.
PRANGE, H. y SCHMIDT-NIELSEN, K. 1970. The metabolic cost of swimming in ducks. IN: J. Exp. Biol. 53:763–777.
RINGUELET, R.A. 1961. Rasgos fundamentales de la zoogeografía de la Argentina. EN: Physis. Buenos Aires, Argentina. 63:151–170.
SCHMIDT-NIELSEN, K. 1976. EN: Fisiología animal. Barcelona, España, Ed. Omega. 499 p.
TUCKER, V.A. 1970. Energetic cost of locomotion in animals. IN: Comp. Biochem. Physiol. 34:841–846.
TUCKER, V.A. 1971. Flight energetics in birds. IN: Am. Zoologist. 11:115–124.
ZAR, J.H. 1968. Standard metabolism comparisons between orders of birds. IN: Condor. 70:278.
ZAR, J.H. 1970. On the fitting of ecuations relating avian standard metabolism to body weight. IN: Condor. 72(2):247.
Cuadro 1. peso en gramos del material considerado.
| ESPECIE | № | X | m | M | S | X+- |
| Phalacrocorax o.olivaceus | 21 | 1405.0 | 1020 | 2220 | 269.8 | 122.8 |
| Podiceps major | 19 | 1151.7 | 860 | 1610 | 186.3 | 89.8 |
| Sterna trudeaui | 9 | 166.7 | 50 | 205 | 18.0 | 13.9 |
CUADRO II
VALORES OBTENIDOS A PARTIR DE LAS TEMPERATURAS MEDIAS MENSUALES (T) Y LA NUMEROSIDAD MENSUAL DE AVES (N)
| MES | T | N | DEB (Kcal) | DDI (g) | PC (Kg) | ||||||||
| (°C) | P.o. | P.m. | S.t. | P.o. | P.m. | S.t. | P.o. | P.m. | S.t. | P.o. | P.m. | S.t. | |
| Enero | 22.2 | 541 | 19 | 6 | 189.12 | 167.16 | 46.69 | 202 | 179 | 50 | 3392 | 105 | 9 |
| Febrero | 20.4 | 937 | 6 | 7 | 196.30 | 173.78 | 49.80 | 210 | 186 | 53 | 5508 | 31 | 10 |
| Marzo | 20.6 | -979 | 33 | - | 195.50 | 173.03 | - | 209 | 185 | - | 6346 | 189 | - |
| Abril | 17.4 | 199 | 22 | 3 | 208.56 | 185.08 | 55.13 | 223 | 198 | 59 | 1332 | 131 | 5 |
| Mayo | 14.7 | 31 | 32 | - | 219.89 | 195.54 | - | 235 | 209 | - | 226 | 208 | - |
| Junio | 9.9 | 3 | 45 | - | 240.75 | 214.80 | - | 258 | 230 | - | 23 | 310 | - |
| Julio | 10.2 | 7 | 157 | - | 239.42 | 213.57 | - | 256 | 228 | - | 56 | 1112 | - |
| Agosto | 11.1 | 40 | 4 | - | 235.45 | 209.91 | - | 252 | 225 | - | 312 | 28 | - |
| Septiembre | 14.3 | 52 | 24 | - | 221.60 | 197.11 | - | 237 | 211 | - | 370 | 152 | - |
| Octubre | 15.5 | 63 | 43 | 6 | 216.50 | 192.41 | 58.51 | 232 | 206 | 63 | 452 | 274 | 12 |
| Noviembre | 17.3 | 75 | 27 | 5 | 208.97 | 185.46 | 55.31 | 224 | 198 | 59 | 503 | 161 | 9 |
| Diciembre | 22.5 | 87 | 11 | 3 | 187.94 | 166.06 | 46.18 | 201 | 178 | 49 | 542 | 61 | 5 |
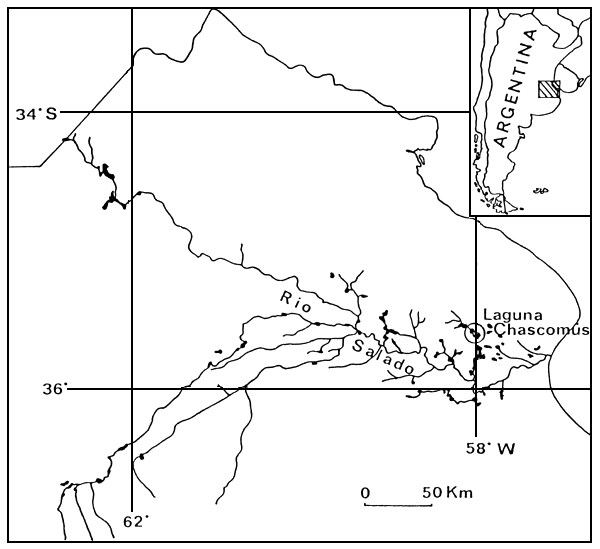
Figura 1. Laguna de Chascomús.
![]()
![]()
![]()