![]()
![]()
![]()
4.3.1. Harvesting and distribution
4.3.2. Cold storage
4.3.3. Nutritional quality
4.3.4. Enrichment with nutrients
4.3.5. Enrichment for disease control
4.3.6. Applications of Artemia for feeding different species
4.3.7. Literature of interest
4.3.8. Worksheets
Greet Merchie
Laboratory of Aquaculture & Artemia Reference
Center
University of Gent, Belgium
After hatching and prior to feeding the nauplii to fish/crustacean larvae, they should be separated from the hatching wastes (empty cyst shells, unhatched cysts, debris, microorganisms and hatching metabolites). Five to ten minutes after switching off the aeration, cyst shells will float and can be removed from the surface, while nauplii and unhatched cysts will concentrate at the bottom (Fig. 4.3.1.).
Figure 4.3.1. Hatching container at harvest.
Since nauplii are positively phototactic, their concentration can be improved by shading the upper part of the hatching tank (use of cover) and focusing a light source on the transparent conical part of the bottom. Nauplii should not be allowed to settle for too long (i.e., maximum 5 to 10 min.) in the point of the conical container, to prevent dying off due to oxygen depletion. Firstly, unhatched cysts and other debris that have accumulated underneath the nauplii are siphoned or drained off when necessary (i.e. when using cysts of a lower hatching quality). Then the nauplii are collected on a filter using a fine mesh screen (< 150 µm), which should be submerged all the time so as to prevent physical damage of the nauplii. They are then rinsed thoroughly with water in order to remove possible contaminants and hatching metabolites like glycerol. Installation of automated systems simplify production techniques in commercial operations, (i.e. by the use of a concentrator/rinser; Fig 4.3.2.) that enables fast harvesting of large volumes of Artemia nauplii and allows complete removal of debris from the hatching medium. This technique results in a significant reduction of labour and production costs.
Figure 4.3.2. Concentrator/rinser in use (Photo from Sorgeloos and Léger, 1992).
As the live food is suspected to be a source of bacterial infections eventually causing disease problems in larval rearing, microbial contamination should be kept to a minimum. During the hatching of Artemia cysts, bacterial numbers increase by 103 to 105 compared to the initial population before the breaking of the cysts. This bacterial population remains well established and cannot be removed from the nauplii by rinsing with seawater or freshwater; rinsing only having a diluting effect on the water surrounding the nauplii. However, hatching nauplii from cysts that have been submitted to a disinfection procedure successfully reduces the bacterial numbers after harvesting compared to standard hatching techniques using non-disinfected cysts (Fig. 4.3.3.); in particular Vibrio levels are reduced below 103 CFU.g-1. At the moment of writing a new disinfected cyst product has become commercially available (namely DC-cysts, INVE Aquaculture NV, Belgium) which has proved to result in low bacterial numbers after hatching.
Since instar I nauplii completely thrive on their energy reserves they should be harvested and fed to the fish or crustacean larvae in their most energetic form, (i.e. as soon as possible after hatching). For a long time farmers have overlooked the fact that an Artemia nauplius in its first stage of development can not take up food and thus consumes its own energy reserves. At the high temperatures applied for cyst incubation, the freshly-hatched Artemia nauplii develop into the second larval stage within a matter of hours. It is important to feed first-instar nauplii to the predator rather than starved second-instar meta-nauplii which have already consumed 25 to 30% of their energy reserves within 24 h after hatching (Fig. 4.3.4.). Moreover, instar II Artemia are less visible as they are transparent, are larger and swim faster than first instar larvae, and as a result consequently are less accessible as a prey. Furthermore they contain lower amounts of free amino acids, and their lower individual organic dry weight and energy content will reduce the energy uptake by the predator per hunting effort. All this may be reflected in a reduced growth of the larvae, and an increased Artemia cyst bill as about 20 to 30% more cysts will be needed to be hatched to feed the same weight of starved meta-nauplii to the predator (Léger et al., 1986). On the other hand, instar II stages may be more susceptible to digestive enzyme breakdown in the gut of the predator since these enzymes can also penetrate the digestive tract of the Artemia through the opened mouth or anus.
Molting of the Artemia nauplii to the second instar stage may be avoided and their energy metabolism greatly reduced (Fig. 4.3.4.) by storage of the freshly-hatched nauplii at a temperature below 10°C in densities of up to 8 million per liter. Only a slight aeration is needed in order to prevent the nauplii from accumulating at the bottom of the tank where they would suffocate. In this way nauplii can be stored for periods up to more than 24 h without significant mortalities and a reduction of energy of less than 5%. Applying 24-h cold storage using styrofoam insulated tanks and blue ice packs or ice packed in closed plastic bags for cooling, commercial hatcheries are able to economize their Artemia cyst hatching efforts (i.e., reduction of the number of hatchings and harvests daily, fewer tanks, bigger volumes). Furthermore, cold storage allows the farmer to consider more frequent and even automated food distributions of an optimal live food. This appeared to be beneficial for fish and shrimp larvae as food retention times in the larviculture tanks can be reduced and hence growth of the Artemia in the culture tank can be minimized. For example, applying one or maximum two feedings per day, shrimp farmers often experienced juvenile Artemia in their larviculture tanks competing with the shrimp postlarvae for the algae. With poor hunters such as the larvae of turbot Scophthalmus maximus and tiger shrimp Penaeus monodon, feeding cold-stored, less active Artemia furthermore results in much more efficient food uptake.
The nutritional effectiveness of a food organism is in the first place determined by its ingestibility and, as a consequence by its size and form. Naupliar size, varying greatly from one geographical source of Artemia to another, is often not critical for crustacean larvae, which can capture and tear apart food particles with their feeding appendages. For marine fish larvae that have a very small mouth and swallow their prey in one bite the size of the nauplii is particularly critical. For example, fish larvae that are offered oversized Artemia nauplii may starve because they cannot ingest the prey. For at least one species, the marine silverside Menidia menidia, a high correlation exists between the naupliar length of Artemia and larval fish mortality during the five days after hatching: with the largest strains of Artemia used (520 µm nauplius length), up to 50% of the fish could not ingest their prey and starved to death whereas feeding of small Artemia (430 µm) resulted in only 10% mortality (Fig. 4.3.5.). Fish that produce small eggs, such as gilthead seabream, turbot and grouper must be fed rotifers as a first food because the nauplii from any Artemia strain are too large. In these cases, the size of nauplii (of a selected strain) will determine when the fish can be switched from a rotifer to an Artemia diet. As long as prey size does not interfere with the ingestion mechanism of the predator, the use of larger nauplii (with a higher individual energy content) will be beneficial since the predator will spend less energy in taking up a smaller number of larger nauplii to fulfill its energetic requirements. Data on biometrics of nauplii from various Artemia strains are presented in Table 4.1.2. (see chapter 4.1.) and ranges given in Fig 4.3.6.
Figure 4.3.5. Correlation of mortality rate of Menidia menidia larvae and nauplii length of Artemia from seven geographical sources offered as food to fish larvae (modified from Beck and Bengtson, 1982).

Figure 4.3.6. Schematic diagram of the biometrical variation in freshly-hatched instar I Artemia nauplii from different geographical origin (size =nauplius length; volume index = CoulterCounter)

Another important dietary characteristic of Artemia nauplii was identified in the late 1970s and early 1980s, when many fish and shrimp hatcheries scaled up their production and reported unexpected problems when switching from one source of Artemia to another. Japanese, American and European researchers studied these problems and soon confirmed variations in nutritional value when using different geographical sources of Artemia for fish and shrimp species. The situation became more critical when very significant differences in production yields were obtained with distinct batches of the same geographical origin of Artemia.
Studies in Japan and the multidisciplinary International Study on Artemia revealed that the concentration of the essential fatty acid (EFA) 20:5n-3 eicosapentaenoic acid (EPA) in Artemia nauplii was determining its nutritional value for larvae of various marine fishes and crustaceans (Léger et al., 1986). Various results were obtained with different batches of the same geographical Artemia source, containing different amounts of EPA and yielding proportional results in growth and survival of Mysidopsis bahia shrimps fed these Artemia. Levels of this EFA vary tremendously from strain to strain and even from batch to batch (Table 4.3.1.), the causative factor being the fluctuations in biochemical composition of the primary producers available to the adult population. Following these observations, appropriate techniques have been developed for improving the lipid profile of deficient Artemia strains (see further). Commercial provisions of Artemia cysts containing high EPA levels are limited and consequently, these cysts are very expensive. Therefore, the use of the high-EPA cysts should be restricted to the feeding period when feeding of freshly-hatched nauplii of a small size is required.
In contrast to fatty acids, the amino acid composition of Artemia nauplii seems to be remarkably similar from strain to strain, suggesting that it is not environmentally determinedi n the manner that the fatty acids are.
Table 4.3.1. Intra-strain variability of 20:5n-3 (EPA) content in Artemia. Values represent the range (area percent) and coefficient of variation of data as compiled by Léger et al. (1986).
|
Cyst source |
20:5n-3 range |
Coefficient of variation |
|
San Francisco Bay, CA-USA |
0.3-13.3 |
78.6 |
|
Great Salt Lake (South arm), UT-USA |
2.7-3.6 |
11.8 |
|
Great Salt Lake (North arm), UT-USA |
0.3-0.4 |
21.2 |
|
Chaplin Lake, Canada |
5.2-9.5 |
18.3 |
|
Macau, Brazil |
3.5-10.6 |
43.2 |
|
Bohai Bay, PR China |
1.3-15.4 |
50.5 |
The presence of several proteolytic enzymes in developing Artemia embryos and Artemia nauplii has led to the speculation that these exogenous enzymes play a significant role in the breakdown of the Artemia nauplii in the digestive tract of the predator larvae. This has become an important question in view of the relatively low levels of digestive enzymes in many first-feeding larvae and the inferiority of prepared feeds versus live prey.
Table 4.3.2. Amino acid composition of Artemia nauplii (mg.g-1 protein) (modified from Seidel et al., 1980).
|
|
Macau, Brazil |
Great Salt Lake, UT-USA |
San Pablo Bay, CA-USA |
|
aspartic acid |
110 |
113 |
141 |
|
threonine |
52 |
48 |
60 |
|
serine |
45 |
54 |
77 |
|
glutamic acid |
131 |
135 |
102 |
|
proline |
57 |
59 |
49 |
|
glycine |
60 |
60 |
74 |
|
alanine |
46 |
49 |
42 |
|
valine |
53 |
52 |
55 |
|
methionine |
22 |
37 |
26 |
|
isoleucine |
56 |
68 |
54 |
|
leucine |
89 |
100 |
84 |
|
tyrosine |
105 |
66 |
77 |
|
phenylalanine |
51 |
85 |
104 |
|
histidine |
49 |
27 |
35 |
|
lysine |
117 |
93 |
87 |
|
arginine |
115 |
97 |
98 |
Artemia cysts (San Francisco Bay) were analysed for the content of various vitamins and were found to contain high levels of thiamin (7-13 µg.g-1), niacin (68-108 µg.g-1), riboflavin (15-23 µg.g-1), pantothenic acid (56-72 µg.g-1) and retinol (10-48 µg.g-1). A stable form of vitamin C (ascorbic acid 2-sulphate) is present in Artemia cysts. This derivative is hydrolysed to free ascorbic acid during hatching, the -ascorbic acid levels in Artemia nauplii varying from 300 to 550 µg g-1 DW. The published data would appear to indicate that the levels of vitamins in Artemia are sufficient to fulfill the dietary requirements recommended for growing fish. However, vitamin requirements during larviculture, are still largely unknown, and might be higher due to the higher growth and metabolic rate of fish and crustacean larvae.
As mentioned previously, an important factor affecting the nutritional value of Artemia as a food source for marine larval organisms is the content of essential fatty acids, eicosapentaenoic acid (EPA: 20:5n-3) and even more importantly docosahexaenoic acid (DHA: 22:6n-3). In contrast to freshwater species, most marine organisms do not have the capacity to biosynthesize these EFA from lower chain unsaturated fatty acids, such as linolenic acid (18:3n-3). In view of the fatty acid deficiency of Artemia, research has been conducted to improve its lipid composition by prefeeding with (n-3) highly unsaturated fatty acid (HUFA)-rich diets. It is fortunate in this respect that Artemia, because of its primitive feeding characteristics, allows a very convenient way to manipulate its biochemical composition. Thus, since Artemia on molting to the second larval stage (i.e. about 8 h following hatching), is non-selective in taking up particulate matter, simple methods have been developed to incorporate lipid products into the brine shrimp nauplii prior to offering them as a prey to the predator larvae. This method of bioencapsulation, also called Artemia enrichment or boosting (Fig. 4.3.7.), is widely applied at marine fish and crustacean hatcheries all over the world for enhancing the nutritional value of Artemia with essential fatty acids.
Figure 4.3.7. Schematic diagram of the use of Artemia as vector for transfer of specific components into the cultured larvae.

British, Japanese, French and Belgian researchers have also developed other enrichment products, including unicellular algae, w-yeast and/or emulsified pre-parations, compound diets, micro-particulate diets or self-emulsifying concentrates. Apart from the enrichment diet used, the different techniques vary with respect to hatching conditions, pre-enrichment time (time between hatching and addition of enrichment diet), enrichment period, and temperature. Highest enrichment levels are obtained when using emulsified concentrates (Fig. 4.3.8., Table 4.3.3.).
Figure 4.3.8. HUFA-levels in Great Salt Lake (Utah, USA) Artemia (meta-) nauplii enriched with Super Selco® (INVE Aquaculture NV, Belgium) (modified from Dhont et al., 1993).

Table 4.3.3. Enrichment levels (mg.g-1 DW) in Artemia nauplii boosted with various products
|
|
DHA |
EPA |
(n-3) HUFA |
|
Super Selco (INVE Aquaculture NV) |
14.0 |
28.6 |
50.3 |
|
DHA Selco (INVE Aquaculture NV) |
17.7 |
10.8 |
32.7 |
|
Superartemia (Catvis) |
9.7 |
13.2 |
26.3 |
|
SuperHUFA (Salt Creek) |
16.4 |
21.0 |
41.1 |
Figure 4.3.9. HUFA levels in 24-h Super Selco®-enriched Artemia metanauplii during storage at 10 and 25°C (modified from Dhont et al., 1993).

In view of the importance of DHA in marine fish species a great deal of effort has been made to incorporate high DHA/EPA ratios in live food. To date, the best results have been obtained with enrichment emulsions fortified with DHA (containing a DHA/EPA ratio up to 7), yielding Artemia meta-nauplii that contain 33 mg DHA.g-1 DW. Compared to enrichment with traditional products, a maximum DHA/EPA ratio of 2 instead of 0.75 can be reached using standard enrichment practices.
The reason for not attaining the same ratio is the inherent catabolism of DHA upon enrichment within the most commonly used Artemia species (i.e. A. franciscana). The capability of some Chinese Artemia strains to reach high DHA levels during enrichment and to maintain their levels during subsequent starvation might open new perspectives to provide higher dietary DHA levels and DHA/EPA ratios to fish and crustacean larvae.
Apart from EFA, other nutrients such as vitamins and pigments can be incorporated in Artemia. Fat soluble vitamins (especially vitamin A and vitamin E) were reported to accumulate in Artemia over a short-term (9 h) enrichment period with vitamin A levels increasing from below 1 IU.g-1 (WW basis) to over 16 IU.g-1 and vitamin E levels increasing from below 20 µg.g-1 to about 250 µg.g-1. Recently tests have also been conducted to incorporate ascorbic acid into live food. Using the standard enrichment procedure and experimental self-emulsifying concentrates containing 10, 20 and 30% (on a DW basis) of ascorbyl palmitate (AP) in addition to the triglycerides, high levels of free ascorbic acid (AA) can be incorporated into brine shrimp nauplii (Fig. 4.3.11.). For example, a 10%-AP inclusion in the emulsion enhances AA levels within freshly-hatched nauplii by 50% from natural levels (500 µg g-1 DW). By contrast, however, a 20 or 30% addition increases AA levels in Artemia 3-fold and 6-fold respectively after 24 h enrichment at 27°C; with (n-3) HUFA levels remaining equal compared to normal enrichment procedures. Moreover, these AA concentrations do not decrease when the enriched nauplii are stored for 24 h in seawater (Fig. 4.3.11.).
Figure 4.3.11. Ascorbic acid enrichment in Artemia nauplii.

Table 4.3.4. Variability in DHA, EPA and total (n-3) HUFA levels in enriched Artemia nauplii sampled in the laboratory (A) using a standard procedure and in three sea bream hatcheries (B) according to the in-house method (mean and sd) (modified from Lavens et al., 1995)
|
|
DHA |
EPA |
(n-3) HUFA |
|||
|
area % |
mg g-1 |
area % |
mg g-1 |
area % |
mg g-1 |
|
|
A: |
|
|
|
|
|
|
|
applied by the same person (n=10) |
||||||
|
|
7.1 ± 2.5 |
12.5 ± 6.5 |
13.8 ± 2.2 |
24.2 ± 5.7 |
23.5 ± 4.5 |
41.9 ± 13.1 |
|
applied by different people over a 2-month period (n=5) |
||||||
|
|
6.2 ± 0.9 |
11.3 ± 2.6 |
14.5 ± 4.1 |
27.0 ± 9.9 |
23.3 ± 5.1 |
43.0±12.9 |
|
applied by different people over a 2-year period (n=13) |
||||||
|
|
7.8 ± 2.2 |
17.0 ± 5.8 |
16.7 ± 2.3 |
35.7 ± 7.6 |
26.7 ± 4.8 |
57.4 ± 14.2 |
|
B: |
|
|
|
|
|
|
|
1 (n=2) |
3.8 ± 2.5 |
8.1 ± 6.3 |
9.9 ± 4.0 |
20.3 ± 11.2 |
16.1 ± 7.1 |
33.2 ± 19.7 |
|
2 (n=3) |
5.9 ± 2.4 |
8.1 ± 1.4 |
10.5 ± 1.1 |
15.9 ± 5.4 |
20.0 ± 5.8 |
28.5 ± 6.4 |
|
3 (n=3) |
6.1 ± 0.6 |
12.6 ± 1.5 |
14.2 ± 0.8 |
29.1 ± 2.3 |
12.6 ± 1.5 |
46.6 ± 4.0 |
The incidence of microbial diseases has increased dramatically along with the degree of intensification in the larval production of aquaculture species. Treating microbial infections in fish and shrimp larvae is most often carried out by dissolving relatively high doses of broad spectrum antibiotics in the culture water. A major disadvantage of this method is that large amounts of expensive drugs are used and subsequently discharged into the environment, and thereby placing the animal and human health at risk. However, a direct treatment through the food chain (i.e. through oral administration) using much smaller quantities has proven to be more effective and safer for the environment. In this respect the possibility of loading Artemia nauplii with doses of up to 300 µg.g-1 DW of the therapeutic mixture Trimetoprim: Sulfamethoxazole (1:5), using self-emulsifying concentrates containing 10% of the mixture, has been demonstrated (Table 4.3.5.). This bioencapsulation technique eventually yielded levels up to 20 µg.g-1 antibiotics within European sea bass larvae 3 h after feeding one dosage of antibiotic-enriched Artemia meta-nauplii (Fig. 4.3.12.). In turbot larvae even higher tissue levels have been obtained, with a maximum tissue concentration of 90 µg antibiotics.g-1 was reached 4 h post feeding. Prophylactic and therapeutic efficiency was tested by feeding medicated Artemia respectively prior to and after an oral challenge with a pathogenic Vibrio anguillarum strain. In both cases mortality was significantly reduced in the treated turbot compared to the untreated controls. Of course, enrichment levels as well as therapeutic efficiency will depend on the antibiotics used. In fact, the same enrichment procedure can also be used to incorporate and transfer vaccines to fish larvae, and by so doing facilitating oral vaccination.
Table 4.3.5. Accumulation of trimetoprim (TMP) and sulfamethoxazole (SMX) in Artemia nauplii after 24 h enrichment using an enrichment emulsion containing TMP:SMX (1:5).
|
|
ng.mg-1 protein |
ng.mg-1 dry weight |
|
TMP |
212.1 |
77.8 |
|
SMX |
579.3 |
212.4 |
|
TMP + SMX |
791.4 |
291.1 |
4.3.6.1. Penaeid shrimp
4.3.6.2. Freshwater prawn
4.3.6.3. Marine fish
4.3.6.4. Freshwater fish
4.3.6.5. Aquarium fish
Artemia is generally used for feeding the late larval and postlarval stages of penaeids. Freshly-hatched nauplii are usually offered at the start of the first mysis stage, and sometimes even earlier at the zoea-mysis molt with some authors even recommending the introduction of Artemia during the second zoea stage.
Table 4.3.6. Typical feeding regime for Penaeus (P. vannamei) larvae.
|
Substage |
Chaetoceros neogracile |
Tetraselmis chuii |
Artemia |
|
N5 or N6 |
60000 |
0-15000 |
0 |
|
P1 |
100000-120000 |
30000 |
0 |
|
P2 |
120000 |
35000 |
0 |
|
P3 |
120000 |
35000 |
0-0.5 |
|
M1 |
100000 |
30000 |
0.2-1.5 |
|
M2 |
75000 |
20000 |
1.5-5.0 |
|
M3 |
50000-75000 |
20000 |
3-8 |
|
PL1 to PL5 |
20000-75000 |
5000-20000 |
6-20 |
Increased survival and growth have been confirmed for several penaeid speciesfed (n-3) HUFA-enriched diets, although often the effects of diet composition only become apparent in later stages (Fig. 4.3.13). A good illustration of this is the resistance to salinity stress in PL-10 stages of a batch of Penaeus monodon larvae fed on three different larval diets that varied in (n-3) HUFA levels. Thus, although no significant differences in survival were observed between treatments before the stress test, pronouned differences in PL-quality (expressed as their ability to survive the salinity stress applied) were observed (Fig 4.3.14).
This criterion of resistance to salinity shocks which can easily be applied at the hatchery level is now commonly being used as a quality criterion for determining the appropriate time for PL-transfer from the hatchery to the pond. Recent studies exploring quantitative dietary requirements as well as the relative importance of selected HUFA (i.e. DHA) showed that feeding Artemia enriched with medium levels of 12.5 mg HUFA.g-1 DW (DHA/EPA ratio of 0.4) considerably enhanced the survival of P. monodon PL-15 and the osmotic resistance of PL-10. This has recently been confirmed with the production characteristics of P. monodon PL-10 and PL-20 being significantly improved when HUFA-fortified Artemia (32 mg.g-1 DW) were administered in comparison to low-HUFA Artemia (4 mg.g-1 DW). However, no significant differences were revealed in function of various DHA/EPA ratios for the production output, apparently indicating that there is no specific requirement for DHA over EPA in postlarval shrimp.
Figure 4.3.14. Survival of P. monodon PL10 cultured on larval diet combinations containing low, medium and high levels of (n-3) HUFA after 60 min transfer from 35 to 7 g.l-1 seawater (modified from Sorgeloos and Léger, 1992).

Artemia nauplii is the most successful diet employed for the larval rearing of freshwater prawn larvae. In contrast to penaeid shrimp, Macrobrachium can initially be fed with freshly-hatched Artemia nauplii, at densities higher than 0.1 nauplii.ml-1 to ensure proper ingestion (Table 4.3.7).
Energy intake in M. rosenbergii was directly proportional not only to Artemia concentration but also to Artemia size the (n-3) HUFA-requirements of Macrobrachium were anticipated not to be very critical in view of the fact that these animals spend most of their life in freshwater.
Table 4.3.7. Variations of food amount per larva per day during larval rearing (Aquacop, 1983).
|
Day |
Artemia nauplii |
Pellets (µg DW) |
|
3 |
5 |
0 |
|
4 |
10 |
0 |
|
5-6 |
15 |
0 |
|
7 |
20 |
0 |
|
8 |
25 |
0 |
|
9 |
30 |
0 |
|
10-11 |
35 |
0 |
|
12 |
40 |
70 |
|
13-14 |
45 |
80-90 |
|
15-24 |
50 |
100-180 |
|
25-30 |
45 |
200 |
|
30+ |
40 |
200 |
Table 4.3.8. Effect of vitamin C enrichment in Artemia nauplii on the larviculture success of the giant freshwater prawn Macrobrachium rosenbergii (day 28) (Merchie et al., 1995)
|
|
experiment 1 |
experiment 2 |
|||
|
dietary ascorbic acid (µg g-1) |
529 |
2920 |
656 |
1305 |
2759 |
|
survival (%) |
72.1 |
48.4 |
57.5 |
57.8 |
57.1 |
|
ind. length (mm) |
9.31 |
9.34 |
9.67 |
9.73 |
9.58 |
|
ind. dry weight (µg) |
831 |
888 |
1130 |
1200 |
1310 |
|
metamorphosis (%) |
12.9 |
16.2 |
40.6 |
53.3 |
49.1 |
|
survival after osmotic stress (%) |
8.7 |
32.7 |
40.0 |
62.0 |
74.0 |
|
ascorbic acid in larvae (µg.g-1 DW) |
365 |
552 |
352 |
448 |
507 |
|
ascorbic acid in PL (µg.g-1 DW) |
288 |
325 |
255 |
389 |
432 |
The larvae of many species of marine fish, such as gilthead seabream, grouper, and turbot, can only be offered an Artemia diet after an initial period on a smaller prey, such as the rotifer, Brachionus plicatilis. However, n contrast to crustacean larvae, marine fish larvae are usually cultured on Artemia for a much longer period of time, (i.e. from 20 to 40 days; Table 4.3.9.). Consequently, Artemia cyst consumption can be among the highest in marine fish larviculture, ranging from 200 to 500 g per 1000 fry produced. In general, instar I nauplii are fed for several days as a transition from the rotifer diet to the larger 24-h enriched preys.
The variability of the nutritional value of Artemia nauplii as a food source for marine fish larvae has been well documented. As mentioned previously, the application of HUFA enrichment of the Artemia diet has been found to have a significant effect in marine fish larviculture, and has generally resulted in increased survival and reduced variability in fish hatchery production. The latter is particularly important since it was the missing link in the development of commercial production. Furthermore, the quality of the fry in terms of stress resistance, better pigmentation, reduced deformities, better swimbladder inflation, and increased vigor, appears to have been directly correlated with the (n-3) HUFA enrichment of their larval diet.
Table 4.3.9. Typical example of feeding regime for seabass (Dicentrarchus labrax) reared from hatching to juveniles
|
Initial fish density is ±100 larvae per l; 10-20 larvae per l during
weaning; temperature 18-20°C, salinity 35-37g.1-1 Artemia
in millions per metric ton culture volume per day; compound diets in gram
per metric ton culture volume per day or otherwise indicated in percent
of fish wet weight per day |
||||||
|
Age (days) |
Wet weight |
Artemia |
Lansy compound diets (INVE Aquaculture NV, Belgium) |
|||
|
small instar I |
Selco-enriched |
Lansy R1 |
Lansy A2 |
Lansy W3 |
||
|
0-7 |
|
|
|
|
|
|
|
8-11 |
|
1 |
|
|
|
|
|
12 |
|
2.5 |
|
|
|
|
|
13-16 |
|
2.7-3.0 |
|
|
|
|
|
17-19 |
|
5.0-7.0 |
|
2-5 |
|
|
|
20-23 |
|
3.0-4.3 |
3-11 |
5-7 |
|
|
|
24-27 |
|
0 |
14-17 |
7-10 |
|
|
|
28-29 |
|
|
17-20 |
10-15 |
|
|
|
30-34 |
|
|
20 |
10-15 |
10-15 |
|
|
35-37 |
|
|
20 |
0 |
20-30 |
|
|
38-41 |
0.05 |
|
20 |
|
30-40 |
|
|
Start of weaning |
||||||
|
|
|
|
20-15 |
|
40-50 |
|
|
|
0.08 |
|
15-10 |
|
45-55 |
15-25 |
|
|
|
|
10-0 |
|
45-55 |
45-55 |
Similarly, for good growth and survival in gilthead sea bream (Sparus aurata) larvae, the feeding regime of rotifers and brine shrimp should contain high levels of both 20:5n-3 and 22:6n-3. Moreover, the best resistance to stress conditions (i.e., activity test) was displayed by larvae fed the 22:6n-3-enriched live feed. More recently, the best growth was achieved with a diet rich in (n-3) HUFA and having a high DHA/EPA ratio of 2 during the first two weeks after hatching.
Furthermore, with turbot (Scophthalmus maximus) (n-3) HUFA enrichment and dietary DHA/EPA ratio may also be involved in larval pigmentation. For example, recent investigations on isolated turbot cells have demonstrated that the conversion from EPA to DHA is very slow in turbot, and that direct supplementation with DHA might be beneficial for the larvae of this species. However, the dosage and boosting with DHA during the early larval stages has to be considered with extreme care since the requirements of the larvae may not only depend upon their ontogenetic stage but also on their fatty acid reserves from the yolk-sac which may in turn vary with the broodstock diet.
The necessity of incorporating DHA in the larval diet of Japanese flounder has also been proven (Table 4.3.10.): the use of DHA resulting in a much higher survival and growth rate than in the control treatment and also facilitating enhanced resistance to stress conditions (day 50). Identical experiments have also been conducted with red seabream and have been even more conclusive: the growth of DHA-fed larvae being 50% better than the control group by day 38.
Table 4.3.10. Survival, growth and stress resistance of Japanese flounder Paralichthys olivaceus (day 50) fed either unenriched rotifers and Artemia (control) or high-DHA Superselco-enriched live food (DHA) Devresse et al., 1992.
|
|
control |
DHA |
|
survival (%) |
1.8 |
21.5 |
|
length (mm) |
19.1 |
28.7 |
|
stress resistance (% survival) |
40.0 |
93.0 |
Until early 1988, culture trials with mahi-mahi larvae (Coryphaena hippurus) had only been successful when the larvae were fed natural copepods or other zooplankton; culture tests with newly-hatched Artemia not being successful at that time. However, In 1988 and 1989, significant progress in the larviculture of this fast-growing aquaculture species was achieved by various research groups in the U.S.A. and Australia. In particular, larvae fed Artemia enriched with high levels of (n-3)HUFA, and in particular DHA, resulted in more consistent larviculture outputs in terms of survival, larval growth, and health as compared to larvae cultured with other zooplankton as food.
Freshwater fish larviculture is often carried out in ponds with natural zooplankton as the larval food. The salmonids, perhaps the group cultured most widely on an intensive basis, have a relatively well-developed digestive tract at first feeding and are usually fed formulated diets from start-feeding. Nevertheless, many species of freshwater fish are fed on Artemia. Whitefish larvae (family Coregonidae) are often fed Artemia until they metamorphose and can be switched to a dry diet. Walleye (Stizostedion vitreum) larvae raised on diets of either Artemia, natural zooplankton, or fish larvae preferred Artemia as a first food. Consequently, a 15-day feeding period on brine shrimp for walleye larvae prior to being fed on traditional artificial diets. Similarly, Artemia nauplii are increasingly being used within the USA as a first food for striped bass larvae (Morone saxatilis). Interestingly, although these fish are reared in freshwater or very low-salinity water, recent evidence suggests that they may have the fatty acid requirements of a marine fish (which they eventually become at adulthood). The larvae are typically fed Artemia from about 5 days post-hatching until about day 20, and then weaned onto an artificial diet by day 30, after which Artemia feeding ceases.
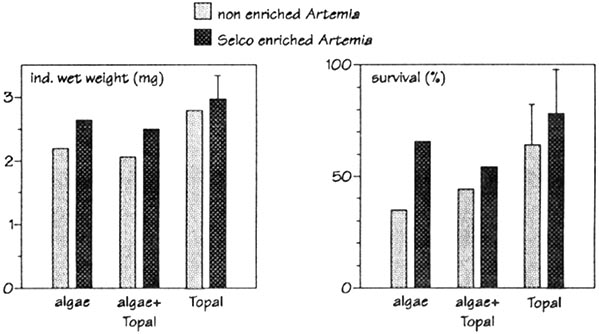
Moreover, African catfish (Clarias gariepinus) larvae have been found to perform significantly better when fed an Artemia diet as compared to co-feeding with Artemia and a semi-purified diet (ratio 80:20). Dietary supplementation with ascorbate in the form of vitamin C-boosted Artemia, was also found to have a significantly positive effect on the growth of catfish larvae one week after first-feeding; the evaluation of the physiological condition of the larvae showing significantly higher resistance with increasing dietary vitamin C concentration (Fig. 4.3.16).
A major drawback in feeding Artemia to freshwater organisms is that the Artemia die after 30 to 60 minutes in freshwater. As a consequence, they are not continuously available to the predator as they would be within marine systems, and must therefore be fed intermittently every 2 to 3 hours.
Figure 4.3.16. Lineair relationship between the larval dry weight (mg) and either the dietary ascorbic acid (µg.g-1 DW) or the ascorbic acid incorporated (µg.g-1 DW) in the Clarias gariepinus larvae (day 8).

Both live and frozen adult Artemia are used as food for aquarium fish species. Cysts are also purchased by these users and hatched for feeding as nauplii. Survival, vigor and pigmentation have been reported to be significantly improved in several tropical species when (n-3) HUFA levels were increased through boosting.
Aquacop, 1983. Intensive larval rearing of Macrobrachium rosenbergii in recirculating system. 1st International Biennal Conference on Warm Water Aquaculture - Crustacea, February 9-11, 1983. Brigham Young University, Hawaii, USA.
Beck, A.D., Bengtson, D.A. 1992. Study on Artemia XXII. Nutrition in Aquatic toxicology - diet quality of geographical strains of Artemia, p.161-169. In: Aquatic toxicology and hazard assessment. 5th Conf., ASTM STP 766. Pearson, J.G., Foster, R.B., Bishop, W.E. (Eds). Amer. Soc. Testing and Materials, Philadelphia, USA, 400 p.
Bengtson, D.A., Léger, P. and Sorgeloos, P. 1991. Use of Artemia as a food source for aquaculture. In: Artemia Biology. Browne, R.A., P. Sorgeloos and C.N.A. Trotman (Eds), CRC Press, Inc., Boca Raton, Florida, USA, pp 225-285.
Chair, M., Dehasque, M., Sorgeloos, P., Nelis, H. and De Leenheer, A.P. 1995. Live food mediated drug delivery as a tool for disease treatment in larviculture: a case study with turbot Scophthalmus maximus. J. World Aquaculture Soc., 26(2):217-219
Chair, M., Romdhane, M., Dehasque, M., Leenheer, A.P. and Sorgeloos, P.-1991. Live food mediated drug delivery as a tool for disease treatment in larviculture. II. A case study with European seabass. In: Larvi’91. Fish & Crustacean Larviculture Symposium, Lavens, P., P. Sorgeloos, E. Jaspers and F. Ollevier (Eds), European Aquaculture Society, Special Publication N°15, Ghent, Belgium, 1991, pp 412-414.
Dehasque, M., Ooghe, B., Wille, M., Candreva, Y. and Lavens, P. 1995. Automation of live food in industrial zootechnics and economics. In: Larvi’95. Lavens, P., E. Jaspers and I. Roelants (Eds), European Aquaculture Society, Spec. Publ. No 24, Gent, Belgium, pp 325-327.
Devresse, B., Romdhane, M., Buzzi, M., Rasowo, J., Léger, Ph., Brown, J. and Sorgeloos, P. 1990. Improved larviculture outputs in the giant freshwater prawn Macrobrachium rosenbergii fed a diet of Artemia nauplii enriched with n3-HUFA and phospholipids. World Aquaculture, 21(2):123-125.
Devresse, B., Leger, B., Sorgeloos, P., Murata, O., Nasu, T., Ikeda, S., Rainuzzo, J.R., Reitan, K.I., Kjorsvik, E. And Olsen, Y. 1992. Improvement of flat fish pigmentation through the use of DHA enriched rotifers and Artemia. Poster presentation at the 5th International Symposium on Fish Nutrition and Feeding, 7-10 September 1992, Santiago, Chile.
Dhert, Ph., Lavens, P., Duray, M. and Sorgeloos, P. 1990. Improved larval survival at metamorphosis of Asian seabass (Lates calcarifer) using w3-HUFA-enriched live food. Aquaculture, 90:63-74.
Dhont, J., Lavens, P., and Sorgeloos, P. 1993. Preparation and use of Artemia as food for shrimp and prawn larvae. In: CRC Handbook in Mariculture, Vol. I. Crustacean Aquaculture (2nd edition). McVey, J. (Ed.), CRC Press, Boca Raton, Florida, USA, pp 61-93.
Lavens, P., Coutteau, P. and Sorgeloos, P. 1995. Laboratory and field variation in HUFA enrichment of Artemia nauplii. In: Larvi’95. Lavens, P., E. Jaspers and I. Roelants (Eds), European Aquaculture Society, Spec. Publ. No 24, Gent, Belgium, pp 137-140.
Lavens, P., Sorgeloos, P., Dhert, Ph. and Devresse, B. 1995. Larval Foods. In: Broodstock Management and Egg and Larval Quality, Bromage, N.R. and R.J. Roberts (Eds), Blackwell Science Ltd., Oxford, UK, pp 373-397.
Léger, Ph., Vanhaecke, P. and Sorgeloos, P. 1983. International Study on Artemia XXIV. Cold storage of live Artemia nauplii from various geographical sources: Potentials and limits in aquaculture. Aquacultural Eng., 2:69-78.
Léger, Ph., Bengtson, D.A., Simpson, K.L. and Sorgeloos, P. 1986. The use and nutritional value of Artemia as a food source. Oceanogr. Mar. Biol. Ann. Rev., 24:521-623.
Léger, Ph., Naessens-Foucquaert, E. and Sorgeloos, P. 1987a. International study on Artemia. XXXV. Techniques to manipulate the fatty acid profile in Artemia nauplii and the effect on its nutritional effectiveness for the marine crustacean Mysidopsis bahia (M.). In: Artemia research and its applications. Vol. 3. Ecology, culturing, use in aquaculture. Sorgeloos, P., D.A. Bengtson, W. Decleir and E. Jaspers (Eds), Universa Press, Wetteren, Belgium, pp. 411-424.
Léger, Ph., Ferraz De Queiroz, J. and Sorgeloos, P. 1987. Improved hatchery production of postlarval Penaeus vannamei through application of innovative feeding strategies with an algal substitute and enriched Artemia. Paper presented at the 18th Ann. Meeting Guayaquil (Ecuador), January 18-23, 1987.
Léger, Ph. and Sorgeloos, P. 1992. Optimized feeding regimes in shrimp hatcheries. In: Culture of Marine Shrimp: Principles and Practices. A.W. Fast, Lester, L.J. (Eds), Elsevier Science Publishers, New York, USA, pp 225-244.
Merchie, G., Lavens, P., Dhert, Ph., Dehasque, M., Nelis, H., De Leenheer, A. and Sorgeloos, P. 1995. Variation of ascorbic acid content in different live food organisms. Aquaculture, 134: 325-337.
Romdhane, M.S., Devresse, B., Léger, Ph. and Sorgeloos, P. 1995. Effects of feeding (n-3) HUFA-enriched Artemia during a progressively increasing period on the larviculture of freshwater prawns. Aquaculture International, 3:236-242.
Seidel, C.R., Schaver, P.S., Katayama, T. And Simpson, K.L. 1980. Culture of Atlantic silversides fed on artificial diets and brine shrimp nauplii. Bull.Jap.Soc.Sci.Fish., 46(2):237-245.
Sorgeloos, P., Dehasque, M. Dhert, Ph. and Lavens, P. 1995. Review of some aspects of marine fish larviculture. ICES mar. Sci. Symp., 201:138-142.
Sorgeloos, P. and Léger, Ph. 1992. Improved larviculture outputs of marine fish, shrimp and prawn. J. World Aquaculture Soc., 23(4):251-264.
Verpraet, R., Chair, M., Léger, Ph., Nelis, J., Sorgeloos, P. and De Leenheer, A.P. 1992. Live food mediated drug delivery as a tool for disease treatment in larviculture. 1. The enrichments of therapeutics in rotifers and Artemia nauplii. Aquacultural Engineering, 11:133-139.
Worksheet 4.3.1.: Standard enrichment for Great Salt Lake Artemia.
|
· Seawater disinfection · add 1 mg. l-1 NaOCl (100 µl bleach solution. 10 l-1 of 0.45 µm filtered seawater) · Cyst disinfection · cylindroconical tube · Hatching · 2 cylindroconical tubes · Enrichment (triplicate) · take volume required for 200,000 nauplii · Harvesting · count survival, i.e. count dead nauplii (no lugol) and total nauplii (+lugol) from 3 × 250 µl sample per cone · Results · initial proportion instar I/II · Timing · Day 1 - filter + disinfect seawater 1 h + aerate |
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}