![]()
![]()
![]()
Birds may affect the incidence of disease among fish populations in a number of ways:
their presence at a water body may be necessary to the completion of a trixenic parasite life cycle which also involves fish;
they may be responsible for the spread of disease organisms from one fish population to another through migratory/foraging behaviour;
they may inflict wounds on fish, thus facilitating secondary infection by pathogens, and
they may stress fish, increasing susceptibility to disease.
Birds can act as final hosts to a number of common metazoan endoparasites of freshwater fish, particularly flukes (Class: Digenea) and tapeworms (Class: Cestoda) (Needham and Wooten, 1978). In Table 13, some of the more common parasites of European freshwater fishes which have birds as their final hosts are listed. The usual final host of fish cestode species appears to be the heron, whilst among fish digenean parasites, herons, gulls, grebes and goosanders are prevalent final hosts.
Some of the parasites listed, such as Diplostomum spathaceum, Ligula intestinalis and Diphyllobothrium spp., have a wide geographical distribution and affect a large number of fish species, whilst others, such as Valipora spp., A. muehlingi and M. yokogawi appear to be more restricted in range and choice of intermediary host species. Moreover, some of the parasites, such as P. cuticola and Tylodelphys spp., have not been reported from fish farmed in land-based (i.e., tank, pond, raceway) systems and appear restricted to open waters and caged fish.
The occurrence of some of the parasites among both wild and farmed fish population can be extremely high. In one study Fuhrmann (1979) found that 40% of one-year old silver carp (Hypophthalmicthys molitrix) stocked in ponds as fry were infected with P. cuticola. In another study McGuigan and Sommerville (1985) conducted a survey of the parasites of caged (Salmo gairdneri) and wild fishes from Loch Fad, Scotland, and found that all roach (Rutilus rutilus) and perch (Perca fluviatilis) and 65% of the feral rainbow trout examined had Diplostomum spp. Amongst the caged fish population, the prevalence of Diplostomum spp. was somewhat lower (30–37%).
Table 13
Common parasites of Europen freshwater fishes which have birds as final hosts
| Species | Fish Intermediary | Habitat | Final host | Description | References |
| (a) Cestodes | |||||
Ligula intestinalis | Cyprinids catastomids | Lakes, reservoirs fishponds | Ardea | Large (up to 20 cm), fleshy plericeroids found in body cavity. Cause compression and distortion of viscera, inhibition of gonadal development, disturbance of behaviour (do not enter spawning shoals, swim poorly) | Arme and Owen, 1968; Hoole and Arme, 1982 |
Diphyllobothrium dentriticum, D. ditremum, D. latum | Salmonids, coregonids | Lakes, reservoirs | Ardea, man, D. latum | Encyst in viscera and musculature, causing adhesion of viscera, sterility and even death. Long-living stage in fish, hence highest densities found in older fish | Fraser, 1960; Wooten and Smith, 1979; Halvorsen and Andersen, 1984 |
| Valipora campylanaristrota | Cyprinids | Lakes, reservoirs, fishponds | Ardea | Moravec, 1984 | |
| (b) Digeneans | |||||
Posthodiplostomum cuticola | Cyprinids | Lakes, reservoirs, fishponds | Ardea, Larus | Metacercariae found in sking, causing black spot disease | Fuhrmann, 1979 |
Diplostomum spathaceum | Cyrpinids, salmonids | Lakes, reservoirs, ponds, raceways, tanks | Larus, Gavia stellata | Metacercariae found in the eyes (retina, vitreous humour, lens). Causes cataracts, even blindness, and changes in behaviour | Willomitzer, 1980; Okulewicz, 1984; McGuigan and Sommerville, 1985 |
| D. gasterostei | |||||
Tylodelphus clavata | Percids, esocids, cyprinids, salmonids | Lakes reservoirs | Podiceps | Metacercariae found in the humour or retina, not in lens | Niewiadomska, 1963; Kennedy and Burrough, 1977 |
| T. prodicipina | |||||
Apophalus muehlingi | Cyprinids | Lakes | Podiceps, Larus | Metacercariae found in the iris | Okulewicz, 1984 |
Metagonimus yokogawi | Cyprinids | Lakes, reservoirs | Larus, Podiceps, Ardea | Metacercariae occur in the skin | Needham and Wooten, 1978 |
The effects of the parasites on the fish range from the largely cosmetic to those which influence host mortality. P. cuticola, which causes the unsightly black spot disease in carps, probably has little adverse affect on the host, unless at extremely high densities (Needham and Wooten, 1978). T. clavata is also believed to have little adverse effect on the growth or survivorship of the host fish species (Kennedy, 1984), for although it occurs in the eyes, and sometimes in considerable numbers, it is not found in the lens, and thus probably does not severely affect fish vision.
Some of the parasites listed in Table 13, however, are known to adversely affect their host, causing disruption of behaviour and physiological processes. Heavy infestations of L. intestinalis can severely affect behaviour and cause increased susceptibility to predation, whilst infestation of the eyes by metacercariae of Diplostomum spp., can cause cataracts and blindness which affects behaviour in general, and feeding (Crowden and Broom, 1980) and anti-predator behaviour (Brassard, Rau and Curtis, 1982) in particular.
Few of the parasites listed, however, are known to directly kill their hosts, the exception being D. spathaceum (Brassard, Rau and Curtis, 1982a).
The parasites listed in Table 13 may also cause problems for other reasons. Fish infected by Diplostomum spp. may not take artificial lures, whilst fish with heavy infestations of Diphyllobothrium spp. or L. intestinalis may be repellant to anglers, thus adversely affecting commercial sport fisheries. D. latum, although not as common as other Diphyllobothrium species, is also of concern, since the plericercoids which encyst in the viscera of the fish host are able to pass to man and re-encyst if the fish is eaten raw or is not properly cooked (Meyer, 1970).
Whilst many authors have reviewed the parasite fauna of freshwater fish communities and commented on the possible consequences, there have been few attempts to quantify host mortalities or economic losses. Anderson and Gordon (1982) suggest that by plotting data on parasite burden versus host age, quantitative data on host mortality could be obtained. They suggested that this method was particularly applicable to digeneans. However, their approach was later rejected by Kennedy (1984) who concluded that “the problem of detecting parasite-induced host mortality unambiguously in the field is still unsolved and may be insoluble”.
Thus, birds such as herons, gulls and grebes are important final hosts for a number of parasitic organisms, particularly trixenic cestodes and digeneans, which in their larval stages infest both farmed and wild fish populations. Such organisms are dominant among the parasite fauna of many freshwater lakes (Wooten, 1973; McGuigan and Sommerville, 1985), particularly the more eutrophic type, and a number of authors have considered this to be related to the relative abundance of piscivorous aquatic birds and mammals (Esch, 1971; Bayanov, 1980) (see also above).
Trixenic cestode and digenean parasites are often less common among farmed than wild fish, in terms of the range of species reported and in the prevalence and intensities of infection, and it has been suggested that this is due to physical separation of the fish from the snail or crustacean intermediary host (Jurewicz, 1959; Zitnan and Cankovic, 1970; Wooten and Smith, 1980; McGuigan and Sommerville, 1985). In lake-based cages, however, separation from crustacean zooplankton does not occur and thus Diphyllobothrium spp. infestations may be readily transmitted to the caged fish (Matheson, 1979). In land-based farms where cestode and digenean infestations of fish have been reported, these have often originated from the wild fish supplying the unit (Wooten and Smith, 1980).
In conclusion, quantitative evidence for adverse effects on growth, survivorship and economic value of parasites on wild fish populations do not exist and may be impossible to obtain. Whilst evidence for parasite-induced damage to farmed fish may be easier to collect and assess, most of the reports to date tend to be qualitative and anecdotal in nature.
The transfer of potentially pathogenic organisms by birds from one fish population to another has not been well studied. It is known that viruses such as infectious pancreatic necrosis (IPN), viral haemorrhagic septicaemia (VHS) and spring viraemia of carp (SVC) can be isolated from beaks or from regurgitated food several hours after birds had been feeding on infected fish (Olesen and Vestergaard Jorgensen, 1982; Peters and Neukirch, 1986). Orally administered IPN has also been re-isolated from the faeces of birds, even from those produced after several days ingestion (Sonstegard and McDermott, 1972; Eskildsen and Vestergaard Jorgensen, 1973; Peters and Neukirch, 1986). Recently Peters and Neukirch (1986) successfully demonstrated that it was possible to induce an IPN infection in trout fry held in an aquarium (201) after small quantities (0.2 g) of faeces collected from herons which had been feeding on infected fish were dropped in the water.
Whilst it has been shown that birds can act as mechanical vectors of viruses and whilst there is some epidemiological evidence to support the supposition that they can transfer viral diseases on fish farms (Bregnballe, 1981), the importance of birds as vectors of fish viral diseases has yet to be properly assessed.
It has been demonstrated that birds, such as gulls which scavenge on rubbish tips can pick up and disseminate pathogenic micro-organisms such as salmonellae and faecal coliforms (Williams, Richards and Lewis, 1976; Gould, 1977; Gould and Fletcher, 1978; Benton et al., 1983). Increases in bacterial numbers in reservoirs have been correlated with roosting gull numbers (Benton et al., 1983). Whilst some of the bacterial species isolated from bird faeces, such as Edwardsiella tarda, have been implicated in severe outbreaks of septicaemia and enteritis in salmonids (Roberts and Shepherd, 1986), no clear link between bird faecal contamination of water bodies and fish diseases has been established.
The ability of birds to transmit protozoan diseases has been demonstrated by Taylor and Lott (1978). They fed mallards (Anas platyrhynchos) and herons (Nycticorax nycticorax) trout infected with the myxosporodan Myxosoma cerebralis, which causes whirling disease in salmonids, and deposited the faeces in tanks of trout, some of which subsequently developed the disease.
There are a number of metazoan parasites whose larval stages infect fish and whose adult stages occur in birds (see above), and although it seems likely that parasites are transferred from one water body to another by bird hosts, this has neither been demonstrated, nor has its importance as a means of dissemination of parasitic fish diseases been evaluated.
Attacks on fish by piscivorous birds are not always successful (see above) and the resultant wounds may be sufficient to kill the fish directly, act as sites of secondary infection by pathogenic micro-organisms, or result in rejection of the fish for sale. However, there has been little study of the nature of the damage inflicted by birds or assessment of the magnitude of the problem.
Descriptions of characteristic wounds inflicted on farmed rainbow trout by herons and cormorants are given in Ranson (1982) and Ranson and Beveridge (1983). Herons attack from above and cause either scale removal on the sides of the fish by grabbing, or a single puncture wound in the head or back with minimal associated scale removal, as a result of stabbing with the beak closed (see Figure 3). These wounds were observed in both pond and caged fishes. Cormorant attacks on caged fish resulted in a puncture mark with some associated scale loss on one side of the fish, and an area of scale removal on the opposite side (Figure 3). There are no descriptions of wounds inflicted on fish by other avian species.
Studies of fishing in rivers by herons have shown that the rate of injury is positively correlated with the quantities of fish successfully captured (Geiger, 1984). However, this differs from the farmed situation. In studies of nocturnal feeding by herons at carp ponds, Draulans and van Vessem (1985) showed that the success rate of attacks was low (<70%) on arrival, increased to a maximum (92%) 6–10 minutes after arrival and thereafter declined. Catch rate was also negatively correlated with flock size.
Studies on fish predation by cormorants at carp ponds have shown that the success rate of capture of larger fish (550–700 g) is much lower than that of smaller individuals, and that fish of up to 700 g can be severely damaged (Moerbeek et al., 1987).
It must be borne in mind that not all failed attacks result in wounding: the proportion of failed attacks that do result in damage will depend on size/age and species involved, as well as the nature of the environment (e.g., farmed/open waters; cage/pond). For example, Ranson and Beveridge (1983) concluded that few, if any, attacks by herons and cormorants on caged fish were likely to be successful, whereas those on fish reared in ponds or raceways certainly are.
There are no published data on the extent of bird-induced wounding amongst fishes of open waters. Estimates from one cage rainbow trout farm on the west coast of Scotland, which was visited by both herons and cormorants, showed that on average 2.2% of all live fish examined had been wounded by birds, although the proportion varied with time of year, size, duration of time the fish had spent on the farm and mesh size (Ranson, 1982). In a study of trout ponds visited by herons, less than 2% of fish showed signs of having been attacked (Meyer, 1980; 1981).
There have been few estimates of economic losses caused by wounding, although Meyer (1980) has discussed the proportion of fish rejected for sale on these grounds (see above). Neither the effects of scale loss nor the invasion of wound sites by pathogenic organisms have been studied. From observations on cage rainbow trout farms, however, it was observed that more than 4% of dead fish retrieved by staff from cages had bird-inflicted wounds (2% of healthy stock, see above) suggesting that wounded fish were almost twice as likely to die as individuals which had not been physically damaged (Ranson, 1982).
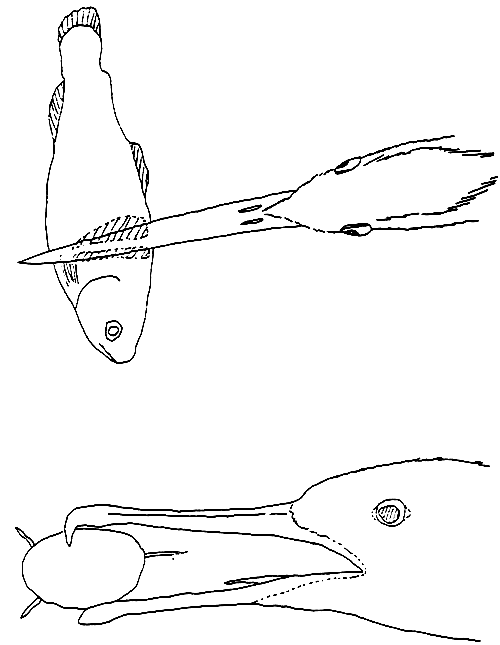
Figure 3 Sketch showing damage inflicted by herons and cormorants. Shaded area indicates scale removal (from Ranson and Beveridge, 1983)
It has been demonstrated under laboratory conditions that the presence of piscivorous birds can disrupt the normal swimming and foraging behaviour of fish (Milinski and Heller, 1978; Giles, 1983), although this has not yet been studied in wild or farmed fish populations (see above). Physiology-based studies of stress in fish caused by birds have also yet to be carried out.
Approaches to alleviating problems caused by birds at inland water fisheries, hatcheries and fish farms, fall into two broad categories; prevention and cure. Prevention of bird problems can be achieved through careful site selection, but, of course it is an option which is only open to hatcheries and farms, unless, perhaps, reservoirs are included. However, economic and logistic considerations in reservoir siting will far outweigh any foreseeable problems that may be caused by birds, and so remedial measures are likely to be the only option available to managers of open waters.
Preventative measures have been recommended as a means of forestalling bird problems in the USA. Lagler (1939) and Salyer and Lagler (1946) noted that the incidence of kingfisher predation was much greater at hatcheries built on or close to the Great Lakes than those sited further inland, and suggested that fish culture establishments should be located away from the major migration routes or flyways of fish-eating birds, and outside the flight range of large nesting or roosting colonies. Similar advice has more recently been propounded by the US Fish and Wildlife Service and Department of the Interior (Salmon and Conte, 1982).
Whilst it sometimes may be possible to follow such advice, there are very likely to be over-riding economic considerations why this may not be practicable. Moreover, some species, such as the heron, can fly considerable distances in order to forage for food (Marion, 1984), and colonies are so common, that in part of Europe it may just not be possible to avoid them. There are also difficulties posed by species such as the cormorant and the black-headed gull, which are greatly extending their ranges in Europe (Cramp et al., 1977; 1983; Hansen, 1984; Im and Hafner, 1984). For example, the pond-based farm at Lelystad, the Netherlands, which was completed in 1963, had no problems with cormorant predation until a colony became established in nearby IJsselmeer in 1978 (Moerbeek et al., 1987).
A number of methods for reducing existing problems caused by birds have been proposed and/or evaluated, ranging from attempts to remove or dissuade visiting birds to reducing the vulnerability of fish to attack.
The shooting of birds visiting fish farms or lakes was once an almost universally practiced panacea (see Lagler, 1939; Mott, 1976; Dombeck, Hammill and Bullon, 1984, for reviews). Today the majority of bird species are protected by law in most, although not all (see below), parts of North America and Europe, and whilst this has had an undoubted effect on reducing the incidence of birds shot, shooting is still resorted to by many fish farmers. For example, although the heron is protected by law in England, Meyer (1981) estimates that some 3 600–5 000 birds are shot each year on fish farms, representing a significant proportion of the British population (estimated at around 5 400 breeding pairs in the late 1970s). In some parts of Europe, such as Scotland and Denmark, herons may be shot quite legally (Mills, 1979; Moller and Olesen, 1984), albeit that a licence may be required. In Denmark it is estimated that approximately 8% of the heron population are killed each year at fish farms (Moller and Olesen, 1984).
The reasons for the appeal of shooting as a method of control are patently obvious: it is an immediate and apparently effective response to a perceived problem which probably engenders a feeling of satisfaction in the perpetrator. However, numerous studies have shown that the efficacy of shooting is dubious, being either totally ineffective, or resulting in only temporary remission. It is also costly.
Researchers in North America have shown that it is possible to control, or even eliminate, goosander populations by shooting and that this can cause a marked improvement in fish catches (White, 1939; Huntsman, 1941; Elson, 1962; Smith, 1968; Erskine, 1972). However, Elson's studies showed that goosander numbers had to be reduced to around one-third of the initial density, involving massive investments in manpower, before shooting proved effective, whilst Smith's eradication of birds programme was carried out simultaneously with a lake fertilization programme and the extermination of piscivorous mammals, making it difficult to evaluate the effectiveness of the avian control component.
Studies in the UK, Germany and Belgium have all shown that shooting of herons has little real impact on the number of fish taken, as immigration of replacement birds rapidly takes place (Meyer, 1980, 1981, 1982; Ranson and Beveridge, 1983; Utschick, 1983; van Vessem, Draulans and de Bont, 1985). Moreover, as Draulans and van Vessem (1985) have demonstrated, the intensity of predation is greatest in the first few minutes after landing at the ponds and rapidly decreased with time, and so birds would have to be shot as soon as possible after arrival in order to effectively reduce the quantity of fish taken. They also showed that success in prey capture was significantly higher among small groups of feeding birds, thus negating the effects of shooting, in part at least (see above).
Other methods for removal of birds, such as trapping, poisoning and capture and release have all been tried, usually with similar results to shooting. Van Vesem, Draulans and de Bont (1985) tried releasing herons 30–150 km from the site of capture. Although none of the adults reappeared, many of the juveniles were observed back at the farm within one month.
A large number of bird scaring devices, including both audio and visual deterrents, sometimes in combination, have been tried with mixed success. Audio deterrents include loud noises (gun-shots, humlines, automatic gas canons, fireworks) taped human voices and distress or alarm calls. Certain types of noise, such as those caused by passing vehicles, have been shown to be ineffective in reducing bird numbers (Draulans and van Vessem, 1985). Lagler (1939), Salyer and Lagler (1946) and Spanier (1979, 1980) have shown that loud noises are only temporarily effective in deterring visiting birds, whilst Hewitt (1936) and Moerbeek et al., (1987) claim that they are useless. In a recent review, Salmon and Conte (1982) state that most loud noise scaring devices are of limited value and stress the importance of choosing automatic equipment which emits noises at irregular intervals and whose position or direction of sound emission can be changed frequently.
The use of loud noise scarers is not recommended if the site is close to human habitation or if there are livestock or game animals in the vicinity (Salmon and Conte, 1982; Im and Hafner, 1984). Salmon and Conte also state that loud noises can disturb spawning catfish.
Taped alarm or distress calls have been successfully used to deter a variety of visiting birds from open waters (Benton et al., 1983; Shedden, 1983) and fishponds (Spanier, 1979; Salmon and Conte, 1982; Utschick, 1983). Reaction varied with species, location, size of area and time of year, and experience has shown that this method is probably most effective if the calls are broadcast as the birds begin to arrive. Salmon and Conte (1982) state that birds soon become used to these calls. However, whilst Utschick's (1983) study of herons corroborates their views, studies of gulls and night herons have shown this method to be effective over many months or even years (Spanier, 1979, 1980; Benton et al., 1983; Shedden, 1983).
A wide range of visual deterrents, including foil and cloth strips, flags, balloons, flashing lights, model aircraft, scarecrows and model raptors have been used. Again, results have been variable, depending upon species, time of year and location (Lagler, 1939; Salyer and Lagler, 1946; Meyer, 1981; Salmon and Conte, 1982; Utschick, 1983).
In conclusion, the effectiveness of most visual and many audio deterrents is questionable. Greater success has been achieved with some species, such as gulls, than others (e.g., cormorants) and it is important that equipment is mobile and that audio emissions are varied in direction and given at irregular intervals. A number of studies have shown that large flocks return more quickly after disturbance than small flocks and that it is more difficult to deter birds if there are no alternative feeding or roost sites nearby (Ranftl and zur Muhlen, 1977; Draulans and van Vessem, 1985). It is also more difficult to deter piscivorous species if the ponds are heavily stocked (Utschick, 1983; Barlow and Bock, 1984). A combination of the above deterrent techniques is probably more effective than reliance on any single strategy. Other deterrents which have been less frequently tested include water spray systems and electrically charged wires. The former has proved particularly effective against gulls roosting on reservoirs in North America, whilst the latter has been shown to be largely ineffective (Emigh, 1962, in Mott, 1976; Salmon and Conte, 1982).
There are a number of methods available for reducing vulnerability to attack by piscivorous birds. The most commonly employed involves the construction of some kind of physical barrier. The complete enclosure of some aquaculture facilities by netting or mesh has been shown to be very effective in eliminating predation by all types of piscivorous birds. In order to protect caged stock, 5–10 cm mesh top nets must be firmly secured to the cages and curtains of similar sized mesh must be hung around the cages (Ranson, 1982; Ranson and Beveridge, 1983; Beveridge, 1987). The anti-predator net curtain must be hung at least 30–40 cm distance from the cage bag and should extend for several metres below the cage floor. Even then, damage by diving birds can still occur (Ranson, 1982). Moreover, anti-predator nets are expensive, reduce current flow through cages and add to management problems, as well as trapping and killing occasional birds.
Tanks and small raceways can be effectively and inexpensively protected by covering with netting or chicken mesh (Hewitt, 1936; Lagler, 1939; Mott, 1978; Salmon and Conte, 1982; Martin, 1982). Mesh size must be related to the size of the piscivore and the mesh or netting should be attached to a wooden or pipework frame so that it can be readily removed to facilitate routine operations.
Small ponds, densely stocked with valuable fry or fingerlings, may be cost-effectively protected by entirely enclosing in netting, or wire or plastic mesh, supported by a wooden or metal framework of posts and beams. The resultant cage can be entered through a screened gate and should be high enough to allow staff to work without discomfort. Such caging is compulsory at Danish fish farms (Meyer, 1981).
Where large ponds or open waters must be protected, partial enclosure using overhead wires or lines is less expensive and is much more practicable than complete enclosure. Wire, braided, mono- and multi-filament man-made fibres are commonly used. Choice of material and system of enclosure should be based upon cost, expected duration of damage, size of facility, extent and nature of predation, management, location and aesthetics. Various paterns of l iens have been tried: parallel, grid, irregular and radial. However, few experiments have been tried to determine the most cost-effective system. Moerbeek et al. (1987) found no significant differences in effectiveness between a regular grid pattern, irregularly placed lines, or a radial pattern of lines.
The distances between lines should be determined by species. According to Salmon and Conte (1982) 120 cm spacings are effective in deterring gulls, 60 cm spacings should be used against goosanders and 30 cm for herons. Moerbeek et al. (1987) found that 20 m spacings were as effective as 10 m spacings in deterring cormorants. The recommended height of the lines above the water is 40–60 cm (Lagler, 1939; Moerbeek et al., 1987).
Partial screening suffers from a number of disadvantages. It is useless against very small birds, such as kingfishers (Salmon and Conte, 1982) and is only partly effective against other species. Moerbeek et al. (1987) showed that only 5–11% of cormorants visiting their fish farm left without landing as a result of the lines, and that few of the birds encountered problems either during landing or take off, irrespective of the system of lines used. Although Im and Hafner's (1984) study of cormorant predation in the Camargue showed that a system of lines was successful in repelling most visiting birds, it was not made clear whether there werer unprotected ponds with attractive stock close by.
Partial screening has high associated maintenance costs and can make routine operations around the farm difficult, particularly if lines have been laid out at the ends of the system in order to prevent wading birds from simply walking in under the wires. This type of screening is also considered by many as unsightly and is not practicable for protecting large open areas of water.
If wading birds are the principal problem species, then extremely simple systems may be employed to protect farm stock. Lagler (1939) suggested that marginal chicken wire fences, 60 cm high, would be sufficient to protect raceways against predators. More recently, Meyer (1980, 1981, 1982) has claimed that a single or double cord fence, 25–35 cm high, in combination with a floatline laid around the margins is an inexpensive and effective method of protecting pond stocks against herons. A similar system has been suggested by Salmon and Conte (1982).
Alternative methods of reducing the vulnerability of valuable stock to avian predators have involved the introduction of a buffer population of fish, either by mixing smaller fishes with more valuable larger ones, or by introducing a low value, easily caught species, such as the mosquito fish (Gambusia affinis) (Lagler, 1939; Utschick, 1983; Barlow and Bock, 1984). Lagler (1939) also suggested building small ponds close to the fish culture ponds and stocking them with frogs and toads.
Fraser (1974) attempted to reduce the vulnerability of brook trout to predation by training hatchery reared fishes to avoid an electrified model bird moving through the water in the hatchery raceway. Low survivorship rates of released fish suggest that the training was of little value.
Finally, it has been suggested that predation by birds can be reduced by modifying the ways in which farms are designed and managed. It is argued that tank, raceways or cage systems are much less vulnerable and easy to protect than large earth ponds (Salmon and Conte, 1982; Beveridge, 1987). Deeper ponds (1 m) with steep-sided banks have been shown to deter waders such as herons (Meyer, 1980, 1981, 1982). However, deep ponds may not be cost-effective and steep banks are prone to erosion, although this can be reduced by compaction with heavy clay. Banks and levees should be cleared of vegetation which may be used as cover, and any facility that can possibly be used as a perch removed (Cottam and Uhler, 1936). Salmon and Conte (1982) recommend that raceways with vertical walls rising at least 1 m above the water surface will prevent many species from taking fish.
Good management practices can help. Feed bags should not be left outside and dead fish should be removed and carefully disposed of (Beveridge, 1987). All screens and netting on cages, tanks and raceways must be secured and maintained in good order. Changes in management practices may also be beneficial. It has been suggested that valuable or vulnerable fishes should be stocked in ponds close to farm office buildings, as proximity to man will deter many species (Salmon and Conte, 1982; Moerbeek et al., 1987). Encouraging the public on to the farm to fish in the ponds may serve not only as an added source of revenue, but may also help keep birds away.
Reductions in stocking density, although not always practicable, can minimize the incidence of predation (Barlow and Bock, 1984). Changes in the timing of stocking may be more effective. Moerbeek et al. (1987) suggest delaying transplanting carp fry from the hatchery to open ponds until late June. As a result, not only will the period when they are vulnerable to predation be shorter, but the number of visiting cormorants should also be greatly reduced. In Canada, Mace (1983) reduced the incidence of predation by gulls on hatchery-reared juvenile salmonids to 5–15% by delaying the release until most of the predatory migrating birds had disappeared, restricting the release to tides and times of day when the birds were least effective at feeding, and by decreasing the duration of the release period in order to minimize the accumulation of predators.
In conclusion, a number of preventative and ameliorative measures are available to fisheries managers and fish farmers for reducing problems associated with birds. However, few options other than the complete enclosure of stock are likely to be completely effective, and a combination of strategies is likely to be superior to reliance on any single measure. The cost-effectiveness of any measures should be carefully considered.
Changes in public attitudes and in the law means that killing or maiming as a means of reducing problems is now unacceptable. Indeed, conservation of many bird species traditionally viewed as enemies by fishermen and fishfarmers is of increasing importance as there is growing public interest in their aesthetic and recreational value (Cottam and Uhler, 1936; Mott, 1978; Dombeck, Hammill and Bullon, 1984). In the USA, the Fisheries and Wildlife Service has recently instigated a research programme into making fish-eating birds and fish rearing more compatible. In the UK, Mills (1979) reports that one fish farmer has turned an apparent problem into profit by encouraging the public to visit his farm to view a rare pair of ospreys fish trout from the ponds.
I would like to thank the following people for providing me with data and references:
Dr P. Monaghan and Dr N. Metcalfe (University of Glasgow), Ms C. Benton and Mr F.R. Kahn (Strathclyde Regional Council Water Department), Dr R. Sibly (University of Reading) and Dr R.K. Elston.
Adams, S.M., B.L. Kimmell and G.R. Ploskey, 1983. Sources of organic matter for reservoir fish production. A trophic-dynamic analysis. Can.J.Fish.Aquat.Sci., 40:1480–95
Anderson, R.M. and D.M. Gordon, 1982. Processes influencing the distribution of parasite members within host populations with special emphasis on parasite-induced host mortalities. Parasitology, 85:373–98
Anderson, A., 1970. Food habits and predation of an inland breeding population of the herring gull, Larus argentatus in southern Sweden. Ornis Scand., 1:75–81
Arme, C. and R.W. Owen, 1968. Occurrence and pathology of Ligula intestinalis infections in British fishes. J.Parasitol., 54:272–80
Backiel, T. and E.D. Le Cren, 1967. Some density relationships for fish population parameters. In The biological basis of freshwater fish production, edited by S.D. Gerking. Oxford, Blackwell, pp. 261–93
Barlow, C.G. and K. Bock, 1984. Predation of fish in farm dams by cormorants. Aust.Wildl.Res., 11(3):559–65
Bauer, K., 1965. Zur Nahrungsokologie einer binnenlandischen. Population der Flusseeshwalbe (Sterna hirundo). Egretta, 8:35–51
Bauer, K.M. and U.N.G. von Blotzheim, 1986. Handbuch der Vogel Mitteleuropas Vol. I. Graviiformes. Phoenicopteriformes. Frankfurt am Main, Akademia, Verlag, 483 p.
Bayanov, M.G., 1980. Circulation of helminths in aquatic-paludal birds in water bodies of different trophic levels and mineral content in the Southern Urals, Russian SFSR. Ekologiya, 1980:56–62 (in Russian)
Benton, C., et al., 1983. The contamination of a major water supply by gulls (Larus spp.): a study of the problem and remedial action taken. Water Res., 17:789–98
Bergman, G., 1961. The food of birds of prey in Fenno Scandia. Br.Birds, 54:307–20
Beveridge, M.C.M., 1987. Cage aquaculture. Farnham, Surrey, Fishing News Books, Ltd.
Beveridge, M.C.M., M. Beveridge and J.F. Muir, 1982. Cage fish culture and Loch Lomond. Stirling, Scotland, Institute of Aquaculture, University of Stirling, 68 p.
Brasard, P., M.E. Rau and M.A. Curtis, 1982. Parasite-induced susceptibility to predation in diplostomiasis. Parasitology, 85:495–501
Brasard, P., 1982a. Infection dynamics of Diplostomum spathaceum cercariae and parasite induced mortality of fish hosts. Parasitology, 85:489–93
Bregnballe, F., 1981. Can the heron transmit Egtved virus? Medd.Fors., (64):22 p.
Brown, L.H. and E.K. Urban, 1969. The breeding biology of the great white pelican Pelecanus onocioralus roseus at Lake Shala, Ethiopia. Ibis, 111:199–237
Cook, D.C., 1978. Foraging behaviour and food of grey herons on the Ythan estuary. Bird Stud., 25:17–22
Cottam, C. and F.M. Uhler, 1936. The role of fish-eating birds. Prog.Fish-Cult., 14:1–14
Cottam, C., 1945. Birds in relation to fishes. Wildl.Leafl.US.Fish.Wildl.Serv., (272)
Cramp, S. (ed.), et al., 1977. Handbook of the birds of Europe, the Middle East and North Africa. Vol. 1. Ostrich to ducks. Oxford, University Press, 722 p.
Cramp, S. (ed.), 1980. Handbook of the birds of Europe, the Middle East and North Africa. Vol. 2. Hawks to bustards. Oxford, University Press, 695 p.
Cramp, S. (ed.), 1983. Handbook of the birds of Europe, the Middle East and North Africa. Vol. 3. Waders to gulls, Oxford, University Press, 913 p.
Cramp, S. (ed.), 1985. Handbook of the birds of Europe, the Middle East and North Africa. Vol. 4. Terns to woodpeckers. Oxford, University Press, 960 p.
Creutz, G., 1958. Wo Fehlt der Haussperling als Brutvogel? Fake, 5:98–107, 116–9
Crowden, A.E. and D.M. Broom, 1980. Effects of the eye fluke, Diplostomum spathaceum, or the behaviour of dace (Leuciscus leuciscus). Anim.Behav., 28:287–94
Curry-Lindal, K., 1969. Nordic fauna: mammals, birds, reptiles, amphibics and fish. Stockholm, Forum, 464 p.
Danell, K. and K. Sjoberg, 1980. Foods of wigeon, teal, mallard and pintail during the summer in a northern Swedish lake. Swed.Wildl.Res., 11:141–67
Deckx, L., 1978. Recollections of the ospreys on the fish-farm ponds (8, 10 and 2 hectares resp.) at Postel (Mol.). Wielewaal, 44:305–7
Dementiev, G.P. and N.A. Gladkov, 1957. Ptitsy Sovietskogo soyuza. (Birds of the Soviet Union). Vol. 1. Moscow, Sovietskoya Nanka Publishing House, 668 p. English translation published by Israel Program for Scientific Translations, Jerusalem, IPST Cat. No. 1120:728 p. (1966)
Deufel, J., 1984. Kormorane-ein problem für die fischerei. Z.Binnenfisch.D.D.R., 33(3): 19–22
Din, N.A. and S.K. Eltringham, 1974. Ecological separation between white and pink-backed pelicans in the Ruwenzori National Park, Uganda. Ibis, 116:28–43
Dobrolowski, K.A., 1973. Role of birds in Polish wetland ecosystems. Pol.Arch.Hydrobiol., 20:217–21
Dobrolowski, K.A., R. Halba and J. Nowicki, 1976. The role of birds in eutrophication by import and export of trophic substances of various waters. Limnologica, 10:543–9
Dombeck, M., J. Hammill and W. Bullon, 1984. Fisheries management and fish dependent birds. Fisheries, 9(2):2–4
Doornbos, G., 1979. Winter food habits of smew (Mergus albetlus) on Lake Yssel, the Netherlands: species and size selection in relation to stocks. Ardea, 67:42–8
Draulans, D. and J. van Vessem, 1985. The effect of disturbance on nocturnal abundance and behaviour of grey herons (Ardea cinerea) at a fish farm in winter. J.Appl.Ecol., 22:19–28
Draulans, D., 1985a. Age related differences in the use of time and space by radio-tagged grey herons Ardea cinerea in winter. J.Anim.Ecol., 54:771–80
Eadie, J.McA. and A. Keast, 1982. Do goldeneye and perch compete for food? Oecologia, Berl., 55:225–30
Elson, P.F., 1962. Predator-prey relationships between fish-eating birds and Atlantic salmon. Bull.Fish.Res.Board Can., (133):87 p.
Eriksson, M.O.G., 1979. Competition between freshwater fish and goldeneye Bucephala clangula (L.) for common prey. Oecologia, Berl., 41:99–107
Eriksson, M.O.G., 1984. Acidification of lakes: effects on waterbirds in Sweden. Ambio, 13:260–2
Eriksson, M.O.G., 1985. Prey detectability for fish-eating birds in relation to fish density and water transparency. Ornis Scand., 16:1–7
Eriksson, M.O.G., 1986. Reproduction of black-throated diver, Gavia artica in relation to fish density in oligotrophic lakes in southeastern Sweden. Ornis Scand., 17:245–8
Erskine, A.T., 1972. Populations, movements and seasonal distribution of mergansers. Can.Wildl.Serv.Rep., (17):36 p.
Esch, G.W., 1971. Impact of ecological succession on the parasite fauna of centrarchids from oligotrophic and eutrophic ecosystems. Am.Midl.Nat., 86:160–8
Eskildsen, U.K. and P.E. Vestergaard Jorgenson, 1973. On the possible transfer of trout pathogenic viruses by gulls. Riv.Ital.Piscic.Ittiopatol., 8:104–5
Fraser, J.M., 1974. An attempt to train hatchery-reared brook trout to aovid predation by the common loon. Trans.Am.Fish.Soc., 103:815–8
Fraser, P.G., 1960. The occurrence of Diphyllobothrium in trout, with special reference to an outbreak in the west of England. J.Helminthol., 34:59–72
Fuhrmann, B., 1979. Posthodiplostomum cuticola als parasit beim silverkarpfen (Hypophthalmichthys molitrix). Z.Binnenfisch.D.D.R., 26:202–4
Gaevskaya, N.S. (Transl. D.G.M. Muller), 1969. The role of hyler aquatic plants in the nutrition of the animals of freshwater basins. Boston Spa, England, National Lending Library for Science Technology. 3 Vols. (Transl. of Russian ed. (1966))
Geiger, C., 1984. Graureihers Ardea cinerea und fischbesland in fliesgewassern. Ornithol.Beob., 81:111–31
Geiger, W., 1957. Die Nahrung der Haubenfaucher (Podiceps cristatus) des Bielsesse. Ornithol.Beob., 54:97–133
Geroudet, P., 1974. Les 18. Nos Oiseaux, 32:188–207
Giles, N., 1983. Behavioural effects of the parasite Schistocephalus solidus (Cestoda) on an intermediate host, the three-spined stickleback, Gasterosteus aculeatus L.. Anim.Behav., 31:1192–4
Glutz von Blotzheim, U.N., K.M. Bauer and E. Bezzel, 1971. Handbuch der Vogel Mitheleuropus. Vol. 4. Frankfurt am Main, Akademia Verlag
Goethe, F., 1975. Zur Biologie der Herringsmowe Larus fuscus unter besonderer Berucksichtigung ethologischer Aspeck. 1. Nahrung und Nahrungserwerb. Ornis Fenn., 52:5–12
Gould, D.J., 1977. Gull droppings and their effects on water quality. WRC Tech.Rep.Medmenham, U.K., (TR37):24 p.
Gould, D.J. and M.R. Fletcher, 1978. Gull droppings and their effects on water quality. Water Res., 12:665–72
Hachler, E.M., 1959. Von Zug der Wildganse im Teichgebiet von Lednice. Sylvia, 16:113–27
Hafner, H. and M. Moser, 1980. Les herons et la pisciculture en Camargue. Bull.Mens.Off.Natl.Chasse, Paris, No. Spec., pp. 255–60
Hakkinen, 1978. Diet of the osprey Pandion haliaetus in Finland. Ornis Scand., 9:111–6
Hallet, C., 1977. Contribution à l'étude du régime alimentaire du martinpêcheur (Alcedo atthis) dans la vallée de la Lesse. Aves, 14:128–44
Hallet, C., 1978. Etude de la taille des poissons capturés per le martinpêcheur. Héron, 1978:84–8
Hallet, C., 1982. Prey size selection by the European kingfisher Alcedo atthis: the sizes of bullheads (Cottus gobio) and trout (Salmo trutta) most frequently taken. Terre Vie, 36:211–2
Halvorsen, O. and K. Andersen, 1984. The ecological interaction between Arctic charr, Salvelinus alpinus (L.), and the plericercoid stage of Diphyllobothrium ditremum. J.Fish Biol., 25:305–16
Hanson, K., 1984. The distribution and numbers of the southern cormorant Phalacrocorax carbo sinensis in Europe. Dan.Ornthol., Foren.Tidsskr., 78:29–40
Harris, M.P., 1965. Food of some Larus gulls. Ibis, 107:43–53
Henderson, H.F., R.A. Ryder and A.W. Kudhongania, 1973. Assessing fishery potential of lakes and reservoirs. J.Fish.Res.Board Can., 30:2000–9
Hewitt, E.R., 1936. Fish-eating birds have no place in trout waters. Prog.Fish-Cult., 16:11–2
Hickling, R.A.O., 1977. Inland wintering of gulls in England and Wales, 1973. Bird Stud., 24:79–88
Hoole, D. and C. Arme, 1982. Ultrastructural studies on the cellular response of roach, Rutilus rutilus, L., to the plericercoid larva of the pseudophyllidean cestode, Ligula intestinalis. J.Fish.Dis., 5:131–44
Hunter, M.J. Jr., et al., 1986. Duckling responses to lake acidification: do black ducks and fish compete? Oikos, 47:26–32
Huntsman, A.G., 1941. Cyclical abundance and birds versus salmon. J.Fish.Res.Board Can., 5:227–35
Im, B.H. and H. Hafner, 1984. Impact des oiseaux piscivores et plus particulièrement du Grand Cormoran (Phalacrocorax carbo sinensis) sur les exploitations piscicoles en Camargue, France. Rapport. Arles, France, Station Biologique de la Tour du Valat, 84 p.
Iribarren, I.B. and L.D. Nevado, 1982. Diet of the kingfisher (Alcedo atthis). Alauda, 50:81–91
Jacoby, H., G. Knotzsch and S. Schuster, 1970. Die Volgerl des Bodenseegebietes. Ornithol.Beob., 67(Suppl.):260 p.
Jenkins, D. and M.V. Bell, 1985. Vertebrates, except salmon and trout, associated with the River Dee. In The biology and management of the River Dee, edited by D. Jenkins. Huntingdon, U.K. Institute of Terrestial Ecology, pp. 83–93
Junor, J.F.R., 1972. Estimation of the daily food intake of piscivorous birds. Ostrich, 43(1):193–205
Jurewicz, E.S., 1959. Expansion of cercariae of Diplostomum spathaceum. Rud. 1819, a common parasite of fishes in the littoral zone of the lake. Pol.Arch.Hydrobiol., 6:105–16
Kale, H.W. II, 1965. Ecology and biochergetics of the long-billed marsh wren (Telmatodytes palustris griseus) in Georgia salt marshes. Publ.Nuttal Ornithol.Club, 5:142 p.
Kennedy, C.R., 1984. The use of frequency distributions in an attempt to detect host mortality induced by infections of diplostomatid metacercariae. Parasitology, 89:109–220
Kennedy, C.R. and R. Burrough, 1977. The population biology of two species of eye fluke, Diplostomum gasterostei and Tylodelphus clavata in perch. J.Fish Biol., 11:619–33
Keve, A., 1962. Der zug des Austerfischers in Karpathenbecken. Allat.Kozlem., 49:75–9
Korodi Gal, J., 1964. Data on the Dalmation pelican's territorial extension, biometry and nutrition in Romania. Aquila, (69–70):71–82
Kramer, A., 1984. Zum einflus des graureihers Ardea cinerea auf den fischbesland von forellen bachen. Ornithol.Beob., 81:149–58
Lack, D. and D.F. Owen, 1955. The food of the swift. J.Anim.Ecol., 24:120–36
Lagler, K.F., 1939. The control of fish predators at hatcheries and rearing stations. J.Wildl.Manage., 3:169–79
Latta, W.C. and R.F. Sherkey, 1966. Feeding behaviour of the American merganser in captivity. J.Wildl.Manage., 30:17–23
Leentvaar, P., 1967. Observations on guanotrophic environments. Hydrobiologia, 29:441–89
Lemmetyinen, R., 1973. Feeding ecology of Sterna paradisaea Pontopp. and S. hirundo L. in the archipelago of south western Finland. Ann.Zool.Fenn., 10:507–25
Linn, I.J. and K.L.I. Campbell, 1986. Cormorants and fisheries. A report on the biology of the white-breasted cormorant, Phalacrocorax carbo, as it affects the commercial fisheries of Lake Malawi. Colchester, U.K., University of Essex, 51 p.
Linnman, G., 1983. Seasonal eutrophication by wildfowl in basins isolating from the sea: a working hypothesis. Hydrobiologia, 103:159–63
Llewellyn, L.C., 1983. Movements of cormorants in south-eastern Australia and the influence of floods on breeding. Aust.Wildl.Res., 10:149–67
Mace, P.M., 1983. Bird predation on juvenile salmonids in the Big Qualicum Estuary, Vancouver Island. Can.Tech.Rep.Fish.Aquat.Sci., (1176):79
Mack, G., 1941. Cormorants and the Gippsland lakes fishery. Mem.Natl.Mus.Vict., (12):95–117
Madsen, F.J., 1957. On the food habits of some fish-eating birds in Denmark. Dan.Rev.Game Biol., 3:19–38
Madsen, F.J. and R. Sparck, 1950. On the feeding habits of the southern cormorant (P. carbo sinensis Shaw) in Denmark. Dan.Rev.Game Biol., 1:45–76
Marion, L., 1983. Etat de nos connaissances sur les relations oiseaux piscivores et populations piscicoles naturelles et artificielles et propositions de recherches prioritaires pour la période 1984–88. Rapport du Groupe de travail “Oiseaux piscivores” du Ministère de l'Environnement francais. Paris, Ministere de l'Environnement, 92 p.
Marion, L., 1984. Mise en evidence par biotélémétrie de territoires alimentaires individuels chez un oiseau colonial, le héron cendré Ardea cinerea. Mécanisme de répartitien et de régulation des effectifs des colonies de hérons. L'Oiseau, Rev.Fr.Ornithol., 54:1–78
Martin, M., 1982. Impact of predators on fish farming. 1 and 2. Aquacult.Mag., 8(2–3):36–7; 44–7
Matheson, A.A., 1979. An investigation of the occurrence and control of plericeroids of Diphyllobothrium spp. in farmed Atlantic salmon. Unpubl. M.Sc. Thesis, University of Stirling, Scotland.
Mattingley, A.H.E., 1927. Cormorants in relation to fisheries. Condor, 36:182–7
McColl, J.G. and J. Burger, 1976. Chemical inputs by a colony of Franklin's Gulls nesting in cattails. Am.Midl.Nat., 96:270–80
McGuigan, J.B. and C. Sommerville, 1985. Studies on the effects of cage culture of fish on the parasite fauna in a lowland freshwater loch in the west of Scotland. Z. Parasitenkd., 71:673–82
McIntosh, R., 1978. Distribution and food of the cormorant on the lower reaches of the River Tweed. Fish.Manage., 9:107–13
Meyer, J., 1980. Herons at fish farms. In Proceedings of the Institute of Fisheries Management Eleventh Study Course, 23–25 September 1980, University of Sussex. London, Janssen Services, pp. 58–63
Meyer, J., 1981. Easy pickings: whether herons need be a menace at fish farms. Birds, RSPB Mag., Summer issue: 51–3
Meyer, J., 1982. Room for birds and fish. Fish Farmer, 4:23–6
Meyer, M.C., 1970. Cestode zoonoses of aquatic animals. J.Wildl.Dis., 6:249–54
Milinski, M. and R. Heller, 1978. Influence of a predator on the optimal foraging behaviour of sticklebacks (Gasterosteus aculeatus L.). Nature, Lond., 275:642–4
Mills, D.H., 1962. The goosander and red-breasted merganser as predators of salmon in Scottish waters. Freshwat.Salmon Fish. Res., 29:10 p.
Mills, D.H., 1962a. The goosander and red-breasted merganser in Scotland. Annu.Rep.Wildfowl Trust, (13):79–92
Mills, D.H., 1964. The ecology of the young stages of the Atlantic Salmon in the River Bran, Ross-Shire. Freshwat. Salmon Fish.Rev., 32:1–58
Mills, D.H., 1965. The distribution and food of the cormorant in Scottish inland waters. Freshwat.Salmon.Fish.Rev., 35:16 p.
Mills, D.H., 1979. Bird predation - current views. In Proceedings of the Institute of Fisheries Management Tenth Annual Study Course, 18–20 September 1979, University of Nottingham. London, Janssen Services, pp. 264–71
Mills, D.H., 1980. Heron - public enemy number 1. Fish Farmer, 3(2):16
Moerbeek, D.J., et al., 1987. Cormorant damage prevention at a fish farm in the Netherlands. Biol.Conserv., 39(1):23–38
Moller, N.W. and N.S. Olesen, 1984. The grey heron (Ardea cinerea) in Denmark. Dan.Vildtunders., 36(5–7):4–23
Moravec, F., 1984. Occurrence of endoparasitic helminths in carp (Cyprinus carpio) from the Macha Lake fishpond system (Czechoslovakia). Vestn.Cesk.Spol.Zool., 48:261–78
Mott, D.F., 1976. Methods of preventing bird predation on fish. An annotated bibliography. Denver, Colorado, Denver Wildlife Research Center, U.S. Fish and Wildlife Service, 9 p.
Mott, D.F., 1978. Control of wading bird predation at fish rearing facilities. In Wading birds, edited by A. Sprunt, J.C. Ogden and S. Winckler. Res.Rep.Audubon Soc.N.Y., 7:131–2
Muller, R., 1984. Magenuntersuchungen an graureihern Ardea cinerea. Ornithol.Beob., 81:159–63
Murton, R.K., 1971. Man and birds. London, Collins, 364 p.
Needham, E. and R. Wooten, 1978. The parasitology of teleosts. In Fish pathology, edited by R.J. Roberts. London, Bailliere Tindall, pp. 144–82
Niewiadomska, K., 1963. Remarks on the discussion on certain species of the genus Tylodelphys Dies. (Trematoda, Diplostomatidae). Acta Parasitol.Pol., 11:307–13
Nilsson, S.G. and I.N. Nilsson, 1976. Numbers, food consumption and fish predation by birds in Lake Mockeln, southern Sweden. Ornis Scand., 7:61–70
Okulewicz, J., 1984. Bird trematodes of Lower Silesia. 2. Intestinal parasites of red-throated diver (Gavia stellata) Wiad.Parazytol., 30:503–20
Olesen, N.J. and P.E. Vestergaard Jorgensen, 1982. Can and do herons serve as vectors for Egtved virus? Bull.Eur.Assoc.Fish.Pathol., 3:48
Olsson, V., 1972. Revir, biotop och boplatsval hos svenska havsornar Haliaeetus albicilla. Var.Fagelvarld, 31:89–95
Peters, F. and M. Neukirch, 1986. Transmission of some fish pathogenic viruses by the heron (Ardea cinerea). J.Fish Dis., 9(6):539–44
Pough, R.H., 1941. The fish eating bird problem at the fish hatcheries of the northeast. Trans.N.Am.Wildl.Conf., 5:203–6
Ranftl, H. and P. zur Muhlen, 1977. Graureiher (Ardea cinerea) und fischerei-vorlaufiger Versuchsergebnisse. Ber.Dtsch.Sekt.Int.Rat Vogelkd., 16:69–74
Ranson, K., 1982. Bird predation at a cage fish farm. Unpubl. B.Sc. Thesis, University of Stirling, Scotland, 90 p.
Ranson, K. and M.C.M. Beveridge, 1983. Raiders from the skies. Bird predation at a freshwater cage farm. Fish Farmer, 6(1):22–3
Roberts, R.J. and C.J. Shepherd, 1986. Hanbook of trout and salmon diseases. Farnham, Surrey, Fishing News Books, Ltd., 222 p.
Salmon, T.P. and F.S. Conte, 1982. Control of bird damage at aquaculture facilities. U.S.Fish Wildl.Serv., (475):12 p.
Salyer, J.C. II and K.F. Lagler, 1946. The eastern belted kingfisher Megaceryle alcyon alcyon (Linn.) in relation to fish management. Trans.Am.Fish.Soc., 76:97–117
Scanlon, P.F., L.A. Helfrich, and R.G. Stultz, 1978. Extent and severity of avian predation at federal fish hatcheries in the United States. Proc.Conf.Southeast.Assoc.Game Fish Wildl.Ag., 32:470–3
Serventy, D.L., 1938. The feeding habits of cormorants in southwestern Australia. Emu, 38:293–316
Shedden, C.B., 1983. Feeding and roosting behaviour of gulls: implications for water storage. Unpubl. Ph.D. Thesis, University of Glasgow
Smith, M.W., 1968. Fertilization and predator control to increase growth rate and yield of trout in a natural lake. J.Fish Res.Board Can., 25:2011–36
Sonstegard, R.A. and L.A. McDermott, 1972. Epidemiological model for passive transfer of IPN virus by homeotherms. Nature, Lond., 237:104–5
Spaans, A.L., 1971. On the feeding ecology of the herring gull (Larus argentalus Pont.) in the northern part of the Netherlands. Ardea, 59:73–188
Spanier, E., 1979. Experiments in repelling night herons from fishponds by the use of distress calls. Bamidgeh, 31:11–24
Spanier, E., 1980. The use of distress calls to repel night herons (N.nycticorax) from fishponds. J.Appl.Ecol., 17:287–94
Stead, D.G., 1954. The role of the cormorant or shag in a state of nature. Aust.Wildl., 2:40–8
Taylor, R.L. and M. Lott, 1978. Transmission of salmonid whirling disease by birds fed trout infected with Myxosoma cerebralis. J.Protozool., 25:105–6
Tjomlid, S.A., 1973. Food preferences of the pied kingfisher, Ceryle rudis.Ornis.Scand., 4:145–51
Utschick, H., 1983. Abwehrstrategie und Abwehrmassnakem gegen den Graureiher (Ardea cinerea) an Fischgewassern. Garm. Vogelkd.Ber., 12:18–58
van Dobben, W.H., 1952. The food of the cormorant in the Netherlands. Ardea, 40:1–63
van Vessem, J., D. Draulans and A.F. de Bont, 1985. The effect of killing and removal on the abundance of grey herons at fish farms. In Proceedings of the Seventeenth Congress of the International Union of Game Biologists, Brussels, 17–21 September 1985, pp. 337–43
Vasvari, M., 1954. Food ecology of the common heron, the great white egret and little egret. Aquila, (55–58):23–38
Vernon, J.D.R., 1972. Feeding habitats and food of the black-headed and common gulls. Bird Stud., 19:173–86
Vlug, J.J., 1974. Zom enconcentraties van der Fuut Podiceps cristatus aan ze ande riese kust tussen Staveren en De Lemmer. Limosa, 47:16–22
Vollonweider, R.A., 1968. Scientific fundamentals of the eutrophication of lakes and flowing waters, with particular reference to nitrogen and phosphorus as factors in eutrophication. Paris, OECD Report DAS/CSI/68.27:182 p.
West, B., D. Cabot and M. Greer-Walkob, 1975. The food of the cormorant (P.carbo) at some breeding colonies in Ireland. Proc.R.Irish Acad., 75:285–305
White, H.C., 1957. Food and natural history of mergansers on salmon waters in the maritime provinces of Canada. Bull.Fish Res.Board Can., (116):63 p.
Willgohs, J.F., 1961. The white-tailed eagle (Haliaeetus albicilla) in Norway. Univ.Bergen Arb.(Mat.-Nat.Serv.), 12:1–212
Williams, B.M., D.W. Richards and J. Lewis, 1976. Salmonella infection in the herring gull (Larus argentatus). Vet.Rec., 98:51
Willomitzer, I., 1980. Seasonal dynamics of parasitozes in grass carp (Ctenopharyngdon idella) fry and fingerlings. Acta Vet., Brno, 49:269–77
Wilson, M.F. and J.C. Harmeson, 1973. Seed preferences and digestive efficiencies of the cardinals and song sparrows. Condor, 75:225–34
Wooten, R., 1973. The metazoan parasite fauna of fish from Hanningfield Reservoir, Essex, in relation to the features of the habitat and host populations. J.Zool.,Lond., 171:323–31
Wooten, R. and J.W. Smith, 1979. The occurrence of plericercoids of Diphyllobothrium spp. In wild and cultured salmonids from Loch Awe area. Scott.Fish.Res.Rep., (13)
Wooten, R., 1980. Studies on the parasite fauna of juvenile Atlantic salmon, Salmo salar L., cultured in fresh water in eastern Scotland. Z.Parasitenkd., 63:221–31
Woynarovich, E., 1980. Raising ducks on fishponds. ICLARM Conf.Proc., (4):129–34
Zitnan, R. and M. Cankovic, 1970. Comparison of the epizoological importance of the parasites of Salmo gairdneri irideus in the two coast areas of Bosna and Herzegovina. Helminthologia, 11:161–6
![]()
![]()
![]()