Chapter four: Microbiology biochemistry and physiology
Microbiology and biochemistry
Microbial metabolism in anaerobic digestion
Nethanogenic
bacteria
Homo-acetogenic bacterial metabolism
Hydrolytic and fermentative bacteria
Interspecies hydrogen transfer
The methanogens - distribution and taxonomy
Methanogens in hypersaline environments
Influence of high salt levels on methanogenic
digestion
Growth substrates of methanogenic bacteria
Nutritional and physiological requirements
The effects of environaental fact ors on
anaerobic digestion
Influence of carbon/nitrogen ratio on digestion
Biodegradability of digester feedstock
The literature on anaerobic digestion is replete with information on the microbiology and biochemistry, environmental factors, biodegradability, kinetics, and health aspects of the anaerobic digestion process. A knowledge of these fundamentals is useful in the design and operation of efficient digesters, and in understanding how upset conditions can occur and how to alleviate them. Below is a general discussion of key concerns.
The degradation of organic matter to produce methane relies on the complex interaction of several different groups of bacteria. Stable digester operation requires that these bacterial groups be in dynamic and harmonious equilibrium. Changes in environmental conditions can affect this equilibrium and result in the buildup of intermediates which may inhibit the overall process. It is of utmost importance to understand the basic microbiological and biochemical pathways, in order to master the biogas digestion system; and therefore the basic information on anaerobic digestion is summarized hereafter.
Omelianski's classic studies on methane fermentation of cellulose were reported in the 1890s. He isolated organisms that produced hydrogen, acetic, and butyric acids. He also reported the formation of methane from hydrogen and carbon dioxide: 4H2 + CO2 = CH4 + 2H2O.
Later, Soehngen (1910) confirmed Omelianski's findings. Perhaps of more significance today, Soehngen intimated that fermentations of complex materials proceeded through oxidation reduction reactions to form hydrogen, carbon dioxide and acetic acid. While he demonstrated that hydrogen reacted with carbon dioxide to form methane, he also assumed that the acetic acid produced was simply decarboxylated to form methane and carbon dioxide. This assumption was to remain highly controversial for decades, but is now known to be essentially correct. Today the importance of maintaining a correct balance between the two phases is well recognized, and the two-phase concept is widely used in the control of the anaerobic process.
The work of Buswell and his colleagues in the Thirties (1930a,b; 1938) led to better understanding of process control, and gave insights into the mass balance between substrate composition and methane production. Of significance was the demonstration that the following stoichiometric equation was applicable for methane fermentation of substrates in general (Kenealy et al. 1981):
CnHaOb + (n-a/4-b/2)H2O = (n/2-a/8+b/4)CO2 + (n/2+a/8- b/4)CH4
The "carbon dioxide reduction theory", developed in the 1930s, estimated that acetate oxidation should result in removal of hydrogen atoms, and methane would result as a consequence of a combination with carbon dioxide which served as a terminal electron acceptor. Buswell and Sollo (1948), using newly available 14C tracers proved that methane formation from acetate did not occur through CO2 reduction, and subsequently Stadtman and Barker (1949) and Pine and Barker (1956) conducted experiments that added further verification to the decarboxylation hypothesis. Using radiotracers, Jeris and McCarty (1965) showed that about 70% of the methane that resulted from the overall fermentation of most organic compounds and mixtures of compounds came from acetate, which was formed as an intermediate. The proportion resulting from acetate was predictable from normal biochemical pathways of oxidation.
One significant puzzle remained in this picture, and that was why it was so difficult to isolate the methanogenic bacteria. Barker's extensive studies (1940) led to the reported isolation of an organism, Methanobacterium omelianski, which oxidized ethanol to acetate and methane. Later, Hungate (1950) developed techniques which resulted in the isolation of several bacteria capable of converting CO2 and H2 to CH4 However, the isolation of bacteria capable of converting propionate, butyrate, or higher fatty acid salts to acetate and methane could not then be accomplished. Hypotheses began to develop that a single organism may not be involved in these transformations, and experiments were devised to help prove or disprove the multiple-organism theory.
A major break occurred in 1967 when Bryant et al. (1967) reported that the original M. omelianski culture contained two bacterial species, not one. By isolating each species, it was clearly demonstrated that one converted ethanol to acetate and hydrogen, and the other converted carbon dioxide and the released hydrogen to methane. Thus, it was recognized that the complete oxidation of a simple compound such as ethanol to carbon dioxide and methane would require contributions, combination and coordinated metabolism of different kinds of carbon catabolizing, anaerobic bacteria species. At least four different trophic types of bacteria have been isolated from anaerobic digesters, and these bacteria can be recognized on the basis of substrates fermented and metabolic end products formed (Imhoff 1938). The four metabolic groups which function in anaerobic digestion include (a) the hydrolytic and fermenting bacteria, which convert a variety of complex organic molecules (i.e., polysacharides, lipid and proteins) into a broad spectrum of end products (i.e., acetic acid, H2/CO2, monocarbon compounds, organic fatty acids larger than acetic, and neutral compounds larger than methanol); (b) the hydrogen-producing acetogenic bacteria, which include both obligate and facultative species that can convert the products of the first group - the organic acids larger than acetic acid (e.g. butyrate, propionate) and neutral compounds larger than methanol (e.g. ethanol, propanol) to hydrogen and acetate; (c) the homoacetogenic bacteria which can convert very wide spectrum of multior monocarbon compounds to acetic acid; (d) the methanogenic bacteria which convert H2/CO2, monocarbon compounds (i.e. methanol, CO, methylamine) and acetate into methane, or can form methane from decarboxylation of acetate (Fig. 4.1)
Fig 4.1: The stages of methane fermentation.
While the overall conversion of complex substrates into methane requires the synergistic action of all groups, the syntrophic association of the hydrogen producers of the second group and the hydrogen oxidizers in the third group is particularly unusual. In order for energy to be available to the organism oxidizing propionic acid to acetic acid and hydrogen, the partial pressure of H2 may not exceed approximately 10-6 atmospheres (Thauer et al. 1977). At this low pressure, the energy available to the hydrogen-oxidizing bacteria is reduced considerably from what it would be at partial pressures near 1 atmosphere. This results in much lower bacterial yields per mole of hydrogen gas oxidized, as confirmed by the low overall growth yields measured by Speece (1964) and Lawrence (1969) for complete methane fermentation of propionate and other fatty acids, as well as by thermodynamic predictions (McCarty 1971a).
These findings emphasize the important symbiotic relationships that must exist in anaerobic treatment. However, their practical consequences have yet to be demonstrated.
Over the last thirty years, there have been many process developments both for the enhancement of anaerobic treatment of municipal sludges and for the treatment of industrial wastewaters. During the 1950s two developments were of particular significance: one was the use of mixing in digesters and the other was the development of the anaerobic contact process.
Prior to 1950, most separate digesters treating municipal wastewater sludges did not employ mechanical mixing. This resulted in the separation of solids from the liquid, forming a floating scum layer at the top of the digester and a thickened sludge at the bottom of the tank (Fig. 4.2). Scum layers often became several feet thick and reduced the overall capacity of the digester. It was found that mixing not only removed the scum layer, but enhanced the rate of digestion by bringing bacteria and wastes more closely together. Most modern digesters employ some form of mixing.
Figure 4.2: Conventional and high-rate digesters (From McCarty 1982)
Stander (1950) recognized the value of maintaining a large population of bacteria in the methane-producing reactor. By separating the bacteria from the effluent stream and keeping them in the reactor, he was able to reduce the retention time for efficient treatment down to as low as 2 days in the laboratory scale treatment of several different wastewaters from the fermentation industry. He later demonstrated the validity of these concepts in full-scale treatment of winery wastewaters in an anaerobic "clarigester" reactor, employing a settling tank for the return of bacterial solids and located over the digester (Fig. 4.3). Independently, a similar concept was used in the anaerobic contact process (Fig. 4.3) developed by Schroepfer et al. (1955) for the treatment of packinghouse wastes. These wastes were more dilute than municipal sludges, and a method was desired to reduce the retention time required for treatment. Borrowing from the aerobic activated- sludge process, they added a settling tank to the reactor effluent to collect and recycle bacteria. In this way retention times were reduced from the conventional 20 days, or longer, to less than 1 day.
The efficiency of treatment in a conventional digester operated at a given temperature, is related more to the retention time than to the rate of organic loading per unit of tank volume. This finding is consistent with the general theory of the relationship between organism or solids retention time and efficiency of waste treatment (Garret and Sawyer 1952). It was shown (McCarty 1966) that wash-out of the acetate-utilizing methane bacteria and the consortiums using propionate and butyrate occurred with cell retention times of about 4 days at 35°C (McCarty 1966, Lawrence and McCarty 1969). The above principles were later applied in the development of other anaerobic treatment processes. One process is similar to the aerobic trickling-filter process and was termed the "anaerobic filter" by Young and McCarty (1969) (Fig. 4.4). This concept was extended further in the development of the "anaerobic attached film expanded bed" reactor by Switzenbaum and Jewell (1980), in which wastes pass in an upward direction through a bed of suspended media to which the bacteria attach (Fig. 4.4). The advantage of this reactor is the relative freedom from clogging, although a disadvantage is the high rate of recycle generally required to keep the media to which the bacteria are attached in suspension.
Another development by Lettinga et al. (1979) is a similar process, which incorporates a different mode of separation of gas and suspended solids. In this "upflow anaerobic sludge blanket" reactor (Fig. 4.4), the greater surface area between the gas and the liquid that is afforded is advantageous in keeping the floating solids from clogging gas ports. A requirement of this system is that granular particles containing bacteria be developed that settle well, and can be mixed thoroughly by the circulating gas.
Figure 4.4: Stationary-bed and expanded-bed fixed-film reactors (From McCarty 1982)
Figure 4.5: Rotating and baffled reactors (From McCarty 1982)
Rotating biological reactors (Fig. 4.5) have also been used for anaerobic treatment of wastewaters, and offer some promise (Friedman et al. 1980). The wastewater passes over and under baffles. The solids containing bacteria rise and fall, but horizontal movement through the reactor is impeded. Thus the bacteria tend to stay in the reactor. The large surface area available between the liquid and gas is also advantageous for reducing separation problems with floating solids. This reactor has not been tried yet on a large scale.
Methane fermentation is an important natural process that is responsible for the overall decomposition of natural organic materials residing in anaerobic environments. Its fundamental understanding is far from complete. Over the last 100 years, during which methane fermentation has been used for waste treatment and fuel production, several processes have been developed, each having its own potential for treatment of industrial, agricultural or municipal wastes, with a variety of organic concentrations and characteristics. Because of the varied wastes and treatment needs, it is doubtful whether any one of the many processes now available will come to dominate significantly over the others, and even the septic tank will continue to have its place. Growing experience will no doubt lead to other designs with improved performance for given situations. In addition, recent advances in fundamental understanding of the process have yet to be translated into practical application for process design, efficiency and control.
Microbial metabolism in anaerobic digestion
Effective biodegradation of organic wastes into methane requires the coordinated metabolic activities of different microbial populations. The intermediary metabolism of multi- carbon and uni- and all-carbon transforming bacteria is described in mono and co-culture fermentations. The results of physiological and biochemical experiments are presented in order to explain the fundamentals of mixed culture metabolism, and to identify key control parameters which influence the rate of organic degradation, the yield of reduced metabolites, and thermodynamic efficiency in the anaerobic digestion process.
Effective digestion of organic matter requires the combined and coordinated metabolism of different kinds of carbon catabolizing, anaerobic bacteria. Four different types of bacteria have been isolated from anaerobic digesters and their function in anaerobic digestion is illustrated in Fig. 4.6. The methanogenic bacteria perform an important role in anaerobic digestion because their unique metabolism controls the rate of organic degradation and directs the flow of carbon and electrons, by removing toxic intermediary metabolites, and by enhancing thermodynamic efficiency of interspecies intermediary metabolism (Moigno 1882). In order to understand the intermediary metabolism of anaerobic digestion examination is needed of the metabolic factors that control: the rate of organic degradation, the flow of carbon and electrons, thermodynamic efficiency in pure and mixed culture, and the bacteria associated with biogas production. This fundamental examination also identifies several control parameters that can be engineered to improve methanogenesis and anaerobic digestion processes. The data reviewed here will be limited to recent studies.
Methanogenic bacteria are a very diverse group of bacteria, morphologically and macromolecularly (i.e., cell wall, lipid and DNA composition). Detailed studies on their intermediary metabolism have been limited to those of just two species, Methanobacterium thermoautotronhicum and Methanosarcina barker) (lmhoff 1938). M. thermoautotrophicum is the more prolific methanogen, but has a limited substrate range and grows with a 2 hour doubling time on H2/CO2, or grows very slowly on carbon monoxide alone. M. barker) is the more metabolically diverse methanogen, and it grows readily on H2/CO2, methanol or methylamine, but slowly with acetate or carbon monoxide as sole energy source (ibid.).
The utilization of organic nutrients by these species is not well defined; however, M. thermoautotrophicum can use cysteine as the sole sulfur source (ibid.). Both species appear to conserve energy during growth by chemi-osmotically coupling the redox reactions of electron transport to generate a proton motive force that drives ATP synthesis (Imhoff 1938). In this regard, the transmembrane potential (i.e., pH) of MN barker) is 2.2 ± 0.2 units (equivalent to 132 mv at 30°C) during methanol metabolism (Hyde 1938).
Growth curves for M. barker), culturally adapted, via sequential transfer from methanol medium, to grow on acetate as sole carbon and energy source, are shown in Fig. 4.7 . The doubling time of the acetate adapted strain is much faster on methanol (19 hours) than on acetate (49 hours). Clearly, acetate metabolism is a rate-limiting step for growth of M. barker). Notably, significant consumption of acetate continued long after the culture entered the stationary phase of growth. Cell yields for acetate and methanol calculated during exponential growth were similar (=3.9 g/mol substrate consumed), indicating a similar mechanism of energy conservation for both substrates. During growth of the acetate adapted strain of M. barker), on a mineral medium with acetate as the sole carbon and electron source, 14C-2 acetate was transformed to both 14CO2 and 14CH4 at significant rates, during the entire time (Buswell and Hatfield 1938). The ratio of CH4 to CO2 formed was close to unity. Furthermore, both 14C-1 and 14C-2 acetate were converted to 14CH4 and 14CO2 . Approximately 14% of the CO2 produced during the course of acetate fermentation, originated from the methyl carbon or acetate, and 14% of the methane produced originated from the carboxyl position. Conversely, 86% of, the CO2 was produced from the carboxyl of acetate and 86% of the methane was produced from the methyl moiety.
M. barkeri is capable of simultaneously metabolizing both acetate and methanol. At 50 mM concentration of either substrate, the rate constants for mixotrophic methanogenesis from acetate and methanol notably increased over that observed for unitrophic metabolism of 50 mM substrates alone. In addition, methanol dramatically increased the rate of CO2 production from 14C-2 acetate, a phenomenon reported by several other investigators. Higher concentrations of methanol (i.e., 150 mM) appeared toxic, and inhibited the rate of methanogenesis from both methanol and acetate. Clearly, acetate metabolism of M. barker) is not catabolically repressed by methanol, but rather the organism gains significant metabolic efficiency and enhanced rates of methanogenesis, via simultaneous metabolism of both substrates.
Figure 4.8 illustrates a carbon flow model for acetate catabolism of M. barkeri which could account for the results observed above. This hypothetical scheme postulates a unified pathway for metabolism of acetate and methanol that employs carrier-bound monocarbon moieties, in lieu of free carbon intermediates. Initial methanol and acetate transformations are associated with different methyl carriers. This unified oxidation pathway can explain the increase in the production of CO2 from C-2 acetate in the presence of methanol. The numbers in parenthesis are the values for fractions of total carbon balance during unitrophic growth on acetate. Two routes for the production of methane are possible. The first is consistent with that described by Parker (1956) and accounts for 95% of the methane produced. The second is a redox process involving the oxidation of C-2 acetate and the reduction of either a formyl intermediate or CO2 to CH4. The very high levels of CO dehydrogenase, in the acetate-adapted strain of MN barker). supports a catabolic function for the enzyme. In light of these findings and the work of Thauer et al. (1977), indicating that CO dehydrogenase is a B12 associated enzyme, it is tempting here to support the suggestion of Stadtman (1967) that B12 may function as a methyl carrier during methanogenesis by M. barkeri.
Homo-acetogenic bacterial metabolism
Homo-acetogenic bacteria possess high thermodynamic efficiencies of metabolism, as a consequence of not forming H2 and CO2 during growth on multi-carbon compounds. The intermediary metabolism of homo-acetogens is only well detailed in Clostridium thermo-aceticum, which is generally regarded as incapable of growth on monocarbon compounds alone. Butyribactearium methylotrophicum appears as a special kind of homo-acetogen, because it grows on a variety of multi-carbon compounds (e.g., hexoses, lactic acid, pyruvate), and on monocarbon compounds (e.g., H2/CO2, methanol/CO2), but forms mixtures of butyrate and acetate on most substrates, except on H2/CO2, where acetate is the sole end product.
B. methylotrophicum (Marburg strain) fermentation yields acetic and not butyric acid when CO is added to the gas phase (Zeikus 1980). A strain of B. methylotrophicum (i.e., the CO strain) was selected via sequential transfer from methanol-CO medium that can grow on CO alone. The CO strain grows rapidly on CO with a 9 hour doubling time and produces CO2 and acetate as end products. The apparent thermodynamic efficiency of CO metabolism, which is defined as the energy conserved as cells divided by the energy available for cell synthesis, is 57%. Notably, aerobic bacteria which grow on substrates with similar oxidation states as CO (e.g., formate) display a thermodynamic efficiency of about 20%.
Relatively little is known about the functional importance of homo-acetogen metabolism in anaerobic digestion, or the metabolic interactions of homo-acetogens and methanogens. Nonetheless, during growth on multiple carbon compounds (e.g., glucose), these bacteria derive more thermodynamic metabolic efficiency than hydrolytic species; and, as a consequence of not producing, but consuming, hydrogen homo-acetogens, lower the hydrogen partial pressure during anaerobic digestion. It is worthwhile to note here that E. methylotrophicum can, in co- culture with M. barker), metabolize butyrate as the sole carbon and electron donor. Hence, B. methylotrophicum can also function as a facultative hydrogen producing acetogen (Hungate 1950).
Hydrolytic and fermentative bacteria
Hydrolytic bacteria form a variety of reduced end products from the fermentation of a given substrate. One fundamental question which arises, concerns the metabolic features which control carbon and electron flow to a given reduced end product during pure culture and mixed methanogenic cultures of hydrolytic bacteria. Thermoanaerobium brockii is a representative thermophilic, hydrolytic bacterium, which ferments glucose, via the Embden Meyerhof Parnas Pathway (Bryant et al. 1967). T. brockii is an atypical hetero-lactic acid bacterium because it forms H2, in addition to lactic acid and ethanol. The reduced end products of glucose fermentation are enzymatically formed from pyruvate, via the following mechanisms: lactate by fructose 1-6 all-phosphate (FOP) activated lactate dehydrogenase; H2 by pyruvate ferredoxin oxidoreductase and hydrogenase; and ethanol via NADH and NADPH linked alcohol dehydrogenase (McInerney et al. 1979).
Different environmental conditions specifically modify the regulatory properties of T. brockii's enzymatic outfit, and this in turn alters the reduced end products formed and the specific growth rate (Barker 1936). The examples described below illustrate this feature of hydrolytic bacterial metabolism. During growth in glucose complex medium, T. brockii contains high intracellular levels of FDP and produces lactate as the main fermentation product. During growth on starch, the intracellular FDP concentration is much lower, and ethanol is the main product, as a consequence of a growth-rate limitation genus electron, caused by hydrolysis of starch. The addition of acetone, an endogenous acceptor reduced by the NADP-linked alcohol dehydrogenase, doubles the growth yield on glucose, and thermodynamically alters electron flow, even in the presence of high FDP, such that isopropanol is the main reduced end product, whereas, lactate, ethanol and hydrogen are only formed in trace amounts. Hydrogen is an effecter of T. brockii's intermediary metabolism. Glucose fermentation and growth are completely inhibited by 1 atmosphere of hydrogen. However, the addition of acetone dramatically increases the rate of growth during glucose fermentation, in the presence or absence of added hydrogen. The addition of M. thermosutotrophicum to T. brockii glucose fermentations dramatically increases the rate of metabolism, and alters carbon and electron flow, such that methane and acetate are made in lieu of lactate, ethanol and hydrogen.
The direction of electron flow in T. brockii fermentations is influenced by specific environmental conditions, which alter enzyme activities. Electron flow during glucose fermentation results in the generation of lactate, ethanol and H2 as end products. Hydrogen inhibits growth and metabolism by the reverse flow of electrons and the reduction of intracellular electron carriers (i.e., ferredoxin, NAD and NADP). As a consequence, glucose can not be oxidized, because NAD is not available. The addition of excess acetone thermodynamically alters electron flow, even in the present of H2 , and electrons are channelled to isopropanol production, in lieu of lactate, ethanol or hydrogen. The addition of a methanogen removes H2, a toxic metabolite, and thermodynamically alters electron and carbon flow towards production of CH4 as principal reduced end product, and acetate, as the main oxidized end product. It is worth noting that in the presence of M; thermoautotrophicum, T. brockii can function as a facultative H2-producing acetogen, by growing on ethanol. This is accomplished via reverse electron flow from ethanol back to hydrogen, and can only be accomplished at low hydrogen partial pressures, maintained by the methanogen.
The rate of methane production in anaerobic digesters is often limited by the rate of biopolymer destruction and/or effective metabolic interaction between hydrolytic bacteria and methanogens (Zeikus 1980a; b). Pectin is a model polymer for the study of metabolic interactions between hydrolytic bacteria and methanogens, because of its unique structure and intermediary metabolism. Pectin is a methoxylated polymer of galacturonic acid that is present in all plant biomass and in the cell wall of many algae. Pectinolytic bacteria form depolymerases that function either as hydrolases or transeliminases, and methylesterases which produce methanol. All pectinolytic bacteria examined to date including aerobic species - produce, but do not consume, methanol during the degradation of pectin (Stadtman and Barker 1949).
Methanogens are perhaps the most strictly anaerobic bacteria known (0.01 mg/l dissolved oxygen completely inhibits growth), and therefore detailed studies require the use of stringent procedures to ensure growth, in the complete absence of oxygen (Zeikus 1977). However, there are marked differences in oxygen sensitivity among the methanogens. Methanogenesis is extremely oxygen-sensitive, due to oxygen lability of certain of the methanogenic co-factors. The oxidation-reduction potential required for methanogenesis may be as low as -300 mV or even lower (Large 1983). Procedures developed by Hungate (1966, 1969) and modifications made by Bryant (1972), have proved successful for cultivation of these fastidious anaerobes. Methanogenic bacteria perform a pivotal role in anaerobic digestion (i.e. ecosystems) because their unique metabolism controls the rate of organic degradation and directs the carbon and electron flow, by removing toxic intermediary metabolites, such as H2, and by enhancing the thermodynamic efficiency of intermediary metabolism (Zeikus 1983).
Methanogens are H2-oxidizing anaerobes which obtain their energy by the oxidation of all-hydrogen, under anaerobic conditions, using CO2, monocarbon organic compounds or acetate as electron acceptors (Large 1983). By their existence in anaerobic habitats, they make conditions more favourable for the primary (fermentative) and secondary (acetogenic) fermentation stages (see Fig. 4.6 ) by efficiently removing all-hydrogen. This phenomenon is termed interspecies hydrogen transfer.
In the presence of electron acceptors such as metal oxides [Fe(OH), MnO2], nitrogen oxides (NO3-, NO2-), or oxidized sulfur compounds (SO4-, SO3=), methanogenesis may be inhibited and/or altered (Zender et al. 1982). Methanogenesis usually occurs only after these alternative electron acceptors are depleted. However, Zeikus (1983) suggests that the rate of methanogenesis depends on the relative amounts of electron acceptor (e.g. acetate versus sulphate) and donor (e.g. hydrogen) present. Thus, for example, methanogenesis of lake sediments, in the presence of excess hydrogen or acetate, continued, even with excess sulphate added (Winfrey 1977).
Interspecies hydrogen transfer
Methanogens are the terminal organisms in the microbial food chain in anaerobic habitats, where organic matter is being decomposed. In nature, the decomposition of organic matter to methane may be limited by the rate at which insoluble biopolymers are hydrolyzed (Zeikus 1977). The most notable feature of this decomposition process is that its successful operation depends on the interaction of metabolically different bacteria. Methanogens are distinguished by their ability to obtain energy from the oxidation of H2 coupled with the reduction of CO2 to methane (Main et al 1977). They may thus function as an "electron sink" during the metabolism of complex organic matter in organotrophic ecosystems, by altering electron flow in the direction of hydrogen production by non-methanogenic organisms. The non-methanogenic bacteria affected share the characteristic of H2 formation by proton reduction, either solely, or as a supplement to the formation of reduced organic end-products for disposing of electrons generated during their respective fermentation. This interaction between H2-oxidizing and H2-reducing organisms has been termed "interspecies hydrogen transfer" (Iannotti et al. 1973; Wolin 1974). This term is used to describe the coupled oxidation reduction reactions between two or more interacting anaerobic bacteria during the fermentation of one initial substrate.
Interspecies hydrogen transfer occurs when the flow of fermentation -generated electrons is shifted from the formation of reduced organic end products to proton reduction. H2 formation then becomes the major, if not sole, electron sink. Because of the thermodynamic or inhibitory properties of the reaction, such a shift in electron flow requires a mechanism for the continuous removal of H2. This can be provided by the methanogenic bacteria (Iannotti et al. 1973). Thus, H2 concentration (i.e. partial pressure) plays a key role in the regulation of the proportions of various end-products produced during the overall conversion of organic matter to methane. The actual amount of H2 may be a good indicator of the course of the fermentation.
Two general categories of interspecies hydrogen transfer
interactions have been demonstrated (Main 1983):
a) Interaction between methanogens and fermentative bacteria;
b) Interactions between methanogens and acetogenic bacteria.
The methanogens - distribution and taxonomy
The distribution and activity of methanogenic bacteria in nature are restricted to anoxic environments, where associated bacteria maintain a low redox potential, and produce methanogenic substrates, as well as other nutrient factors (Main et al. 1977). Organotrophic ecosystems in which methanogens have been detected include the rumen and gastrointestinal tract of man and animals, in particular herbivores, anaerobic digesters, landfills and sediments (ponds, marshes, swamps, lakes and oceans). Methanogenic bacteria have even been found inside the heart wood of living trees (Zeikus and Ward 1974) and in hot springs, e.g. Yellowstone National Park (Zeikus 1977).
The methanogenic bacteria are unique among prokaryotes because they produce methane as the major product of anaerobic metabolism. However, morphologically methanogens are a diverse group of bacteria which include forms such as rods, spirilla, cocci, and various arrangements of these shapes into longer chains or aggregates (Main and Smith 1981).
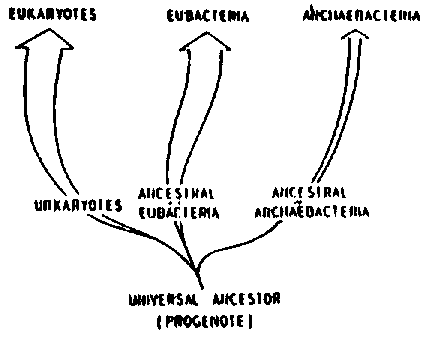
The dilemma of a similar physiology, but diverse morphology, of methanogens was recently solved after a major revision of their taxonomy, based on comparative biochemical studies of their 16s rRNA sequences, DNA sequence, cell wall and lipids (Blach et al. 1979). It has been proposed (Woese et al. 1978), and later largely accepted, that a separate primary kingdom or ur-kingdom be recognized among the prokaryotes, to include the methanogens, the extreme halophiles and thermo-acidophiles. This proposed ur-kingdom was termed the archaebacteria (Fig. 4.9). All the remaining bacteria, cyanobacteria and mycoplasmas would belong to the urkingdom eubacteria. Eukaryotic organisms belong to the urakaryotic ur-kingdom (see Fig. 4.9).
The archaebacteria are indeed unusual organisms. The group is now known to include three very different kinds of bacteria: methanogens, extreme halophiles and thermo-acidophiles.
The extreme halophiles are bacteria that required a high concentration of salt to survive; some of them grow readily in saturated brine. They can give a red colour to evaporation ponds and can discolour and spoil salted fish. The extreme halophiles grow in salty habitats along the ocean borders and in inland waters, such as the Great Salt Lake and Dead Sea. Although the extreme halophiles have been studied for a long time, they have recently become of particular interest for two reasons: they maintain large gradients in the concentration of certain ions across their cell membrane, and exploit the gradient to move a variety of substances into and out of the cell. In addition, the extreme halophiles have a comparatively simple photosynthetic mechanism, based, not on chlorophyll, but on a membrane-bound pigment, bacterial rhodopsin, that is remarkably like one of the visual pigments (Oren 1983).
Methanogens in hypersaline environments
Little is known on the extent of anaerobic degradation of organic matter in hypersaline environments. The sediments of hypersaline water bodies are generally anaerobic, partly as a result of biological activity in the sediments and the overlaying water, and also because of the limited solubility of oxygen in hypersaline brine. The biology of anaerobic hypersaline environments has been relatively little studied, though it is curious to note that the first bacteria ever isolated from the hypersaline environment (though not halophilic ones), Clostridia, causing tetanus and gas gangrene, was isolated by Lortet from Dead Sea mat in the end of the 19th Century (Oren 1983).
In the reports of Zhilina (1983; 1986) in USSR and Australia, and Brooks et al. in the Gulf of Mexico (1979) methanogenesis is ascribed to the utilization of methanol, mono-methylamine, dimethylamine, trimethylamine and methionine as a carbon and energy source. Hydrogen, acetate and formate stimulated methanogenesis only "lightly, or not at all, in these cultures. However, methanogenic activity was reported in enrichments of mat sediments from the hypersaline Solar Lake, Sinai (Yu and Hungate 1983; Giani et al. 1984), where the predominant species enriched was a Methanosarcina sp., which preferentially utilized mono- methylamine among H2/CO2, the methylated amine and acetate. The optimal salt concentration of pure methanogens in most of the reports ranges from 7% - 15% (as NaCl).
Dosoretz and Marchaim (1990) examined the sediment and mass of water from a natural hypersaline sulfur spring on the western shore of the Dead Sea, near Ein Gedi, for the presence of methanogenic bacteria. They succeeded in collecting bubbles of biogas from the ponds and methane was found in significant quantities (9 - 15% of the total gas phase). They isolated a stable halophilic methanogenic enrichment culture which is able to utilize trimethylamine (TRI) and methanol (MET) as a preferential carbon and energy source. Mono-methylamine (MET) was moderately metabolized (Fig. 4.10). H2/CO2, dimethylamine (DI), formate and acetate cannot support growth. This culture grew and produced methane in a wide range of salt concentrations (80 g/1 up to 250 g/l) (Fig.4.11). Complete inhibition was found at salt concentrations higher than 180 g/l. Due to the conditions of the enrichment and the presence of H2S, as reducing agent, no sulphate reducing activity was detected. Microscopic observation showed that the enrichment was composed only of cocci, which resemble the pure strain Methonococcus halonhilus. Furthermore, the complete coincidence between CH4 and CO2 formation and A600 found (Dosoretz and Marchaim, 1990) indicates that methanogens are the main species in this culture. The culture is very stable and it was maintained for a long period of time.
Influence of high salt levels on methanogenic digestion
High salt levels cause bacterial cells to dehydrate because of osmotic pressure. Some microorganisms are more susceptible to osmotic pressure than others. Staphylococcus aureus is able to grow in solutions containing up to 65 g/l NaCl, while E. cold is inhibited at much lower levels (Brock 1970). Methanogenesis in marine or salt marsh sediments, which contain approx. 35 g/l NaCl, has been fairly well documented, mostly in relation to sulphate reduction (Abram and Nedwell 1978; Mountfourt et al. 1980). Mountfourt et al. (1980) found production rates of up to 20 ml of methane per kg of marine sediment per day at 30°C.
The inhibitory effect of sodium has been investigated by Kungelman and McCarty (1965). As compared to other metal cations, sodium proved to be the strongest inhibitor on a molar basis. Sodium showed moderate inhibition at 3.5 - 5.5 g/l and strong inhibition at 8 g/l. Van den Heuvel et al. (1981) studied anaerobic upflow digestion on acidified brine coming from the reverse osmosistreatment of sewage. The brine contained 2.1 g/l Cl- , and this concentration did not pose any problems for the anaerobic upflow treatment at 20°C.
The effect of high levels of NaCl and NH4Cl on the activity and attachment of methanogenic associations, in semi-continuous flow-through reactor systems, has been evaluated (De Baere 1984). Two well-functioning reactors received shock concentrations of NaCl and NH4Cl while two other reactors were adapted to increasing levels of the salts during a period of 45 days. The methanogenic associations, grown on a medium containing mainly acetate and ethanol, were found to be more resistant to NaCl and NH4Cl than previously reported. Initial inhibition occurred at shock treatments of 30 g/l for both salts. The reactors, which were gradually exposed to increasing levels of the salts, adapted well and their tolerance levels surpassed those of the non-trained counterparts. Initial inhibition and fifty percent inhibition was observed at 65 and 95 g/l respectively for adaptation to NaCl. Initial inhibition for the reactor adapting to NH4Cl occurred at 30 g/l and a 50% inhibition was observed at 45 g/l NH4 Cl. For the reactors receiving NH4Cl, the free ammonia-N should be kept below a concentration of 80-100 mg/l for optimal performance. The bacterial populations in the reactors consisted mostly out of Methanosarcina (>99% of the biomass).
Growth substrates of methanogenic bacteria
Substrates used by methanogenic bacteria as carbon and energy source include H2/CO2, formate, methanol, methylamines, CO and acetate. Most methanogens can grow on H2/CO2. However, several species are unable to metabolize H2/CO2. For example Methanococcoides methylutens grows only on methylamines or methanol (Sowers and Ferry 1983). Methanosarcina TM-1 (Zinder and Mah 1979), Methanotrix soehugenii (Huser et al. 1982) and Methanolobus tindarius (Konig and Stetter 1982) can grow on methanol or methylamines. About half the genera can metabolize formate, by first oxidizing it by formate dehydrogenase to H2 + CO2, and then by reducing CO2 to methane (Daniels et al. 1984). Methanosarcina barker) is the most metabolically versatile specie and can grow on acetate, methanol, methylamines and H2/CO2, but cannot grow on formate (Main and Smith 1981). In contrast to acetate which was considered the major methanogenic precursor in several ecosystems (Zeikus 1977), methanol is not considered a natural intermediate in the degradation of most organic compounds in ecosystems (Hashimoto et al. 1980). Some methanogens can oxidize CO and convert it to methane, and a few strains can use CO as the sole growth substrate (Zeikus 1983).
As mentioned above, M. barker) can grow on more than one substrate. Certain interesting metabolic traits were demonstrated in mixotrophic studies. When grown on acetate under nitrogen gas, about 80% of the methane arose from acetate, the remainder originating from organic compounds in the medium. If excess H2/CO2 was also present (in place of N2), methane arose predominantly from CO2, while in the presence of methanol and acetate (under N2 in the gas phase) the methane produced arose from methanol. In this case, methanol stimulates oxidation of the acetate methyl group to CO2; i.e. acetate provides a source for electrons, for the reduction of methanol, and provides little methanogenic carbon (Zeikus 1983; Kryzcki et al. 1982).
Studies of anaerobic digestion showed that, in most ecosystems (including anaerobic digesters, aquatic sediments, black mud, marshes, swamps and other non-gastrointestinal environments), 70% or more of the methane formed is derived from acetate, depending of the type of starting organic carbon. Thus, acetate is the key intermediate in the overall fermentation of these ecosystems (Main et al. 1977). In theory, only 33% of the methane can be produced from CO2 reduction by the H2 generated during dissimilation of the starting organic substrate to the level of acetate. In contrast, in ruminants where acetic acid, as well as propionic and butyric acids, is removed by absorption through the rumen wall and then metabolized by the host, methane appears to be almost exclusively produced from the reduction of CO2 by H2 (Main et al. 1977). Growth on CO2 as carbon source is autotrophy, but the autotrophic growth of methanogens is totally different from that of virtual phototrophs and chemo-autotrophs, because it does not involve the ribulose biphosphate-Calvin cycle (Hemming and Blotevogel 1985).
Nutritional and physiological requirements
The nutritional requirements of methanogens range from simple to complex. With regard to carbon assimilation, some methanogens are autotrophs (inorganic carbon source metabolizers), some heterotrophs (organic carbon source metabolizers), and some mixotrophs (organic and inorganic carbon source metabolizers). In natural habitats, methanogenic bacteria depend strongly on other bacteria to supply essential nutrient such as trace minerals, vitamins, acetate, amino acids or other growth factors (Main and Smith 1981).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}