![]()
![]()
![]()
The role of iron in human metabolic processes
Iron requirements
Iron absorption
Iron deficiency
Iron during pregnancy and lactation
Iron supplementation and fortification
The evidence for estimating the recommended nutrient intake for iron
Recommendations for iron intake for infants, children, younger and older adults, and pregnant and lactating women
Future research
References
Iron has several vital functions in the body. It serves as a carrier of oxygen to the tissues from the lungs by red blood cell haemoglobin, as a transport medium for electrons within cells, and as an integrated part of important enzyme systems in various tissues. The physiology of iron has been extensively reviewed (1-6).
Most of the iron in the body is present in the erythrocytes as haemoglobin, a molecule composed of four units, each containing one heme group and one protein chain. The structure of haemoglobin allows it to be fully loaded with oxygen in the lungs and partially unloaded in the tissues (e.g., in the muscles). The iron-containing oxygen storage protein in the muscles, myoglobin, is similar in structure to haemoglobin but has only one heme unit and one globin chain. Several iron-containing enzymes, the cytochromes, also have one heme group and one globin protein chain. These enzymes act as electron carriers within the cell and their structures do not permit reversible loading and unloading of oxygen. Their role in the oxidative metabolism is to transfer energy within the cell and specifically in the mitochondria. Other key functions for the iron-containing enzymes (e.g., cytochrome P450) include the synthesis of steroid hormones and bile acids; detoxification of foreign substances in the liver; and signal controlling in some neurotransmitters, such as the dopamine and serotonin systems in the brain. Iron is reversibly stored within the liver as ferritin and hemosiderin whereas it is transported between different compartments in the body by the protein transferrin.
Basal iron losses
Iron is not actively excreted from the body in urine or in the intestines. Iron is only lost with cells from the skin and the interior surfaces of the body - intestines, urinary tract, and airways. The total amount lost is estimated at 14 µg/kg body weight/day (7). In children, it is probably more correct to relate these losses to body surface. A non-menstruating 55-kg women loses about 0.8 mg Fe/day and a 70-kg man loses about 1 mg. The range of individual variation has been estimated to be ±15 percent (8).
Earlier studies suggested that sweat iron losses could be considerable, especially in a hot, Humid climate. However, new studies which took extensive precautions to avoid the interference of contamination of iron from the skin during the collection of total body sweat have shown that these sweat iron losses are negligible (9).
Growth
The newborn term infant has an iron content of about 250-300 mg (75 mg/kg body weight). During the first 2 months of life, haemoglobin concentration falls because of the improved oxygen situation in the newborn infant compared with the intrauterine foetus. This leads to a considerable redistribution of iron from catabolised erythrocytes to iron stores. This iron will cover the needs of the term infant during the first 4-6 months of life and is why iron requirements during this period can be provided by human milk, that contains very little iron. Because of the marked supply of iron to the foetus during the last trimester of pregnancy, the iron situation is much less favourable in the premature and low-birth-weight infant than in the term infant. An extra supply of iron is therefore needed in these infants even during the first 6 months of life.
In the full-term infant, iron requirements will rise markedly after age 4-6 months and amount to about 0.7-0.9 mg/day during the remaining part of the first year. These requirements are therefore very high, especially in relation to body size and energy intake (Table 39) (10).
Table 39
Iron intakes required for growth under the age of 18 years, median basal iron losses, menstrual losses in women, and total absolute iron requirements
|
Group |
Age |
Body weight |
Required Iron intakes for Growth
|
Basal Iron losses |
Menstrual losses |
Total Absolute Requirements † |
||
|
Mean |
Median |
Median |
95th percentile |
Median |
95th percentile |
|||
|
(Years) |
(kg) |
(mg/day) |
(mg/day) |
(mg/day) |
(mg/day) |
(mg/day) |
(mg/day) |
|
|
Children
|
0.5-1 |
9 |
0.55 |
0.17 |
|
|
0.72 |
0.93 |
|
1-3 |
13.3 |
0.27 |
0.19 |
|
|
0.46 |
0.58 |
|
|
4-6 |
19.2 |
0.23 |
0.27 |
|
|
0.50 |
0.63 |
|
|
7-10 |
28.1 |
0.32 |
0.39 |
|
|
0.71 |
0.89 |
|
|
Males
|
11-14 |
45 |
0.55 |
0.62 |
|
|
1.17 |
1.46 |
|
15-17 |
64.4 |
0.60 |
0.90 |
|
|
1.50 |
1.88 |
|
|
18+ |
75 |
|
1.05 |
|
|
1.05 |
1.37 |
|
|
Females |
11-14 b |
46.1 |
0.55 |
0.65 |
|
|
1.20 |
1.40 |
|
11-14 |
46.1 |
0.55 |
0.65 |
0.48 c |
1.90 c |
1.68 |
3.27 |
|
|
15-17 |
56.4 |
0.35 |
0.79 |
0.48 c |
1.90 c |
1.62 |
3.10 |
|
|
18+ |
62 |
|
0.87 |
0.48 c |
1.90 c |
1.46 |
2.94 |
|
|
Post-menopausal |
|
62 |
|
0.87 |
|
|
0.87 |
1.13 |
|
Lactating |
|
62 |
|
1.15 |
|
|
1.15 |
1.50 |
In the first year of life, the full-term infant almost doubles its total iron stores and triple its body weight. The change in body iron during this period occurs mainly during the first 6-12 months of life. Between 1 and 6 years of age, the body iron content is again doubled. The requirements for absorbed iron in infants and children are very high in relation to their energy requirements. For example, in infants 6-12 months of age, about 1.5 mg of iron need to be absorbed per 4.184 MJ and about half of this amount is required up to age 4 years.
Table 40
The recommended nutrient intakes for iron based on varying dietary iron bio-availabilities
|
Group
|
Age |
Mean Body weight |
|
|||
|
(years)
|
(kg)
|
|
Recommended Nutrient Intakea |
|||
|
% Dietary Iron Bio-availability |
||||||
|
15 |
12 |
10 |
5 |
|||
|
Children |
0.5-1 |
9 |
[6.2]b |
[7.7]b |
[9.3]b |
[18.6]b |
|
1-3 |
13.3 |
3.9 |
4.8 |
5.8 |
11.6 |
|
|
4-6 |
19.2 |
4.2 |
5.3 |
6.3 |
12.6 |
|
|
7-10 |
28.1 |
5.9 |
7.4 |
8.9 |
17.8 |
|
|
|
|
|
|
|
|
|
|
Males
|
11-14 |
45 |
9.7 |
12.2 |
14.6 |
29.2 |
|
15-17 |
64.4 |
12.5 |
15.7 |
18.8 |
37.6 |
|
|
18+ |
75 |
9.1 |
11.4 |
13.7 |
27.4 |
|
|
|
|
|
|
|
|
|
|
Females
|
11-14 c |
46.1 |
9.3 |
11.7 |
14 |
28 |
|
11-14 |
46.1 |
21.8 |
27.7 |
32.7 |
65.4 |
|
|
15-17 |
56.4 |
20.7 |
25.8 |
31 |
62 |
|
|
18+ |
62 |
19.6 |
24.5 |
29.4 |
58.8 |
|
|
Post-menopausal |
|
62 |
7.5 |
9.4 |
11.3 |
22.6 |
|
Lactating |
|
62 |
10 |
12.5 |
15 |
30 |
In the weaning period, the iron requirements in relation to energy intake are the highest of the lifespan except for the last trimester of pregnancy, when iron requirements to a large extent have to be covered from the iron stores of the mother (see section on iron and pregnancy). The rapidly growing weaning infant has no iron stores and has to rely on dietary iron. It is possible to meet these high requirements if the diet has a consistently high content of meat and foods rich in ascorbic acid. In most developed countries today, infant cereal products are the staple foods for that period of life. Commercial products are regularly fortified with iron and ascorbic acid, and they are usually given together with fruit juices and solid foods containing meat, fish, and vegetables. The fortification of cereal products with iron and ascorbic acid is important in meeting the high dietary needs, especially considering the importance of an optimal iron nutrition during this phase of brain development.
Iron requirements are also very high in adolescents, particularly during the period of rapid growth (11). There is a marked individual variation in growth rate and the requirements may be considerably higher than the calculated mean values given in Table 39. Girls usually have their growth spurt before menarche, but growth is not finished at that time. Their total iron requirements are therefore considerable. In boys during puberty there is a marked increase in haemoglobin mass and concentration, further increasing iron requirements to a level above the average iron requirements in menstruating women (Figure 22). (The calculations in Figure 22 are based on references 8 and 12-16.)
Figure 22. Iron requirements of boys and girls
Menstrual iron losses
Menstrual blood losses are very constant from month to month for an individual but vary markedly from one woman to another (16).The main part of this variation is genetically controlled by the content of fibrinolytic activators in the uterine mucosa even in populations which are geographically widely separated (Burma, Canada, China, Egypt, England, and Sweden) (17, 18). These findings strongly suggest that the main source of variation in iron status in different populations is not related to a variation in iron requirements but to a variation in the absorption of iron from the diets. (This statement disregards infestations with hookworms and other parasites.) The mean menstrual iron loss, averaged over the entire menstrual cycle of 28 days, is about 0.56 mg/day. The frequency distribution of physiologic menstrual blood losses is highly skewed. Adding the average basal iron loss (0.8 mg) and its variation allows the distribution of the total iron requirements in adult women to be calculated as the convolution of the distributions of menstrual and basal iron losses (Figure 23). The mean daily total iron requirement is 1.36 mg. In 10 percent of women it exceeds 2.27 mg and in 5 percent it exceeds 2.84 mg (19). In 10 percent of menstruating (still-growing) teenagers, the corresponding daily total iron requirement exceeds 2.65 mg, and in 5 percent of the girls it exceeds 3.2 mg/day. The marked skewness of menstrual losses is a great nutritional problem because personal assessment of the losses is unreliable. This means that women with physiologic but heavy losses cannot be identified and reached by iron supplementation. The choice of contraceptive method greatly influences menstrual losses. The methods of calculating iron requirements in women and their variation were recently re-examined (19).
Figure 23. Distribution of daily iron requirements in menstruating adult women and teenagers: the probability of adequacy at different amounts of iron absorbed

Note: Left: basal obligatory losses that amount to 0.8 mg; right: varying menstrual iron losses. This graph illustrates that growth requirements in teenagers vary considerably at different age and between girls.
In postmenopausal women and in physically active elderly people, the iron requirements per unit of body weight are the same as in men. When physical activity decreases as a result of ageing, blood volume and haemoglobin mass also diminish, leading to a shift of iron from haemoglobin and muscle to iron stores. This implies a reduction of the daily iron requirements. Iron deficiency in the elderly is therefore seldom of nutritional origin but is usually caused by pathologic iron losses. The absorbed iron requirements in different groups are given in Table 39. Dietary iron requirements will be discussed below. The iron situation during pregnancy and lactation are dealt with separately below.
With respect to the mechanism of absorption, there are two kinds of dietary iron: heme iron and non-heme iron (20). In the human diet the primary sources of heme iron are the haemoglobin and myoglobin from consumption of meat, poultry, and fish whereas non-heme iron is obtained from cereals, pulses, legumes, fruits, and vegetables. The average absorption of heme iron from meat-containing meals is about 25 percent (21) The absorption of heme iron can vary from about 40 percent during iron deficiency to about 10 percent during iron repletion (22). Heme iron can be degraded and converted to non-heme iron if foods are cooked at a high temperature for too long. Calcium (see below) is the only dietary factor that negatively influences the absorption of heme iron and does so to the same extent that it influences non-heme iron (Table 41) (23).
Table 41
Factors influencing dietary iron absorption
|
HEME IRON ABSORPTION |
|
|
NON-HEME IRON ABSORPTION |
|
|
Enhancing factors |
Inhibiting factors |
Reducing substances (i.e., substances that keep iron in the ferrous form) must be present for iron to be absorbed (28). The presence of meat, poultry, and fish in the diet enhance iron absorption. Other foods contain factors (ligands) that strongly bind ferrous ions, that subsequently inhibit absorption. Examples are phytates and certain iron-binding polyphenols.
Inhibition of iron absorption
Phytates are found in all kinds of grains, seeds, nuts, vegetables, roots (e.g., potatoes), and fruits. Chemically, phytates are inositol hexaphosphate salts and are a storage form of phosphates and minerals. Other phosphates have not been shown to inhibit non-heme iron absorption. In North American and European diets, about 90 percent of phytates originate from cereals. Phytates strongly inhibit iron absorption in a dose-dependent fashion and even small amounts of phytates have a marked effect (29, 30).
Bran has a high content of phytate and strongly inhibits iron absorption. Whole-wheat flour, therefore, has a much higher content of phytates than does white wheat flour (31). In bread some of the phytates in bran are degraded during the fermentation of the dough. Fermentation for a couple of days (sourdough fermentation) can therefore almost completely degrade the phytate and increase the bio-availability of iron in bread made from whole-wheat flour (32). Oats strongly inhibit iron absorption because of their high phytate content, that results from native phytase in oats being destroyed by the normal heat process used to avoid rancidity (33). Sufficient amounts of ascorbic acid can counteract this inhibition (34). By contrast, non-phytate-containing dietary fibre components have almost no influence on iron absorption.
Almost all plants contain phenolic compounds as part of their defence system against insects, animals, and humans. Only some of the phenolic compounds (mainly those containing galloyl groups) seem to be responsible for the inhibition of iron absorption (35). Tea, coffee, and cocoa are common plant products that contain iron-binding polyphenols (36-39). Many vegetables, especially green leafy vegetables (e.g., spinach), and herbs and spices (e.g., oregano) contain appreciable amounts of galloyl groups, that strongly inhibit iron absorption. Consumption of betel leaves, common in areas of Asia, also has a marked negative effect on iron absorption.
Calcium, consumed as a salt or in dairy products interferes significantly with the absorption of both heme and non-heme iron (40-42). Because calcium and iron are both essential nutrients, calcium cannot be considered to be an inhibitor in the same way as phytates or phenolic compounds. The practical solution for this competition is to increase iron intake, increase its bio-availability, or avoid the intake of foods rich in calcium and foods rich in iron at the same meal (43).
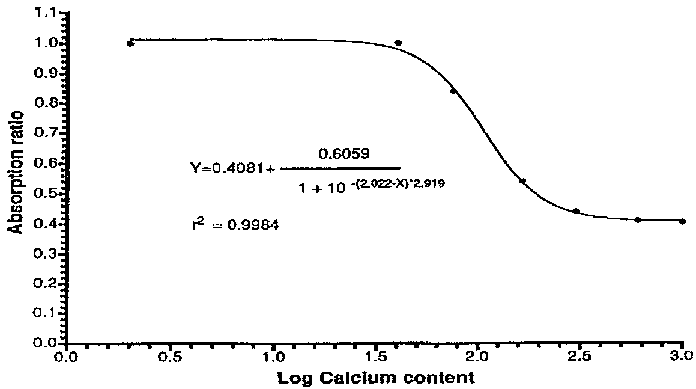
The mechanism of action for absorption inhibition is unknown, but the balance of evidence strongly suggest that the inhibition is located within the mucosal cell itself at the common final transfer step for heme and non-heme iron. Recent analyses of the dose-effect relationship show that no inhibition is seen from the first 40 mg of calcium in a meal. A sigmoid relationship is then seen, reaching a 60 percent maximal inhibition of iron absorption by 300-600 mg calcium. The form of this curve suggests a one-site competitive binding of iron and calcium (Figure 24). This relationship explains some of the seemingly conflicting results obtained in studies on the interaction between calcium and iron (44).
For unknown reasons, the addition of soy protein to a meal reduces the fraction of iron absorbed (45-48). This inhibition is not solely explained by the high phytate content of soy protein. However, because of the high iron content of soy proteins, the net effect on iron absorption of an addition of soy products to a meal is usually positive. In infant foods containing soy proteins, the inhibiting effect can be overcome by the addition of sufficient amounts of ascorbic acid. Some fermented soy sauces, however, have been found to enhance iron absorption (49, 50).
Figure 24. Effect of different amounts of calcium on iron absorption
Enhancement of iron absorption
Ascorbic acid is the most potent enhancer of non-heme iron absorption (34, 51-53). Synthetic vitamin C increases the absorption of iron to the same extent as the native ascorbic acid in fruits, vegetables, and juices. The effect of ascorbic acid on iron absorption is so marked and essential that this effect could be considered as one of vitamin C’s physiologic roles (54). Each meal should preferably contain at least 25 mg of ascorbic acid and possibly more if the meal contains many inhibitors of iron absorption. Therefore, a requirement of ascorbic acid for iron absorption should be taken into account when establishing the requirements for vitamin C, that are set only to prevent vitamin C deficiency (especially scurvy).
Meat, fish, and seafood all promote the absorption of non-heme iron (55-58). The mechanism for this effect has not been determined. It should be pointed out that meat also enhances the absorption of heme iron to about the same extent (21). Meat promotes iron nutrition in two ways: it stimulates the absorption of both heme and non-heme iron and it provides the well-absorbed heme iron. Epidemiologically, the intake of meat has been found to be associated with a lower prevalence of iron deficiency.
Organic acids, such as citric acid, have in some studies been found to enhance the absorption of non-heme iron (29). This effect is not observed as consistently as is the effect of ascorbic acid (47, 52). Sauerkraut (59) and other fermented vegetables and even some fermented soy sauces (49, 50) enhance iron absorption. The nature of this enhancement has not yet been determined.
Iron absorption from meals
The pool concept (see above) in iron absorption implies that there are two main pools in the gastrointestinal lumen - one pool of heme iron and another pool of non-heme iron - and that iron absorption takes place independently from these two pools (24). The pool concept also implies that the absorption of iron from the non-heme iron pool results from all ligands present in the mixture of foods included in a meal. The absorption of non-heme iron from a certain meal not only depends on its iron content but also, and to a marked degree, on the composition of the meal (i.e., the balance among all factors enhancing and inhibiting the absorption of iron). The bio-availability can vary more than 10-fold among meals with a similar content of iron, energy, protein, fat, etc. (20). Just the addition of certain spices (e.g., oregano) or a cup of tea may reduce the bio-availability by one-half or more. However, the addition of certain vegetables or fruits containing ascorbic acid may double or even triple iron absorption, depending on the other properties of the meal and the amounts of ascorbic acid present.
Iron absorption from the whole diet
There is limited information about the total amounts of iron absorbed from the diet because no simple method is available to measure iron absorption from the whole diet. It has been measured by chemical balance studies using long balance periods or by determining the haemoglobin regeneration rate in subjects with induced iron deficiency anaemia and a well-controlled diet over a long period of time.
A method was recently developed to measure iron absorption from the whole diet. In the first studies all non-heme iron in all meals over periods of 5-10 days was homogeneously labelled to the same specific activity with an extrinsic inorganic radioiron tracer (43, 60). Heme iron absorption was then estimated. In a further study, heme and non-heme iron were separately labelled with two radioiron tracers as biosynthetically labelled haemoglobin and as an inorganic iron salt (22). New information could be obtained, for example, about the average bio-availability of dietary iron in different types of diets, overall effects of certain factors (e.g., calcium) on iron nutrition, and regulation of iron absorption in relation to iron status. Iron absorption from the whole diet is the sum of the absorption of iron from the single meals included in the diet. It has been suggested that the iron absorption of single meals may exaggerate the absorption of iron from the diet (61, 62). Iron absorption from single meals can never represent iron absorption from the whole diet, but iron absorption from a single meal was the same when the meal was served in the morning after an overnight fast or at lunch or supper (63). The same observation was made in another study when a hamburger meal was served in the morning or 2-4 hours after a breakfast (42).
Because energy expenditure and energy intake set the limit for the amount of food eaten and for meal size, it is practical to relate the bio-availability of iron in different meals to energy content (bio-available nutrient density). The use of bio-available nutrient density is a feasible way to compare different meals, construct menus, and calculate recommended intakes (64).
Intake of energy and essential nutrients such as iron was probably considerably higher for early humans than it is today (65-67). The present low iron intake associated with a low-energy lifestyle implies that the interaction between different factors influencing iron absorption, will be more critical. For example, the interaction between calcium and iron absorption probably had no importance in the nutrition of early humans, who had a diet with ample amounts of both iron and calcium.
Iron balance and regulation of iron absorption
The body has three unique mechanisms for maintaining iron balance and preventing iron deficiency and iron overload. The first is the continuous re-utilisation of iron from catabolised erythrocytes in the body. When an erythrocyte dies after about 120 days, it is usually degraded by the macrophages of the reticular endothelium. The iron is released and delivered to transferrin in the plasma, which brings the iron back to red blood cell precursors in the bone marrow or to other cells in different tissues. Uptake and distribution of iron in the body is regulated by the synthesis of transferrin receptors on the cell surface. This system for internal iron transport not only controls the rate of flow of iron to different tissues according to their needs but also effectively prevents the appearance of free iron and the formation of free radicals in the circulation.
The second mechanism is the access of the specific storage protein, ferritin, which can store and release iron to meet excessive iron demands. This iron reservoir is especially important in the third trimester of pregnancy.
The third mechanism involves the regulation of absorption of iron from the intestines, with an increased iron absorption in the presence of decreasing body iron stores and a decreased iron absorption when iron stores increase. Iron absorption decreases until an equilibrium is established between absorption and requirements. For a given diet this regulation of iron absorption, however, can only balance losses up to a certain critical point beyond which iron deficiency will develop (68). About half of the basal iron losses are from blood, primarily in the gastrointestinal tract. Both these losses and the menstrual iron losses are influenced by the haemoglobin level; during the development of an iron deficiency, menstrual and basal iron losses will successively decrease when the haemoglobin level decreases. In a state of more severe iron deficiency, skin iron losses may also decrease. Iron balance (absorption equals losses) may be present not only in normal subjects but also during iron deficiency and iron overload.
The three main factors that affect iron balance are absorption (intake and bio-availability of iron), losses, and amount in stores. The interrelationship among these factors was recently been described in mathematical terms, making it possible to predict, for example, the amount of stored iron when iron losses and bio-availability of dietary iron are known (69). With increasing iron requirements or decreasing bio-availability, the regulatory capacity to prevent iron deficiency is limited (68). However, to prevent iron overload with increasing dietary iron intake or bio-availability, the regulatory capacity seems to be extremely good (69).
Populations at risk for iron deficiency
Worldwide, the highest prevalence of iron deficiency is found in infants, children, adolescents, and women of childbearing age, especially pregnant women. The weaning period in infants is especially critical because of the very high iron requirements in relation to energy requirements. Thanks to better information and access to fortified cereals for infants and children, the iron situation has markedly improved in these groups in most industrialized countries where the highest prevalences of iron deficiency today are observed in menstruating and pregnant women and adolescents of both sexes. In developing countries, however, the iron situation is very critical in many groups, especially in the weaning period. Iron nutrition is of great importance for the adequate development of the brain and other tissues such as muscles, which are finally differentiated early in life.
Iron deficiency and iron deficiency anaemia are often incorrectly used as synonyms. A definition of these terms may clarify some confusion about different prevalence figures given in the literature (70). The cause of the problem is the very wide distribution of the haemoglobin concentration in healthy, fully iron-replete subjects (in women, 120-160 g/l; in men, 140-180g/l [71]). During the development of a negative iron balance in subjects with no mobilisable iron from iron stores (no visible iron in technically perfect bone marrow smears or a serum ferritin concentration <15 µg/l), there will be an immediate impairment in the production of haemoglobin with a resulting decrease in haemoglobin and different erythrocyte indexes (e.g., mean corpuscular haemoglobin and mean corpuscular volume). In turn this will lead to an overlap of the distributions of haemoglobin in iron-deficient and iron-replete women (Figure 25). The extent of overlap depends on the prevalence and severity of iron deficiency. In populations with more severe iron deficiency, for example, the overlap is much less marked.
Figure 25. Distribution of haemoglobin concentration in a sample of 38-year-old women with and without stainable bone marrow iron

Note: The main fraction (91 percent) of the iron-deficient women in this sample had haemoglobin levels above the lowest normal level for the population: 120 g/l (mean ± 2 SD) (68). The degree of overlap of the two distributions depends on the severity of anaemia in a population.
In women, anaemia is defined as a haemoglobin <120 g/l. For a woman who has her normal homeostatic value set at 150 g/l, haemoglobin level must decrease to 119 g/l (by 26 percent) before she is considered to be anaemic, whereas for a woman who has her normal haemoglobin set at 121 g/l, haemoglobin level must only decrease by 1.5 percent to 119 g/l. Iron deficiency anaemia is a rather imprecise concept for evaluating the single subject and has no immediate physiologic meaning. By definition, this implies that the prevalence of iron deficiency anaemia is less frequent than iron deficiency and that the presence of anaemia in a subject is a statistical rather than a functional concept. The main use of the cut-off value is in comparisons between population groups (72). In practical work, iron deficiency anaemia should be replaced by the functional concept of iron deficiency. Anaemia per se is mainly important when it becomes so severe that oxygen delivery to tissues is impaired. An iron deficiency anaemia which develops slowly in otherwise healthy subjects with moderately heavy work output will not give any symptoms until the haemoglobin level is about 80 g/l or lower (71). The reason for the continued use of the concept of iron deficiency anaemia is the ease of determining haemoglobin. Therefore, in clinical practice, knowledge of previous haemoglobin values in a subject is of great importance for evaluating the diagnosis.
Iron deficiency being defined as an absence of iron stores combined with signs of an iron-deficient erythropoiesis implies that in a state of iron deficiency there is an insufficient supply of iron to various tissues. This occurs at a serum ferritin level <15 µg/l. Iron can then no longer be mobilised from iron stores and insufficient amounts of iron will be delivered to transferrin, the circulating transport protein for iron. The binding sites for iron on transferrin will therefore contain less and less iron. This is usually described as a reduction in transferrin saturation. When transferrin saturation drops to a certain critical level, erythrocyte precursors, which continuously need iron for the formation of haemoglobin, will get an insufficient supply of iron. At the same time, the supply of iron by transferrin to other tissues will also be impaired. Liver cells will get less iron, more transferrin will be synthesised, and the concentration of transferrin in plasma will then suddenly increase. Cells with a high turnover rate are the first ones to be affected (e.g., intestinal mucosal cells with a short life span). The iron-transferrin complex is bound to transferrin receptors on cell surfaces and the whole complex is then taken up by special receptors on the surface of various cells and tissues. The uptake of iron seems to be related both to transferrin saturation and the number of transferrin receptors on the cell surface (73, 74). There is a marked diurnal variation in the saturation of transferrin because the turnover rate of iron in plasma is very high. This fact makes it difficult to evaluate the iron status from single determinations of transferrin saturation.
Indicators of iron deficiency
The absence of iron stores (iron deficiency) can be diagnosed by showing that there is no stainable iron in the reticuloendothelial cells in bone marrow smears or more easily by a low concentration of ferritin in serum (£15 µg/l). Even if an absence of iron stores per se may not necessarily be associated with any immediate adverse effects, it is a reliable and good indirect indicator of iron-deficient erythropoiesis and of an increased risk of a compromised supply of iron to different tissues.
Even before iron stores are completely exhausted, the supply of iron to the erythrocyte precursors in the bone marrow is compromised, leading to iron-deficient erythropoiesis (70). A possible explanation is that the rate of release of iron from stores is influenced by the amount of iron remaining. As mentioned above it can then be assumed that the supply of iron to other tissues needing iron is also insufficient because the identical transport system is used. During the development of iron deficiency haemoglobin concentration, transferrin concentration, transferrin saturation, transferrin receptors in plasma, erythrocyte protoporphyrin, and erythrocyte indexes are changed. All these methods, however, show a marked overlap between normal and iron-deficient subjects, that makes it impossible to identify the single subject with mild iron deficiency by using any of these methods. Therefore, these tests have been used in combination (e.g., for interpreting results from the second National Health and Nutrition Examination Survey in the United States of America [75, 76]). The diagnostic specificity then increases but the sensitivity decreases, and thus the true prevalence of iron deficiency is markedly underestimated if multiple diagnostic criteria are used. By definition in screening for iron deficiency, the more tests that are used the higher is the diagnostic specificity but the lower is the sensitivity of the procedure. Fortunately, a low serum ferritin, £15µg/l is always associated with an iron-deficient erythropoiesis. The use of serum ferritin alone as a measure will also underestimate the true prevalence of iron deficiency but to a lesser degree than when the combined criteria are used.
A diagnosis of iron deficiency anaemia can be suspected if anaemia is present in subjects who are iron-deficient as described above. Preferably, to fully establish the diagnosis, the subjects should respond adequately to iron treatment. The pitfalls with this method are the random variation in haemoglobin concentrations over time and the effect of the regression towards the mean when a new measurement is made.
The use of serum ferritin has improved the diagnostic accuracy of iron deficiency. It is the only simple method available to detect early iron deficiency. Its practical value is somewhat reduced, however, by the fact that serum ferritin is a very sensitive acute-phase reactant and may be increased for weeks after a simple infection with fever for a day or two (77). Several other conditions, such as use of alcohol (78, 79), liver disease, and collagen diseases, may also increase serum ferritin concentrations. Determination of transferrin receptors in plasma has also been recommended in the diagnosis of iron deficiency. Its advantage is that it is not influenced by infections. Its main use is in subjects who are already anaemic and it is not sensitive for the early diagnosis of iron deficiency. The use of a combination of determinations of serum ferritin and serum transferrin receptors has also been suggested (80).
Causes of iron deficiency
Nutritional iron deficiency implies that the diet cannot cover physiologic iron requirements. Worldwide this is the most common cause of iron deficiency. In many tropical countries, infestations with hookworms lead to intestinal blood losses that may be considerable. The severity of the infestations with hookworms varies considerably between subjects and regions. The average blood loss can be well estimated by egg counts in stools. Usually the diet in these populations is also limited with respect to iron content and availability.
In clinical practice a diagnosis of iron deficiency must always lead to a search for pathologic causes of blood loss (e.g., tumours in the gastrointestinal tract or uterus, especially if uterine bleedings have increased or changed in regularity). Patients with achlorhydria absorb dietary iron less well (a reduction of about 50 percent) and patients who have undergone gastric surgery, especially if the surgery was extensive, may eventually develop iron deficiency because of impaired iron absorption. Gluten enteropathy is another possibility to consider, especially in young patients.
Prevalence of iron deficiency
Iron deficiency is probably the most frequent nutritional deficiency disorder in the world. A recent estimate based on World Health Organization (WHO) criteria indicated that around 600-700 million people worldwide have a marked iron deficiency anaemia (81). In industrialized countries, the prevalence of iron deficiency anaemia is much lower and usually varies between 2 percent and 8 percent. However, the prevalence of iron deficiency, including both anaemic and non-anaemic subjects (see definitions above), is much higher. In industrialized countries, for example, an absence of iron stores or subnormal serum ferritin values is found in about 20-30 percent of women of fertile age. In adolescent girls the prevalence is even higher.
It is difficult to determine the prevalence of iron deficiency more exactly because representative populations for clinical investigation are hard to obtain. Laboratory methods and techniques for blood sampling need careful standardization. One often neglected source of error, for example, when materials in different regions or at different times are compared, is the fact that there are still reagent kits on the market for determining serum ferritin which are not adequately calibrated at different concentrations against the international WHO standards. In addition, seasonal variations in infection rates influence the sensitivity and specificity of most methods used.
Worldwide, the highest prevalence figures for iron deficiency are found in infants, children, teenagers, and women of childbearing age. Thanks to better information and access to fortified cereals for infants and children, the iron situation has markedly improved in these groups in most industrialized countries, where the highest prevalence today is observed in menstruating women and adolescents of both sexes.
In developing countries, where the prevalence of iron deficiency is very high and the severity of anaemia is marked, studies on the distribution of haemoglobin in different population groups can provide important information as a valuable basis for action programmes (72). A more detailed analysis of subsamples may then give excellent information for the planning of more extensive programmes.
Effects of iron deficiency
Studies in animals have clearly shown that iron deficiency has several negative effects on important functions in the body (3). Physical working capacity in rats has been shown to be significantly reduced in iron deficiency, that is especially valid for endurance activities (82, 83). This negative effect seems to be less related to the degree of anaemia than to impaired oxidative metabolism in the muscles with an increased formation of lactic acid, that in turn is due to a lack of iron-containing enzymes which are rate limiting for the oxidative metabolism (84).
The relationship between iron deficiency and brain function is of great importance for the choice of strategy in combating iron deficiency (85-88). Several structures in the brain have a high iron content of the same magnitude as observed in the liver. Of great importance is the observation that the lower iron content of the brain in iron-deficient growing rats cannot be increased by giving iron later on. This fact strongly suggests that the supply of iron to brain cells takes place during an early phase of brain development and that, as such, early iron deficiency may lead to irreparable damage to brain cells.
In humans about 10 percent of brain iron is present at birth; at the age of 10 years the brain has only reached half its normal iron content, and optimal amounts are first reached at the age of 20-30 years.
In populations with long-standing iron deficiency, a reduction of physical working capacity has been demonstrated by several groups with improvement in working capacity after iron administration (84).
Iron deficiency also negatively influences the normal defence systems against infections. The cell-mediated immunologic response by the action of T lymphocytes is impaired as a result of a reduced formation of these cells. This in turn is due to a reduced DNA synthesis depending on the function of ribonucleotide reductase, which requires a continuous supply of iron for its function. The phagocytosis and killing of bacteria by the neutrophil leukocytes is an important component of the defence mechanism against infections. These functions are impaired in iron deficiency. The killing function is based on the formation of free hydroxyl radicals within the leukocytes, the respiratory burst, and results from the activation of the iron-sulphur enzyme NADPH oxidase and probably also cytochrome b (a heme enzyme) (89).
The impairment of the immunologic defence against infections that was found in animals is also regularly found in humans. Administration of iron normalises these changes within 4-7 days. It has been difficult to demonstrate, however, that the prevalence of infections is higher or that their severity is more marked in iron-deficient subjects than in control subjects. This may well be ascribed to the difficulty in studying this problem with an adequate experimental design.
A relationship between iron deficiency and behaviour such as attention, memory, and learning, has been demonstrated in infants and small children by several groups. In the most recent well-controlled studies, no effect was noted from the administration of iron. This finding is consistent with the observations in animals. Therapy-resistant behavioural impairment and the fact that there is an accumulation of iron during the whole period of brain growth should be considered strong arguments for the more active and effective combating of iron deficiency. This is valid for women, especially during pregnancy, for infants and children, and up through the period of adolescence and early adulthood. In a recent well-controlled study, administration of iron to non-anaemic but iron-deficient adolescent girls improved verbal learning and memory (90).
Well-controlled studies in adolescent girls show that iron-deficiency without anaemia is associated with reduced physical endurance (91) and changes in mood and ability to concentrate (92). A recent careful study showed that there was a reduction in maximum oxygen consumption in non-anaemic women with iron deficiency that was unrelated to a decreased oxygen-transport capacity of the blood (93).
Iron requirements during pregnancy are well established (Table 42). Most of the iron required during pregnancy is used to increase the haemoglobin mass of the mother, which occurs in all healthy pregnant women who have sufficiently large iron stores or who are adequately supplemented with iron. The increased haemoglobin mass is directly proportional to the increased need for oxygen transport during pregnancy and is one of the important physiologic adaptations that occurs in pregnancy (94, 95). A major problem for iron balance in pregnancy is that iron requirements are not equally distributed over its duration. The exponential growth of the foetus implies that iron needs are almost negligible in the first trimester and that more than 80 percent relates to the last trimester. The total daily iron requirements, including the basal iron losses (0.8 mg), increase during pregnancy from 0.8 mg to about 10 mg during the last 6 weeks of pregnancy.
Iron absorption during pregnancy is determined by the amount of iron in the diet, its bio-availability (meal composition), and the changes in iron absorption that occur during pregnancy. There are marked changes in the fraction of iron absorbed during pregnancy. In the first trimester there is a marked, somewhat paradoxical, decrease in the absorption of iron, which is closely related to the reduction in iron requirements during this period as compared with the non-pregnant state (see below). In the second trimester iron absorption is increased by about 50 percent, and in the last trimester it may increase by up to about four times. Even considering the marked increase in iron absorption, it is impossible for the mother to cover her iron requirements from diet alone, even if its iron content and bio-availability are very high. It can be calculated that with diets prevailing in most industrialized countries, there will be a deficit of about 400-500 mg in the amount of iron absorbed during pregnancy (Figure 26).
An adequate iron balance can be achieved if iron stores of 500 mg are available. However, it is uncommon for women today to have iron stores of this size. It is therefore recommended that iron supplements in tablet form, preferably together with folic acid, be given to all pregnant women because of the difficulties in correctly evaluating iron status in pregnancy with routine laboratory methods. In the non-anaemic pregnant woman, daily supplements of 100 mg of iron (e.g., as ferrous sulphate) given during the second half of pregnancy are adequate. In anaemic women higher doses are usually required.
Table 42
Iron requirements during pregnancy
|
|
Iron requirements (mg) |
|
IRON REQUIREMENTS DURING PREGNANCY |
300 |
|
Foetus Placenta |
50 |
|
Expansion of maternal erythrocyte mass |
450 |
|
Basal iron losses |
240 |
|
Total iron requirement |
1040 |
|
NET IRON BALANCE AFTER DELIVERY |
|
|
Contraction of maternal erythrocyte mass |
+450 |
|
Maternal blood loss |
-250 |
|
Net iron balance |
+200 |
|
Net iron requirements for pregnancy if sufficient maternal
iron stores are present (1040 - 200 = 840) |
840 |
There is an association between low haemoglobin values and prematurity. An extensive study (96) showed that a woman with a hematocrit of 37 percent had twice the risk of having a premature birth as did a woman with a hematocrit between 41 percent and 44 percent (P£0.01). A similar observation was reported in another extensive study in the United States of America (97). These materials were examined retrospectively and the cause of the lower hematocrit was not examined.
In lactating women, the daily iron loss in milk is about 0.3 mg. Together with the basal iron losses of 0.8 mg, the total iron requirements during the lactation period amount to 1.1 mg/day.
Early in pregnancy there are marked hormonal, haemodynamic, and haematologic changes. There is, for example, a very early increase in the plasma volume, which has been used to explain the physiologic anaemia of pregnancy observed also in iron-replete women. The primary cause of this phenomenon, however, is more probably an increased ability of the haemoglobin to deliver oxygen to the tissues (foetus). This change is induced early in pregnancy by increasing the content of 2, 3-diphospho-D-glycerate in the erythrocytes, which shifts the hemoglobin-oxygen dissociation curve to the right. The anaemia is a consequence of this important adaptation and is not primarily a desirable change, for example, to improve placental blood flow by reducing blood viscosity.
Figure 26. Daily iron requirements and daily dietary iron absorption in pregnancy

Note: The hatched area represents the deficit of iron that has to be covered by iron from stores or iron supplementation.
Another observation has likewise caused some confusion about the rationale of giving extra iron routinely in pregnancy. In extensive studies of pregnant women, there is a U-shaped relationship between various pregnancy complications and the haemoglobin level (i.e., there are more complications at both low and high levels). There is nothing to indicate, however, that high haemoglobin levels (within the normal non-pregnant range) per se have any negative effects. The haemoglobin increase is caused by pathologic hormonal and hemodynamic changes induced by an increased sensitivity to angiotensin II that occurs in some pregnant women, leading to a reduction in plasma volume, hypertension, and toxaemia of pregnancy.
Pregnancy in adolescents presents a special problem because iron is needed to cover the requirements of growth. In countries with very early marriage, a girl may get pregnant before menstruating. The additional iron requirements for growth of the mother are then very high and the iron situation is very serious.
In summary, the marked physiologic adjustments occurring in pregnancy are not sufficient to balance its very marked iron requirements, and the pregnant woman has to rely on her iron stores, if present. The composition of the diet has not been adjusted to the present low-energy-demanding lifestyle in industrialized countries. This is probably the main cause of the critical iron-balance situation in pregnancy today, that is due to absent or insufficient iron stores in women before they get pregnant. The unnatural necessity to give extra nutrients such as iron and folate to otherwise healthy pregnant women should be considered in this perspective.
The prevention of iron deficiency has become even more urgent in recent years with the accumulation of evidence strongly suggesting a relationship between even mild iron deficiency and brain development and especially with the observation that functional defects affecting learning and behaviour cannot be reversed by giving iron later on. As mentioned, iron deficiency is common both in developed and in developing countries. Great efforts have been made by WHO to develop methods to combat iron deficiency.
Iron deficiency can generally be combated by one or more of the following three strategies: 1) iron supplementation (i.e., giving iron tablets to certain target groups such as pregnant women and preschool children); 2) iron fortification of certain foods, such as flour; and 3) food and nutrition education to improve the amount of iron absorbed from the diet by increasing the intake of iron and especially by improving the bio-availability of the dietary iron.
Several factors determine the feasibility and effectiveness of different strategies, such as the health infrastructure of a society, the economy, access to suitable vehicles for iron fortification, etc. The solutions are therefore often quite different in developing and developed countries. There is an urgency to obtain knowledge about the feasibility of different methods to improve iron nutrition and to apply present knowledge. In addition, initiation of local activities should be stimulated while actions from governments are awaited.
To translate physiologic iron requirements, given in Table 30, into dietary iron requirements, the bio-availability of iron in different diets must be calculated. It is therefore necessary to choose an iron status where the supply of iron to the erythrocyte precursors and other tissues starts to be compromised. A state of iron-deficient erythropoiesis occurs when iron can no longer be mobilised from iron stores; iron can no longer be mobilised when stores are almost completely empty. A reduction then occurs, for example, in the concentration of haemoglobin and in the average content of haemoglobin in the erythrocytes (a reduction in mean corpuscular haemoglobin). At the same time the concentration of transferrin in the plasma increases because of an insufficient supply of iron to liver cells. These changes were recently shown to occur rather suddenly at a level of serum ferritin of £15 µg/l (68, 70). A continued negative iron balance will further reduce the level of haemoglobin. Symptoms related to iron deficiency are less related to the haemoglobin level and more to the fact that there is a compromised supply of iron to tissues.
The bio-availability of iron in meals consumed in countries with a Western-type diet has been measured by using different methods. Numerous single-meal studies have shown absorption of non-heme iron ranging from 5 percent to 40 percent (59, 98, 99). Attempts have also been made to estimate the bio-availability of dietary iron in populations consuming Western-type diets by using indirect methods (e.g., calculation of the coverage of iron requirements in groups of subjects with known dietary intake). Such estimations suggest that in borderline iron-deficient subjects the bio-availability from good diets may reach a level around 14-16 percent (15 percent relates to subjects who have a serum ferritin value of <15µg/l or a reference dose absorption of 56.5 percent) (19).
Recently, direct measurements were made of the average bio-availability of iron in different Western-type diets (22, 43, 60). Expressed as total amounts of iron absorbed from the whole diet, it was found that 53.2 µg Fe/kg/day could be absorbed daily from each of the two main meals of an experimental diet which included ample amounts of meat or fish. For a body weight of 55 kg and an iron intake of 14 mg/day, this corresponds to a bio-availability of 21 percent in subjects with no iron stores and an iron-deficient erythropoiesis. A diet common among women in Sweden contained smaller portions of meat and fish, higher phytates, and some vegetarian meals each week was found to have a bio-availability of 12 percent. Reducing the intake of meat and fish further will reduce the bio-availability to about 10 percent (25µg Fe/kg/day). In vegetarians the bio-availability is usually low because of the absence of meat and fish and a high intake of phytate and polyphenols. An average good Western-type whole diet has a bio-availability of about 15 percent but for common diets, especially among women, the bio-availability is around 12 percent or even 10 percent. In countries or for certain groups in a population with a very high meat intake, the bio-availability may rather be around 18 percent. In Western countries, a high bio-availability is mainly associated with a high meat intake, a high intake of ascorbic acid with meals, a low intake of phytate-rich cereals, and no coffee or tea within 2 hours of the main meals (38). Table 43 shows examples of diets with different iron bio-availability.
Table 43. Examples of diet with different iron bio-availability
|
Type of diet |
Bio-availability µg/kg/day |
|
Preagricultural ancestors |
|
|
Very high meat and ascorbic acid |
150 |
|
Very high meat in 2 main meals daily and high ascorbic acid
(theoretical) |
75 |
|
High meat/fish in 2 main meals daily |
66.7 |
|
Moderate meat/fish in 2 main meals daily |
53.2 |
|
Moderate meat/fish in 2 main meals daily, low phytate and
calcium |
42.3 |
|
Meat/fish in 60% of 2 main meals daily, high phytate and
calcium |
31.4 |
|
Lower meat intake, high phytate. Often one main
meal. |
25 |
|
Meat/fish negligible, high phytate. High tannin, low ascorbic
acid. |
15 |
Translation of bio-availability expressed as amount of iron absorbed into percent absorbed for two levels of iron intake
|
Bio-availability, µg/kg/day |
Absorption as mg Fe at no iron stores in women of 55 kg
body weight |
Bio-availability % |
|
|
|
|
15 mg |
17 mg |
|
150 |
8.25 |
55 |
48.8 |
|
75 |
4.13 |
27.5 |
24.4 |
|
66.7 |
3.67 |
24.5 |
21.8 |
|
53.2 |
2.93 |
19.5 |
17 |
|
42.3 |
2.32 |
15.5 |
13.5 |
|
31.4 |
1.73 |
11.5 |
10 |
|
25 |
1.38 |
9.2 |
8.2 |
|
15 |
0.83 |
5.5 |
4.7 |
It is obvious that absorbed iron requirements need to be adjusted to different types of diets, especially in vulnerable groups. The Food and Agriculture Organization of the United Nations (FAO) and WHO recommended, for didactic reasons, three bio-availability levels of 5 percent, 10 percent, and 15 percent (8). For developing countries, it may be realistic to use the figures of 5 percent and 10 percent. In populations consuming more Western-type diets, two levels would be adequate - 12 percent and 15 percent - mainly depending on meat intake.
The amount of dietary iron absorbed is mainly determined by the amount of body stores of iron and by the properties of the diet (iron content and bio-availability). In anaemic subjects the rate of erythrocyte production also influences iron absorption. In a 55-kg woman with average iron losses who consumes a diet with an iron bio-availability of 15 percent, the mean iron stores would be about 120 mg. Under these circumstances approximately 10-15 percent of women would have no iron stores. When a diet with a bio-availability of 12 percent is consumed by a 55-kg woman, iron stores would be approximately 75 mg and about 25-30 percent of women would have no iron stores at all. When the bio-availability of iron decreases to 10 percent, mean iron stores are reduced to about 25 mg and about 40-50 percent of women consuming this diet would have no iron stores. Those consuming diets with an iron bio-availability of 5 percent have no iron stores and they are iron deficient. These calculations are based on a recent study (69).
Tables 39 and 40 showed both the physiologic absorbed iron requirements and the dietary iron requirements. All these figures are for the 95th percentile of iron requirements. The figures are given for women with a body weight of 55 kg and men with a body weight of 70 kg. For example, women with a body weight of 45 kg and men with a body weight of 55 kg have iron requirements that are 20 percent lower than those given in Table 39.
No figures are given for dietary iron requirements in pregnant women because the iron balance in pregnancy depends not only on the properties of the diet but also and especially on the amounts of stored iron.
1. Bothwell, T.H. 1979. Iron metabolism in man. London, Blackwell Scientific Publications.
2. Hallberg, L. 1982. Iron absorption and iron deficiency. Hum Nutr:Clin. Nutr., 36:259-278.
3. Dallman, P.R. 1986. Biochemical basis for the manifestations of iron deficiency. Ann. Rev. Nutr., 6: 13-40.
4. Brock, J.H., Halliday, J.W., M.J.P. & Powell, L.W. 1994. Iron metabolism in health and disease, London, W.B. Saunders Company Ltd.
5. Kühn, L.C. 1996. Control of cellular iron transport and storage at the molecular level. In: Hallberg LA, et al., eds. Iron nutrition in health and disease. p. 17-29. London, John Libbey & Company.
6. Mascotti, D.P., Rup, D. & Thach, R.E. 1995. Regulation of iron metabolism: Translational effects medicated by iron, heme and cytokines. Ann. Rev. Nutr., 15: 239-61.
7. Green, R. 1968. Body iron excretion in man. A colloborative study. Am. J. Med., 45: 336-353.
8. FAO/WHO. 1988. Requirements of vitamin A, iron, folate and vitamin B12. Report of a Joint FAO/WHO Expert Consultation.. Rome: FAO. (FAO Food and Nutrition Series No. 23).
9. Brune, M. 1986. Iron losses in sweat. Am. J. Clin. Nutr., 43: 438-443.
10. European Communities. 1993. Nutrient and energy intakes for the European Community. EG-Report. Brussels Luxembourg: Commission of the European Communities.
11. Rossander-Hulthén, L. & Hallberg L. 1996. Prevalence of iron deficiency in adolescents. In: Hallberg L, Asp N-G, eds. Iron nutrition in health and diseas. p.149-156. London, John Libby& Co.
12. Dallman, P.R. & Siimes, M. 1979. Percentile curves for hemoglobin and red cell volume in infancy and childhood. J. Pediatr., 94: 26-31.
13. Tanner, J.M., Whitehouse, R.H. & Takaishi, M. 1966. Standards from birth to maturity for height, weight, height velocity, and weight velocity in British children, 1965, Part I. Arch. Dis. Child., 41: 454-471.
14. Tanner, J.M., Whitehouse, R.H. & Takaishi, M. 1966. Standards from birth to maturity for height, weight, height velocity, and weight velocity in British children, 1965, Part II. . Arch Dis. Child., 41: 613-632.
15. Karlberg, P. 1976. The somatic development of children in a Swedish urban community. Acta. Paediatr. Scand. Supplement, 258: 5-147.
16. Hallberg, L. 1966. Menstrual blood loss - a population study. Variation at different ages and attempts to define normality. Acta. Obstet. Gynecol. Scand., 45: 320-351.
17. Rybo, G-M. & Hallberg, L. 1966. Influence of heredity and environment on normal menstrual blood loss. A study of twins. Acta. Obstet. Gynecol. Scand., 45: 389-410.
18. Rybo, G-M. 1966. Plasminogen activators in the endometrium. I. Methodological aspects and II. Clinical aspects. Acta. Obstet. Gynecol. Scand., 45: 411-450.
19. Hallberg, L. & Rossander-Hulthénm, L. 1991. Iron requirements in menstruating women. Am. J. Clin. Nutr., 54: 1047-1058.
20. Hallberg, L. 1981. Bio-availability of dietary iron in man. Ann. Rev. Nutr., 1: 123-147.
21. Hallberg, L. 1979. Dietary heme iron absorption. A discussion of possible mechanisms for the absorption-promoting effect of meat and for the regulation of iron absorption. Scand. J. Gastroenterol., 14: 769-779.
22. Hallberg, L., Hulthén, L. & Gramatkovski, E. 1997. Iron absorption from the whole diet in men: how effective is the regulation of iron absorption? Am. J. Clin. Nutr., 66: 347-56.
23. Hallberg, L. 1993. Inhibition of haem-iron absorption in man by calcium. Br. J. Nutr., 69:533-540.
24. Hallberg, L. 1974. The pool concept in food iron absorption and some of its implications. Proc. Nutr. Soc., 33: 285-291.
25. Hallberg, L. 1985. Factors influencing the efficacy of iron fortification and the selection of fortification vehicles. In: Clydesdale FM, Wiemer KL, eds. Iron fortification of foods. p. 17-28. New York: Academic Press Inc. p. 17-28.
26. Hallberg, L. & Björn-Rasmussen, E. 1981. Measurement of iron absorption from meals contaminated with iron. Am. J. Clin. Nutr., 34: 2808-2815.
27. Hallberg, L. 1983. Iron absorption from some Asian meals containing contamination iron. Am. J. Clin. Nutr., 1983, 37:272-277.
28. Wollenberg, P. & Rummel, W. 1987. Dependence of intestinal iron absorption on the valency state of iron. Naunyn-Schmiedeberg´s Arch. Pharmacol., 36: 578 - 582.
29. Gillooly, M. 1983. The effect of organic acids, phytates and polyphenols on absorption of iron from vegetables. Br. J. Nutr., 49: 331-342.
30. Hallberg, L., Brune, M. & Rossander, L. 1989. Iron absorption in man: ascorbic acid and dose-dependent inhibition by phytate. Am. J. Clin. Nutr., 49: 140-144.
31. Hallberg, L., Rossander, L. & Skånberg, A-B. 1987. Phytates and the inhibitory effect of bran on iron absorption in man. Am. J. Clin. Nutr., 45: 988-996.
32. Brune, M. 1992. Iron absorption from bread in Humans: Inhibiting effects of cereal fiber, phytate and inositol phosphates with different numbers of phosphate groups. J. Nutr., 122: 442-449.
33. Rossander-Hulthén, L., Gleerup, A. & Hallberg, L. 1990. Inhibitory effect of oat products on non-haem iron absorption in man. Eur. J. Clin. Nutr., 44: 783-791.
34. Siegenberg, D. 1991. Ascorbic acid prevents the dose-dependent inhibitory effects of polyphenols and phytates on nonheme-iron absorption. Am. J. Clin. Nutr., 53: 537-41.
35. Brune, M., Rossander, L. & Hallberg, L. 1989. Iron absorption and phenolic compounds: importance of different phenolic structures. Eur. J. Clin. Nutr., 43: 547-558.
36. Disler, P.B. 1975. The effect of tea on iron absorption. Gut, 16: 193-200.
37. Derman, D. 1977. Iron absorption from a cereal-based meal containing cane sugar fortified with ascorbic acid. Br. J. Nutr., 38: 261-269.
38. Morck, T.A., Lynch, S.E. & Cook, J.D. 1983. Inhibition of food iron absorption by coffee. Am. J. Clin. Nutr., 37: 416-420.
39. Hallberg, L. & Rossander, L. 1982. Effect of different drinks on the absorption of non-heme iron from composite meals. Hum. Nutr: Appl. Nutr., 36: 116-123.
40. Hallberg, L. 1991. Calcium: effect of different amounts on nonheme-and heme-iron absorption in Humans. Am. J. Clin. Nutr., 53: 112-119.
41. Hallberg, L. 1992. Calcium and iron absorption: mechanism of action and nutritional importance. Eur. J. Clin. Nutr., 46: 317-327.
42. Gleerup, A., Rossander-Hultén, L. & Hallberg, L. 1993. Duration of the inhibitory effect of calcium on non-haem iron absorption in man. Eur. J. Clin. Nutr., 47: 875-879.
43. Gleerup, A. 1995. Iron absorption from the whole diet: comparison of the effect of two different distributions of daily calcium intake. Am. J. Clin. Nutr., 61: 97-104.
44. Hallberg, L. 1998. Does calcium interfere with iron absorption? Am. J. Clin. Nutr., 68: 3-4.
45. Cook, J.D., Morck,T.A. & Lynch, S.R. 1981. The inhibitory effects of soy products on nonheme iron absorption in man. Am. J. Clin. Nutr., 34: 2622-9.
46. Hallberg, L. & Hultén, L. 1982. Effect of soy protein on nonheme iron absorption in man. Am. J. Clin. Nutr., 36: 514-520.
47. Hallberg, L. & Rossander, L. 1984. Improvment of iron nutrition in developing countries: comparison of adding meat, soy protein, ascorbic acid, citric acid, and ferrous sulphate on iron absorption from a simple Latin American-type of meal. Am. J. Clin. Nutr., 39: 577-583.
48. Hurrell, R.F. 1992. Soy protein, phytate, and iron absorption in Humans. Am. J. Clin. Nutr., 56: 573-578.
49. Baynes, R.D. 1990. The promotive effect of soy sauce on iron absorption in Human subjects. Eur. J. Clin. Nutr., 44: 419-24.
50. Macfarlane, B.J. 1990. The effect of traditional oriental soy products on iron absorption. Am. J. Clin. Nutr., 51: 873-80.
51. Cook, J.D. & Monsen, E.R. 1977. Vitamin C, the common cold and iron absorption. Am. J. Clin. Nutr., 30: 235-241.
52. Hallberg, L., Brune, M. & Rossander, L. 1986. Effect of ascorbic acid on iron absorption from different types of meals. Studies with ascorbic-acid-rich foods and synthetic ascorbic acid given in different amounts with different meals. Hum. Nutr: Appl. Nutr., 40: 97-113.
53. Derman, D.P. 1980. Importance of ascorbic acid in the absorption of iron from infant foods. Scand. J. Haematol., 25: 193-201.
54. Hallberg, L., Brune, M. & Rossander-Hultén, L-S. 1987. Is there a physiological role of vitamin C in iron absorption? Ann. York. Acad. Sci., 498: 324-332.
55. Layrisse, M., Martinez-Torres, C. & Roch, M. 1968. The effect of interaction of various foods on iron absorption. Am. J. Clin. Nutr., 21: 1175-1183.
56. Layrisse, M. 1969. Food iron absorption: A comparison of vegetable and animal foods. Blood, 33: 430-443.
57. Cook, J.D. & Monsen, E.R. 1976. Food iron absorption in Human subjects. III. Comparison of the effect of animal proteins on nonheme iron absorption. Am. J. Clin. Nutr., 29: 859-867.
58. Björn-Rasmussen, E. & Hallberg, L. 1979. Effect of animal proteins on the absorption of food iron in man. Nutr. Metab., 23: 192-202.
59. Hallberg, L. & Rossander, L. 1982. Absorption of iron from Western-type lunch and dinner meals. Am. J. Clin. Nutr., 35: 502-509.
60. Hulthén, L. 1995. Iron absorption from the whole diet. Relation to meal composition, iron requirements and iron stores. Eur. J. Clin. Nutr., 49: 794-808.
61. Hallberg, L. & Hulthén, L. 1996. Methods to study dietary iron absorption in man -. an overview. In: Hallberg L, Asp N-G, eds. Iron nutrition in health and disease. p. 81-95. London, John Libbey & Company Ltd.
62. Cook, J.D., Dassenko, S.A. & Lynch, S.R. 1991. Assessment of the role of non heme- iron availability in iron balance. Am. J. Clin. Nutr., 54: 717-722.
63. Taylor, P.G. 1995. Iron bio-availability from diets consumed by different socio-economic strata of the Venezuelan population. J. Nutr., 25: 1860-1868.
64. Hallberg, L. 1981. Bio-available nutrient density: a new concept applied in the interpretation of food iron absorption data. Am. J. Clin. Nutr., 34: 2242-2247.
65. Eaton, S.B. & Konner, M. 1985. Paleolithic nutrition: A consideration of its nature and current implications. N. Engl. J. Med., 312: 283-289.
66. Eaton, S.B. & Nelson, D.A. 1991. Calcium in evolutionary perspective. Am. J. Clin. Nutr., 54: S-281-287.
67. Eaton, S.B., Eaton, III S.B. & Konner, M. 1997. Paleolithic nutrition revisited: a twelve year retrospective on its nature and implications. Eur. J. Clin. Nutr., 51: 207-216.
68. Hallberg, L. 1995. Iron balance in menstruating women. Eur. J. Clin. Nutr., 49: 200-207.
69. Hallberg L., Hulthén L. & Garby L. 1998. Iron stores in man in relation to diet and iron requirements. Eur. J. Clin. Nutr., 52: 623-31.
70. Hallberg, L. 1993. Screening for iron deficiency: an analysis based on bone-marrow examinations and serum ferritin determinations in a population sample of women. Br. J. Haematol., 85: 787-798.
71. Wintrobe, M.M. 1981. Clinical Hematology. (Eighth ed.), Philadelphia, Lea & Febiger.
72. Yip, R., Stolzfus, R.J. & W.K.S. 1996. Assessment of the prevalence and the nature of iron deficiency for populations: the utility of comparing haemoglobin distributions. In: Hallberg L, Asp, N-G., eds. Iron nutrition in health and disease. London, John Libby & Company Ltd.
73. Harford, J.B., Röuault, T.A. & Klausner, R.D. 1994. The control of cellular iron homeostasis. In: Brock JH et al., eds. Iron metabolism in health and disease. p.123-149. London, W.B.Saunders Company Ltd.
74. Baker, E. & Morgan, E.H. 1994. Iron transport. In: Brock JH et al., eds. Iron metabolism in health and disease. p.63-95. London, W.B. Saunders Company Ltd.
75. Pilch, S.M. & Senti, F.R.E. 1984. Assessment of the iron nutritional status of the US population based on data collected in the second National Health and Nutrition Examination Survey, 1976-1980. Prepared for the Food and Drug Administration under Contract no FDA 223-83-2384. Bethesda, MD, Life Sciences Research Office, Federation of American Societies for Experimental Biology.
76. Group ESW. 1985. Summary of a report on assessment of the iron nutritional status of the United States population. Am. J. Clin. Nutr., 2: 1318-1330.
77. Hulthén, L. 1998. Effect of a mild infection on serum ferritin concentration -clinical and epidemiological implications. Eur. J. Clin. Nutr., 52: 1-4.
78. Osler, M., Minman, N. & Heitman, B.L. 1998. Dietary and non-dietary factors associated with iron status in a cohort of Danish adults followed for six years. Eur. J. Clin. Nutr., 52: 459-63.
79. Leggett, B.A. 1990. Factors affecting the concentrations of ferritin in serum in a healthy Australian population. Clin. Chem., 36: 1350 1555.
80. Cook, J.D., Skikne, B. & Baynes, R. 1996. The use of transferrin receptor for the assessment of iron status. In: Hallberg LA, Asp N-G, eds. Iron nutrition in health and disease, London: John Libbey & Co.
81. DeMaeyer, E ., Adiels-Tegman, M. & Raystone, E. 1985. The prevalence of anemia in the world. World Health Stat Q., 38: 302-316.
82. Egderton, V.R. 1972. Iron deficiency anemia and physical performance and activity of rats. J. Nutr., 102: 381-400.
83. Finch, C.A. , 1976. Iron deficiency in the rat. Physiological and biochemical studies of muscle dysfunction. J. Clin. Investig., 58: 447-53.
84. Scrimshaw, N.S. 1984. Functional consequences of iron deficiency in Human populations. J. Nutr. Sci. Vit., 30: 47-63.
85. Lozoff, B., Jimenez, E. & Wolf, A. 1991. Long-term developmental outcome of infants with iron deficiency. N. Engl. J. Med., 325: 687-694.
86. Youdim, M.B.H. 1988. Brain iron:Neurochemical and behavioural aspects. New York, Taylor & Francis.
87. Beard, J.L., Connor, J.R. & Jones, B.C. 1993. Iron in the brain. Nutr. Revs., 1: 157-170.
88. Pollitt, E. 1993. Iron deficiency and cognitive function. Ann. Rev. Nutr., 13: 521-37.
89. Brock, J.H. 1994. Iron in infection, immunity, inflammation and neoplasia. In: Brock JH et al., eds. Iron metabolism in health and disease. p. 353-389. London, W.B.Saunders Company Ltd.
90. Bruner, A.B. 1996. Randomised study of cognitive effects of iron supplementation in non-anaemic iron-deficient adolescent girls. Lancet, 348: 992-96.
91. Rowland, T.W. 1988.The effect of iron therapy in the excersice capacity of non-anemic iron-deficient adolescent runners. Am. J. Dis. Child., 142: 165-169.
92. Ballin, A. 1992. Iron state in female adolescents. Am. J. Dis. Child., 146: 803-805.
93. Zhu ,Y.I. & Haas, J.D. 1997. Iron depletion without anemia and physical performance in young women. Am. J. Clin. Nutr., 66: 334-41.
94. Hallberg, L. 1988. Iron balance in pregnancy. In: Berger H, ed. Vitamins and minerals in pregnancy and lactation. p. 115-127. Nestlé Nutrition Workshop Series Vol 16. New York: Raven Press.
95. Hallberg, L. 1992. Iron balance in pregnancy and lactation. In: Fomon SJ., Zlotkin S, eds. Nutritional anemias. New York: Raven Press, Ltd. p.13-25. Nestlé Nutrition Workshop Series, vol 30).
96. Lieberman, E., Ryan, K.J. & Monsen, R.R. 1988. Association pf maternal hematocrit with premature labour. Am. J. Obstet. Gynecol., 159: 107-114.
97. Garn, S.M., Ridella, S.A., Petzold, A.S. & Falkner, F. 1981. Maternal hematological levels and pregnancy outcome. Semin Perinatol., 5: 155-162.
98. Rossander, L., Hallberg, L. & Björn-Rasmussen, E. 1979. Absorption of iron from breakfast meals. Am. J. Clin. Nutr., 32: 2484-2489.
99. Hallberg, L. & Rossander, L. 1982. Bio-availability of iron from Western-type whole meals. Scand. J. Gastroenterol., 17: 151-160.
100. Galan, P. 1990. Iron absorption from typical West African meals containing contaminating Fe. Br. J. Nutr., 64: 541-546.
101. Acosta, A. 1984. Iron absorption from typical Latin American meals. Am. J. Clin. Nutr., 39: 953-962.
102. Rao, B.S.N., Vijayasarathy, C. & Prabhavathi, T. 1983. Iron absorption from habitual diets of Indians studied by the extrinsic tag technique. Indian J. Med.. 77: 648-657.
103. Aung-Than-Batu, Thein-Than & Thane-Toe. 1976. Iron absorption from Southeast Asian rice-based meals. Am. J. Clin. Nutr., 29: 219-225.
104. Hallberg, L. 1974. Iron absorption from Southeast Asian diets. Am. J. Clin. Nutr., 27: 826-836.
105. Hallberg, L. 1977. Iron absorption from Southeast Asian diets. II. Role of various factors that might explain low absorption. Am. J. Clin. Nutr., 30: 539-548.
106. Hallberg, L. 1978. Iron absorption from South-East Asian diets and the effect of iron fortification. Am. J. Clin. Nutr., 31: 1403-1408.
107. Hallberg, L., Björn-Rasmussen, E. & Rossander, L.R.S. 1979. The measurement of food iron absorption in man. A methodological study on the measurement of dietary non-haem-Fe absorption when the subjects have a free choice of food items. Br. J. Nutr. 41: 283-289.
![]()
![]()
![]()
{kind=link}
{kind=link}