![]()
![]()
![]()
Methods
Results
Discussion: floodplain gillnet fisheries
Selected data and information on eight species of economic importance
i. Experimental gillnet fishing
ii. Additional fish sampling
iii. Selected species
iv. Fish identifications
v. Ichthyological parameters
The gear for the experimental gillnet fishing were monofilament nylon gillnets with knot-to-knot mesh sizes of 10, 20, 30, 40, 50, 60, 70, 80 and 100 mm. These nets each had a length of 10 metres and 2 metres hanging depth. On top, they were equipped with a cork line and underneath with a PVC lead line of 1.2 kg/100m. Sets of these gillnets of all the mentioned mesh sizes were assembled, in order of mesh size, to panels of 90 metres total length. Total surface area per panel was thus 180 m2, with 20 m2 of surface per mesh size. This definition of a panel is the unit used for the calculation of the catch per unit effort (CPUE) data. Panels that were used for fishing at the bottom were additionally weighed and marked with buoys.
The sampling procedure was to set three panels at the surface, and to sink one panel for bottom catches. In the scrubland, the bottom panel could for obvious reasons usually not be used. Catches in the forest near the lakeshore were often impossible because of the presence of the submerged vegetation and the almost perpetual wave and wind action. The nets were set from a small man-powered boat.
One habitat was sampled at a time, and all panels were placed in that habitat. The panels were placed from around 0500 hours on, shortly before sunrise, and were soaked for three hours. The limnology data were collected after the nets were placed. As the nets were being recovered, the fish caught were taken from the net and per mesh size and panel collected in marked plastic bags. The bags were kept on ice until analysis of the catch.
All specimens from the experimental catches were identified and measured for weight, standard and total length. CPUE was calculated using the panels as defined higher as standard gear, and expressed as g.panel-1.hour-1 of soaking time (CPUEw) for a weight- based measurement, and as panel-1.hour-1 (CPUEn) for a figure based on the number of specimens.
On a number of occasions, fish were collected from a variety of sources for additional data collection. This was the case with fish sizes or species that were only occasionally caught by the gillnets, or to assess gear characteristics as selectivity. Fish were collected from traps, from additional gillnets or from local markets where fish sold were caught by a variety of gear (traps, harpoons, hooked lines, cast nets, etc.). Data from these fish were used together with those from the experimental gillnet fishing in the calculation of the length-weight relations, for scale collection and for gear selectivity assessment. Unless collected directly from the source, the habitat where the fish originated from was not traced, and no quantification of fishing effort was attempted for any of these additional sources since reliable information was usually not available.
Eight species were selected for a more detailed study of the nature of the relationship between the fishes and the different habitats. The criteria for the selection were the economical importance of each species in the catches of Cambodia and their occurrence in the experimental catches. The selected species are Cirrhinus microlepis, Cyclocheilichthys apogon, Cyclocheilichthys enoplos, Henicorhynchus siamensis, Osteochilus melanopleurus, Paralaubuca typus, Trichogaster microlepis and Trichogaster trichopterus.
Fish identification based on the FAO species guide for the fishes of the Cambodian Mekong (Rainboth, 1996) and all the scientific and Khmer names used here refer to the names as used in the FAO guide. In a number of cases, the information given on morphology and anatomy for a species did not suffice for certain identification. At that point, other information such as geographical, spatial and temporal distribution as well as data on behaviour and reported feeding habits were taken into consideration. Although this was not a problem in this study, regional variations in the use of the Khmer fish names are a potential source of misidentification.
· Weight
All the fish that were analysed in this study were weighed as soon as possible after retrieval of the catch using precision spring balances. When large quantities of one species occurred in the catches, a random sample of 20 specimens were individually weighed and measured; the rest was counted and weighed in bulk.
· Length
Total length and, except for some samples collected early in the study, standard length of all the fish caught were measured to the nearest millimetre, using measurement boards.
· Condition Factor
In isometrically growing fish, the exponent in the power equation describing the relation between the length of a fish and its weight is three. W = q.L3 describes this relationship between the length L and the weight W. In case of allometry, the exponent b in the more general power equation W = qLb will differ from three. In these equations, q is a constant to be determined empirically.
The length-weight relationships were analysed for the eight species studied in more detail by using all the length and weight data collected over the whole year (September 1996-August 1997). The length-weight relationship was established by calculating the least squares fit through the data points of each species using the equation y = qxb, where q and b are constants, y the weight and x the standard length. This length-weight equation is given for each species in the chart with the data points.
Once the general length-weight equation is determined for a species, it is possible to calculate the factor qi for individual fish of known length and weight. The value of the factor qi can be considered a measure for the condition of the fish, supposing that the weight of a fish is directly linked to its condition or “well-being” (King, 1995). The more a fish weighs at a given length, the greater will be its factor qi.
The general length-weight relationship established using the data from the whole year can be used to predict a posteriori the weight for a fish of a given length. It can also be used to predict the mean weight for a sample of fish, given the mean length of the fish in that sample. This would allow monthly values of mean weight to be compared with the general predicted value for fish of the same mean length.
A mean weight-based condition factor (CFw) can be calculated from this:
CFw = (mean weight).(predicted mean weight)-1
The results of this comparison are given for each of the selected species. The condition factors that are thus calculated from monthly samples can allow the detection of monthly variations in the condition of fish. These variations may be related to fluctuations in food availability, reproductive activity of the species, etc. and would need to be completed with additional information for interpretation.
There are some limitations to this technique. The factors in the length-weight equations are estimates of the populations parameters based on a limited sample. For some species, the fit that was found is not very close. The monthly samples of fish are not of the same size, given that they consisted for a large part of the catches of unpredictable sizes of the experimental gillnet fishing. This source of bias was reduced towards the end of the period when every month large samples (more than 100 specimens) of each species were analysed.
· Scalimetry
Scales were collected from the same area on the bodies. The area between the rostral insertion of the dorsal fin and the lateral line was used to collect the scales for analysis. After removal, the scales were conserved in paper envelopes. Prior to mounting between two microscope slides, the scales were cleansed by rubbing between the top of two fingers and rinsing with clean water. Three to eight scales were mounted per slide, and these were then coded. Per specimen, one scale was selected for analysis. Regenerated scales were disregarded.
The scales were inspected for selection and analysed using a microfiche reader enlarging them 40 times. The median radius (distance from the focus to the edge of the rostral field along the median axis of the scale) was measured.
i. Composition of the gillnet catches
ii. Selectivity of gillnets - comparison of experimental gillnet catches with those by other types of gear
iii. Habitat-related results of the experimental gillnet fishing
The species composition and the total number of specimens of all the experimental gillnet fishing together are given in Table 2.7. Based on information from the literature, two other relevant characteristics are listed for each species: their main source of food and the ability of the species to tolerate low levels of dissolved oxygen.
In total, 7 744 specimens of 46 species were caught and identified. Of these, three species could only be identified with certainty to genus level, one species with 11 specimens only by its Khmer name, and two specimens of one species not at all. Nine of the collected species were unique catches with only one specimen, and 16 species could be considered occasional catches with fewer than ten specimens. Between 10 and 100 specimens were caught of each of the ten more common species, while 11 species are abundant in the catches with up to over 2 000 specimens. Two small schooling cyprinids (Paralaubuca typus and Parachela siamensis) make up over half of the total number of fish in the catch.
Table 2.7 Fish species with number of specimens caught during the experimental gillnet fishing. Codes: P: plankton community, B: benthic community, A: Aufwuchs community, H: plant community, C: prey community,?: unknown (see text)
|
Species |
Total number caught with gillnets |
Percentage of total catch by number |
Percentage of total catch by weight |
Main food source(s) |
Resistance to low dissolved oxygen
concentrations |
|
Anabas testudineus |
33 |
0.4 |
0.9 |
C |
very high[1] |
|
Arius truncatus |
1 |
0.0 |
0.0 |
C |
low (high?) |
|
Barbodes gonionotus |
8 |
0.1 |
0.1 |
H/A |
low (high?) |
|
Cirrhinus microlepis |
3 |
0.0 |
0.8 |
P/H/C |
low |
|
Coilia lindmani |
402 |
5.2 |
4.3 |
P |
low |
|
Crossocheilus reticulatus |
1 |
0.0 |
0.0 |
A/P |
low |
|
Cyclocheilichthys apogon |
362 |
4.7 |
3.4 |
P |
low |
|
Cyclocheilichthys enoplos |
9 |
0.1 |
1.7 |
C/B |
low |
|
Dangila lineata |
32 |
0.4 |
0.9 |
P/A |
low |
|
Dangila spilopleura |
9 |
0.1 |
0.1 |
P/A |
low |
|
Esomus longimanus |
12 |
0.2 |
0.1 |
P/C |
low |
|
Esomus metallicus |
15 |
0.2 |
0.2 |
P/C |
low |
|
Hampala dispar |
2 |
0.0 |
0.2 |
C |
low |
|
Hampala macrolepidota |
16 |
0.2 |
2.1 |
C |
low |
|
Hemipimelodus borneensis |
1 |
0.0 |
0.0 |
B |
high |
|
Henicorhynchus siamensis |
276 |
3.6 |
8.0 |
H/P/A |
low |
|
Hypsibarbus lagleri |
12 |
0.2 |
0.9 |
P/B |
low |
|
Kryptopterus cheveyi |
6 |
0.1 |
0.1 |
C |
low |
|
Kryptopterus schilbeides |
2 |
0.0 |
0.0 |
C |
low |
|
Labeo erythropterus |
1 |
0.0 |
0.1 |
A/P |
low |
|
Luciosoma bleekeri |
1 |
0.0 |
0.0 |
C |
low |
|
Lycothrissa crocodilus |
46 |
0.6 |
0.9 |
C |
low |
|
Macrognathus sp. |
1 |
0.0 |
0.0 |
B |
low |
|
Mystus mysteticus |
14 |
0.2 |
0.2 |
A |
low |
|
Notopterus notopterus |
2 |
0.0 |
0.1 |
C |
very high |
|
Osteochilus hasselti |
105 |
1.4 |
4.2 |
H/A |
low |
|
Osteochilus melanopleurus |
8 |
0.1 |
1.5 |
H/P |
low |
|
Oxyeleotris marmorata |
1 |
0.0 |
0.0 |
C |
high |
|
Pangasius sp. (2) |
2 |
0.0 |
1.6 |
B |
high |
|
Parachela siamensis |
2 358 |
30.4 |
21.4 |
P |
low |
|
Paralaubuca typus |
1 897 |
24.5 |
16.3 |
P |
low |
|
Polynemus longipectoralis |
17 |
0.2 |
0.4 |
C |
low |
|
Pristolepis fasciata |
9 |
0.1 |
0.2 |
H |
high |
|
Pseudambassis notatus |
157 |
2.0 |
0.4 |
C |
high |
|
Puntius brevis |
563 |
7.3 |
4.0 |
B/H |
high |
|
Rasbora aurotaenia |
7 |
0.1 |
0.1 |
C |
low |
|
Rasbora caudimaculata |
2 |
0.0 |
0.2 |
C |
low |
|
Rasbora sp. |
1 |
0.0 |
0.0 |
P (?) |
low |
|
Systomus orphoides |
1 |
0.0 |
0.0 |
H |
low |
|
Thynnichthys thynnoides |
2 |
0.0 |
0.1 |
A/P |
low |
|
Trichogaster microlepis |
514 |
6.6 |
9.2 |
P/C |
very high |
|
Trichogaster pectoralis |
2 |
0.0 |
0.1 |
H (?) |
very high |
|
Trichogaster trichopterus |
454 |
5.9 |
4.0 |
P/C |
very high |
|
Undetermined # 1 |
2 |
0.0 |
0.0 |
? |
? |
|
Undetermined # 2 |
11 |
0.1 |
0.1 |
? |
? |
|
Xenentodon cancila |
364 |
4.7 |
11.1 |
C |
high |
|
Total |
7 744 |
99.7[2] |
100.0 |
|
|
Table 2.8. Relevant food categories and their composition
|
Category |
Composition |
|
Plankton community |
· phytoplankton |
|
Benthic community |
· mud and associated
micro-organisms |
|
Aufwuchs community of epilithic/epiphytic organisms and the
periphyton |
· epiphytic or epilithic
algae |
|
Plant community |
· plants including filamentous
algae and submerged, floating or emergent vascular plants |
|
Prey community |
· neuston, surface living
insects and larvae at the water/air interface |
Source: Welcomme, 1985.The assessment of tolerance for low dissolved oxygen concentrations is made based on the known presence of anatomical or physiological adaptations to use atmospheric oxygen for breathing or of behavioural adaptations to use the better oxygenated surface layers of the water. Unambiguous for those species with clear adaptations of either type, there is a degree of arbitration in the classification of species’ resistance of low oxygen conditions as low or high. This classification was based in this case mainly on information on the timing of movements in and out of the flooded areas and the associated changes in overall water quality. Most species encountered are obligate aquatic respirators, and only a small number showed higher tolerance for low dissolved oxygen levels.
On a number of occasions, samples were taken from commercial traps in the same general area as where the experimental gillnet fishing was done (for timing and location see Annex 3). Of the 46 species caught by the experimental gillnets, 27 were also caught by the traps. The average weight for species caught in common by gillnets and traps is given in Table 2.9. Where sufficiently large numbers (> 5) of specimens were available, a Student’s t-test was done to assess whether the difference in the average weight for fish from the different gear is statistically significant (a = 0.05). This is the case for half of the species for which a t-test could be done (Table 2.9). For only one species (Osteochilus hasselti) the average weight in the gillnet catches is significantly higher than that of the trap catches, but only by about 16 percent; in all other cases there is no significant difference or the average weight from the traps is higher than that from the gillnets. There is large variation in the factor by which the significantly different average weights from both gear differ: fish from traps are 1.18 to 25.8 times heavier than same-species fish from the gillnets.
Table 2.9 Species common to the experimental gillnet catches and the samples collected from traps. All habitats, September 1996-August 1997
|
Species |
Total number caught with gillnets |
Total number caught with traps |
Average weight (g) in gillnets |
Average weight (g) in traps |
Difference significant (p < 0.05) |
|
Anabas testudineus |
33 |
1 |
19.0 |
26.0 |
- |
|
Barbodes gonionotus |
8 |
15 |
4.4 |
113.5 |
yes |
|
Cirrhinus microlepis |
3 |
37 |
183.3 |
139.3 |
- |
|
Coilia lindmani |
402 |
1 |
7.2 |
8.8 |
- |
|
Cyclocheilichthys apogon |
362 |
130 |
6.2 |
12.1 |
yes |
|
Cyclocheilichthys enoplos |
9 |
448 |
124.1 |
72.9 |
no |
|
Dangila lineata |
32 |
207 |
18.9 |
16.1 |
no |
|
Hampala dispar |
2 |
1 |
61.0 |
32.0 |
- |
|
Hampala macrolepidota |
16 |
2 |
88.8 |
39.0 |
- |
|
Henicorhynchus siamensis |
276 |
83 |
19.2 |
50.4 |
yes |
|
Hypsibarbus lagleri |
12 |
120 |
48.9 |
97.2 |
yes |
|
Kryptopterus schilbeides |
2 |
11 |
6.6 |
6.4 |
- |
|
Lycothrissa crocodilus |
46 |
32 |
12.5 |
7.4 |
no |
|
Mystus mysteticus |
14 |
2 |
8.2 |
20.5 |
- |
|
Osteochilus hasselti |
105 |
95 |
26.7 |
23.1 |
yes |
|
Osteochilus melanopleurus |
8 |
76 |
128.4 |
130.2 |
no |
|
Oxyeleotris marmorata |
1 |
4 |
30.8 |
80.3 |
- |
|
Parachela siamensis |
2 358 |
13 |
6.0 |
5.7 |
no |
|
Paralaubuca typus |
1 897 |
251 |
5.7 |
6.7 |
yes |
|
Pristolepis fasciata |
9 |
11 |
15.8 |
22.8 |
no |
|
Pseudambassis notatus |
157 |
1 |
1.8 |
1.8 |
- |
|
Puntius brevis |
563 |
70 |
4.7 |
11.0 |
yes |
|
Rasbora aurotaenia |
7 |
1 |
6.9 |
4.2 |
- |
|
Thynnichthys thynnoides |
2 |
11 |
34.5 |
74.8 |
- |
|
Trichogaster microlepis |
514 |
52 |
11.9 |
12.9 |
no |
|
Trichogaster trichopterus |
454 |
43 |
5.9 |
11.6 |
yes |
|
Xenentodon cancila |
364 |
107 |
20.2 |
19.5 |
no |
|
Total |
7 656 |
1 825 |
|
|
|
The species caught only by traps are rather large in comparison with the average catch by the experimental gillnets; the average standard length for specimens of all the 11 species is more than 100 mm. None of the species only caught by traps are feeding predominantly on the plankton community. Benthic community feeders are as common as species feeding on prey (Table 2.10).
Table 2.10 Fish species caught exclusively by the traps. Codes: P: plankton community, B: benthic community, A: Aufwuchs community, H: plant community, C: prey community,?: unknown (see text)
|
Species |
Total number caught in traps |
Main food source(s) |
Resistance to low dissolved oxygen
concentrations |
|
Belodontichthys dinema |
1 |
C |
low |
|
Heterobagrus bocourti |
4 |
B |
low |
|
Hyporhamphus limbatus |
9 |
C |
high |
|
Micronema apogon |
4 |
C |
low |
|
Monotreta cambodgiensis |
3 |
B/H |
low |
|
Morulius chrysophekadion |
47 |
B/P |
low |
|
Mystus filamentus |
1 |
C |
low |
|
Mystus multiradiatus |
2 |
A |
low |
|
Pangasianodon hypophthalmus |
61 |
C/B/H |
low |
|
Parambassis wolffi |
30 |
C/B |
high |
|
Undetermined # 3 |
1 |
? |
? |
|
|
Habitat |
|||||
|
rice field |
forest |
|||||
|
n |
% n |
% w |
n |
% n |
% w |
|
|
Respiration |
|
|
|
|
|
|
|
Exclusive aquatic |
21 |
95 |
97 |
17 |
100 |
100 |
|
Facultative air |
2 |
5 |
3 |
1 |
0 |
0 |
|
Feeding community |
|
|
|
|
|
|
|
Plankton |
9 |
64 |
53 |
7 |
44 |
50 |
|
Benthic |
4 |
10 |
10 |
5 |
78 |
79 |
|
Aufwuchs |
3 |
32 |
43 |
6 |
8 |
7 |
|
Plants |
6 |
19 |
25 |
7 |
28 |
26 |
|
Prey |
13 |
24 |
27 |
8 |
58 |
53 |
|
Total |
23 |
|
|
18 |
|
|
The results from the experimental gillnet fishing split between surface panels and bottom panels are given in Table 2.12. The results of fishing with additional local gillnets are presented in the same table.
Table 2.12 Catch composition (numbers) by mesh sizes for surface, bottom and panels of local gillnets
|
Mesh size (mm) |
Surface panels |
Bottom panels |
Local gillnets |
|||
|
% of catch (n) within the panel |
% of total exp. gillnets catch (n) |
% of catch (n) within the panel |
% of total exp. gillnets catch (n) |
Number of species |
% of catch (n) within the panel |
|
|
10 |
91.2 |
76.8 |
90.0 |
14.2 |
- |
- |
|
20 |
7.7 |
6.5 |
9.8 |
1.6 |
20 |
94.5 |
|
30 |
1.1 |
0.9 |
0.2 |
0.0 |
9 |
5.5 |
|
40 |
0.0 |
0.0 |
0.0 |
0.0 |
0 |
0.0 |
|
50 |
0.0 |
0.0 |
0.0 |
0.0 |
0 |
0.0 |
|
60 |
0.0 |
0.0 |
0.0 |
0.0 |
- |
- |
|
70 |
0.0 |
0.0 |
0.0 |
0.0 |
- |
- |
|
80 |
0.0 |
0.0 |
0.0 |
0.0 |
- |
- |
|
100 |
0.0 |
0.0 |
0.0 |
0.0 |
- |
- |
Over three quarters of all fish caught in the experimental gillnet fishing were collected from the smallest mesh size section of the surface panels. Over 90 percent of the fish caught in a panel was found in the smallest mesh size, 10 mm for the experimental gillnets and 20 mm for the local gillnets. No fish were caught in the sections of the panels with mesh sizes between 50 and 70 mm, or in the 100 mm section. The catches in the 40 and 80 mm sections consist both of a single fish.
Figure 2.3 Number of species common and specific for the surface and bottom panels in the gillnet catches (upper) and for the surface panels of the experimental gillnets and the local gillnets (lower). The numbers indicate the number of species for each category


Bottom panels caught fewer species than panels at the surface (Fig. 2.2), but mostly they were the same species. The local gillnets produced 22 species, which is less than half of the number of species caught in the experimental gillnets; however, four species were only found in the former.
The fishing effort was not equal for the different panels. Bottom panels were hardly used in certain habitats (scrubland, forest) because they would become entangled in the vegetation. In general, when bottom panels were used, only one panel was sunk to the bottom instead of three as used for the surface. The local gillnet panels were used occasionally, when conditions allowed, one panel at the time, and only on the surface, at a distance away from the experimental panels in the same habitat type.
The largest number of species in the experimental gillnet catches was found in the floodplain pool, while scrubland, lotus field and the coastal lake all recorded the lowest number of species. The very low number of the forest is not very representative since it is based on a single sampling. The floodplain pool catches also hold the largest number of facultative air breathing species. The proportion of air breathing species in the total number does not vary much for any of the habitats.
Table 2.13 Number of species (n) for each habitat and their percentages by number of specimens (%n) and weight (% w) in the gillnets catch, categorized by their facultative air breathing ability and by food community.
|
|
Habitat |
|||||||||||
|
Scrubland |
Grassland |
Floodplain pool |
Lotus field |
|||||||||
|
n |
% n |
% w |
n |
% n |
% w |
n |
% n |
% w |
n |
% n |
% w |
|
|
Respiration |
|
|
|
|
|
|
|
|
|
|
|
|
|
Exclusive aquatic |
13 |
71 |
76 |
22 |
79 |
87 |
23 |
90 |
89 |
13 |
95 |
92 |
|
Facultative air |
3 |
29 |
24 |
3 |
21 |
13 |
4 |
10 |
11 |
3 |
5 |
8 |
|
Feeding community |
|
|
|
|
|
|
|
|
|
|
|
|
|
Plankton |
7 |
82 |
67 |
12 |
85 |
69 |
10 |
72 |
60 |
7 |
66 |
46 |
|
Benthic |
1 |
5 |
2 |
2 |
1 |
2 |
3 |
18 |
15 |
3 |
8 |
30 |
|
Aufwuchs |
4 |
12 |
25 |
6 |
8 |
20 |
5 |
6 |
9 |
2 |
4 |
6 |
|
Plants |
4 |
16 |
24 |
6 |
9 |
25 |
5 |
23 |
19 |
5 |
13 |
11 |
|
Prey |
8 |
40 |
48 |
11 |
31 |
39 |
14 |
19 |
33 |
7 |
25 |
29 |
|
Total |
16 |
|
|
25 |
|
|
27 |
|
|
16 |
|
|
|
|
Habitat |
|||||||||||
|
Rice field |
Forest |
Lake, coastal |
Lake, pelagic |
|||||||||
|
n |
% n |
% w |
n |
% n |
% w |
N |
% n |
% w |
n |
% n |
% w |
|
|
Respiration |
|
|
|
|
|
|
|
|
|
|
|
|
|
Exclusive aquatic |
23 |
96 |
95 |
0 |
0 |
0 |
14 |
96 |
92 |
21 |
86 |
76 |
|
Facultative air |
3 |
4 |
5 |
3 |
100 |
100 |
2 |
4 |
8 |
3 |
14 |
24 |
|
Feeding community |
|
|
|
|
|
|
|
|
|
|
|
|
|
Plankton |
13 |
89 |
83 |
1 |
44 |
29 |
9 |
86 |
79 |
12 |
89 |
81 |
|
Benthic |
3 |
3 |
4 |
0 |
|
|
2 |
8 |
5 |
4 |
4 |
12 |
|
Aufwuchs |
6 |
4 |
9 |
0 |
|
|
3 |
3 |
12 |
4 |
3 |
9 |
|
Plants |
5 |
6 |
10 |
1 |
13 |
22 |
4 |
11 |
18 |
5 |
6 |
14 |
|
Prey |
10 |
9 |
13 |
2 |
88 |
78 |
8 |
8 |
17 |
12 |
21 |
43 |
|
Total |
26 |
|
|
3 |
|
|
16 |
|
|
24 |
|
|
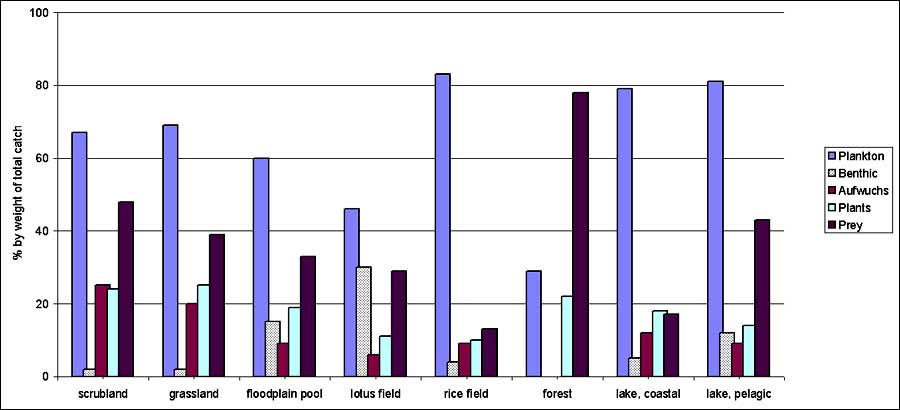
Note: The totals for the feeding communities do not add up to 100 as the categories are not mutually exclusiveSpecies feeding on plankton communities make up the largest group for all habitats, both by number of specimens as by weight. In some habitats (rice field and the two lake habitats), this dominance is more pronounced than in others. The highest number of prey eating species was found in the catches from the scrubland, with half of the species fitting in this category. Also in the floodplain pool, the benthic community plays a considerable role as a food source. Herbivorous species take up only the third rank in most habitats; they are somewhat more present in the coastal area of the lake and in the floodplain pool. The benthic and Aufwuchs communities are of less importance in most habitats. However, fish feeding on the benthic community make up almost one third of the weight of the fish caught in the lotus field, but these are mostly big fish as their share in the total number is much smaller. In the floodplain pool, they make up 15-18 percent of the catch. Species feeding on the Aufwuchs community make up a quarter and one fifth of the weight of the catch in the scrubland and the grassland respectively.
Some caution is required with the interpretation of these feeding community results. The feeding habits are those reported for the encountered species in a variety of ecosystems, but almost no reports on feeding habits of these species in the studied habitats are available. Their classification as belonging to one or more categories does not imply that this is effectively the case in the habitats of the floodplain, or at all times and stages of their development.
Figure 2.4 Number of species in the experimental gillnet catches per habitat and per month
Species composition of the experimental gillnet fishing per habitat shows considerable fluctuations over time. The pelagic lake has the highest number of species present in the catches at any time but its total number of species is lower than that of e.g. the rice field. (Fig. 2.4).
Over 90 percent of all 7 744 specimens and two thirds of the total ichthyomass of 66.459 kg were caught in the section of the panels with the smallest mesh size. No fish were caught in the 50, 60, 70 or 100 mm mesh size sections, and the catches in the 40 and 80 mm sections consist each of one rather big fish.
The average soaking time for the experimental gillnets was three hours and 18 minutes. On average, and with little variation, the nets were set at 0527 hours and retrieved around 0845 hours.
The average catch per effort results are calculated for each habitat based on all the available data from all experimental gillnet fishing in that habitat. The results, based on both total weight of the catch (CPUEw) and on the number of fish specimens in the catch (CPUEn), are given in Fig. 2.6 and 2.7. In general, there is large variation in CPUE. In Table 2.14, the calculated average CPUE-values are presented, together with the total number of panels used in the sampling (each time a panel was used was counted as a unit) and the extreme values for the CPUE that are found. As remarked earlier, the number of samples is rather small for some habitats, especially for the forest.
Table 2.14 CPUE for the habitats and its variation. Units are g.panel-1.hour-1 (CPUEw) and panel-1.hour-1 (CPUEn)
|
Habitat |
Surface panels |
||||||
|
CPUEw |
CPUEn |
Number of panels |
|||||
|
mean |
min. |
max. |
mean |
min. |
max. |
||
|
Scrubland |
94.8 |
21.5 |
261.1 |
11.6 |
1.8 |
35.8 |
24 |
|
Grassland |
148.8 |
2.7 |
374.6 |
15.3 |
0.5 |
29.3 |
24 |
|
Floodplain pool |
106.4 |
11.6 |
218.1 |
15.5 |
0.9 |
29.3 |
30 |
|
Lotus field |
93.5 |
19.0 |
239.1 |
8.3 |
2.4 |
17.7 |
9 |
|
Rice field |
67.3 |
16.3 |
129.2 |
8.0 |
2.4 |
12.9 |
6 |
|
Forest |
7.7 |
1.3 |
12.6 |
0.5 |
1.3 |
0.9 |
3 |
|
Lake, coastal |
189.1 |
5.3 |
732.9 |
19.1 |
1.0 |
65.0 |
18 |
|
Lake, pelagic |
125.9 |
16.6 |
351.9 |
11.9 |
1.8 |
32.9 |
21 |
|
Habitat |
Bottom panels |
||||||
|
CPUEw |
CPUEn |
Number of panels |
|||||
|
mean |
min. |
max. |
mean |
min. |
max. |
||
|
Scrubland |
53.9 |
47.4 |
60.3 |
2.4 |
1.6 |
3.3 |
2 |
|
Grassland |
74.0 |
5.3 |
208.2 |
8.6 |
1.2 |
26.8 |
7 |
|
Floodplain pool |
55.0 |
0.0 |
140.6 |
9.9 |
0.0 |
24.9 |
8 |
|
Lotus field |
56.4 |
3.5 |
149.5 |
5.6 |
0.6 |
14.2 |
3 |
|
Rice field |
30.5 |
17.0 |
44.0 |
4.0 |
1.8 |
6.2 |
2 |
|
Forest |
27.8 |
- |
- |
3.3 |
- |
- |
1 |
|
Lake, coastal |
168.0 |
12.9 |
544.6 |
18.7 |
2.4 |
58.8 |
6 |
|
Lake, pelagic |
102.4 |
8.0 |
249.2 |
9.7 |
0.3 |
25.6 |
7 |
The seasonal variation in CPUE in the floodplain pool follows different patterns depending on the weight or the numbers as basis for calculation. It is clear however that there is a sharp drop in CPUE in August (Fig. 2.8).
The experimental gillnet catches were dominated both in terms of numbers as by species by small cyprinids. Most fish were caught in the smallest mesh sizes. Species of all feeding guilds were present in all habitats but there were considerable differences in the importance each group has in the total catch. The largest number of species was found in the floodplain pool; however, it is not clear whether this is due to the special characteristics of this habitat as discussed above, or simply to the fact that this habitat was the most sampled.
The CPUE values of the experimental gillnet panels do not provide a realistic estimate of the productivity of gillnets in these waters. Ninety percent of the numbers were caught in the section of the gillnets with the smallest mesh size of 10 mm, which made up only one ninth of the total surface of the panels. The CPUE of gillnets consisting only of such mesh size would probably be about ten times higher.
The large variation in CPUE within habitats is in part explained by the presence or absence of schooling species. The number of samples and the time between samples did not allow a rigorous analysis of the sources of variation between the CPUEs as recorded in the different habitats. The differences in CPUE also seem related to the degree of diversity within the habitats: the lowest production was found in the habitats where diversity is actively reduced by agricultural activities (rice and lotus fields).
The floodplain pool shows a sharp drop in CPUE for August. This coincides with the flooding of the pool and is probably due to the “dilution” of the fish present in the pool in the low-in-fish lake water, and to the abrupt change in water quality, which may have resulted in fish mortality.
Gillnets are highly selective fishing gear. The results from the experimental gillnet fishing for numbers of species and CPUE for the different habitats therefore only provide information on the composition of the gillnet catches and the productivity of these gear. They do not allow drawing conclusions about species diversity in the different habitats, or about the potential of such habitats for use of other gear, and about the effects of the conversion of habitat types on catches and biodiversity.
The selectivity of the gillnets as compared with other gear is not only in terms of species but also in terms of size of fish. The composition of the catches based on the feeding communities for the species is shown in Fig. 2.9. Details of the catch composition for all the habitats are discussed in the section on habitats. The two habitats for which the species composition in terms of feeding communities can be compared show considerable differences depending on the gear (Fig. 2.10).
Facultative air breathing species are present in all habitats. The species with higher tolerance to low dissolved oxygen levels and the facultative air breathing species have significant advantages in taking up niches that are created in the freshly flooded floodplain areas. The adaptations to tolerate low levels of dissolved oxygen allow these species to occupy specific niches in the flooded ecosystem that offer particular advantages for reproduction and feeding, opportunities for geographical distribution and protection from predation by less adapted predator fish species (Welcomme 1985, Rainboth 1996, Jobling 1995).
The total number of species encountered in any given habitat is much higher than the number of species found in the same habitat at any given time. There are shifts in the species composition throughout the season, shifts that are closely linked to the water quality in the floodplain habitats and to the migration habits of many species. For the floodplain pool for instance, a total of 27 species was recorded; however, no more than 15 species were found together at any time. The pelagic area of the lake has the highest number of species present in the catches at any time but the total number of species is lower than that of for instance, the rice field. This indicates that the variation in the species composition over time is not the same in all habitats. The difference in species numbers caught with bottom panels and local gillnets is probably attributable to the difference in soaking time of these two gear rather than to specific selective capacity. However, the species composition of the fish population near the bottom may differ from that near the surface.
There is evidence that vegetable detritus is contributing only for a small amount to the growth of at least some groups of detritivorous fish. Most of the carbon constituting these fish originates from phytoplankton (Forsberg et al., 1993; Hamilton et al., 1992), even if phytoplankton constitutes only a small part of their diet. This has been attributed by some authors to the poor digestibility of the vegetable detritus.
i. Cirrhinus microlepis (Sauvage, 1878)
ii. Cyclocheilichthys apogon (Valenciennes, 1842)
iii. Cyclocheilichthys enoplos (Bleeker, 1850)
iv. Henicorhynchus siamensis (deBeaufort, 1927)
v. Osteochilus melanopleurus (Bleeker, 1852)
vi. Paralaubuca typus (Bleeker, 1865)
vii. Trichogaster microlepis (Günther, 1861)
viii. Trichogaster trichopterus (Pallas, 1770)
During the experimental gillnet fishing a substantial amount of data was collected on the biology and the ecology of many species. Mostly, this was an outcome of the activity itself, but in some cases additional specimens were collected for analysis from a variety of sources (local markets, intermediaries, traps, etc.) in Siem Reap. This was done in particular for economically important species that were only occasionally present in the experimental catches.
In this section, the data collected from all sources on fish biology and ecology are presented for the eight species. In addition to the original data, information from the literature is presented, though without the ambition of providing a review of each species. The purpose is rather to provide a fuller picture of these species and on their relationship with the habitats, which are the subject of this study.
The economic importance of the fish in Cambodia has been discussed in the introduction. Table 2.15 gives an overview of the important species of the medium and large-scale fisheries. Three species that are not in this list have been selected to be included here because of their importance in gillnet fisheries.
Table 2.15 Medium and large-scale fisheries. Species composition (top-ten species only) and value of the 1995/96 season (after Deap et al., 1998). Species names and Khmer names as given in original document
|
|
Species |
Percentage of total catch |
|
|
By weight |
By value |
||
|
1 |
Trey riel (Henicorhynchus spp.) |
21 |
9 |
|
2 |
Trey chadaur (Channa micropeltes) |
9 |
19 |
|
3 |
Trey chhkauk (Cyclocheilichthys enoplos) |
9 |
8 |
|
4 |
Trey khnong veng (Dangila spp.) |
6 |
2 |
|
5 |
Trey krum (Osteochilus spp.) |
4 |
2 |
|
6 |
Trey proul (Cirrhinus microlepis) |
3 |
4 |
|
7 |
Trey pra (Pangasius spp.) |
3 |
3 |
|
8 |
Trey chhpin (Barbodes gonionotus) |
3 |
2 |
|
9 |
Trey sluk russey (Paralaubuca typus) |
3 |
1 |
|
10 |
Trey raws (Channa striata) |
2 |
6 |
|
|
Total |
63 |
56 |
Figure 2.11 Cirrhinus microlepis (from Vittayanon, 2000)
Description
Family Cyprinidae, genus Cirrhinus (Oken, 1817). This family is the most developed one in Cambodia in terms of numbers of genera, species and number of individuals.
Cirrhinus microlepis is known by two different names in Khmer: trey krawlang for juveniles and trey proul for adult specimens.
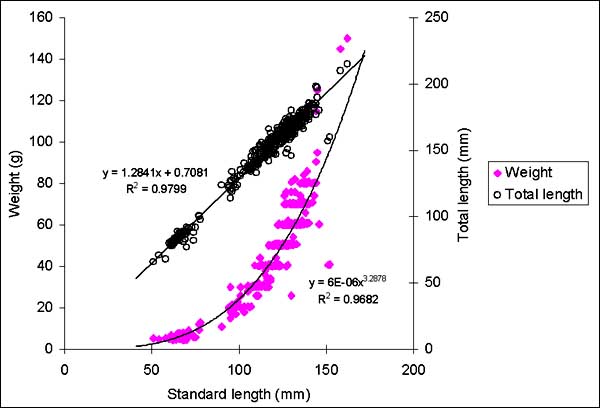
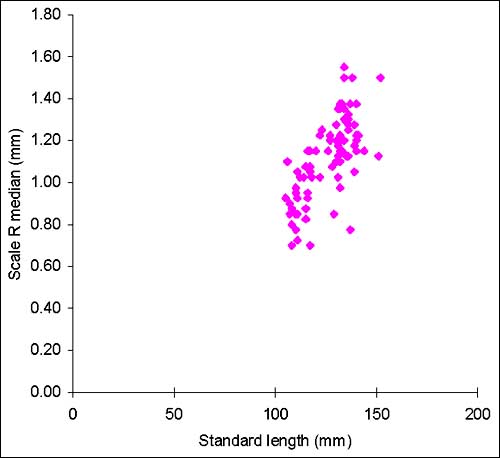
Almost all 331 fish considered for this analysis came from traps in the lake and the floodplain near Siem Reap. Only two specimens were caught in the experimental gillnet fishing. The least squares power trend line of the standard length-weight plot (Fig. 2.12) shows that the growth of the fish analysed is nearly isometric. Maximum total length for the species is reported to be 65 cm (FishBase, 2000). Fig. 2.13 shows that there is large variation in the median radius of the scales for fish of the same standard length. The lifespan of Cirrhinus microlepis is believed not to exceed 11 years (Nguyen and Nguyen, 1991). The species shows slow growth after the seventh year.
Figure 2.13 Cirrhinus microlepis. Body length and scales median radius (n = 109)
Feeding ecology
Cirrhinus microlepis is reported to be essentially vegetarian (leafy plant matter and phytoplankton) but also to eat insects, shrimps and worms. Bardach (1959) classifies the species as microphage, feeding on algae and plant fragments. Its digestive tract shows characteristics that meet the requirements of a herbivorous diet. Nguyen and Nguyen (1991) report that 94-99 percent of gut contents is made up of organic detritus, and the remainder of phytoplankton.
Flooded vegetation provides an important feeding ground for this species. Shifts in its diet composition towards phytoplankton are likely to occur in function of the availability of leafy matter, which is mostly absent during the dry season period.
The condition factor (Fig. 2.14) does not show much variation throughout the year for the sampled fish, except for the four specimens in January 1997 that were markedly lighter than what could be expected based on their length.
Reproductive ecology
All the fish observed for sexual maturity in the markets of Siem Reap from June to September 1997 were immature and in most cases, sex was indeterminable. Cirrhinus microlepis is believed to spawn in the northern Mekong rapids from Sambor to the Khone falls. The fish move with the Mekong floods and re-enter the Tonle Sap mainly for feeding in the rich feeding grounds of the inundated areas. When the water recedes (December to March), they leave the Tonle Sap lake and channel in waves and migrate up the Mekong. Like in the case of many Cambodian migrating fishes, this migration coincides with the waxing moon (Bardach, 1959). There are reports that Cirrhinus microlepis shows four-year cycles of abundance. It appears that clearer water and bottoms other than those that can be found in the Tonle Sap are required for their spawning. The age of sexual maturation and first spawning is reported to be the third or forth year (Nguyen and Nguyen, 1991).
Fisheries
This fish is mostly caught with seines, traps and hook and line. It is almost completely absent from the experimental gillnet catches, possibly due to its ability as a fast swimmer, capable of jumping many feet in the air on order to clear obstacles. Its catches are reported to be declining. Most of the catch of Cirrhinus microlepis is done in the dai fisheries during its spawning migration, and with traps. Fish caught with dais or traps are at times kept alive in cages for later sale. They are usually marketed fresh and sometimes dried and salted (FishBase, 2000).
Van Zalinge and Thuok (1999) write that the catch of medium-sized fishes such as Cirrhinus microlepis has gone down due to overfishing.
Potential for aquaculture
There is contradictory information from the literature about the ability of Cirrhinus microlepis to persist in impoundments. It is not used in aquaculture in Siem Reap, and fish caught in the lake are immediately marketed. It is not mentioned in the list of major aquaculture species in Cambodia (Nam and Thuok, 1999). Recent studies in Lao PDR (Chapman* et al. 1996) show progress in the induced breeding of this species using brood stock caught from the Mekong. Progress is also reported on the techniques for growing Cirrhinus microlepis fry obtained from induced breeding in earthen ponds. The size of this fish, its presumed diet, the fact that it will sexually mature in ponds and the high appreciation of the fish as food make that Cirrhinus microlepis has potential for use as an indigenous species in aquaculture.
* Lamberts:Importance
Page: 4 Get this complete reference from Siem Reap
This fish is important in the fisheries of Cambodia. Cirrhinus microlepis makes up 3 percent of the total catch volume of middle and large-scale fisheries (Deap et al., 1998) and as such is the sixth most important species for inland fisheries. It is one of the most popular food fish, much appreciated because of its attractive coloration and graceful lines as well as its food quality (Smith, 1945). Cirrhinus microlepis is one of the more expensive fish. It is sold at about 2 500 Riel/kg in markets along the lake. It is available from the lake throughout most of the year, except during the low-water period when most fish have migrated to the Mekong for reproduction.
Conclusion
The importance of the flooded area for Cirrhinus microlepis lies in the feeding opportunities it offers. Being primarily detritivorous, habitats with large standing crops of vegetation in the flood area will offer the most opportunities. For this species, preservation of the scrubland and forest vegetation is to be given priority as a management measure at the level of the Tonle Sap lake and canal. The other habitats in the flood area seem to offer less feeding potential for this species. In the floodplain pools and the permanent lake, Cirrhinus microlepis has to rely mostly on phytoplankton for feeding. It is probably not among the first fish to enter the inundated areas but does so during the third phase of flooding (see Part I). Unhindered migration up and down the Mekong is essential for its survival as a dominant species in the Tonle Sap.
Figure 2.15 Cyclocheilichthys apogon (from Vittayanon, 2000)
Description
This cyprinid species belongs to the genus Cyclocheilichthys (Bleeker, 1859), which is well represented in the ichthyofauna of Cambodia. Some species in this taxon are abundant and form an important element in the food supply of the people living around the Tonle Sap lake and the channel. Cyclocheilichthys apogon can be distinguished from the other species of this genus in Cambodia by the absence of barbels (hence the FAO name “beardless barb”). The Khmer name for it used in Siem Reap is trey srawka kdam or trey kros.
Rainboth (1996) mentions Cyclocheilichthys apogon attaining in Cambodia a maximum length of 15 cm, although the species is reported to grow up to 25 cm in Malaysia and India (FishBase, 2000). The standard length of the specimens found in this study varied between 2.7 and 10.2 cm.
Figure 2.17 Cyclocheilichthys apogon. Body length and scales median radius (n = 180)
Feeding ecology
Cyclocheilichthys apogon is reported to be microphage, feeding on plankton, zoobenthos and crustaceans. It is typically found around surfaces, such as plant leaves, branches and roots, where it browses for food.
Reproductive ecology
This species uses the flood area not only for feeding but specifically also for breeding, which occurs late in the high-water season, as water levels peak and begin to decline. Rainboth (1996) speculates that by breeding late in the flooding (September to October), predation by species that move back to permanent water immediately at the onset of water recession, could be avoided.
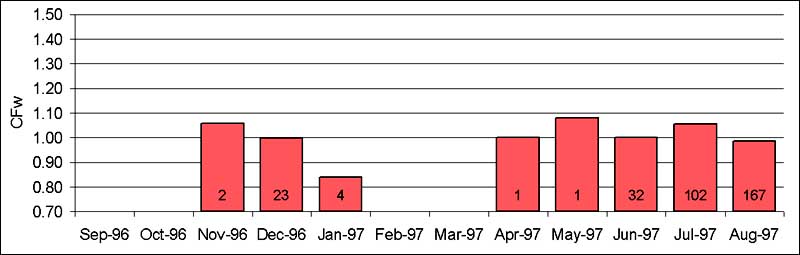
Fish collected during a period of time that are on average heavier than expected by calculation for the same period from the general length-weight relationship, score CFw-values of higher than one. The condition factor shows a sharp decrease in July and August (Fig. 2.18). This coincides with and is probably due to the spawning of the females. Gonads make up a large part of their body mass in the weeks before spawning. Fig. 2.19 shows the gonad weights for female fish around that time. Gonads then made up as much as 30 percent of the body mass of female specimens. For all fish in these samples, average sex ratio (m:f) was 1.10.
Fisheries
Cyclocheilichthys apogon is caught with seines, set nets and traps. The species is abundantly present in the experimental gillnet catches, and is mostly used to make prahoc or marketed fresh for direct consumption. Cyclocheilichthys apogon is available the whole year round. It performs a lateral migration into the flooded area, into both flooded forest as non forested floodplains (Rainboth, 1996).
Potential for aquaculture
This species is known to survive in impoundments and offers thus possibilities as an indigenous aquaculture species. With about 1 000 Riel/kg, it is in the middle-price range.
Importance
The species is indigenous in Cambodia and widely distributed in Southeast Asia. It is an important species in the fisheries of the Tonle Sap, making up about 1 percent by weight of the total catches by middle and large-scale fisheries (Deap et al., 1998).
Conclusion
The available information shows that scrubland and forest are the habitats offering the best opportunities for Cyclocheilichthys apogon. Floodplain pools seem to have few features that are particularly attractive to this species. Lotus fields offer potentially much food but they are readily accessible for migrating or foraging species and do not provide much shelter for fry or adults.
Figure 2.20 Cyclocheilichthys enoplos (from Vittayanon 2000)
Description
Cyclocheilichthys enoplos (Cyprinidae) can be distinguished from the other members of this genus in Cambodia by the lateral line with bifurcate or even multifurcate tubules. However, there is considerable variation in this character. The Khmer name used in Siem Reap is trey chhkok.
It is the largest species of the genus. It is reported to reach a total length of 74 cm, but lengths up to 45 cm are most common (Rainboth, 1996).
Figure 2.22 Cyclocheilichthys enoplos Body length and scales median radius (n = 197)
Fish dissected in July and August 1997 showed high amounts of body fat, often combined with important parasite infections of the intestinal tract and internal organs.
Feeding ecology
The young of Cyclocheilichthys enoplos are reported to feed on zooplankton while the diet of adults includes insect larvae, crustaceans and fish. There are reports of the diet also containing plant roots, bivalves and green algae (FishBase, 2000) and being more that of omnivorous fish. However, Nguyen and Nguyen (1991) report that zoobenthos accounts for up to 95 percent of the food intake.
Reproductive ecology
Rainboth (1996) reports this fish to spawn in the rainy season, without specifying when precisely. There is one report from Thailand (FishBase, 2000) about the presence of mature females of this species from May to August. The main spawning season of the group of anadromously migrating fish for breeding in the Mekong like Cyclocheilichthys enoplos is from May to July (Nguyen and Nguyen, 1991).
Fisheries
The fish is caught with different kinds of nets and traps, but only occasionally with gillnets. It is usually marketed fresh and is among the more expensive fish.
Potential for aquaculture
Cyclocheilichthys enoplos is not found in impoundments. Experimental use for aquaculture is reported from Thailand (FishBase, 2000).
Importance
Cyclocheilichthys enoplos is one of the most important species in the fisheries of Cambodia. With 9 percent of the total volume, it is reported to be the third most important species in the catches by middle and large-scale fishing operations (Deap et al., 1998). It is found in benthopelagic habitats in rivers and the lake, and moves into the flooded plains and inundated riparian forests. It is reported to return from there to the permanent waters from October to December. This fish is considered excellent eating and is highly sought after.
Conclusion
Considering its diet and the lack of adaptations for surviving in waters with low dissolved oxygen levels, Cyclocheilichthys enoplos will probably not move into the inundated areas until water quality has improved after the beginning of the flooding. Low dissolved oxygen levels limit the area where the fish can comfortably survive. This factor, combined with the lack of well-developed intermediary-level communities (zoobenthos, aquatic insects, zooplankton), limits the distribution of Cyclocheilichthys enoplos in the flooded areas during the earlier stages of the flooding. It is one of the first species to return to the permanent water zones as soon as the floodwaters begin to recede.
Figure 2.24 Henicorhynchus siamensis (from Vittayanon, 2000)
Description
The taxonomic status of Henicorhynchus siamensis and of the genus Henicorhynchus has been subject to considerable discussion. FishBase (2000) considers the name Henicorhynchus siamensis (deBeaufort, 1927) a junior synonym to Cirrhinus siamensis (Sauvage, 1881). For consistency, the former is used in this publication. This species is another member of the in Cambodian fresh waters dominantly present family of the Cyprinidae. The Khmer name is trey riel.
There is a gap in the representation of fish of about 80 mm in the fish sampled. The cause for this is not clear. Maximum size reported for this species is 20 cm standard length (Rainboth, 1996).
Feeding ecology
This fish is well known for its annual trophic migrations out to the floodplains in the wet season. It is almost completely absent from the experimental gillnet catches from January until March. This coincides with the period when most of the migration towards the Mekong takes place (Rainboth, 1996). Henicorhynchus siamensis is reported to be herbivorous, feeding on algae, periphyton and phytoplankton.
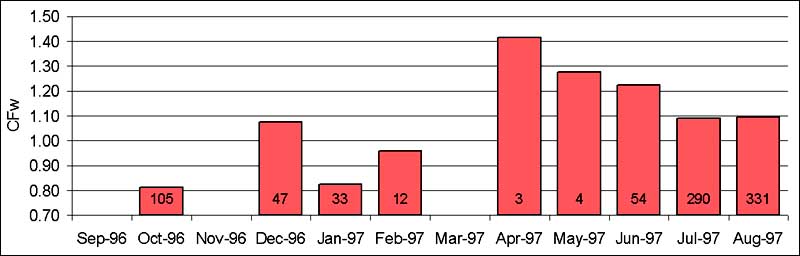
The evolution of the condition factor from April to August appears to confirm the feeding migration into the flooded areas. The first fish entering the inundated habitats (April-May) are rather lean but by the time the floodwaters begin to recede (December), they are about twice as heavy as what can be expected for a fish caught at random throughout the year.
Reproductive ecology
The rise of the condition factor is partly due to the development of gonads in the females (Fig. 2.27). Henicorhynchus siamensis is reported to reproduce in flowing water, also in currents in the core of the Tonle Sap lake and in the floodplain (H. Hy, personal communication, 2000).
Large amounts of body fat were observed in many specimens during the dissections in June and July 1997.
Figure 2.28 Henicorhynchus siamensis. Body length and scales median radius (n = 84)
Fisheries
Most of the catches of Henicorhynchus siamensis are done with seines, cast nets, traps and set nets. The dai fisheries take a large part of the total catch during the annual migration out of the Tonle Sap lake towards the Mekong. The fish is used along the Tonle Sap to make prahoc.
Potential for aquaculture
Henicorhynchus siamensis is not known to prosper in impoundments and seems to have limited potential for aquaculture.
Importance
Henicorhynchus siamensis is the backbone of commercial fishing operations in the Tonle Sap lake and channel. It makes up every fifth kilogram of fish produced by these fisheries, and represents almost 10 percent of the total value generated (Deap et al., 1998). It is no coincidence that its Khmer name is that of the national currency. Despite heavy fishing pressure, the species is not believed to be overfished at present (van Zalinge and Thuok, 1999).
Conclusion
The main function of the inundated area for this species is feeding. This is supported by the increasing condition factor as the flooding starts and goes on. More body fat is also observed during this period. The spawning function is not clear. Its diet makes it rather independent of the kind of habitat it moves to in the flooded area. Habitats such as scrubland and forest with lots of surface that offer opportunities for the development of epiphytic algae seem to offer advantages. Being a rather small fish, the bushy habitats offer considerable shelter from larger predators.
Figure 2.29 Osteochilus melanopleurus (from Vittayanon, 2000)
Description
The cyprinid osteochilids are commonly found in Cambodia, and in some places and at certain times they may be abundant. The Khmer name for Osteochilus melanopleurus is trey krum.
Osteochilus melanopleurus is the largest of the local species of the genus. Its maximum standard length is reported to be 40 cm (FishBase, 2000).
Feeding ecology
This species migrates into the seasonally flooded habitats but returns from there to the Mekong as soon as the water begins to recede in October. The migration reaches a maximum in January.
Figure 2.31 Osteochilus melanopleurus. Body length and scales median radius (n = 95)
The diet of Osteochilus melanopleurus consists mainly of periphyton, aquatic macrophytes and inundated land plants. It also feeds on phytoplankton, filamentous algae and bottom algae.
Reproductive ecology
All 65 specimens that were analysed in this study before and at the onset of the flooding were immature; in only one case the sex of the fish could be determined. Osteochilus melanopleurus probably reproduces in the Mekong.
Fisheries
Osteochilus melanopleurus is caught with hooks, seines, cast nets, set nets and traps. It is reported to be abundant in the catches of the dai fisheries in the Tonle Sap channel. Osteochilus melanopleurus was caught only sporadically with the experimental gillnets. Large individuals are marketed fresh or dried and salted. Small fish are used for the preparation of prahoc. The fish is among the more expensive; prices of 3 500 Riel/kg are no exception.
The results of the monthly condition factor CFw-analysis for Osteochilus melanopleurus should be interpreted with caution since they are based on rather small numbers of fish. At the end of the trophic migration to the Tonle Sap lake (December-January), the fish are heavy.
Potential for aquaculture
Rainboth (1996) reports that large specimens of this species are found in impoundments. Considering this, its high value and its diet, this species could offer opportunities for culture in reservoirs and ponds, provided seed is available.
Importance
Osteochilus melanopleurus represents a significant part of the total catches of commercial fisheries in Cambodia. There are no separate data by species available but the fish of the genus Osteochilus contribute up to 4 percent of the total commercial catches by weight (Deap et al., 1998).
Conclusion
The main reason for the migration of Osteochilus melanopleurus into the flooded area appears to be feeding. Given its preferred diet of periphyton, habitats offering opportunities for the proliferation of periphytic communities will offer the better feeding opportunities. Large surface area on the inundated vegetation is the determining factor in this, which makes that the habitats with a high standing crop of terrestrial vegetation -such as scrubland and forest - are most important. They offer furthermore plenty of feedstuff from inundated land plants and allow the development of epiphytic algae. Grassland and rice fields hold less surface area but they offer feeding opportunities in the stubbles and leaves that remain. Lotus fields show large surface areas but the shading by the leaves of Nelumbo nucifera limits primary production by phytoplankton and attached algae. The flora of the lake itself is not well known but offers probably only limited food resources for this species.
Figure 2.33 Paralaubuca typus (from Vittayanon, 2000)
Description
This species also belongs to the Cyprinidae family. Paralaubuca typus usually occurs in schools.
There is some confusion about the Khmer name of this species, and this is the only case where the nomenclature used in Rainboth (1996) is not entirely followed in this study. In-depth analysis of fresh samples has shown that the fish referred to in Siem Reap (by staff of the Department of Fisheries, by fishers, intermediaries, prahoc producers etc.) as trey slak russey in reality corresponds to the species Parachela siamensis (Günther, 1868). All the discriminative features as mentioned in the species guide by Rainboth (1996) are present: 12 gill rakers on the first arch, dark outline on first few lateral-line canals, dorsal fin origin slightly in front of anal fin, 43-46 lateral-line scales, 25-32 branched anal-fin rays, pectoral fin not extending beyond tip of pelvic fin. The characteristics for the genus Parachela are all present. Resemblance with colour plate No. III, 18 in Rainboth’s guide supports this as well. So either the Khmer name as referred to in the guide is incorrect or there is a local alternative use of this Khmer species name. Simultaneously, the name used in Siem Reap for fish identified as Paralaubuca typus (Bleeker, 1865) is trey chanteas phluk or rather trey chantu phluk. The Khmer name used in Rainboth’s guide for Paralaubuca typus is trey slak russey.
Table 2.16 Local species names
|
Species |
Rainboth (1996) |
Siem Reap |
|
Parachela siamensis (Günther, 1868) |
Trey chanteas phluk |
Trey slak russey |
|
Paralaubuca typus (Bleeker, 1865) |
Trey slak russey |
Trey chanteas phluk |
A normal but not invariable character in this species is the presence of a disconnected lateral line, with the anterior part overlapping the first 6 to 20 scales of the posterior part, which is one row below the anterior part (Smith, 1959).
The maximum length for Paralaubuca typus is reported to be 18 cm but most specimens are smaller. The fit of the power curve for the standard length-weight relation is poor, and this affects the usability of this equation to calculate the monthly condition factor (Fig. 2.35).
Feeding ecology
Paralaubuca typus moves into the flooded area late in the flood cycle when water levels are high, and only leaves the inundated area when the water levels have already considerably declined (Rainboth, 1996). Bardach (1959) reports Paralaubuca typus to be feeding mainly on plankton and crustaceans, but occasionally also on grains, seeds and flying insects. This rather diverse diet permits the species to thrive in several kinds of habitats in the inundated area, and to enter it before the communities of zooplankton and benthos have become fully established.
Reproductive ecology
Smith (1959) reports Paralaubuca typus to be spawning at the beginning of the rains (May-June), the fish reaching full maturity when 13 to 15 cm long. In July and August 1997, all 124 fish analysed for sexual maturity were considered immature, though this may have been a post-spawning phase.
Fisheries
The species is caught by seines, cast nets, set nets, weirs, traps and gillnets.
Figure 2.36 Paralaubuca typus. Body length and scales median radius (n = 58)
Potential for aquaculture
The fish does well in land-locked ponds of limited size.
Importance
This species makes up a significant element of the fish catches in Cambodia but given the confusion about the local names, it is difficult to assess the reported catch figures. Deap et al. (1998) report it to make up 3 percent by weight of the commercial catch, and it is one of the major species in gillnet catches.
Conclusion
Paralaubuca typus performs a shifted migration into the flooded area. It moves into the flooded vegetation rather late, at high water levels, when the water quality in the flooded area has already considerably improved, and the zooplankton communities have had some time to develop. Its late migration out of the flood area into the permanent waters seems to indicate that it endures increasingly adverse water quality conditions to benefit maximally from the food available in the flooded habitats. Habitats that offer favourable conditions for the development of rich zooplankton communities are expected to be of interest for this species. The preferred habitats would be scrubland, forest and lotus fields.
Figure 2.37 Trichogaster microlepis (from Rainboth 1996)
This species, also known as the moonlight gourami, belongs to the family of the Belontiidae or gouramis. It can be distinguished from the other species in the genus by its plain coloration, small scales and reduced number of dorsal-fin spines. The Khmer name for this species is trey kawmphleanh phluk.
A total length of 15 cm is attained.
Feeding ecology
The diet of the species is reported to consist of zooplankton, crustaceans and aquatic insects (Rainboth 1996).
Reproductive ecology
The species of the Trichogaster genus blow masses of glutinous bubbles to form floating nests in which they deposit their eggs. The use of a nest to deposit the eggs keeps them and the newly hatched fry concentrated in a protected locality that is easily defended by one or both parents (Welcomme, 1985). The use of a floating nest also keeps the eggs and young fish in contact with the upper layers of the water, which are better oxygenated. This behaviour allows these fish to reproduce in the highly deoxygenated habitats they usually inhabit. Trichogaster microlepis is a batch spawner species.
Fig. 2.40 shows the development of gonads in female fish towards the end of the study period. The sampling was fragmentary but shows that there is a peak in gonad weight at the end of July, which is at the beginning of the flood season. There may be others throughout the year.
Figure 2.41 Trichogaster microlepis. Body length and scales median radius (n = 51)
Fisheries
Trichogaster microlepis is caught by seines, cast nets, gillnets and traps. A specific kind of folded woven trap (la in Khmer) made out of thin sliced bamboo is used to catch Trichogaster species.
Potential for aquaculture
Trichogaster microlepis is reared in Thailand in captivity for the ornamental fish trade. Its potential for aquaculture in Cambodia is limited due to its low value, certainly in the proximity of the lake. However, it is commonly found in ponds as an accessory species.
Importance
Trichogaster microlepis makes up a significant part of the fisheries in the lake. It is sold fresh at markets but is mostly used for preparation of prahoc. This fish is one of the species that can be seen almost the year round in the shallow sluggish water at the edge of the lake or the flood area. Despite the very poor water quality, it is very common in the floating villages among the boats. As the other species in this genus, it is capable of surviving in waters with low dissolved oxygen levels and high turbidity thanks to the labyrinth organ that originates from the first gill arch in the suprabranchial cavity. As such, it is one of the common species caught with gillnets throughout the year.
Conclusion
Comparatively few piscivorous predators have developed adaptations to survive in deoxygenated waters so that fish that do colonize them are relatively free of predation (Welcomme 1985). Trichogaster microlepis is one of the fishes that can move into the flooded area during the early stages of the flooding. It has adaptations to survive in the deoxygenated waters and uses this period for reproduction. The kind of nests it makes allows the species to reproduce at stages of the flooding when most other fishes are unable to have eggs or fry survive the low oxygen concentrations.
Habitats with a lot of vegetation cover that does not become completely submerged during the flooding offer specific protection for Trichogaster microlepis. They find a large part of their food in the upper layers of the water and make regular trips to the surface layer to benefit from the higher dissolved oxygen concentrations there or to inhale air.
Based on the share of 28.5 percent in the number of specimens in the experimental gillnet catches in the scrubland, this seems to be preferred habitat for Trichogaster microlepis. Stands of open water and submerged rice or grasslands offer less food and protection.
Figure 2.42 Trichogaster trichopterus (from Vittayanon 2000)
Description
This species is of the same genus as Trichogaster microlepis and they have a large number of characteristics in common. The oblique bars on the body and the two black spots, one at the middle of the side, and one at the caudal-fin base, characterize this species. Because of this conspicuous feature, the fish is also known as threespot gourami. The variation in the number of dorsal and anal-fin rays is considerable. Mostly, seven or eight dorsal-fin spines are present, together with eight or nine branched soft rays. Anal-fin rays counts vary between 9 and 12 spines and 33 to 38 branched rays (Rainboth, 1996). The Khmer name for this species is trey kawmphleanh samrai.
Trichogaster trichopterus is reported to reach a maximum length of 15 cm (FishBase, 2000, Rainboth, 1996). All the fish sampled in this study were smaller than 12.5 cm total length.
As in many other species, the condition factor (CFw) shows a peak in December, right after the height of the flood season (Fig. 2.45). Based on this, Trichogaster trichopterus seems to be relatively in better condition (heavier) during the low-water time of the year (February-July) than Trichogaster microlepis.
Figure 2.44 Trichogaster trichopterus. Body length and scales median radius (n = 74)
Feeding ecology
The diet composition of Trichogaster trichopterus is similar to that of T. microlepis: zooplankton, crustaceans and insect larvae. The superior mouth position and the bands of minute fixed teeth support this diet analysis. Mohsin and Ambak (1983) report Trichogaster trichopterus to be usually algae feeders.
Reproductive ecology
The same authors mention the length of the dorsal fin when flexed as a secondary sexual characteristic allowing for the sexing of the fish. This criterion has shown to be of little practical value. Like Trichogaster microlepis, this fish is partial to a weedy environment, which provides protection from predatory fish and birds and offers a suitable location for the bubble nests it builds, usually at the surface. The male usually gathers the eggs, places them in the nest and guards them until they reach the free-swimming stage (FishBase, 2000). Fig. 2.46 shows the development of gonads in female fish towards the end of the study period. The sampling was fragmentary but shows that there is a rise in gonad weight in July and August, which is at the beginning of the flood season. There may be other development periods throughout the year.
Fisheries
The species is an important component of the Cambodian fisheries, and like Trichogaster microlepis with which it is often found, is it mainly used for preparation of prahoc. It is caught in the same way as the other gourami.
Potential for aquaculture
The threespot gourami is a desired aquarium fish. In Thailand it is bread in captivity for this purpose.
Conclusion
Habitat preferences and advantages are similar as those for Trichogaster microlepis.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}