![]()
![]()
![]()
Calcium in health and disease
Le rôle du calcium dans la santé et la maladie
El calcio en relación con la salud y la enfermedad
B.E.C. Nordin
B.E. Christopher Nordin is Professor in the Division of Clinical Biochemistry, Institute of Medical and Veterinary Science, Department of Pathology at the University of Adelaide, South Australia, Australia.
Calcium is an essential but controversial nutrient: there is no consensus on the level of human calcium requirement or the significance of calcium deficiency. The explanation for this must be sought in the history of calcium research since Pommer (1885) first established the difference between osteoporosis and osteomalacia. Pommer showed that rickets and osteomalacia represented a failure to mineralize new bone as it was being formed, leading ultimately to a calcium deficit in the bony tissue itself, whereas osteoporosis was a deficit of whole bone tissue without change in its mineral content. Not long afterwards, Miwa and Stoeltzner (1898) demonstrated that calcium deficiency caused osteoporosis, not osteomalacia, in experimental adult animals. It was Mellanby (1918) who finally identified vitamin D as the factor that prevented and cured rickets. Soon after that, Telfer (1926) showed that vitamin D deficiency led to malabsorption of calcium. From this it was assumed that vitamin D deficiency and calcium deficiency must have the same metabolic consequences. Accepting the concept that both calcium and vitamin D deficiencies led to rickets or osteomalacia, Albright et al. (1938) invoked a new pathogenetic mechanism - failure of new bone formation because of sex hormone deficiency and/or negative nitrogen balance - to explain postmenopausal and other forms of osteoporosis.
This paradigm prevailed for a generation until a seminal paper from Sweden (Carlsson and Lindquist, 1955) showed that vitamin D not only regulated calcium absorption but also independently regulated plasma calcium homeostasis. This made it possible to distinguish vitamin D deficiency from calcium deficiency (Nordin, 1960) and allowed the re-emergence of the concept (Hess, 1929) that calcium deficiency led to osteoporosis and vitamin D deficiency to osteomalacia. Although this concept is increasingly accepted in theory, there is in practice no consensus about the contribution of calcium deficiency to clinical osteoporosis or about the reality of a calcium requirement in humans (Kanis and Passmore, 1989). Clearly the menopause is associated with a rise in bone breakdown rather than a fall in bone formation (Young and Nordin, 1967; Gallagher, Young and Nordin, 1972; Stepan et al., 1987; Nordin and Polley, 1987; Prince et al., 1995). The current issue is whether oestrogens inhibit bone resorption directly or whether they operate through the calcium economy; it is no longer maintained that they promote bone formation. In the meantime, Albright and colleagues' original ideas about osteoporosis survive in relation to corticosteroid-induced osteoporosis and perhaps as part of the explanation for age-related osteoporosis, particularly in men.
PHYSIOLOGY
Calcium is an essential nutrient accounting for about 2 percent of body weight, ranking fifth after oxygen, carbon, hydrogen and nitrogen. Nearly 99 percent of the body's calcium (1200 g or 30 mol) is stored in the skeleton; the remainder is in the teeth and soft tissues (each containing 7 g or 175 mmol) and the extracellular fluid (ECF) (1 g or 25 mmol) (Nordin, 1976). Calcium comprises 25 percent of the dry weight of the skeleton and circulates in the ECF at a concentration of 6 to 7 mg per 100 ml (1.5 to 1.75 mmol per litre). The total calcium concentration within cells is comparable to that in the extracellular water, but the free calcium concentration is about 104 times lower (Robertson, 1988).
The most obvious function of calcium is to provide rigidity to the skeleton by virtue of the insoluble salts it forms with phosphoric acid. Primitive exoskeletons (such as shells) are composed of calcium carbonate formed in seawater, which is high in calcium but low in phosphorus. Fish endoskeletons may be composed of cartilage or bone. All mammalian skeletons are composed of true bone in which rigidity is provided by a particularly insoluble calcium salt analogous to the mineral hydroxyapatite [Ca10 (OH)2 (PO4)6] but containing also a small component of calcium carbonate (Termine and Posner, 1967).
The calcium salts do more than provide rigidity to the skeleton; they also constitute a very large reservoir of calcium for the maintenance of the (ionized) calcium concentration in the extracellular water at or very close to 4.8 mg per 100 ml (1.2 mmol per litre). The protection of this critical concentration by parathyroid hormone and vitamin D reflects the vital role that calcium plays in the neuromuscular system, in regulation of the heart, in enzyme-mediated reactions and in many other metabolic processes. Since only about 1 g (25 mmol) of calcium normally circulates in the ECF, the skeleton can maintain the extracellular ionized calcium against external calcium deprivation for years before it suffers significant damage. It is important to recall that as calcium is removed from bone, the matrix is also resorbed; this is why the result is osteoporosis rather than osteomalacia.
BONE GROWTH AND DECAY
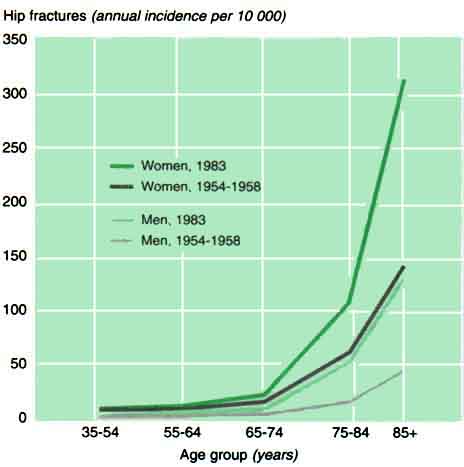
Calcium enters the body through the gastro-intestinal tract, is absorbed mainly in the upper small intestine and is excreted via the bowel, kidneys and skin. During skeletal growth and maturation, i.e. until the age of the early twenties in humans, calcium accumulates in the skeleton at an average rate of 150 mg a day (Figure 1). During maturity, the body - and therefore the skeleton - is more or less in calcium equilibrium. From the age of about 50 in men and from the menopause in women, bone balance becomes negative and bone is lost from all skeletal sites (Figure 2). This bone loss is associated with a marked rise in fracture rates, starting with non-hip fractures in women after the menopause but leading to rising hip fracture rates in both sexes (Figures 3 and 4). The cause of the bone loss associated with menopause and ageing is uncertain. This paper looks at this question, particularly in relation to the body's calcium economy.
THE CALCIUM ECONOMY
Regulation of the calcium economy is somewhat more complex than that of other minerals such as sodium, potassium, phosphorus and magnesium (Marshall, Nordin and Speed, 1976). These other minerals are primarily regulated by a relatively simple homeostatic system in which variations in input modify the concentration in the ECF, which in turn modifies the urinary loss; as input rises or falls, the ECF concentration follows suit, and so does urinary excretion, until a new equilibrium is established at which input and output are equal. This account is of course an oversimplification, but it represents the essential feature of these systems.
The calcium system contains a similar component but differs from the others in that the concentration of (ionized) calcium in the ECF is more vigorously defended, presumably because it is critical to so many physiological functions. Although it is true that raising calcium input raises ECF and urine calcium and therefore tends to equilibrium, the converse process (lowering of ECF and urine calcium when input falls) is limited by the intervention of parathyroid hormone secretion when the ionized calcium approaches the lower limit of the normal range. The result is the maintenance of the ECF ionized calcium and thus of calcium excretion via the kidneys and the bowel. The immediate effect of this parathyroid response is to increase renal tubular resorption of calcium (and so to conserve calcium) and to stimulate the synthesis of calcitriol [1,25(OH)2D3 in the kidneys to promote calcium absorption. If these mechanisms are insufficient to maintain the concentration of ionized calcium, parathyroid hormone secretion rises and increases bone resorption. This is the response of the organism to calcium deprivation, and it leads sooner or later to osteoporosis in experimental animals (Nordin, 1960). Osteoporosis can also be produced experimentally by oophorectomy, but the magnitude of the effect depends on the calcium intake (Hodgkinson et al., 1978; Shen et al., 1995) and can be amplified by sodium feeding, which increases the obligatory calcium loss (Goulding and Campbell, 1983). Since oestrogen insufficiency is regarded as the most important single risk factor for osteoporosis - at least in women - it is important to establish whether the negative calcium balance that follows the menopause is the cause or the result of the bone loss. This question requires consideration of the calcium requirement and therefore of the relationship among calcium intake, absorption and excretion.
1 Accumulation of total bone mineral and whole body calcium in boys and girls as a function of age
Source: J. Zanchetta, unpublished Norland bone densitometry data.
Note: Densitometry data are normally expressed in total bone mineral terms (left axis), whereas nutritionists normally consider the body content of elemental calcium (right axis). Bone mineral contains approximately 40 percent calcium.
2 Percentage loss of bone at four different sites as a function of age in men and women (Females)
2 Percentage loss of bone at four different sites as a function of age in men and women (Males)
Source: Calculated from Wahner and Fogelman, 1994.
3 Wrist fracture rates as a function of age in men and women in two European cities
Source: Miller and Grimley Evans, 1985.
Source: Calculated from Wahner and Fogelman, 1994.
Intake
Calcium consumption varies greatly worldwide, ranging from 800 mg per day or more in industrialized countries to 200 to 300 mg in some developing countries (FAO, 1991) (Table 1). The differences are mainly due to variation in the intake of dairy products, which are the richest sources of calcium. In countries where milk is scarce, calcium may be obtained from certain cereals, notably millets. Despite this large variation in calcium consumption, there is no evidence that osteoporosis is more common in developing than in industrialized countries - rather the reverse. In fact, the prevalence of hip fracture, as measured by mortality from fractures in the elderly (Eddy, 1972), is positively rather than negatively related to calcium intake (Figure 5). This paradox, and evidence of adaptation to low calcium intakes in Norwegian male prisoners (Malm, 1958), led a 1960 FAO/WHO Expert Committee to recommend a calcium allowance of only 500 mg (FAO, 1962). This recommendation is now being seriously questioned if not abandoned as the influence of other dietary constituents becomes increasingly apparent (Nordin et al., 1996).
Absorption
Calcium is absorbed from the small intestine by a saturable active transport system with a diffusion component (Wilkinson, 1976; Marshall, 1976; Bronner, 1987; Heaney, Saville and Recker, 1975) (Figure 6). At low calcium intakes, most calcium travels by the active pathway, but as intake rises, the amount transported by diffusion (6 percent of intake) gradually becomes dominant. In the calcium intake range of 500 to 1 000 mg (12.5 to 25 mmol), active transport is the main component, but at higher intakes (e.g. when calcium supplements are taken), the additional calcium is mainly absorbed by diffusion. It follows that true fractional calcium absorption is inversely related to calcium intake and falls from about 70 percent for very low intakes to 10 percent or less for high intakes (Nordin and Marshall, 1988). However, there is always loss of calcium in the faeces from the digestive juices (endogenous faecal calcium), and it follows that the net calcium absorbed (the difference between calcium intake and faecal calcium) is negative at zero intake, becomes zero when intake matches endogenous loss and then increases with intake, rapidly at first and then more slowly. This relationship [based on 212 calcium balances in 84 normal Western subjects (Marshall, Nordin and Speed, 1976)] is shown in Figure 6; the equation is:
y = [491x/(287 + x)] + 0.06x - 206 mg,
where y represents net absorbed calcium and x represents intake (Nordin and Marshall, 1988).
TABLE 1. Food consumption, latitude, hip fractures and standard of living in 19 countries
|
Country |
Calcium consumptiona |
Animal proteina |
Latitude of capital city |
Mortality from falls in women over age 75b |
Gross domestic product |
|
Japan |
354 |
52.2 |
19.2 |
2.9 |
21 140 |
|
Jamaica |
319 |
27.5 |
17.6 |
3.4 |
3 400 |
|
Guatemala |
262 |
7.6 |
14.5 |
4.3 |
3 440 |
|
Chile |
466 |
27.1 |
33.0 |
7.2 |
8 890 |
|
Brazil |
406 |
24.5 |
16.5 |
7.4 |
5 400 |
|
Australia |
921 |
65.7 |
35.1 |
7.4 |
18 120 |
|
Argentina |
603 |
63.6 |
34.4 |
7.9 |
8 720 |
|
United Kingdom |
855 |
53.0 |
51.3 |
8.5 |
17 970 |
|
Ecuador |
529 |
22.3 |
1.0 |
8.7 |
4 190 |
|
Colombia |
519 |
25.6 |
4.4 |
9.3 |
5 330 |
|
New Zealand |
771 |
70.2 |
41.2 |
11.6 |
15 870 |
|
Canada |
898 |
62.7 |
45.3 |
12.4 |
19 960 |
|
Israel |
704 |
55.1 |
32.1 |
13.4 |
15 300 |
|
Netherlands |
997 |
64.7 |
52.2 |
16.6 |
18 750 |
|
Finland |
1 267 |
61.6 |
60.1 |
18.0 |
16 150 |
|
Italy |
637 |
57.3 |
41.5 |
21.6 |
18 460 |
|
France |
969 |
76.1 |
48.5 |
24.6 |
19 670 |
|
Norway |
1 002 |
64.7 |
59.6 |
27.0 |
20 210 |
|
Denmark |
974 |
66.2 |
55.5 |
29.0 |
19 880 |
aFAO, 1991.
bWHO, 1990-1995.
cWorld Bank, 1996.
The principal regulator of calcium absorption is vitamin D [generally derived from the effect of sunlight on the skin (Holick, 1995)] in the form of its two active metabolites, 25-hydroxy vitamin D (calcidiol) and 1,25-dihydroxyvitamin D (calcitriol) (Holick, 1995), acting almost certainly on the active transport component. It would therefore be expected that the absorption line in Figure 6 would move up or down with vitamin D status (see below).
Excretion
Apart from the loss of calcium through the bowel, the principal routes of calcium excretion are via the kidneys and the skin. In normal individuals in zero calcium balance, calcium excreted via the skin and urine must equal net calcium absorbed. The slope of urinary calcium (y) to dietary calcium (x) in normal Western subjects is: y = 0.051x + 128 mg per day (Nordin and Marshall, 1988). However, skin loss of some 40 mg (1 mmol) per day (Charles et al., 1983) also needs to be taken into account. If this skin loss is assumed to be independent of intake, it can be added to the urine calcium component, and the equation of skin plus urine calcium to intake becomes y = 0.051x + 168 mg per day, as represented by the central excretion line in Figure 6. It must be emphasized that this relationship is based on Western data. Urine calcium is closely related to phosphate and sodium excretion, and variation from an average Western diet will move the urine calcium line up or down as indicated in Figure 6 and discussed below.
Calcium requirement
The mean calcium intake at which urine plus skin calcium equals net absorbed calcium can be taken as the calcium requirement of adults. As calculated by Nordin and Marshall (1988) from 212 Western calcium balances, this requirement amounts to about 840 mg (21 mmol) daily (Figure 6). It is clear that calcium consumption in most developing countries falls well short of this value (Table 1), which would lead to the conclusion either that the values derived from balance studies are misleading and do not take sufficient account of adaptation (which was the view of the FAO/WHO Expert Committee) or that there are special circumstances that reduce the calcium requirement of poorer nations. The reduction could be related to reduced calcium excretion and/or increased calcium absorption.
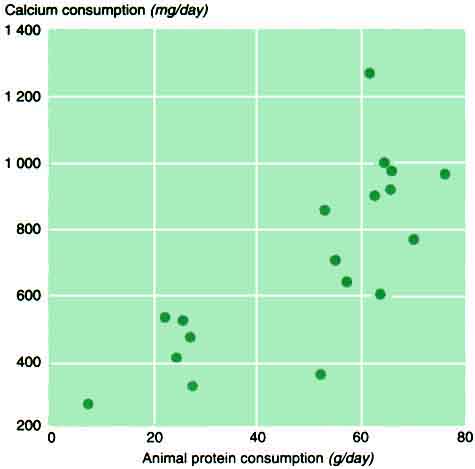
Reduced calcium excretion. At least two factors modify urine calcium at any given calcium intake: dietary protein and sodium. A positive relationship between urine calcium and protein (particularly animal protein) intake has been well documented (Linkswiler et al., 1981; Margen et al., 1974) and is probably due to the complexing of calcium by phosphate in the renal tubules. This explains the significant correlation between urine calcium and phosphate in normal subjects on free diets (Nordin and Polley, 1987), but the quantitative relation between protein intake and urine calcium has not been precisely defined. However, in one small study (Nordin et al., 1996) in which calcium intake was held constant, raising the animal protein intake from 40 to 80 g increased the urine calcium by 40 mg (1 mmol) in six postmenopausal women. If this relationship operates at all calcium intakes (which is not certain), the addition of 40 g of animal protein to a basal intake of 60 g would double the calcium requirement. Conversely, reducing the animal protein intake from 60 to 20 g would be expected to reduce the calcium requirement from 840 to 480 mg per day (21 to 12 mmol) (Figure 6). Since animal protein and calcium intakes are strongly related (Table 1 and Figure 7), populations that have low calcium intakes may be at least partially protected from negative calcium balance by their low protein consumption.
6 Calcium absorbed and excreted as a function of calcium intake in normal subjects
Note: The intercept of the excretion and absorption lines represents the calcium requirement, as shown by the interrupted lines.
7 Relation between calcium and protein consumption in the countries in Table 1
Urine calcium is also greatly influenced by sodium intake, because of competition between calcium and sodium ions for renal tubular resorption; 2.3 g of sodium (100 mmol) results in excretion of 1 mmol of calcium (40 mg) in the urine (Nordin and Polley, 1987; Nordin et al., 1996; Goulding, 1981; Nordin et al., 1993; Nordin and Need, 1994). Varying the sodium intake by this amount therefore has the same effect on urine calcium as 40 g of animal protein (Figure 6). It is likely that these two effects are additive (since the mechanisms are different) and that populations on low sodium and low protein intakes would have even lower calcium requirements, but sodium data from poorer countries are scanty (Intersalt Cooperative Research Group, 1988). In hot climates, there may also be loss of sodium in sweat, which would tend to divert sodium from the urine and reduce renal calcium excretion and therefore the calcium requirement. However, this effect might be offset by greater losses of calcium in sweat (Klesges et al., 1996), which would have the opposite effect. This is relatively unexplored territory.
Increased calcium absorption. Higher calcium absorption must also lower the calcium requirement. Since calcium absorption is a function of the serum calcitriol level (Morris et al., 1991), and since the serum concentration of its precursor calcidiol is a function of latitude (because of the angle of incidence of the sun's rays) (Table 2 and Figure 8), it is very likely, although unproven, that calcium absorption also falls with latitude. However, serum calcidiol is also a function of racial factors (skin pigmentation) (Matsuoka et al., 1995), social habits and climate (exposure to sunlight) (van der Wielen et al., 1995), diet (McKenna, 1992) and age (Baker, Peacock and Nordin, 1980; MacLaughlin and Holick, 1985), all of which contribute to the scattered pattern in Figure 8. Hip fracture rates (inferred from mortality from falls in the elderly) are also a positive function of latitude (Figure 9) and are thus more closely related to vitamin D status than to wealth or animal protein intake (Table 1). Moreover, low vitamin D status is a well-documented feature of hip fracture cases in the West (Aaron et al., 1974; Baker et al., 1979; Lips et al., 1982; Morris et al., 1984), and the risk of hip fracture was significantly reduced by vitamin D with calcium in a controlled study in France (latitude 48°) (Chapuy et al., 1992). Vitamin D supplementation was also shown to inhibit bone loss in a controlled study of elderly women in Leeds, United Kingdom (latitude 53°) (Nordin et al., 1985).
TABLE 2. Latitude and vitamin D status in 27 cities
|
City |
Latitude |
Serum calcidiol |
Reference |
|
Recife, Brazil |
8 |
103 |
Linhares et al., 1984 |
|
Curaçao, Netherlands Antilles |
12 |
116 |
Dubbelman et al., 1993 |
|
Hong Kong SAR, China |
22 |
60 |
Pun et al., 1990 |
|
Brisbane, Australia |
28 |
90 |
Personal communication |
|
Houston, USA |
30 |
125 |
Breslau et al., 1982 |
|
Perth, Australia |
32 |
86 |
Personal communication |
|
Sydney, Australia |
34 |
110 |
Personal communication |
|
Adelaide, Australia |
35 |
90 |
Personal communication |
|
Buenos Aires, Argentina |
36 |
53 |
Personal communication |
|
Melbourne, Australia |
37 |
95 |
Personal communication |
|
Saint Louis, USA |
38 |
58 |
Villareal et al., 1991 |
|
Baltimore, USA |
39 |
92 |
Sherman, Hollis & Tobin, 1990 |
|
Indianapolis, USA |
40 |
60 |
Personal communication |
|
Philadelphia, USA |
40 |
63 |
Matsuoka et al., 1995 |
|
Madrid, Spain |
40 |
59 |
Moreiras et al., 1992 |
|
Omaha, USA |
41 |
75 |
Personal communication |
|
New York, USA |
42 |
60 |
Meier, 1991 |
|
Hobart, Australia |
43 |
66 |
Personal communication |
|
Paris, France |
48 |
60 |
Chapuy, Durr & Chapuy, 1983 |
|
Brussels, Belgium |
50 |
61 |
Bouillon et al., 1987 |
|
Cardiff, UK |
51 |
56 |
Compston et al., 1981 |
|
London, UK |
51 |
18 |
Corless et al., 1985 |
|
Groningen, Netherlands |
52 |
50 |
Dubbelman et al., 1993 |
|
Leeds, UK |
53 |
50 |
Personal communication |
|
Copenhagen, Denmark |
55 |
73 |
Hartwell, Riis & Christiansen, 1990 |
|
Oslo, Norway |
59 |
75 |
Falch, Oftebro & Haug, 1987 |
|
Helsinki, Finland |
60 |
47 |
Von Knorring et al., 1982 |
8 Relation between serum calcidiol and latitude in the cities in Table 2
MENOPAUSE
Although the above account may offer a possible explanation for the apparent differences in calcium requirement and hip fracture rates among nations, it leaves the important role of the menopause unexplained. The menopause is associated with a rise in obligatory calcium excretion (Young and Nordin, 1967; Gallagher, Young and Nordin, 1972; Stepan et al., 1987; Nordin and Polley, 1987; Prince et al., 1995) and a probable decrease in calcium absorption (Heaney et al., 1989; Nordin, 1997) which are commonly attributed to a cytokine-induced increase in bone resorption secondary to oestrogen deficiency (Manolagas et al., 1993; Pacifici, 1996). This aspect tends to separate postmenopausal bone loss from the rest and to imply a need for hormones or other expensive treatment regimens. However, it can be argued that menopausal bone loss results from an increase in calcium requirement and can be managed accordingly. The increase in calcium loss at the menopause is some 20 to 40 mg (0.5 to 1.0 mmol) (Young and Nordin, 1967; Gallagher, Young and Nordin, 1972; Stepan et al., 1987; Nordin and Polley, 1987; Prince et al., 1995), which is comparable to the salt and protein effects described above and shown in Figure 6.
9 Mortality from falls in women over age 75 as a function of latitude in the countries in Table 1
This argument, fully reviewed elsewhere (Nordin, 1997), relies on the experimental evidence that ovariectomy-induced osteoporosis in experimental animals is modifed by calcium and sodium intake (Hodgkinson et al., 1978; Shen et al., 1995; Goulding and Campbell, 1983) and on physiological evidence that the increase in calcium excretion in women at the menopause is due to a renal calcium "leak" (Nordin et al., 1991; Nordin et al., 1994) rather than to an overflow of calcium lost from the skeleton by a primary process. This paradigm is supported by 20 prospective bone density trials in which mean bone loss was close to zero in 855 calcium-treated postmenopausal women and highly significant in 625 controls (P < 0.001) (Nordin, 1997). Although treatment with calcium is not quite as effective as treatment with oestrogen [which produces a small gain of bone (Prince et al., 1991; Christiansen and Riis, 1990), which was not seen in the calcium trials], this may be because oestrogens simultaneously lower urinary calcium (Gallagher and Nordin, 1973; Reginster et al., 1993; Prince, 1994; Selby et al., 1985) and improve calcium absorption (Civitelli et al., 1988; Horowitz et al., 1993), thus having a greater effect than simple calcium supplementation. This issue is not yet resolved, but the old paradigm is changing, and the concept that menopausal bone loss is (at least in part) secondary to an increase in calcium requirement is gaining support. Recent consensus conferences (United States National Institutes of Health, 1994; American Journal of Medicine, 1993) have recommended a calcium allowance of 1500 mg for postmenopausal women, without however explicitly stating that postmenopausal osteoporosis is, or could be, caused by calcium deficiency.
CONCLUSIONS
It is remarkable that there should be so much controversy over the roles of calcium and vitamin D in human nutrition in general and in osteoporosis in particular, given that both are acknowledged to be essential nutrients. Classical work on rickets and osteomalacia casts a long shadow. No sooner have osteomalacia and osteoporosis been satisfactorily distinguished than evidence of their overlapping aetiologies becomes apparent. The theses put forward in this paper are that primary negative calcium balance is the major, but not the only, cause of osteoporosis; that such a negative balance may be the result of inadequate calcium intake, low calcium absorption or high calcium excretion; that low calcium absorption may be the result of moderate vitamin D insufficiency; and that high calcium excretion may be due to dietary factors (such as protein and sodium intakes) or to hormonal effects (such as oestrogen deficiency). Seen in this light, the worldwide pattern of osteoporosis becomes comprehensible, but carefully targeted fieldwork - exemplified by studies on the relationship between calcium intake and bone density in the Gambia (Prentice, 1994) - will be required to fill in the picture.
REFERENCES
Aaron, J.E., Gallagher, J.C., Anderson, J., Stasiak. L., Longton, E.B. & Nordin, B.E.C. 1974. Frequency of osteomalacia and osteoporosis in fractures of the proximal femur. Lancet, 2: 229-233.
Albright, F., Bloomberg, E., Drake, T. & Sulkowitch, H.W. 1938. A comparison of the effects of A.T.10 (dihydrotachysterol) and vitamin D on calcium and phosphorus metabolism in hypoparathyroidism. J. Clin. Invest., 17: 317-329.
American Journal of Medicine. 1993. Consensus development conference: diagnosis, prophylaxis, and treatment of osteoporosis. 94: 646-650.
Baker, M.R., McDonnell, H., Peacock, M. & Nordin, B.E.C. 1979. Plasma 25-hydroxy vitamin D concentrations in patients with fractures of the femoral neck. Br. Med. J., 1: 589.
Baker, M.R., Peacock, M. & Nordin, B.E.C. 1980. The decline in vitamin D status with age. Age Ageing, 9: 249-252.
Bouillon, R.A., Auwerx, J.H., Lissens, W.D. & Pelemans, W.K. 1987. Vitamin D status in the elderly: seasonal substrate deficiency causes 1,25-dihydroxycholecalciferol deficiency. Am. J. Clin. Nutr., 45: 755-763.
Boyce, W.J. & Vessey, M.P. 1985 Rising incidence of fracture of the proximal femur. Lancet, 2: 150-151.
Breslau, N.A., McGuire, J.L., Zerwekh, J.E. & Pak, C.Y.C. 1982. The role of dietary sodium on renal excretion and intestinal absorption of calcium and on vitamin D metabolism. J. Clin. Endocrinol. Metab., 55: 369-373.
Bronner, F. 1987. Intestinal calcium absorption: mechanisms and applications. J. Nutr., 11: 1347-1352.
Carlsson, A. & Lindquist, B. 1955. A comparison of the intestinal and skeletal effect of vitamin D in relation to dosage. Acta Physiol. Scand., 35: 53-55.
Chapuy, M.C., Arlot, M.E., Duboeuf, F., Brun, J., Crouzet, B., Arnaud, S., Delmas, P.D. & Meunier, P.J. 1992. Vitamin D3 and calcium to prevent hip fractures in elderly women. N. Engl. J. Med., 327: 1637-1642.
Chapuy, M.-C., Durr, E. & Chapuy, P. 1983. Age-related changes in parathyroid hormone and 25 hydroxycholecalciferol levels. J. Gerontol., 38: 19-22.
Charles, P., Taagehoj, F., Jensen, L., Mosekilde, L. & Hansen, H.H. 1983. Calcium metabolism evaluated by Ca47 kinetics: estimation of dermal calcium loss. Clin. Sci., 65: 415-422.
Christiansen, C. & Riis, B.J. 1990. 17b -Estradiol and continuous norethisterone: a unique treatment for established osteoporosis in elderly women. J. Clin. Endocrinol. Metab., 71: 836-841.
Civitelli, R., Agnusdei, D., Nardi, P., Zacchei, F., Avioli, L.V. & Gennari, C. 1988. Effects of one-year treatment with estrogens on bone mass, intestinal calcium absorption, and 25-hydroxyvitamin D-1a -hydroxylase reserve in postmenopausal osteoporosis. Calcif. Tissue Int., 42: 77-86.
Compston, J.E., Vedi, S., Merrett, A.L., Clemens, T.L., O'Riordan, J.L.H. & Woodhead, J.S. 1981. Privational and malabsorption metabolic bone disease: plasma vitamin D metabolic concentrations and their relationship to quantitative bone histology. Metab. Bone Dis. Rel. Res., 3: 165-170.
Corless, D., Dawson, E., Fraser, F., Ellis, M., Evans, S.J., Perry, J.D., Reisner, C., Silver, C.P., Beer, M., Boucher, B.J. et al. 1985. Do vitamin D supplements improve the physical capabilities of elderly hospital patients? Age Ageing, 14: 76-84.
Dubbelman, R., Jonxis, J.H.P., Muskiet, F.A.J. & Saleh, A.E.C. 1993. Age-dependent vitamin D status and vertebral condition of white women living in Curaçao (the Netherlands Antilles) as compared with their counterparts in the Netherlands. Am. J. Clin. Nutr., 58: 106-109.
Eddy, T.P. 1972. Deaths from domestic falls and fractures. Br. J. Prev. Soc. Med., 26: 173-179.
Falch, J.A., Oftebro, H. & Haug, E. 1987. Early postmenopausal bone loss is not associated with a decrease in circulating levels of 25-hydroxyvitamin D, or vitamin D-binding protein. J. Clin. Endocrinol. Metab., 64: 836-841.
FAO. 1962. Calcium requirements. Report of an FAO/WHO Expert Group. FAO Nutrition Meetings Report Series No. 30. Rome.
FAO. 1991. Production Yearbook 1990, Vol. 44. Rome.
Gallagher, J.C. & Nordin, B.E.C. 1973. Oestrogens and calcium metabolism. In P.A. van Keep & C. Lauritzen, eds. Ageing and estrogens: frontiers of hormone research. Vol. 2, p. 98-117. Basel, Switzerland, Karger.
Gallagher, J.C., Young, M.M. & Nordin, B.E.C. 1972. Effects of artificial menopause on plasma and urine calcium and phosphate. Clin. Endocrinol., 1: 57-64.
Goulding, A. 1981. Fasting urinary sodium/creatinine in relation to calcium/creatinine and hydroxyproline/creatinine in a general population of women. N.Z. Med. J., 93: 294-297.
Goulding, A. & Campbell, D. 1983. Dietary NaCl loads promote calciuria and bone loss in adult oophorectomized rats consuming a low calcium diet. J. Nutr., 113: 1409-1414.
Hartwell, D., Riis, B.J. & Christiansen, C. 1990. Comparison of vitamin D metabolism in early healthy and late osteoporotic postmenopausal women. Calcif. Tissue Int., 47: 332-337.
Heaney, R.P., Recker, R.R., Stegman, M.R. & Moy, A.J. 1989. Calcium absorption in women: relationships to calcium intake, estrogen status, and age. J. Bone Miner. Res., 4: 469-475.
Heaney, R.P., Saville, P.D. & Recker, R.R. 1975. Calcium absorption as a function of calcium intake. J. Lab. Clin. Med., 85: 881-890.
Hess, A.F. 1929. Rickets, including osteomalacia and tetany. London, UK, Lea & Febiger.
Hodgkinson, A., Aaron, J.E., Horsman, A., McLachlan, M.S.F. & Nordin, B.E.C. 1978. Effect of oophorectomy and calcium deprivation on bone mass in the rat. Clin. Sci. Mol. Med., 54: 439-446.
Holick, M.F. 1995. Vitamin D: photobiology, metabolism, and clinical applications. In L.J. DeGroot, ed. Endocrinology, p. 990-1014. Philadelphia, Pennsylvania, USA, W.B. Saunders. 3rd ed.
Horowitz, M., Wishart, J.M., Need, A.G., Morris, H.A. & Nordin, B.E.C. 1993. Effects of norethisterone on bone related biochemical variables and forearm bone mineral in postmenopausal osteoporosis. Clin. Endocrinol., 39: 1-5.
Intersalt Cooperative Research Group. 1988. Intersalt: an international study of electrolyte excretion and blood pressure. Results for 24 hour urinary sodium and potassium excretion. Br. Med. J., 297: 319-328
Kanis, J.A. & Passmore, R. 1989. Calcium supplementation of the diet. Br. Med. J., 298: 137-140, 205-208.
Klesges, R.C., Ward, K.D., Shelton, M.L., Applegate, W.B., Cantler, E.D., Palmieri, G.M.A., Harmon, K. & Davis, J. 1996. Changes in bone mineral content in male athletes. J. Am. Med. Assoc., 276: 226-230.
Linhares, E.R., Jones, D.A., Round, J.M. & Edwards, R.H.T. 1984. Effect of nutrition on vitamin D status: studies on healthy and poorly nourished Brazilian children. Am. J. Clin. Nutr., 39: 625-630.
Linkswiler, H.M., Zemel, M.B., Hegsted, M. & Schuette, S. 1981. Protein-induced hypercalciuria. Fed. Proc., 40: 2429-2433.
Lips, P., Netelenbos, J.C., Jongen, M.J., van Ginkel, F.C., Althuis, A.L., van Schaik, C.L., van der Vijgh, W.J., Vermeiden, J.P. & van der Meer, C. 1982. Histomorphometric profile and vitamin D status in patients with femoral neck fracture. Metab. Bone. Dis. Rel. Res., 4:85-93.
MacLaughlin, J. & Holick, M.F. 1985. Aging decreases the capacity of human skin to produce vitamin D. J. Clin. Invest., 76: 1536-1538.
Malm, O.J. 1958. Calcium requirement and adaptation in adult men. Scand. J. Clin. Lab. Invest., 10(Suppl. 36): 1-289.
Manolagas, S.C., Girasole, G., Passeri, G., Bellido, T., Williams, D.C. & Jilka, R.L. 1993. Estrogen, cytokines, and the control of osteoclast formation and bone resorption in vitro and in vivo. Osteoporosis Int., Suppl. 1: S144-S166.
Margen, S., Chu, J.Y., Kaufman, N.A. & Calloway, D.H. 1974. Studies in calcium metabolism -1. The calciuretic effect of dietary protein. Am. J. Clin. Nutr., 27: 584-589.
Marshall, D.H. 1976. Calcium and phosphate kinetics. In B.E.C. Nordin, ed. Calcium, phosphate and magnesium metabolism, p. 257-297. Edinburgh, UK, Churchill Livingstone.
Marshall, D.H., Nordin, B.E.C. & Speed, R. 1976. Calcium, phosphorus and magnesium requirement. Proc. Nutr. Soc., 35: 163-173.
Matsuoka, L.Y., Wortsman, J., Chen, T.C. & Holick, M.F. 1995. Compensation for the interracial variance in the cutaneous synthesis of vitamin D. J. Lab. Clin. Med., 126(5): 452-457.
McKenna, M.J. 1992. Differences in vitamin D status between countries in young adults and the elderly. Am. J. Med., 93:69-77.
Meier, D.E., Luckey, M.M., Wallenstein, S., Clemens, T.L., Orwoll, E.S. & Waslien, C.I. 1991. Calcium, vitamin D, and parathyroid hormone status in young white and black women: association with racial differences in mass. J. Clin. Endocrinol. Metab., 72: 703-710.
Mellanby, E. 1918. The part played by an "accessory factor" in the production of experimental rickets. A further demonstration of the part played by accessory food factors in the aetiology of rickets. J. Physiol., 52: 11-53.
Miller, S.W.M. & Grimley Evans, J. 1985. Fractures of the distal forearm in Newcastle: an epidemiological survey. Age Ageing. 14: 155-158.
Miwa, S. & Stoeltzner, W. 1898. Über die bei jungen Hunden durch kälkarme Futterung enstehende Knochenerkrankung. Beitr. Pathol. Anat., 24: 578.
Moreiras, O., Carbaja, A., Perea, I. & Varela-Moreiras, V. 1992. The influence of dietary intake and sunlight exposure on the vitamin D status in an elderly Spanish group. Int. J. Vitam. Nutr. Res., 62(4): 303-307.
Morris, H.A., Morrison, G.W., Burr, M., Thomas, D.W. & Nordin, B.E.C. 1984. Vitamin D and femoral neck fractures in elderly South Australian women. Med. J. Aust., 140: 519- 521.
Morris, H.A., Need, A.G., Horowitz, M., O'Loughlin, P.D. & Nordin, B.E.C. 1991. Calcium absorption in normal and osteoporotic postmenopausal women. Calcif. Tissue Int., 49: 240-243.
Nordin, B.E.C. 1960. Osteomalacia, osteoporosis and calcium deficiency. Clin. Orthop., 17: 235-258.
Nordin, B.E.C. 1976. Nutritional considerations. In B.E.C. Nordin, ed. Calcium, phosphate and magnesium metabolism, p. 1-35. Edinburgh, UK, Churchill Livingstone.
Nordin, B.E.C. 1997. Calcium and osteoporosis. Nutrition, 13(7-8): 664-686.
Nordin, B.E.C., Baker, M.R., Horsman, A. & Peacock, M. 1985. A prospective trial of the effect of vitamin D supplementation on metacarpal bone loss in elderly women. Am. J. Clin. Nutr., 42: 470-474.
Nordin, B.E.C., Horowitz, M., Need, A.G. & Morris, H.A. 1994, Renal leak of calcium in postmenopausal osteoporosis. Clin. Endocrinol., 41: 41-45.
Nordin, B.E.C. & Marshall, D.H. 1988. Dietary requirements for calcium. In B.E.C. Nordin, ed. Calcium in human biology, p. 447-471. Berlin, Germany, Springer-Verlag.
Nordin, B.E.C., Morris, H.A., Need, A.G. & Horowitz, M. 1996. Dietary calcium and osteoporosis. In P. Pietinen, C. Nishida & N. Khaltaev. eds. Nutrition and quality of life. Proceeding of the 2nd WHO Symposium on Health Issues for the 21st Century, Kobe. Japan, 1993, p. 181-198. Geneva, Switzerland, World Health Organization.
Nordin, B.E.C. & Need, A.G. 1994. The effect of sodium on calcium requirement. Adv. Nutr. Res., 9: 209-230.
Nordin, B.E.C., Need, A.G., Horowitz, M. & Robertson, W.G. 1991. Evidence for a renal calcium leak in postmenopausal women. J. Clin. Endocrinol. Metab., 72: 401-407.
Nordin, B.E.C., Need, A.G., Morris, H.A. & Horowitz, M. 1993. The nature and significance of the relation between urine sodium and urine calcium in women. J. Nutr., 123: 1615-1622.
Nordin, B.E.C & Polley, K.J. 1987. Metabolic consequences of the menopause. A cross-sectional, longitudinal, and intervention study on 557 normal postmenopausal women. Calcif. Tissue Int., 41:S1 - S60.
Pacifici, R. 1996. Review: Estrogen, cytokines and pathogenesis of postmenopausal osteoporosis. J. Bone Miner. Res., 11: 1043-1051.
Pommer, G. 1885. Undersuchungen uber Osteomalacie und Rachitis. Leipzig, Germany, Vogel.
Prentice, A. 1994. Calcium intakes and bone densities of lactating women and breast-fed infants in the Gambia. Adv. Exp. Med. Biol., 352: 243-255.
Prince, R.L. 1994. Counterpoint: Estrogen effects on calcitropic hormones and calcium homeostasis. Endocr. Rev., 15: 301-309.
Prince, R.L., Dick, I., Devine, A., Price, R.I., Gutteridge, D.H., Kerr, D., Criddle, A., Garcia-Webb, P. & St John, A. 1995. The effects of menopause and age on calcitropic hormones: a cross-sectional study of 655 healthy women aged 35 to 90. J. Bone Miner. Res., 10: 835-842.
Prince, R.L., Smith, M., Dick, I.M., Price, R.I., Webb, P.G., Henderson, N.K. & Harris, M.M. 1991. Prevention of postmenopausal osteoporosis: a comparative study of exercise, calcium supplementation and hormone-replacement therapy, N. Engl. J. Med., 325: 1189-1195.
Pun, K.K., Wong, F.H., Wang, C., Lau, P., Ho, P.W., Pun, W.K., Chow, S.P., Cheng, C.L., Leong, J.C. & Young, R.T. 1990. Vitamin D status among patients with fractured neck of femur in Hong Kong. Bone, 11: 365-368.
Reginster, J.Y., Christiansen, C., Dequinze, B., Deroisy, R., Gaspard, U., Taquet, A.N. & Franchimont, P. 1993. Effect of transdermal 17b -estradiol and oral conjugated equine estrogens on biochemical parameters of bone resorption in natural menopause. Calcif. Tissue Int., 53: 13-16.
Robertson, W.G. 1988. Chemistry and biochemistry of calcium. In B.E.C. Nordin, ed. Calcium in human biology, p. 1-26. Berlin, Germany, Springer-Verlag.
Selby, P.L., Peacock, M., Barkworth, S.A., Brown, W.B. & Taylor, G.A. 1985. Early effects of ethinyloestradiol and norethisterone treatment in postmenopausal women on bone resorption and calcium regulation hormones. Clin. Sci., 69: 265-271.
Shen, V., Birchman, R., Xu, R., Lindsay, R. & Dempster, D.W. 1995. Short-term changes in histomorphometric and biochemical turnover markers and bone mineral density in estrogen and/or dietary calcium-deficient rats. Bone, 16: 149-156.
Sherman, S.S., Hollis, B.W. & Tobin, J.D. 1990. Vitamin D status and related parameters in a healthy population: the effects of age, sex and season. J. Clin. Endocrinol. Metab., 71: 405-413.
Stepan, J.J., Posphichal, J., Presl, J. & Pacovsky, V. 1987. Bone loss and biochemical indices of bone remodeling in surgically induced postmenopausal women. Bone, 8: 279- 284.
Telfer, S.V. 1926. Studies in calcium and phosphorus metabolism. Q. J. Med., 20: 1-6.
Termine, J.D. & Posner, A.S. 1967. Amorphous/crystalline interrelationships in bone mineral. Calcif. Tissue Res., 1: 8- 23.
United States National Institutes of Health. 1994. Optimal calcium intake. NIH Consensus Conference. J. Am. Med. Assoc., 272: 1942-1948.
van der Wielen, R.P.J., Löwik, M.R.H., van den Berg, H., de Groot, L.C.P.G.M., Haller, J. & Moreiras, O. 1995. Serum vitamin D concentrations among elderly people in Europe. Lancet, 346: 207-210.
Villareal, D.T., Civitelli, R., Chines, A. & Avioli, L.V. 1991. Subclinical vitamin D deficiency in postmenopausal women with low vertebral bone mass. J. Clin. Endocrinol. Metab., 72: 628-634.
Von Knorring, J., Slatis, P., Weber, T.H. & Helenius, T. 1982. Serum levels of 25-hydroxy vitamin D, 24,25-dihydroxy vitamin D and parathyroid hormone in patients with femoral neck fracture in southern Finland. Clin. Endocrinol. 17: 189- 194.
Wahner, H.W. & Fogelman, I. 1994. The evaluation of osteoporosis: dual energy X-ray absorptiometry in clinical practice. London, UK, Martin Dunitz Ltd.
Wilkinson, R. 1976. Absorption of calcium, phosphorus and magnesium. In B.E.C. Nordin, ed. Calcium, phosphate and magnesium metabolism. Edinburgh, UK, Churchill Livingstone, p. 36-112.
World Bank. 1996. World Development Report 1996. New York, NY, USA, Oxford University Press.
World Health Organization (WHO). 1990-1995. World Health Statistics Annual. Geneva, Switzerland.
Young, M.M. & Nordin, B.E.C. 1967. Effects of natural and artificial menopause on plasma and urinary calcium and phosphoms. Lancet, 2:118-120.
Summary
There is a remarkable amount of controversy about the roles of calcium and vitamin D in human nutrition in general and in osteoporosis in particular. This article reviews the functions of calcium in the body, highlights major findings about calcium during the past century and explains the current debates.
The calcium salts that provide rigidity to the skeleton also constitute a large reservoir for the maintenance of the calcium concentration in the body's extracellular water. The protection of this concentration by parathyroid hormone and vitamin D reflects the vital role of calcium in the neuromuscular system, in regulation of the heart, in enzyme-mediated reactions and in many other metabolic processes.
During the period of skeletal growth and maturation, i.e. until the age of the early twenties in humans, calcium accumulates in the skeleton. During maturity, the body is more or less in calcium equilibrium; from the age of about 50 years in men and from the menopause in women, bone balance becomes negative and bone is lost from all skeletal sites. This bone loss is associated with a marked rise in fracture rates. The cause of the bone loss associated with menopause and ageing is uncertain.
Calcium consumption varies greatly worldwide, ranging from 800 mg per day or more in industrialized countries to 200 to 300 mg per day in developing countries. Paradoxically, osteoporosis is more common in the industrialized countries.
The theses put forward in this paper are that primary negative calcium balance is the major, but not the only, cause of osteoporosis; that such a negative balance may be the result of inadequate calcium intake, low calcium absorption or high calcium excretion; that low calcium absorption may be the result of moderate vitamin D insufficiency; and that high calcium excretion may be correlated with dietary factors (such as protein and sodium intakes) or hormonal effects (such as oestrogen deficiency). Seen in this light, the worldwide pattern of osteoporosis becomes comprehensible, but carefully targeted field work will be required to fill in the picture.
Résumé
Le rôle du calcium et de la vitamine D dans l'alimentation humaine en général, et dans l'ostéoporose en particulier, est très controversé. Le présent article examine les fonctions du calcium dans le corps, souligne les principales conclusions des recherches entreprises au cours du siècle dernier et explique les débats actuels.
Les sels de calcium, qui assurent la rigidité au squelette, constituent également un vaste réservoir destiné à maintenir la concentration du calcium dans le liquide extracellulaire (LEC) du corps. L'hormone parathyroïde et la vitamine D protègent cette concentration qui reflète le rôle vital joué par le calcium dans le système neuromusculaire, dans le fonctionnement cardiaque, dans les réactions dépendant d'une activité enzymatique et dans beaucoup d'autres processus métaboliques.
Pendant la période de croissance et de maturation du squelette (dans les 20 premières années environ), le calcium s'accumule dans le squelette. A maturité, le corps est plus ou moins en équilibre calcique; à partir de 50 ans chez l'homme, et à partir de la ménopause chez la femme, le bilan osseux devient négatif et on enregistre une perte de masse osseuse dans toutes les parties du squelette, qui se traduit par une augmentation sensible de la fréquence des fractures. Les raisons de la perte osseuse liée à la ménopause et au vieillissement ne sont pas encore bien connues.
La consommation de calcium varie considérablement dans le monde: elle se situe entre 800 mg par jour ou plus dans les pays industrialisés, et à 200-300 mg par jour dans les pays en développement. Paradoxalement, l'ostéoporose est plus fréquente dans les pays industrialisés.
Les thèses exposées dans le présent article sont les suivantes: i) un bilan calcique primaire négatif est la raison principale, mais non la seule, de l'ostéoporose; ii) ce bilan peut être dû à un apport insuffisant de calcium, à une faible absorption ou à une excrétion importante de calcium; iii) une faible absorption de calcium peut provenir d'une carence modérée en vitamine D; et iv) une excrétion importante de calcium peut être due à des facteurs alimentaires (apports protéiques et sodiques, par exemple) ou hormonaux (tels qu'une insuffisance œstrogénique). Dans ce contexte, on comprend mieux le profil mondial de l'ostéoporose, mais on n'obtiendra une image complète de la situation que si on réalise des études de terrain soigneusement ciblées.
Resumen
La función del calcio y de la vitamina D en la nutrición humana en general y en la osteoporosis en particular ha suscitado notables polémicas. En el presente artículo se examinan las funciones del calcio en el organismo, se describen las principales conclusiones de los estudios sobre el calcio realizados durante el último siglo y se exponen los debates en curso.
Las sales de calcio que proporcionan rigidez al esqueleto constituyen además una reserva muy amplia para el mantenimiento de la concentración de calcio en el agua extracelular del organismo. La hormona paratiroidea y la vitamina D protegen esta concentración, lo que evidencia la función decisiva que el calcio desempeña en el sistema neuromuscular, en la regulación cardíaca, en las reacciones en las que intervienen enzimas y en muchos otros procesos metabólicos.
El calcio se acumula en el esqueleto durante el período de crecimiento y maduración de éste, es decir hasta que el individuo tiene poco más de 20 años. Durante la edad adulta, el calcio en el organismo se mantiene en equilibrio más o menos estable; a partir de los 50 años aproximadamente en los hombres y de la menopausia en las mujeres, el equilibrio óseo se altera y en todas las partes del esqueleto se producen pérdidas óseas. Este proceso está asociado con un aumento notable del índice de fracturas. No se conoce con certeza la causa de la pérdida ósea asociada con la menopausia y el envejecimiento.
El consumo de calcio varía enormemente en el mundo, oscilando entre 800 mg/día o más en los países industrializados y 200-300 mg/día en los países en desarrollo. Paradójicamente, la osteoporosis es más frecuente en los países industrializados.
Las tesis que se formulan en el presente artículo son que la alteración del equilibrio primario del calcio es la causa principal de la osteoporosis, pero no la única; que esta alteración del equilibrio puede ser el resultado de una ingesta insuficiente, de una absorción baja o de una excreción alta de calcio; que una absorción baja de calcio puede ser el resultado de una insuficiencia moderada de vitamina D; y que una excreción alta de calcio puede deberse a factores alimentarios (como la proteína y el sodio) o a efectos hormonales (como la carencia de estrógenos). Desde esta perspectiva, la tipología de la osteoporosis a nivel mundial resulta comprensible, si bien será necesario realizar una labor sobre el terreno con objetivos cuidadosamente seleccionados para completar el cuadro.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}