![]()
![]()
![]()
by
V.S. Kirpichnikov
Candidate of Biological Science
I. Introduction
Selection is the improvement of hereditary features of plants and animals for man's benefit. As a result of long-term selective work, breeds of domesticated animals and species of cultivated plants have been developed which more or less differ from their ancestors. The growth of the population of the earth, together with development of industry, makes domestication of animals and plants as well as rearing of more productive breeds and species more and more important. We have seen the completion of the process of transition from obtaining animals by hunting to their cultivation in all the countries of the world. The only exception is fishery management, the bulk of fish products still being obtained by “hunting”, mostly in open seas and oceans.
Until recently, utilization of fish resources in the sea had been inadequate and there is still ample potential for expansion of ocean fishing. The situation in inland water fishing is quite different.
The downward trend of catches in inland waters is quite obvious.
The development of industry and pollution of water bodies thus inhibiting normal fish stock reproduction, are the main causes.
The only way out is domestication and breeding of new races of fresh-and salt-water fish adapted for habitation in ponds, small lakes and water reservoirs, as well as for living and feeding in tanks and live-boxes
Up to now, comparatively few species of fish have been domesticated. The carp (Cyprinus carpio L.) and the closely related species C. fossicola are the most widely distributed species in the countries of Europe and Asia. Different races of the carp reared in a number of countries are used for culturing.
In China and Japan a great number of gold-fish races, obviously descending from the silver crucian (Carassius auratus gibelio Bl.), have been cultured. These races are exclusively decorative and we shall not list them.
The rainbow trout (Salmo g. irideus Gibb), is being cultured in North America, Europe and Asia on a large scale. This pond fish species was produced by crossing two or three species (or sub-species) of the American trout. The selection of rainbow trout proved to be rather fruitful.
The other species of fish, which are cultured in ponds and are domesticated to a certain degree, do not differ at all from their “wild” ancestors. Such species are:
Tench (Tinca tinca L.) German Democratic Republic (G.D.R.), Poland, and the U.S.S.R.
White and black amur (Ctenopharyngodon idellus) Val.) Mylopharyngodon piceus (Rich.) - The Chinese People's Republic, Viet-Nam and neighboring countries.
Hypophthalmichthys molitrix Val., Aristichthys nobilis (Rich.). Same countries.
Indian carp (Catla, Labeo) - India.
Gourami (Osphronemus gourami L.) - Indonesia and the Philippines.
Tilapia (Tilapia mossambica and other species) - tropical and north Africa, south Asia.
Sunfishes (Micropterus salmonides Lac., Lepomis macrochirus Mitch., etc.) - North America.
A few species with insignificant local distribution and some species whose domestication was started only recently could complete the list. Among the U.S.S.R. fish, two species of the Siberian whitefish, Coregonus peled Gm. and C. nasus Pall, are included in this group.
Actually, domestication of the Far East phytophagous fish, the white amur, and Hypophthalmichthys molitrix Val. started only in 1960, when propagation of these species in ponds became possible.
Carp selection should undoubtedly be considered the most effective and important in fish selection practice. Cultured races of carp are bred on a wide scale in many countries of the world and some countries have developed completely independent races (the U.S.S.R., German Democratic Republic, China, Indonesia, Viet-Nam). For this reason we shall specially dwell upon methods and results of carp selection and the problems facing carp culturists. The problems of trout and other pond fish selection will also be discussed briefly.
II. Carp Culturing and Races in Different Countries of the World
China's carp culturing is evidently the most ancient, its history covering about 2,000 years. The main object of Chinese pondfish culturing is not the carp but two species of the amur and Hypophthalmichthys molitrix Val. and Aristichthys nobilis Rich. The Chinese carp apparently differs but little from the wild river carp. In China, as far as we know, there are local dwarf froms of carp which reach sexual maturity while small in size and are adapted to the conditions of rice field pools.
The methods applied in Vietnamese carp culturing are also semi-cultivated, carp fry being mostly collected in rivers. According to data by Trân Tinh Trong (unpublished), there are no less than six varieties of carp in North Viet-Nam. All these varieties, according to F. Trong and V. Kirpichnikov (unpublished), belong to another species of Cyprinus (C. fossicola) genus which differs substantially from the European or east Chinese wild carp in a number of morphologic characters. This species has not yet been studied sufficiently well.
Carp culturing in Indonesia is also several centuries old, being particularly well developed in Java. There are descriptions of several races of carp, reared by the natives as follows: Sinjonja, Ikan mas, Tjuko, Punten, etc. (Iashouv, 1956; Steffens, 1962). These races were developed as a result of a long selection process but we know nothing of the methods used in Indonesia. The growth rate of the European mirror carp translocated to Java is lower than that of the local carp under conditions at low altitudes in the tropical climate but it grows faster than the local variety in the mountains. (Buschkiel, 1933).
The position of the Indonesian carp in fish systematics has not been established.
European cultured carp differ from their ancestors, the Danube wild carp, in a higher growth rate, their body shape, their mouths are larger, their intestines are longer, their plant feed mixture conversion is better (Schaperclaus, 1958; Rudzinsky, 1961; Steffens, 1964a, 1965b). Though the above differences are obvious and the carp was domesticated as early as six centuries ago, culturing of new carp races has made but little progress.
Until 1941 German fish culturists would refer to five “races” of the carp, which were actually race groups, differing in ranges and some exterior characteristics (Walter, 1901; Eleonsky, 1946).
The so-called “Eischgrund” or “Plate” carp, had the most distinct race characteristics but at the present time it has lost nearly all its importance. Based on the mixing of the remnants of several “races”, the German sparse-scaled carp is being successfully cultured in the German Democratic Republic.
Selection of the carp has begun in Poland. In other countries of Eastern Europe selection of the carp is in the embryonic stage.
Certain achievements have been scored in carp culture in the U.S.S.R. We can list several races and race groups cultivated in this country within recent years. They are:
Ramchaty (frame-scaled) and scaled Ukrainian carp, which were cultured and improved by a group of Ukrainian specialists headed by A.I. Kuzena for over 35 years.
Ukrainian carp is a race widely spread in the south of the U.S.S.R. and famous for its high growth rate.
Hybrid race groups, created on the basis of hybridization of the cultured carp (evidently of Galician origin) with the Amur wild carp of the Far East subspecies of this species (Cyprinus carpio hacmatopterus Temen and Schl.). The Amur wild carp with the cultured carp and hybrid rearing made carp culturing feasible in the northwestern parts of the U.S.S.R. and Siberia.
Most progress in hybrid selection (the fifth generation in 1964) was made in the northwest near Leningrad, where there are already several thousands of mature hybrid spawners (“Ropsha” carps, c.f. Kirpichnikov, 1965).
The second way of utilizing hybrids of the carp with the Amur wild carp is commercial crossbreeding, i.e., commercial rearing of the first generation hybrids. In this case heterosis higher vitality and growth rate of hybrids is used for attaining the goal. (Kirpichnikov, 1959). Commercial crossbreeding is practiced now in several republics: the Ukrainian SSR, Belorussian SSR and Latvian SSR.
West Ukrainian carp culturing was started by A.I. Kusema about 15 years ago. The west Ukrainian stock was obtained by crossing the Ukrainian carp with the local low bred carp; the selection work is still going on.
In addition to the above race groups, there are spawner stocks in the U.S.S.R. which, despite their potentialities, have been of no practical value so far. They include crosses between the Ukrainian and Ropsha (Northern) carp (created in the Ukraine), some spawner stocks near Moscow (subject of K. A. Golovinskaya's work), partially productive “triple” hybrids cultured by A.K. Kusema (the result of successive crossing of the carp with the common crucian and gold fish), the Belorussian carp created by D.P. Poliksenov and some other stocks.
III. Trout Races and Their Selection
The rainbow trout is the most popular fish cultivated in Denmark and the U.S.A., and it is also cultivated in Czechoslovakia, Japan, Korea and some other countries. Trout culturing has also started in the U.S.S.R. Fish culturists of North America have achieved great success in this field. During their 25 years experience they created a race characterized by a very high growth rate (the growth rate was doubled) (Donaldson and Olson, 1956). Good results have also been scored in Denmark, the Danish trout being one of the best races in the world. Morphologic characters of the Danish and American trout have not been studied yet and it is impossible to say whether they may be considered new races. In other countries, the U.S.S.R. particularly, trout selection has just started and it would be premature to discuss the results of this work.
The great diversity of species and varieties within the Salmo genus as well as the trout responsiveness to selection looks promising for culturing new trout races in many areas in the future.
IV. Goals of Fish Selection
The following problems are major tasks facing fish selection experts:
Culturing of fish with better feed conversion.
Culturing of fish which ensure more complete utilization of pond feed organisms.
Culturing of races resistant to various unfavorable influences of the environment (high or low temperature, oxygen deficiency, higher salinity, etc.).
Culturing of races resistant to various parasitic and infectious diseases.
Improving the taste qualities and raising the nutritional value, thus improving the commercial qualities of the fish.
Better feed conversion and higher resistance of fish to unfavorable conditions should be the major determining factors in selection work. Solution of these problems will ensure reaching the chief goal, the raising of productivity of rearing fish in tanks and live-boxes.
In various countries and climatic zones, the aims of selection of the same fish could be quite different. Thus, in the north and central U.S.S.R. the carp culturist faces the very important task of raising the cold resistance of the carp and developing the ability of feeding and growing at low temperatures. In more southern countries, like the Chinese People's Republic, Korea, Viet-Nam, Israel and Japan, it is essential to condition the carp to higher temperatures and to develop a slower maturing rate (Kessler, Wohlfarth; Lahmana, Moav, 1961).
The latest accomplishments in genetics, immunology and bio-chemistry offer good prospects for raising fish resistance to serious infectious diseases, such as, fish measles in carp, twist disease in salmon, ichthyophthiriasis, etc. Resistance to a certain parasite in some animals and plants was obtained by only one mutation, which brought about albuminous incompatibility of the tissues of the parasite and host. Speeding up the process of mutation, one may expect similar results with fish.
The problems confronting the selectionists dealing with various species of fish have much in common, though specific in each particular case. The methods of selection are much alike in many respects and we now consider them.
V. Methods of Fish Selection
a) Mass selection 1
When practicing mass selection, only the best specimens for the characters being selected are preserved for propagation. The easiest method is selection by the length and weight of the body.
When selecting according to exterior characters (body shape), it is always necessary to fully realize what particular exterior is the best. Mass selection by vitality is effective, provided that the majority of specimens perish, but this condition is often impracticable.
Direct mass selection according to such characters as the degree and rate of food assimilation, intensity of metabolism, etc. is very difficult for practical realization but the above characters also improve when fast growing specimens are selected. Selection by biochemical composition of meat and taste qualities is still more difficult, because it requires working out methods for biochemical analysis of meat during the lifetime of the fish.
Mass selection can be easily expressed quantitatively. For this purpose it is necessary to introduce some symbols.
Rate of selection (p) - percentage of specimens preserved for the stock to specimen initial number:
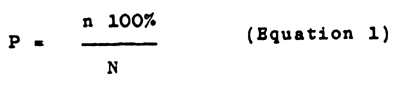
Where, N and n - numbers of specimens before and after selection.
Selection differential (S) - value, characterizing the change in the character as a result of selection. For instance, if out of 1,000 carp specimens (average weight 500 g) reared in one pond, 100 specimens (average weight - 600 g) are selected, we have
S = 100 g
and p = 10 percent
Intensity of selection (i) - selection differential expressed by means of standard deviations:
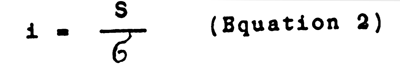
This value is more convenient than the selection differential when comparing selection in different fish populations and selecting according to different characters.
Effectiveness of selection (R) - value by which the character changes in the next generation of the selected specimens progeny. This value depends on the selection differential and the degree of heritability of the character in question:
R = S h2 (Equation 3)
where, h2 - heritability expressed in fractions of the unity: Substituting S = i . σ , we obtain: R = i. σ h2
This equation is particularly important because it shows that the greater genetic variability in the population (expressed by the product h2), the greater the selection effectiveness is. The magnitude of “i” depends on the rate of selection. For instance, at p = 20 percent the intensity of selection (with ordinary “normal” variation of the character) equals 2, but at p = 0.2 percent (selection of two specimens out of 1,000) the intensity of selection exceeds 3.
Formulas three and four suggest that the effectiveness of selection may be raised in the three ways:
By raising the rate of selection (by decreasing the value of “p”) and as a result of it, by increasing its intensity “i”.
by increasing the character variability (σ), provided that it does not entail a reduction in heritability, and
by increasing heritability (h2), i.e., by reducing the influence of the environment upon the character in question.
A high rate of selection (p = 1 percent and less) is readily attainable with highly fecund fish (the carp, crucian, amur Hypophthalmichthys molitrix and pike perch). The white fish and salmon lay a limited quantity of eggs, for which reason the rate of selection in their case cannot rise very high.
The character variability (genetic variability is implied) increases as the result of crossings, whereas in the case of in-breeding it decreases, crossing being a very important factor in selection.
Heritability corresponds to the share of herditary (genotypic) variability: in aggregate (phenotypical) variability:
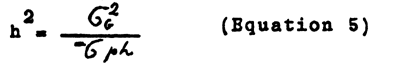
where σG and σph standard deviations of genotypical and phenotypical variability 1. Squares of deviations (Variances σ2) define variability better than simple variances (σ).
Heritability of weight difference with various species, races and varieties of fish varies within a wide range, changing with age as well. For the carp of the Q + and I + age group, the weight heritability does not, evidently, exceed 0.2 (Kirpichnikov 1958a; Moav and Wohlfarth, 1963).
1 Heritability is determined more exactly according to the formula:
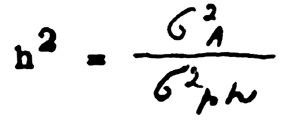
where, - variance of the so-called “additive” i.e., arithmetically summed hereditary variability. The value σ2A is less than σ2G but the difference is relatively insignificant and in many cases substitution of one variance by another is permissible
We have every reason to suppose that the weight heritability in the group of “record-breakers” may be still far less. (Nakoroku and Kasahara, 1955, 1957; Moav and Wohlfarth, 1963).
According to other characters, the magnitude of the heritability index is different. For instance, in the case of the number of vertebrae of the carp, it approximates 0.5 to 0.6 (Nenashev, 1965).
Heritability may be increased in two ways by distant crossings, leading to the increase of σ2G or by providing more uniform rearing conditions (by decreasing the magnitude of σ2ph).
The following is required when breeding carp:
Spawning should be quick and timely, otherwise it should be substituted by artificial fertilization.
Similar conditions should be provided for all specimens, from incubation of the spawn to the rearing of stock material.
Stocking of ponds should be simultaneous or as close as possible.
There should be no intermixing of the stocks from different ponds and of different age groups before selection.
There should be sufficient feeding and, consequently, less competition for food.
The principal aim attained by the above measures is minimizing hereditary differences caused by different environments or competition. Such “paratypical” differences may sometimes be correlated with genetic differences, but most often in the case of game fishes, such correlation is insignificant. Due to food competition, therefore, paratypical variability is chiefly increased and heritability is decreased. Even under the most favorable feeding conditions, the paratypical variability of fishes remains rather high. This being so, the most drastic method of raising heritability is crossing, which results in more heterogeneity of the population.
Until recently, mass selection was the main (if not the only) method of selection in European carp culture. The results of long-term carp selection practices appeared to be rather modest. The carp has not changed much since the time domestication measures were first taken. The effectiveness of selection was low for a number of reasons, the principal ones being:
failure to ensure the uniformity of the material.
Discrepancies in the growth rate of the carp at different ages.
Inbreeding of the carp.
Heterozygous balance.
Inadequate uniformity of the environment and mixing of the material prior to selection adversely affected the results of selection in the past.
However, there exists a possibility of reducing the inhabiting effect of these factors to a minimum in the future.
Irregularity of the growth rate of the carp at different ages is manifested in the fact that fish exceptionally large in their first year may become average size or even undersized in the second year and later.
In such cases, fingering selection will not improve two-year-old fish but the opposite. It is not difficult to avoid a lower selection effectiveness in such a case by concentrating on the age at which the character is to be improved.
Inbreeding causes a sharp drop in genetic variability and quite often a general depression.
There are methods applicable to animal selection to avoid the disadvantageous inbreeding effect. We shall consider them later.
The incidence of heterozygous may sometimes bring selection results to naught. This phenomenon, as it has now become clear (Lerner, 1954) is incident in animal populations quite often. The essence of it is that the higher heterozygosity of specimens turns out to be advantageous (in terms of better survival) and, as a result, a better portion of the population consists of heterozygotes. In such cases only more heterozygous specimens will be selected and selection will but maintain the already attained leval of growth and vitality. In such cases, good results can be achieved only by new, sufficiently distant crossings to upset the balance. In our estimation, mass selection as a method of selective breeding will undoubtedly remain one of the basic methods of race perfection in fish culture. Specific ways of its application depend on the individual features of the object and the goals set for the selectionist. Mass selection will always be expedient in the case of the high heritability of the character and, particularly, if a high rate of selection and, consequently, a greater selection differential could be attained. It is possible to determine, though only approximately, the effectiveness of mass selection and to compare it with that of the other selection methods by means of the above formulas.
Mass selection by such indices as the growth rate is expedient only with a sufficiently high selection differential, whereas selection by characters of higher heritability may be effective with a low rate of selection as well. But in the majority of cases it is most expedient to combine mass selection with genotypic selection, which is described further here.
b) Genotypic selection
Genotypic selection, i.e., according to the qualities inherent in relatives, is often referred to as “individual”, in Russian scientific literature.
As it may lead to misunderstanding, we shall not use the term. We shall consider two most important methods of genotypic selection.
Family selection. Family selection is comparing several families and selecting the best ones. A comparative appraisal of the families requires a large number of ponds. If a total increase in the fish output is to be evaluated per unit of pond area, separate rearing will be required. In this case, for a triple evaluation of only four families of the carp, 12 nursery, 11 fingerling rearing and 12 feeding ponds will be needed; in trout culturing, ponds may be replaced with the same number of live boxes and running water tanks. But four families constitute a very small number and one cannot expect that a substantial selection differential can be attained.
When selecting by characters which require dissection, the family selection is converted to sib-selection, i.e., estimation and selection of brothers and sisters. A certain number of specimens of every family are examined; brothers and sisters (sibs) of the dissected specimens from better families are preserved for propagation.
The main reason for practicing family selection is the low heritability inherent in the character under selection.
This type of selection yields good results only in case various families are provided with as uniform conditions as possible. The following procedure of family selection may be suggested to provide such uniform conditions:
Five to ten couples are crossed with the aid of artificial fertilization. Requirements for females are the same age and rearing conditions.
Eggs are incubated in apparatus, on frames or in crystallizers (the procedure should be repeated two or three times), the conditions being thoroughly equalized.
Additional separate rearing of larvae in aquaria, tanks and small ponds (the procedure should be repeated two or three times).
Regular rearing in ponds or live boxes under the same conditions. Separate rearing will require 15 to 30 ponds, whereas joint rearing (every pond is stocked with 5 to 6 generations) will require 3 to 6 ponds (when the procedure is repeated three times). Joint rearing is possible, subject to marking all of the fishes and their weights being equal at the time of stocking. Food competition can be avoided only by separate rearing.
Family selection is hindered by technicalities in many spheres of fish culture and particularly in the sphere of carp selection.
Progeny testing has become the most important method of breed improving in many branches of live stock breeding. Experiments of carp spawner evaluation were conducted by A.G. Kusema (Kusema, 1961, 1962) and V.S. Kirpichnikov (Kirpichnikov, 1959, 1960, 1961, 1965). The test pattern of checking males in our experiments was as follows and the same pattern is applicable when checking females:
We used a pituitary injection and artificial fertilization, dividing eggs into portions, their numbers corresponding to the number of males. A detailed description of the test method has been published, therefore, it should be added only, that comparison of the progeny by every male under test should be carried out under thoroughly levelled conditions of rearing at all stages and the procedure should be repeated, as required.
One of the baffling effects involved in evaluation of females is the strong “mother effect”. The quality of eggs tells on the progeny quality during a comparatively long period (2 to 3 months) and the progenesis have to be bred until two years of age.
The common shortcoming in progeny testing of males is the requirement of 1 to 2 years for obtaining exact data.
As to fish species whose males reach sexual maturity earlier than females (carp, for instance) males may be progeny tested without slowing down the selection process. In carp culture we recommend combined selection which makes up for time losses. Combined selection comprises the following stages:
Mass selection among the progeny obtained after the initial crossings;
Progeny testing of the best males of the first generation;
Family selection based on the material of 5 to 6 crossings of the phenotypically best females with the genotypically best males;
Mass selection in the best families of the second generation, etc.
We know the selection effectiveness is determined by the equation
R = S - h2
In the case of genotypical selection and, particularly, family selection, S decreases (it is impossible to breed many families), whereas h2 shows a sharp rise (not a single specimen but the whole family as evaluated). The heritability of the family mean values may be brought nearly to I, provided the procedure repetition rate is sufficient and the ponds are similar. It promotes better results of selection, even when a small number of families are involved in the experiment. Combined selection may yield still better results but its application is limited.
c) Crossbreeding, heterosis and inbreeding
Any animal race is heterogenic and fish are no exception. (Kirpichnikov, 1958). It is also true of fish that increased heterozygosity, in most cases, brings about a somewhat higher vitality and growth rate. Consequently, in selection work it is necessary to practice crossbreeding widely. We know of several such applications.
1. Commercial hybridization, i.e., crossbreeding of two races or lines for the purpose of commercial rearing the first heterosis generation. Inbred lines are often used in such cases. In fish culturing, crossing of highly inbred lines is possible only in the cases of species with fast developing sexual maturity. As to species like the carp and amur, selection may be confined to moderate crossing of inbred races or even outbred ones, provided their relation is distant enough. Our experiments (Kirpichnikov, 1957; Andrinsheva, 1965) as well as investigations by other authors (Kusema and Tomilenko, 1962) show that even in such cases a considerable heterosis may be incident.
2. Crossbreeding for New Synthetic Race Development
Quite often the aim of crossing is to combine in hybrids the characters of two existing races or varieties.
Crossbreeding of the European carp with the Amur wild carp and the subsequent selection of hybrids which developed the race group of the Ropsha carps (Kirpichnikov, 1965) may serve as an example of such selection. Initial crossing was conducted in order to obtaine a progeny combining the best qualities of the European carp and wild carp - the high growth rate and cold resistance.
It should be emphasized that synthetic crossing entails a long-term selection (for no less than 5 to 7 generations), required for the stabilization of the characters of the newly-bred race.
3. Crossbreeding for Raising Genetic Variability and Killing Inbreeding Depression
Outbreeding and often rather distant crossbreeding, increase gentic variability (the magnitude of σG rises). This provides the selectionist with material and is therefore of vital importance.
It is essential to select a system of rearing that would maintain sufficient heterogeneity and preclude inbreeding depression.
It is impossible to offer a universal method of selection but the two or three line system of breeding should be considered the most effective in carp culturing. It requires two or three parallel layers, each permitting moderate inbreeding; it also implies selection and evaluation of spawners. Following 3 to 4 generations, the best spawners of the unrelated layers are crossed to found new isolated lines or layers.
d) Other methods of Selection
Direct application of data on fish genetics may play a very important role in selection. Thus the removal of gene S from the Ropsha scaled hybrid carp, which is responsible for the formation of mirror scale, became feasible due to the previous check backcrosses:
SS × ss
and
Ss × ss
Having checked over 400 spawners, we could single out S-homozygous specimens. They were selected for reproduction and we managed to get rid of splitting completely (Kirpichnikov, 1961; 1965). Unfortunately, fish genetics has not yet been sufficiently developed and but few species have been investigated.
Speeding of the mutation process has not been tested on fishes so far but, as powerful chemical mutagenes are available, considerable achievments can be expected in this field in the not too distant future.
Gynogenesis (development without the sperm nucleus) and androgenesis (development without ovum nucleus) may contribute to the solution of some special problems of selection. Gynogenesis was effective in carp culturing (Golovinskaya, Romashov, Chefras, 1963) due to application of large X ray doses for destruction of sperm nuclei (up to 200,000 R).
Distant hybridization has been utilized repeatedly in fish selection. Some hybrids were made fecund (crosses of the carp with the crucian carp and gold fish). Crosses between Coregonus peled (Gm) and Coregonus nasus (Pall.) were fecund from the very beginning (unpublished data by G.A. Golovkova and G.I. Kiakova). New synthetic races can be cultured on the basis of such crossbreeding, provided normal fecundity is restored.
These are some special selection methods applied in fish culture.
Conclusions
1. The number of domesticated freshwater fish species is limited, whereas cultured races are still fewer (the carp, rainbow trout, tench, silver crucian). A great number of species are being domesticated.
2. The main objectives of fish selection:
creation of races with the best food conversion,
raising of fish resistance to unfavorable influences of the environment.
3. Mass selection was the principal method of selective breeding in the past and is similarly important today; but it calls for meeting some requirements entailing a higher effectiveness of selection.
4. Genotypical evaluation in fish breeding is hampered by great technical difficulties. The best solution of the problem in many cases is combined selection, a combination of mass selection with evaluation of families (family selection) and progeny checking of spawners.
5. Commercial crossbreeding is useful when the first generation hybrids display a considerable heterosis. Commercial crossbreeding has not been largely applied in fish culture but it is a method with great prospects.
When practicing synthetic crossbreeding with a view to subsequently improve hybrids, the heterogeneity of the race should be preserved through a number of generations. The best solution would be the development of 2 to 3 lines (layers) and rearing the lines with periodical interline crossbreedings: in such cases no “inbreeding depression” occurs, which is so frequent in inbreeding.
7. As to the other selection methods, utilization of fish genetics data should be noted as well as speeding up of the mutation process, gynogenesis and distant hybridization. The first method was applied when selecting the Ropsha carp; the remaining ones have not found application so far.
BIBLIOGRAPHY
Andriasheva, M.A., 1965 Heterosis at infraspecific inter-breeding of carp. “Information of the State Research Institute of Fresh Water Fisheries” (in press).
Buschkiel, A.L., 1933 Teichwirtschaftliche Erfahrungen mit Karpfen in den Tropen. Ztschr.f.Fischer., Bd. 31
Donaldson, L.R. and P.R. Olson, 1956 Development of rainbow trout brood stock by selective breeding. Trans.Am.Fish.Soc. Vol. 85
Eleonskaya, A.N., 1946 Pond fish culture. Moscow.
Falconer, D.S., 1960 Introduction to quantitative genetics. Edinburgh and London.
Golovinskaya, K.A., D.D. Komashov and N.B. Chefras, 1963 On the radiation gynogenesis in carp culturing. “Transactions of the State Research Institute of Pond Fisheries”, Vol. 12
Jashouv, A., 1955 The Punten carp and its attributes. Bamidgeh. Vol. 7, N 3
Kessler, S., et al., 1961 Monosex culture of carp. Bamidgeh, Vol. 13, N 3–4
Kirpichnikov, V.S., 1957 Creation of a new northern hybrid carp race. “Transactions of the Fish Culture Meeting”, Moscow.
Kirpichnikov, V.S., 1958 Heterogenic grade of wild carp populations and hybrids of wild carp with cultured carp. “Reports of the Academy of Science of the U.S.S.R.”, Vol. 122, N 4
Kirpichnikov, V.S., 1959a Genetic methods of the fish selection. “Bulletin of the Moscow Nature Society”, Biological Section, Vol. 64, N 1
Kirpichnikov, V.S., 1959b Hybridization of the cultured carp with the Amur wild carp applied for the western parts of the U.S.S.R. “Transactions of the V Inland Baltic Fisheries”. Minsk.
Kirpichnikov, V.S., 1960 Genetic methods of the fish hybrid selection. “Transactions of the Plant and Animal Hybridization Meeting”, Moscow.
Kirpichnikov, V.S., Die genetischen Methoden der Selektion in der Karpfensucht. Ztschr.f.Fischer, N.F., Bd. 10, H. 1–3
Kirpichnikov, V.S., 1965a First results and tasks for breeding business organization in the northwest of the U.S.S.R. Information of the State Research Institute of the Fresh Water Fisheries (in press).
Kirpichnikov, V.S., 1965b Checking methods of the spawners by the generations in the carp farms. “Information of the State Research Institute of the Fresh Water Fisheries”. (in press).
Lerner, I.M., 1954 Genetic homeostasis. N.-J.
Moav, R. and G.W. Wohlfarth, 1963 Breeding schemes for the genetic improvement of edible fish. Progress Report 1962, Jerusalem.
Nenashev, G.A., (in press) Heritability of some morphological (diagnostic) characteristics of the Ropsha carp. “Information of the State Research Institute of the Fresh Water Fisheries”.
Nakoroku, N. and S. Kasahara, 1955 On the earliest stage at which the shoot carp appears. Bull. Japan Soc.Sci.Fish., Vol. 21, N 2
Nakoroku, N., 1957 On the result of culturing the modal group and the growth of carp reared individually. Bull.Japan Soc.Sci.Fish., Vol. 22, N 11
Rudzinesky, E., 1961 Vergleichende Untersuchungen über den Wildkarpfen der Donau und den Teichkarpfen. Ztschr.f.Fischer, N.F. Bd. 10, H. 1–3
Schaperclaus, W., 1958 Lehrbuch der Teichwirtschaft, Berlin, 2 Auflage.
Steffens, W., 1962 Der Karpfen. Berlin, 2 Auflage.
Steffens, W., 1964a Der heutige Stand der Zuchtleistungen beim Karpfen in unseren Teichwirtschaften. Deutsche Fischer, Ztg., N11
Steffens, W., 1964b Vergleichende anatomisch-physiologische Untersuchungen an Wild - und Teichkarpfen (Cyprinus carpio L.). Ztschr.f.Fischer. N.F. Bd. 12, N 8–10
Walter, E., 1907 Die Karpfenrassen (in Knaute, 1907. Das Susswasser. Neudamm).
![]()
![]()
![]()