![]()
![]()
![]()
INTRODUCTION
Nutrition of domestic ruminants in the tropics is mainly based on the exploitation of rangeland resources which are subject to high quantitative and qualitative variations over the year. Fodder trees and shrubs are an integral part of the diet of these animals and constitute the main source of proteins, minerals and vitamins during the dry season.
Selection and intake of diet depends not only on the available plant resources but also on the feeding behaviour of the animals. Better understanding of feeding behaviour allows the development of management strategies aimed at maximizing the use of ecosystems for increased animal production.
Among domestic ruminants, camels are classified as browsers, goats as intermediate selective feeders with preference for browse, sheep as nonselective intermediate feeders with preference for grasses and buffalos, cattle and donkeys as grazers (McDowell, 1988; Schwartz and Schafft, 1988). These dietary preferences are however influenced by the constantly changing enviornment and human intervention in livestock management.
This paper presents an account of the effects of environmental factors on the feeding behaviour of domestic ruminants and the contribution of browse to their productivity. Based on this review, recommendations have been formulated which are aimed at maximizing the use of fodder trees and shrubs to increase animal production in developing countries.
FEEDING BEHAVIOUR
Most behaviour studies have been carried out by visual observation of grazing activity but also by examination of faeces, rumen contents and intake samples collected through oesophageal fistulae and evaluation of the forage offered minus the quantity left over. These studies have shown that feeding behaviour is controlled by factors of animal and plant interaction. The animal factors including species, physiological status, previous experience with the vegetation and habitat exploration, while the plant variables relate, among other things, to availability and the physical and qualitative characteristics of plant biomass. In grazing situations, the relative importance of all these factors can be modified by environmental variables such as human interventions which determine strategies of land use and stock management.
Dietary preference of domestic ruminants and effects of management decisions on feeding behaviour.
Some results of the behaviour studies carried out with cattle, sheep and goats by one of the authors in the arid and semi-arid programme of International Livestock Centre for Africa (Dicko et al., 1983; Dicko and Sangare, 1984) will be used here to describe the complex interaction between animal, forage availability and management decisions.
The study site in the sahelian zone of Mali has an average annual rainfall of 580mm occurring during the July-September period. The notable features are: (i) an irrigated land of 45,000 ha devoted to rice cultivation where a few trees and shrubs are found along irrigation ditches as well as around villages; and (ii) a dryland with villages and millet fields. The behaviour studies concerned:
The preference of the cattle for grass rather than for browse is obvious in Figures 1 and 2, particularly during the wet season when the cattle spent more than 95% of their grazing time on grass despite the simultaneous availability of abundant browse with the grass. Browsing started to increase at the beginning of the dry season (October) but this intake was not sustained by the cattle under semi-sedentary management (Figure 1) because of their shift to millet, then to rice fields where there were relatively few trees and shrubs. Conversely, with the cattle under the sedentary system (Figure 2), browsing in the dry season increased steadily, with a peak at the close of the season (June) because of the flush of a great number of trees.
The increase in the grazing time during the dry season in Figure 1 and the reverse trend in Figure 2 are due to the influence of herd management on grazing activity. In the semi-sedentary system, the herdsman always settled on an open pasture near a watering point convenient to both his family and cattle, while in the sedentary system the permanent kraal and watering point had an eccentric position relative to pastures. In consequence, the semi-sedentary cattle spent less time walking to and from pastures than did the sedentary animals. In addition, they were able to practise night grazing because of the closeness of the pasture lands while in the sedentary system, long walks and lack of night grazing had a depressive effect on grazing activity.
Figure 3a and 3b constitute good examples for classifying sheep as intermediate feeders with preference for grasses and goats as intermediate feeders with preference for browse (Schwartz and Schafft, 1988). While grazing together, these two animal species behaved according to their dietary preferences. The time spent by the sheep in browsing varied with the season from 2 to 76% of their grazing time while that spent by the goats ranged from 73 to 93%. The sheep, like the sedentary cattle, ate browse mainly during the dry season, with the time spent on browse increasing throughout the season.
Figures 4a and 4b demonstrate the influence that low availability of browse plants has on the feeding behaviour of the sheep and particularly the goats. Because of limited number of fodder trees and shrubs on the irrigated land, the time the goats spent on browse varied over the year from 4 to 93% of their grazing time. A browsing duration of only 4% of their daily grazing time is abnormal with goats. Such a great modification of feeding behaviour was brought about in this system by human intervention.
Among domestic ruminants, the most effective browsers are camels. In the arid zones of Kenya, Lusigiet al. (1984) have found that more than 50% of the tree species and 55% of the shrub species were utilized by camels, while only 14 to 19% of the perennial grasses were eaten. There is a great overlap between the diet of camels and goats but little competition in browse utilization occurred between the two species owing to the superior reach of the camels (Keya et al., 1991).
Preference for specific plant species and seasonal changes in palatability of trees and shrubs are also some features of the feeding behaviour of domestic animals. In addition, some intrinsic characteristics of trees and shrubs such as glabrousness, toughness, hairs, thorns and presence of deleterious substances may also influence palatability.
A better understanding of the feeding behaviour of domestic animals allows the development of management strategies aimed at maximizing the use of ecosystems for increased animal production.
Distribution of grazing time among various types of forages.
FIGURE 1. Semi-sedentary cattle.

FIGURE 2. Sedentary cattle in dry areas.
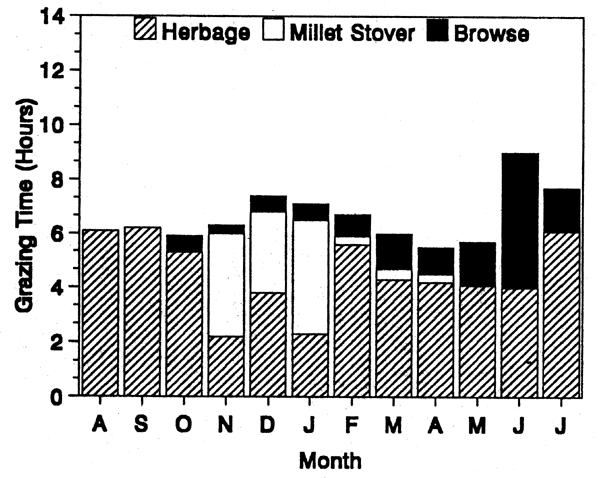
Distribution of grazing time among various types of forages.
FIGURE 3a. Sedentary sheep in dry areas.

FIGURE 3b. Sedentary goats in dry areas.
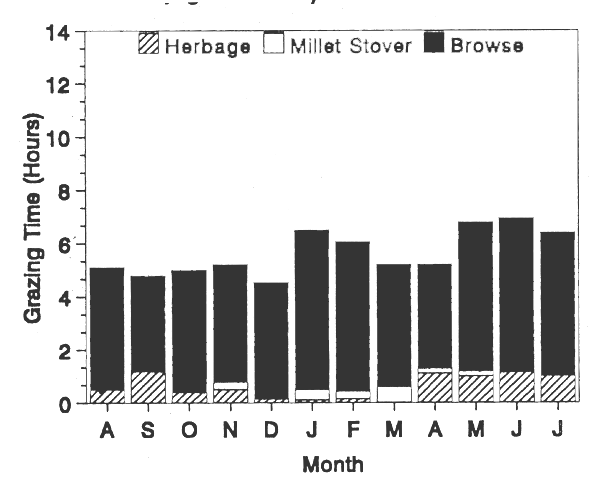
Distribution of grazing time among various types of forages.
FIGURE 4a. Sedentary sheep in irrigated areas.

FIGURE 4b. Sedentary goats in irrigated areas.
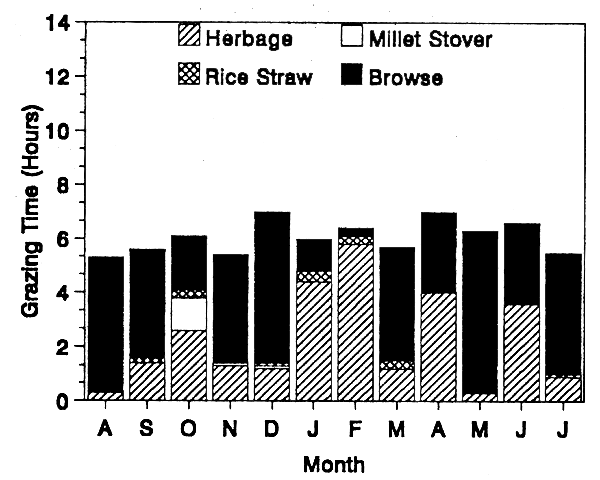
NUTRITIVE VALUE
Compared to grasses, fodder trees and shrubs have relatively higher concentrations of crude protein, minerals and neutral detergent fibre plus acid detergent lignin, while their average concentration in acid detergent fibre, as well as their average dry matter digestibility, are both lower. These nutrient contents are subject to less variation than with grasses and this particularly enhances their value as dry season feeds for livestock (Wilson, 1977; Ibrahim, 1981).
Protein
A significant variation in crude protein (CP) content occurs between species of trees and shrubs and even between edible parts of the same plant. Guérin (1987) reported a variation of CP content from 6 to 23% in dry matter (DM).
In general, leaves are higher in CP than twigs, almost twice as much in the case of southern African browse according to Walker (1980). They also contain more CP on average than pods but the latter were found with higher organic matter and digestibility (Göhl, 1981).
Leguminous species were found to contain 25 to 50% more crude protein than non-leguminous plants (Wilson, 1969; Nitis, 1989). Leaves of phyllodenous acacias tend to have higher crude fibre, lower crude protein, phosphorus, organic matter and digestibility than other acacias (Goodchild and McMeniman, 1987).
In Saudi Arabia, Mirreh and al Daraan (1991) found that perennial shrubs retained high CP content for a longer period than did annual shrubs. The CP in perennials varied from 14% in spring to 7% in winter while, in annual ones, a variation from 22% to 8% was recorded during the same spring season.
In addition to seasonal changes, environmental factors such as fire, substratum and amount of tree cover also influence nitrogen content. Barbero et al. (1991) reported a difference of 20% between the CP contents of an underwood calcicolous stand and an open silicicolous stand in a Mediterranean environment. Lay (1967) found that CP content in browse increased by 42.2% after burning. However, most of the benefits disappeared within a year or two.
Minerals and vitamins
The concentration of calcium and potassium is usually higher than that of other minerals; the average being around 1 to 1.5% for southern African browse (Walker, 1980). However, abnormally high concentrations of some minerals such as sodium chloride and selenium may be found in browse (Underwood, 1981; Olsson and Welin-Berger, 1989).
Vitamin A, B, C and PP are present in varying concentrations in leaves, fruits and seeds of some trees and shrubs (Bergeret, 1986). Carotene content is usually high during early growth and declines rapidly with maturity, except in evergreen shrubs which tend to retain it for long periods (Ibrahim, 1981).
Digestibility
Dry matter digestibility (DMD), which is related to nutrient composition, varied widely among tree and shrub species. Skarpe and Borgstrom (1986), working in Botswana with Kalahari woody species reported a range in digestibility from 38 to 78%. Similar findings were reported by Wilson (1977).
The digestibility of cellulose and cell walls decreased as the lignin to cellulose or lignin to acid detergent fibre ratio increased (Robbins and Moer, 1975). The digestibility of crude protein did not always match the high CP content which characterizes fodder trees and shrubs. For example Wilson (1977) found an apparent digestibility as low as 14% for Heterodendrum oleifolium containing 12.5% of CP while Atriplex vesicaria, also with 12.5% CP, had a nitrogen digestibility of 71.4%. Also there was not always a correlation between intake and digestibility; highly digestible stuff may be poorly consumed and vice versa (Wilson 1977).
Deleterious substances
Tannins, mimosine, cyanoglucosides, fluoroacetate, oxalate, selenium, saponin and sodium chloride are among substances which have the potential of decreasing feed intake, animal productivity and even causing toxicity at high levels ingestion.
Tannins relate to a large number of phenolic compounds whose deleterious effects are not uniform. Some compounds like condensed tannins (CT) even have positive effects on nitrogen utilization by protecting protein from microbial degradation. According to Barry and Manley (1985), CT concentration of 20–40mg/kg in DM is ideal. Higher levels (76–90mg) were found to be detrimental to the health of animals.
Detannification techniques include heating, wilting, treatment with chemicals, storage of fresh leaves in polythene bags and bio-treatment using the fungus Sprotrichum pulverulentum (Makkar, 1991).
Mimosine is found in Leucaena leucocephala. Its concentration, which varies with sub-species, is also affected by environmental factors such as soil and climate. Studies have shown that Leucaena can be used without harm as asupplementary feed for ruminants at a rate of 20–50% of the ration (Gupta et al., 1991; Atta-Krah and Reynolds, 1989). Differences in minosine concentration could explain such variation in results.
Domestic ruminants, however, were seen to be better adapted to the consumption of trees and shrubs with high content of deleterious substances, provided there was agradual ingestion of the browse. This phenomenon of adaptation, as well as the whole metabolism of antinutritional substances, still need further investigation (Schwartz and Schafft, 1988).
INTAKE
Domestic ruminants utilize trees and shrubs both as browse in situ and “cut and carried” branches in the stall. On rangelands, animals have the advantage of selecting from a wide choice of browse and obtaining high quality feed. Consumption of various types of forage reduces chances of poisoning. However, the effective contribution of trees to the nutrition of livestock is not always realised due to inability to reach the browsable canopy of tall trees.
The cut and carry system obviates the limitation of inaccessibility of the browse. The system also facilitates rational usage of trees and shrubs. Its disadvantages include damage to trees due to injudicious lopping and the imposition of a limited variety of feed which may increase the risk of poisoning.
Use of fodder trees and shrubs in stall feeding.
The period of adaptation (i.e., up to intake stabilisation) to new feed may be quite long, as shown in the studies of Le Houérou (1991) in Libya. The intake of 9 species of native and exotic shrubs offered either alone or in a mixture to ewes increased with time and stabilized after 3 to 5 months at levels 3 to 5 times the initial intake (increasing from 25 65g/kg0.75/day to 35–145 g/kg0.75/day). The author also reported that consumption of mixed shrubs was higher than that of a single species. Wilting for 24 to 48 hours also increased the intake.
Many trials on the use of shrubs and leaves of trees to supplement either natural grasses or crop residues gained positive responses in livestock performance.
Nitis (1989) reported a case where sheep and goats fed on Pennisetum purpureum supplemented with 0.3 to 1.8 kg Gliricidia per day gained 17– 27% more weight than the unsupplemented animals. With Bali cattle fed on 80% of natural grasses plus 9% of Leucaena and Musa, and 11% of tree leaves, the increase in weight was 58% more than that of the control group.
ILCA studies in Nigeria established that the use of Leucaena and Gliricidia as supplementary feeds significantly increased the growth and survival rates of lamb. Statistical analyses of the data showed that each 100g of browse DM consumed per day raises the productivity index by 1.41 kg lamb weaned/dam/year (Atta-Krah and Reynolds, 1989).
Unfortunately, the interesting findings on supplementary feeding are not adopted by small-scale farmers, particularly in sub-Saharan Africa. Investigations of the limitations on the adoption of this practice is recommended as a way to alleviate inadequate nutrition in domestic ruminants.
Exploitation of fodder trees and shrubs on rangeland
Owing to their dietary preference and the influence of animal size on the vertical utilization of plants, there is little competition among domestic ruminants in the exploitation in situ of fodder trees and shrubs. According to Ibrahim (1981), the proportion of browse eaten by cattle and sheep varies from 2–30% of their intake. With goats, the intake ranges from 25–50% in the rainy season and up to 75% or more during dry periods. However, as the above feeding behaviour study demonstrates, these estimates may vary widely.
Studies on the contribution of fodder trees and shrubs to the productivity of grazing animals are still limited. Reasons for this include difficulties encountered when implementing trials with paired areas (i.e., plots with trees and those without) and the negative effects of lopping or deforestation on the environment.
A study by Rees (quoted by Bingham, 1983) on the evaluation of browse potential in cattle production is one of the few reports available. She used steers on 3 types of paddocks: natural, lopped (all trees lopped) and cleared (all trees removed).
Table 1 shows that lopped paddocks had the highest fodder potential for increased animal production, particularly when young steers (1–2 years) were used. Comparison of the performance of the 2–3 year old steers suggests that the efficiency of habitat exploitation and feed utilization increased with age.
| Age of cattle | Woodland | Lopped | Cleared |
|---|---|---|---|
| 1–2 | 13.7 | 90.8 | 40.6 |
| 2–3 | 46.6 | 100.0 | 97.5 |
Source: Bingham (1983)
Further monitoring of the effect of lopping showed that, within 3 years, most of the trees managed for browse either die or grow beyond the reach of animals.
This study confirms that fodder trees and shrubs have potential for increased livestock production but has shown that the magnitude of the contribution to the productivity of grazing animals greatly depends upon accessibility of the browse. High accessibility is not often achieved in the grazing situation prevailing in the dry tropics which carry the majority of domestic ruminants. Increasing accessibility of browse entails human interventions which may be detrimental to woody plants. There is therefore a need to develop appropriate woodland management systems which will ensure optimum utilization concomitant with persistence of browse production in grazing systems.
RECOMMENDATIONS
In order to enhance the utilization of fodder trees and shrubs by domestic animals, there is a need to:
assess the suitability of animal species to their environment through better knowledge of their feeding behaviour. This will allow the development of management strategies aimed at maximizing the use of ecosystems;
carry out an exhaustive assessment of the nutritive values (including deleterious substances) of fodder trees and shrubs, particularly of those intended to be used in stall feeding;
conduct socio-economic surveys on the limitations of on-farm supplementation in certain developing countries;
work out appropriate woodland management systems which will ensure optimum utilization, concomitant with the persistence of fodder trees and shrubs in grazing systems.
Bibliography
Atta-Krah, A. N. and Reynolds, L. 1989. Utilization of pasture and fodder shrubs in the nutrition of sheep and goats in the humid tropics of West Africa. In: Sheep and goat meat production in the humid tropics of West Africa. FAO animal production paper 70.
Barbero, M., Hubert, B., Lebreton, P.H., Nader, S. and Quezel, P. 1991. Structure des écosystèmes buissonnants et arbustifs: variabilité phénologique et écologique au milieu méditerranéen. IVth International Rangeland Congress, Montpellier, France (in press).
Barry, T. N. and Manley, T. R. 1985. The role of condensed tannins in the nutritional value of Lotus pendunculatus for sheep. 3: Rates of body and wool growth. British Journal of Nutrition 54: 211–217.
Bergeret, A. 1986. Rôle alimentaire des arbres et arbustes et de quelques plantes herbaceés. VIIe séminaire d'Economie et Sociologie, Montpellier, France. 28 pp.
Bingham, M.G. 1983: Prospect for livestock production in northern Zambia. Seminar on soil productivity in high rainfall areas. Lusaka, 8–10 February 1983.
Dicko, M.S., Lambourne, J., de Leeuw, P.N. and de Haan, C. 1983. Nutrition. In: Recherches sur les systèmes des zones arides du Mali, Résultats préliminaires. ILCA, Addis Ababa, Ethiopia pp. 95–104.
Dicko, M. S. and Sangare, M. 1984. Feeding behaviour of domestic ruminants in Sahelian zone. Proceedings of 2nd International Rangelands Congress. Adelaide, Australia pp. 388–390.
Göhl, B. 1981: Tropical Feeds. FAO, Rome.
Goodchild, A. V. and McMeniman, N.P. 1987. Nutritive value of Acacia foliage and pods for animal feeding. In: Australian Acacias in Developing Countries. J.W. Turbull (ed), Australian Centre for Integrated Agricultural Research, Canberra. Proceedings Series no. 16. pp. 101–106.
Guérin, H. 1987. Alimentation des Ruminants domestiques sur pâturages naturels sahéliens et sahélo - soudaniens: étude méthodologique de la région du Ferlo au Sénégal. Thèse. École Nationale Supérieure Agronomique de Montpellier, France. 211p.
Gupta, B.K., Singh, A. and Malick, N.S. 1991. Effect of feeding dried Leucaena to buffalo and crossbred cow calves. IVth International Rangeland Congress, Montpellier, France (in press).
Ibrahim, K.M. 1981. Shrubs for fodder production. In: Advances in food producing systems for arid and semi-arid lands. Academic press Inc. pp. 601–642.
Keya, G. A., Woie, B. M. and Cheruiyot, H.K. 1991. Utilization of rangeland resources by camels in the arid zones of Kenya. IVth International Rangeland Congress, Montpellier, France (in press).
Lay, D. W. 1967. Browse palatability and the effects of prescribed burning in southern pine forests. Jour nal of Forestry 65: 826–828.
Le Houérou, H. N. 1991. Feeding shrubs to sheep in the Mediterranean arid zone: intake, performance and feed value. IVth International Rangeland Congress, Montpellier, France.
Lusigi, W. J., Nkurunzize, E.R. and McDowell, R.E. 1988. Importance of crop residues in feeding livestock in smallholders farming systems. In: Plant breeding and the nutritive value of crop residues. Proceedings of a Workshop held at ILCA, Addis Ababa, Ethiopia. 7–10 Dec 1987.
Makkar, H.P.S. 1991. Utilization of Oak tree leaves as livestock feed: an overview. IVth International Rangeland Congress, Montpellier, France (in press).
Masheti, S. 1984. Forage preferences of livestock in the arid lands of northern Kenya. Journal of Range Management 37: 542–548.
Mirreh, M.M. and al Daraan, M.S. 1991. Nutritional adequacy of native shrubs in northern Saudi Arabia. IVth International Rangeland Congress, Montpellier, France (in press).
Nitis, I. M. 1989. Fodder trees and livestock production under harsh environment. Asian Livestock, October 1989: 116–120.
Olsson, A. C. and Welin - Berger, S. 1989. The potential of local shrubs as feed for livestock and mineral contents of some soils and soillicks in central Tanzania. Two minor field studies. Arbetsrapport, Sveriges Lantbruksuniversitet No. 125, 25pp.
Robbins, C.T., and Moer, A.N. 1975. Composition and digestibility of several deciduous browses in the Northeast. Journal of Wildlife Management 39: 337–341
Schwartz, H.J. and Schafft, H. 1988. Integrated live stock tree cropping systems: A review of potential and constraints arising from the biology of animals. Proceedings of the International Livestock Tree cropping Workshop, 5–9 December 1988, Serdang, Malaysia pp 24–38.
Skarpe, C. and Bergstrom, R. 1986. Nutrient content and digestibility of forage plants in relation to plant phenology and rainfall in the Kalahari, Botswana. Journal of Arid Environments 11: 147–164.
Underwood, E.J. 1981. Mineral nutrition of livestock. Commonwealth Agricultural Bureau, UK
Walker, B. H. 1980. A review of browse and its role in livestock production in Southern Africa. In: Browse in Africa, the current state of knowledge. Le Houérou, H.N. (ed.), ILCA Addis Ababa, Ethiopia.
Wilson, A.D. 1969. A review of browse in the nutrition of grazing animals. Journal of Range Management 22: 23–28.
Wilson, A.D. 1977. The digestibility and voluntary intake of the leaves of trees and shrubs by sheep and goats. Australian Journal of Agricultural Research 28: 501–508.
![]()
![]()
![]()