![]()
![]()
![]()
4.1.1. Introduction
4.1.2. Biology and ecology of Artemia
4.1.3. Literature of interest
Gilbert Van Stappen
Laboratory of Aquaculture & Artemia Reference
Center
University of Gent, Belgium
Among the live diets used in the larviculture of fish and shellfish, nauplii of the brine shrimp Artemia constitute the most widely used food item. Annually, over 2000 metric tons of dry Artemia cysts are marketed worldwide for on-site hatching into 0.4 mm nauplii. Indeed, the unique property of the small branchiopod crustacean Artemia to form dormant embryos, so-called ‘cysts’, may account to a great extent to the designation of a convenient, suitable, or excellent larval food source that it has been credited with. Those cysts are available year-round in large quantities along the shorelines of hypersaline lakes, coastal lagoons and solar saltworks scattered over the five continents. After harvesting and processing, cysts are made available in cans as storable ‘on demand’ live feed. Upon some 24-h incubation in seawater, these cysts release free-swimming nauplii that can directly be fed as a nutritious live food source to the larvae of a variety of marine as well as freshwater organisms, which makes them the most convenient, least labour-intensive live food available for aquaculture. Although Artemia has been known to man for centuries, its use as a food for the culture of larval organisms apparently began only in the 1930’s, when several investigators found that it made an excellent food for newly-hatched fish larvae. During the 1940’s, most commercially available brine shrimp cysts represented collections from natural saline lakes and coastal saltworks. With the growing interest for tropical hobby fish in the late 1940’s, commercial value was attached to brine shrimp, thereby establishing a new industry. Early pioneers exploited in 1951 the cyst production of Artemia at the Great Salt Lake in Utah, USA. First harvests of the lake yielded 16 tons of finished product. During the mid-1950’s, commercial attention for brine shrimp was turned to controlled sources for production in the San Francisco Bay region. Here it was found that brine shrimp and their cysts could be produced as a by-product of solar saltworks. Since salt production entails management of the evaporation process, yearly cyst and biomass productions could be roughly predicted. In the 1960’s, commercial provisions originated from these few sources in North America and seemed to be unlimited. However, with the expansion of aquaculture production in the 1970’s, the demand for Artemia cysts soon exceeded the offer and prices rose exponentially, turning Artemia into a bottleneck for the expansion of the hatchery aquaculture of marine fishes and crustaceans. In particular, many developing countries could hardly afford to import the very expensive cysts.
At the Kyoto FAO Technical Conference on Aquaculture in 1976 it was claimed that the cyst shortage was an artificial and temporary problem. During the following years research efforts were made to prove the possibility of local production of Artemia in developing countries.
At present, Artemia is being produced and exploited on the five continents. Despite this, a large part of the cyst market is still supplied by harvests from one location, the Great Salt Lake. This situation makes the market still extremely vulnerable to climatological and/or ecological changes in this lake, which has been illustrated by the unusually low cyst harvests in the seasons 1993-1994 and mainly 1994-1995.
Already in the late seventies it appeared that the nutritional value of Artemia, especially for marine organisms, was not constant but varied among strains and within batches of each strain, causing unreliable outputs in marine larviculture. Through multidisciplinary studies in the eighties both the causes for the nutritional variability in Artemia and the methods to improve poor-quality Artemia were identified. Genotypic and phenotypic variation (i.e. cyst size, cyst hatching characteristics, caloric content and fatty acid composition of the nauplii) determine if a particular cyst product is suitable for hatchery use of specific fish or shrimp species.
By bio-encapsulating specific amounts of particulate or emulsified products rich in highly unsaturated fatty acids in the brine shrimp metanauplii, the nutritional quality of the Artemia can be further tailored to suit the predators’ requirements. Application of this method of bio-encapsulation, also called Artemia enrichment or boosting, has had a major impact on improved larviculture outputs, not only in terms of survival, growth and success of metamorphosis of many species of fish and crustaceans, but also with regard to their quality, e.g. reduced incidence of malformations, improved pigmentation and stress resistance. The same bio-encapsulation method is now being developed for oral delivery of vitamins, chemotherapeutics and vaccines.
Furthermore, a better knowledge of the biology of Artemia was at the origin of the development of other Artemia products, such as disinfected and decapsulated cysts, various biomass preparates, which presently have application in hatchery, nursery and broodstock rearing. All these developments resulted in optimized and cost-effective applications of this live food in hatchery production.
4.1.2.1. Morphology and life cycle
4.1.2.2. Ecology and natural distribution
4.1.2.3. Taxonomy
4.1.2.4. Strain-specific characteristics
In its natural environment at certain moments of the year Artemia produces cysts that float at the water surface (Fig. 4.1.1.) and that are thrown ashore by wind and waves. These cysts are metabolically inactive and do not further develop as long as they are kept dry. Upon immersion in seawater, the biconcave-shaped cysts hydrate, become spherical, and within the shell the embryo resumes its interrupted metabolism. After about 20 h the outer membrane of the cyst bursts (= “breaking”) and the embryo appears, surrounded by the hatching membrane (Fig. 4.1.2.). While the embryo hangs underneath the empty shell (= “umbrella” stage) the development of the nauplius is completed and within a short period of time the hatching membrane is ruptured (= “hatching”) and the free-swimming nauplius is born (Fig.4.1.3.).
Figure 4.1.1. Harvesting of brine shrimp cysts from a saltpond.
Figure 4.1.2. Cyst in breaking stage. (1) nauplius eye.
The first larval stage (instar I; 400 to 500 µm in length) has a brownish-orange colour, a red nauplius eye in the head region and three pairs of appendages: i.e. the first antennae (sensorial function), the second antennae (locomotory + filter-feeding function) and the mandibles (food uptake function). The ventral side is covered by a large labrum (food uptake: transfer of particles from the filtering setae into the mouth). The instar I larva does not take up food as its digestive system is not functional yet; it thrives completely on its yolk reserves.
After about 8 h the animal molts into the 2nd larval stage (instar II). Small food particles (e.g. algal cells, bacteria, detritus) ranging in size from 1 to 50 µm are filtered out by the 2nd antennae and ingested into the functional digestive tract.
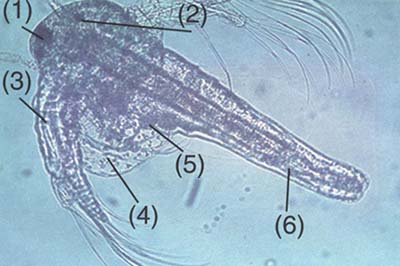
The larva grows and differentiates through about 15 molts. Paired lobular appendages are appearing in the trunk region and differentiate into thoracopods (Fig. 4.1.4.). On both sides of the nauplius lateral complex eyes are developing (Fig. 4.1.5. and 4.1.6.). From the 10th instar stage on, important morphological as well as functional changes are taking place: i.e. the antennae have lost their locomotory function and undergo sexual differentiation. In males (Fig. 4.1.6. and 4.1.8.) they develop into hooked graspers, while the female antennae degenerate into sensorial appendages (Fig. 4.1.11.). The thoracopods are now differentiated into three functional parts (Fig. 4.1.13.), namely the telopodites and endopodites (locomotory and filter-feeding), and the membranous exopodites (gills).
Adult Artemia (± 1 cm in length) have an elongated body with two stalked complex eyes, a linear digestive tract, sensorial antennulae and 11 pairs of functional thoracopods (Fig. 4.1.10. and 4.1.11.). The male (Fig. 4.1.10.) has a paired penis in the posterior part of the trunk region (Fig. 4.1.9.). Female Artemia can easily be recognized by the brood pouch or uterus situated just behind the 11th pair of thoracopods (Fig. 4.1.9. and 4.1.11.). Eggs develop in two tubular ovaries in the abdomen (Fig. 4.1.7.). Once ripe they become spherical and migrate via two oviducts into the unpaired uterus.
Figure 4.1.6. Head and thoracic region of young male. (1) antenna; (2) telopodite; (3) exopodite.
Fertilized eggs normally develop into free-swimming nauplii (= ovoviviparous reproduction) (Fig. 4.1.12.) which are released by the mother. In extreme conditions (e.g. high salinity, low oxygen levels) the embryos only develop up to the gastrula stage. At this moment they get surrounded by a thick shell (secreted by the brown shell glands located in the uterus), enter a state of metabolic standstill or dormancy (diapause) and are then released by the female (= oviparous reproduction) (Fig. 4.1.14.). In principle both oviparity and ovoviviparity are found in all Artemia strains, and females can switch in-between two reproduction cycles from one mode of reproduction to the other. The cysts usually float in the high salinity waters and are blown ashore where they accumulate and dry. As a result of this dehydration process the diapause mechanism is generally inactivated; cysts are now in a state of quiescence and can resume their further embryonic development when hydrated in optimal hatching conditions.
Under optimal conditions brine shrimp can live for several months, grow from nauplius to adult in only 8 days time and reproduce at a rate of up to 300 nauplii or cysts every 4 days.
Figure 4.1.9. Artemia couple in riding position. (1) uterus; (2) penis.
Artemia populations are found in about 500 natural salt lakes and man-made salterns scattered throughout the tropical, subtropical and temperate climatic zones, along coastlines as well as inland (Fig. 4.1.15.). This list still remains provisional as more extensive survey work should lead to the discovery of many more Artemia biotopes in different parts of the world (Table 4.1.1.). The distribution of Artemia is discontinuous: not all highly saline biotopes are populated with Artemia. Although brine shrimp thrive very well in natural seawater, they cannot migrate from one saline biotope to another via the seas, as they depend on their physiological adaptations to high salinity to avoid predation and competition with other filter feeders. Its physiological adaptations to high salinity provide a very efficient ecological defense against predation, as brine shrimp possess:
· a very efficient osmoregulatory system;· the capacity to synthesize very efficient respiratory pigments to cope with the low O2 levels at high salinities;
· the ability to produce dormant cysts when environmental conditions endanger the survival of the species.
Artemia therefore, is only found at salinities where its predators cannot survive (³ 70 g.l-1). As a result of extreme physiological stress and water toxicity Artemia dies off at salinities close to NaCl saturation, i.e. 250 g.l-1 and higher.
Different geographical strains have adapted to widely fluctuating conditions with regard to temperature (6-35°C), salinity and ionic composition of the biotope. Thalassohaline waters are concentrated seawaters with NaCl as major salt. They make up most, if not all, of the coastal Artemia habitats where brines are formed by evaporation of seawater in salt pans. Other thalassohaline habitats are located inland, such as the Great Salt Lake in Utah, USA. Athalassohaline Artemia biotopes are located inland and have an ionic composition that differs greatly from that of natural seawater: there are sulphate waters (e.g. Chaplin Lake, Saskatchewan, Canada), carbonate waters (e.g. Mono Lake, California, USA), and potassium-rich waters (e.g. several lakes in Nebraska, USA).
Artemia is a non-selective filter feeder of organic detritus, microscopic algae as well as bacteria. The Artemia biotopes typically show a very simple trophical structure and low species diversity; the absence of predators and food competitors allows brine shrimp to develop into monocultures. As high salinity is the common feature determining the presence of Artemia, the impact of other parameters (temperature, primary food production, etc.) may at most affect the abundance of the population and eventually cause a temporary absence of the species.
As Artemia is incapable of active dispersion, wind and waterfowl (especially flamingos) are the most important natural dispersion vectors; the floating cysts adhere to feet and feathers of birds, and when ingested they remain intact for at least a couple of days in the digestive tract of birds. Consequently the absence of migrating birds is probably the reason why certain areas that are suitable for Artemia (e.g. salinas along the northeast coast of Brazil) are not naturally inhabited by brine shrimp.
Next to the natural dispersion of cysts, deliberate inoculation of Artemia in solar salt works by man has been a common practice in the past. Since the seventies man has been responsable for several Artemia introductions in South America and Australia, either for salt production improvement or for aquaculture purposes. Additionally, temporal Artemia populations are found in tropical areas with a distinct wet and dry season (monsoon climate), through inoculation in seasonal salt operations (e.g. Central America, Southeast Asia).
Figure 4.1.15. The world distribution of Artemia.
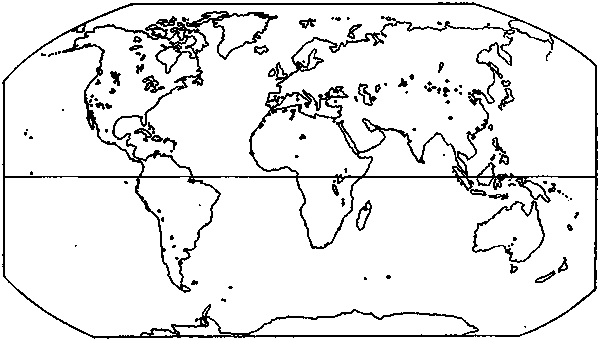
Table 4.1.1. World distribution of Artemia.
|
Country |
Locality |
Sex |
Species |
|
Artemia sites in Africa |
|
|
|
|
Algeria |
Chegga Oase |
- |
- |
|
Chott Djeloud |
- |
- |
|
|
Chott Ouargla |
- |
- |
|
|
Dayet Morselli |
- |
- |
|
|
Gharabas Lake |
- |
- |
|
|
Sebket Djendli |
- |
- |
|
|
Sebket Ez Zemouk |
- |
- |
|
|
Sebket Oran |
- |
- |
|
|
Tougourt |
- |
- |
|
|
Egypt |
Port Fouad |
B |
A. sal |
|
Wadi Natron |
B |
A. sal |
|
|
Qarun Lake |
P |
A. par |
|
|
Kenya |
Elmenteita |
- |
- |
|
Libya |
Mandara |
B |
A. sp |
|
Ramba-Az-Zallaf (Fezzan) |
- |
- |
|
|
Quem el Ma |
- |
- |
|
|
Trouna |
- |
- |
|
|
Gabr Acun (Fezzan) |
- |
- |
|
|
Madagascar |
Salins de Diego Suarez |
- |
- |
|
Ankiembe saltworks |
P(3n) |
A. par |
|
|
Ifaty saltworks |
B |
A. fra |
|
|
Morocco |
Larache |
P |
A. par |
|
Moulaya estuary |
- |
- |
|
|
Qued Ammafatma |
- |
- |
|
|
Qued Chebeica |
- |
- |
|
|
Sebket Bon Areg |
- |
- |
|
|
Sebket Zima |
- |
- |
|
|
Mozambique |
Lagua Quissico |
P |
A. par |
|
Namibia |
Vineta Swakopmund |
P(2n, 4n) |
A. par |
|
Niger |
Teguidda In Tessoun |
- |
- |
|
Senegal |
Dakar |
- |
- |
|
Lake Kayar |
- |
- |
|
|
Lake Retba |
- |
- |
|
|
South Africa |
Couga Salt Flats |
- |
- |
|
Swartkops |
- |
- |
|
|
Tunisia |
Bekalta |
B |
A. sal |
|
Chott Ariana |
B |
A. sal |
|
|
Chott El Djerid |
- |
- |
|
|
Megrine |
B |
A. sal |
|
|
Sebket Kowezia |
- |
- |
|
|
Sebket mta Moknine |
B |
A. sal |
|
|
Sebket Sidi el Hani |
- |
- |
|
|
Sfax |
B |
A. sal |
|
|
Artemia sites in Australia and New Zealand |
|||
|
New Zealand |
Lake Grassmere |
B |
A. fra |
|
Queensland |
Bowen |
- |
- |
|
Port Alma |
B |
A. fra |
|
|
Rockhampton |
B |
A. fra |
|
|
South Australia |
Dry Creek, Adelaide |
P |
A. par |
|
West Australia |
Dampier |
- |
- |
|
Lake Mc Leod |
- |
- |
|
|
Port Hedland |
P |
A. par |
|
|
Rottnest Island |
P |
A. par |
|
|
Shark Bay |
P,B |
A.par,A.fra |
|
|
Artemia sites in North America |
|||
|
Canada |
Akerlund Lake |
B |
A. sp |
|
Alsask Lake |
B |
A. sp |
|
|
Aroma Lake |
B |
A. sp |
|
|
Berry Lake |
B |
A. sp |
|
|
Boat Lake |
B |
A. sp |
|
|
Burn Lake |
B |
A. sp |
|
|
Ceylon Lake |
B |
A. sp |
|
|
Chain Lake |
B |
A. sp |
|
|
Chaplin Lake |
B |
A. fra |
|
|
Churchill |
B |
A. sp |
|
|
Coral Lake |
B |
A. sp |
|
|
Drybore Lake |
B |
A. sp |
|
|
Enis Lake |
B |
A. sp |
|
|
Frederick Lake |
B |
A. sp |
|
|
Fusilier Lake |
B |
A. sp |
|
|
Grandora Lake |
B |
A. sp |
|
|
Gull Lake |
B |
A. sp |
|
|
Hatton Lake |
B |
A. sp |
|
|
Horizon Lake |
B |
A. sp |
|
|
Ingerbright Nath |
B |
A. sp |
|
|
Landis Lake |
B |
A. sp |
|
|
La Perouse |
B |
A. sp |
|
|
Little Manitou Lake |
B |
A. fra |
|
|
Lydden Lake |
B |
A. sp |
|
|
Mawer Lake |
B |
A. sp |
|
|
Meacham Lake |
B |
A. sp |
|
|
Muskiki Lake |
B |
A. sp |
|
|
Neola Lake |
B |
A. sp |
|
|
Oban Lake |
B |
A. sp |
|
|
Richmond Lake |
B |
A. sp |
|
|
Shoe Lake |
B |
A. sp |
|
|
Snakehole Lake |
B |
A. sp |
|
|
Sybouts Lake-East |
B |
A. sp |
|
|
Sybouts Lake-West |
B |
A. sp |
|
|
Verlo West |
B |
A. sp |
|
|
Vincent Lake |
B |
A. sp |
|
|
Wheatstone Lake |
B |
A. sp |
|
|
Whiteshore Lake |
B |
A. sp |
|
|
USA Arizona |
Kiatuthlana Red Pond |
B |
A. fra |
|
Kiatuthlana Green Pond |
B |
A. fra |
|
|
USA California |
Carpinteria Slough |
B |
A. sp |
|
Chula Vista |
B |
A. sp |
|
|
Mono Lake |
B |
A.f. mon |
|
|
Moss Landing, Monterey Bay |
B |
A. fra |
|
|
Owens Lake |
B |
A. sp |
|
|
San Diego |
B |
A. sp |
|
|
San Francisco Bay |
B |
A. fra |
|
|
San Pablo Bay |
B |
A. fra |
|
|
Vallejo West Pond |
B |
A. sp |
|
|
USA Hawaii |
Christmas Islands |
B |
A. sp |
|
Hanapepe |
B |
A. sp |
|
|
Laysan Atoll |
B |
A. fra |
|
|
USA Nebraska |
Alkali Lake |
B |
A. sp |
|
Ashenburger Lake |
B |
A. sp |
|
|
Antioch (Potash)Lake |
B |
A. fra |
|
|
Cook Lake |
B |
A. sp |
|
|
East Valley Lake |
B |
A. sp |
|
|
Grubny Lake |
B |
A. sp |
|
|
Homestead Lake |
B |
A. sp |
|
|
Jesse Lake |
B |
A. fra |
|
|
Johnson Lake |
B |
A. sp |
|
|
Lilly Lake |
B |
A. sp |
|
|
Reno Lake |
B |
A. sp |
|
|
Richardson Lake |
B |
A. fra |
|
|
Ryan Lake |
B |
A. sp |
|
|
Sheridan County Lake |
B |
A. sp |
|
|
Sturgeon Lake |
B |
A. fra |
|
|
USA Nevada |
Fallon Pond |
B |
A. fra |
|
USA North Dakota |
Miller Lake |
B |
A. sp |
|
Stink (Williams) Lake |
B |
A. sp |
|
|
USA New Mexico |
Laguna del Perro |
B |
A. sp |
|
Loving Salt Lake |
B |
A. sp |
|
|
Quemado |
B |
A. fra |
|
|
Zuni Salt Lake |
B |
A. fra |
|
|
USA Oregon |
Lake Abert |
B |
A. sp |
|
USA Texas |
Cedar Lake |
B |
A. fra |
|
McKenzies Playa |
B |
A. sp |
|
|
Mound Playa |
B |
A. sp |
|
|
Playa Thahoka |
B |
A. sp |
|
|
Raymondville |
B |
A. sp |
|
|
Rich Playa |
B |
A. sp |
|
|
Snow drop Playa |
B |
A. sp |
|
|
USA Utah |
Great Salt Lake |
B |
A. fra |
|
USA Washington |
Cameron Lake |
B |
A. fra |
|
Deposit Thirteen |
B |
A. fra |
|
|
Penley Lake |
B |
A. fra |
|
|
Hot (Bitter) Lake |
B |
A. fra |
|
|
Omak Plateau |
B |
A. sp |
|
|
Soap Lake |
B |
A. sp |
|
|
Artemia sites in Central America |
|||
|
Bahamas |
Great Inagua |
B |
A. sp |
|
Long Island |
B |
A. sp |
|
|
San Salvador |
B |
A. sp |
|
|
Brit. Virgin Islands |
Anegada |
B |
A. sp |
|
Carribean Islands |
Antigua |
B |
A. sp |
|
St. Kitts |
B |
A. sp |
|
|
St. Martin |
B |
A. sp |
|
|
South Caicos |
B |
A. sp |
|
|
Costa Rica |
Gulfo Nicova |
B |
A. sp |
|
Bahia salinas, Guanacaste |
B |
A. fra |
|
|
Dominican Republic |
Isla Cabra |
B |
A. sp |
|
Las Calderas |
B |
A. sp |
|
|
Monte Cristi |
B |
A. sp |
|
|
Puerto Alejandro |
B |
A. sp |
|
|
Punta Salinas |
B |
A. sp |
|
|
Haiti |
Grandes salines |
B |
A. fra |
|
Mexico Baja Calif. Norte |
San Quintin |
B |
A. fra |
|
Mexico Baja Calif. Sur |
Pichilingue, La Paz |
B |
A. fra |
|
Guerrero Negro |
B |
A. fra |
|
|
Isla del Carmen |
B |
A. fra |
|
|
Mexico Sonora |
Laguna de Yavaros |
B |
A. fra |
|
Mexico Coahuila |
Salinas 5 km SE Cuatrocienegas |
B |
A. sp |
|
Mexico Chiapas |
Laguna del Mar Muerto |
B |
A. sp |
|
La Joya |
B |
A. sp |
|
|
Buenavista |
B |
A. sp |
|
|
Los Palos |
B |
A. sp |
|
|
Solo Dios |
B |
A. sp |
|
|
Carretas |
B |
A. sp |
|
|
Pereyra |
B |
A. sp |
|
|
Chanchuto |
B |
A. sp |
|
|
Panzacola |
B |
A. sp |
|
|
Mexico Estado de Mexico |
Brine El Caracol, Sosa Texcoco |
B |
A. sp |
|
Mexico Oaxaca |
Ponds W. Salina Cruz |
B |
A. sp |
|
Mexico San Luis Potosi |
Las Salinas |
B |
A. sp |
|
Mexico Sinaloa |
Bahia de Ceuta |
B |
A. sp |
|
Mexico Yucatan |
San Crisanto |
B |
A. sp |
|
Celestun |
B |
A. sp |
|
|
Chuburna |
B |
A. sp |
|
|
Xtampu |
B |
A. sp |
|
|
Las Coloradas |
B |
A. sp |
|
|
Netherlands Antilles |
Aruba |
B |
A. sp |
|
Bonaire Duinmeer |
B |
A. fra |
|
|
Gotomeer |
B |
A. sp |
|
|
Pekelmeer |
B |
A. sp |
|
|
Martinus |
B |
A. sp |
|
|
Slagbaai |
B |
A. sp |
|
|
Curaçao Fuik |
B |
A. sp |
|
|
Rifwater |
B |
A. sp |
|
|
Nicaragua |
Salinas Grandes, Leon |
B |
A. fra |
|
Puerto Rico |
Bahia Salinas |
B |
A. fra |
|
Bogueron |
B |
A. sp |
|
|
Cabo Rojo |
B |
A. sp |
|
|
La Parguera |
B |
A. sp |
|
|
Ponce |
B |
A. sp |
|
|
Tallaboa salterns |
B |
A. fr |
|
|
Artemia sites in South America |
|||
|
Argentina |
Bahia Blanca |
B |
A. sp |
|
Buenos Aires |
B |
A. per |
|
|
Hidalgo |
B |
A. per |
|
|
Mar Chiquita |
B |
A. sp |
|
|
Bolivia |
Lake Canapa |
B |
A. sp |
|
Lake Chulluncani |
B |
A. sp |
|
|
Lake Hedonia |
B |
A. sp |
|
|
Lake Poopo |
B |
A. sp |
|
|
Brazil |
Aracati |
B |
A. sp |
|
Cabo Frio |
B |
A. fra |
|
|
Fortaleza |
B |
A. sp |
|
|
Icapui |
B |
A. sp |
|
|
Macau |
B |
A. fra |
|
|
Mundau |
B |
A. sp |
|
|
Chile |
Salar de Surire |
B |
A. sp |
|
Playa Yape (Iquique) |
B |
A. sp |
|
|
Salar de Pintados (I Region) |
B |
A. sp |
|
|
Salar de Llamara (II Region) |
B |
A. sp |
|
|
B |
A. fra |
||
|
Puerto Viejo (Copiapo) |
B |
A. sp |
|
|
La Pampilla (Coquimbo) |
B |
A. sp |
|
|
Palo Colorado (Los Vilos) |
B |
A. sp |
|
|
Salinas de Cahuil (Pichilemu) |
B |
A. sp |
|
|
Salinas de Constitución (VII Región) |
B |
A. sp |
|
|
Colombia |
Galerazamba |
B |
A. sp |
|
Manaure |
B |
A. sp |
|
|
Ecuador |
Galapagos (S.Salvador) |
B |
A. fra |
|
Pacoa |
B |
A. sp |
|
|
Salinas |
B |
A. sp |
|
|
Peru |
Caucato |
B |
A. sp |
|
Chicama |
B |
A. sp |
|
|
Chilca |
B |
A. sp |
|
|
Estuario de Virrila |
B |
A. sp |
|
|
Guadalupe |
B |
A. sp |
|
|
Pampa de Salinas |
B |
A. sp |
|
|
Pampa Playa Chica |
B |
A. sp |
|
|
Puerto Huarmey |
B |
A. sp |
|
|
Tumbes |
B |
A. sp |
|
|
Venezuela |
Boca Chica |
B |
A. sp |
|
Coya Sal |
B |
A. sp |
|
|
Coche |
B |
A. sp |
|
|
Coro Coastline |
B |
A. sp |
|
|
La Orchila |
B |
A. sp |
|
|
Las Aves |
B |
A. sp |
|
|
Los Roques |
B |
A. sp |
|
|
Port Araya |
B |
A. sp |
|
|
Tucacas |
B |
A. sp |
|
|
Artemia sites in Asia |
|||
|
P.R. China Liaoning |
Jinzhou |
P |
A. par |
|
Yingkou |
P(2, 4, 5n) |
A. par |
|
|
Dongjiagou |
P(2n) |
A. par |
|
|
Pulandian |
P(2n) |
A. par |
|
|
Lushun |
P(2, 4, 5n) |
A. par |
|
|
Fuzhouwan |
P |
A. par |
|
|
P.R. China Hebei |
Nanpu |
P(2n) |
A. par |
|
Luannan |
P |
A. par |
|
|
Daqinghe |
P |
A. par |
|
|
Huanghua |
P(2n) |
A. par |
|
|
Shangyi |
B |
A. sin |
|
|
Zhangbei |
B |
A. sin |
|
|
Kangbao |
B |
A. sin |
|
|
P.R. China Tianjin |
Hangu |
P(2n) |
A. par |
|
Tanggu |
P(2, 4, 5n) |
A. par |
|
|
P.R. China Shandong |
Chengkou |
P(2n) |
A. par |
|
Yangkou |
P(2n) |
A. par |
|
|
Dongfeng |
P(2, 5n) |
A. par |
|
|
Gaodao |
P |
A. par |
|
|
Xiaotan |
P |
A. par |
|
|
Nanwan |
P |
A. par |
|
|
Jimo |
P |
A. par |
|
|
P.R. China Jiangsu |
Xuyu |
P |
A. par |
|
Lianyungang |
P |
A. par |
|
|
P.R. China Zhejiang |
Zhanmao |
P |
A. par |
|
Shunmu |
P |
A. par |
|
|
Zhujiajian |
P |
A. par |
|
|
P.R. China Fujian |
Shanyao |
P |
A. par |
|
Xigang |
P |
A. par |
|
|
Huian |
P |
A. par |
|
|
P.R. China Guangdong |
|
P |
A. par |
|
P.R. China Hainan |
Dongfang |
P |
A. par |
|
Yinggehai |
P(2, 4, 5n) |
A. par |
|
|
P.R. China Xinjiang |
Aibi |
P(2, 4n) |
A. par |
|
Dabancheng |
P(2, 3, 4, 5n) |
A. par |
|
|
Balikun |
P(2,4n) |
A. par |
|
|
Aletai |
B |
A. sp |
|
|
P.R. China Tibet |
Yanjing |
B |
A. sp |
|
Shenzha |
B |
A. sp |
|
|
Bange |
- |
- |
|
|
Gaize |
- |
- |
|
|
Geji |
- |
- |
|
|
Zhangchaka |
- |
- |
|
|
Wumacuo |
- |
- |
|
|
Jibuchaka |
- |
- |
|
|
Dongcuo |
- |
- |
|
|
P.R. China Qinghai |
Gahai |
P(2n) |
A. par |
|
Xiaocaidan |
P |
A. par |
|
|
Dacaidan |
P |
A. par |
|
|
Suban |
P |
A. par |
|
|
Keke |
P(4n) |
A. par |
|
|
Chaka |
P |
A. par |
|
|
Tuosu |
P |
A. par |
|
|
P.R. China Gansu |
Gaotai |
B |
A. sp |
|
P.R. China Inner Mongolia |
Haolebaoji(Y) |
B |
A. sin |
|
(Y = Yimeng Area) |
Haotongyin(Y) |
B |
A. sin |
|
(X = Ximeng Area) |
Taigemiao(Y) |
B |
A. sin |
|
Ejinor(X) |
B |
A. sin |
|
|
Beidachi(Y) |
B |
A. sin |
|
|
Jilantai |
B |
A. sin |
|
|
Wuqiangi |
B |
A. sin |
|
|
Shanggendalai(X) |
B |
A. sin |
|
|
Dagenor(X) |
B |
A. sin |
|
|
Bayannor(X) |
B |
A. sin |
|
|
Zhunsaihannor |
B |
A. sin |
|
|
Erendabusen |
B |
A. sin |
|
|
Chagannor(X) |
B |
A. sin |
|
|
Huhetaolergai(Y) |
B |
A. sin |
|
|
Hangjinqi |
B |
A. sin |
|
|
P.R. China Ningxia |
|
- |
- |
|
P.R. China Shaanxi |
Dingbian |
- |
- |
|
P.R. China Shanxi |
Yuncheng |
B |
A. sin |
|
India Rajasthan |
Didwana |
- |
- |
|
Sambhar Lake |
- |
- |
|
|
India Gujarat |
Gulf of Kutch |
P |
A. par |
|
Balamba salterns |
P |
A. par |
|
|
Mithapur |
P |
A. par |
|
|
Jamnagar |
- |
- |
|
|
India Bombay |
Vadala |
- |
- |
|
Bhayander |
P |
A. par |
|
|
Bahinder |
- |
- |
|
|
India Madras |
Kelambakkam |
- |
- |
|
Vedaranyam |
- |
- |
|
|
India Tuticorin |
Veppalodai |
- |
- |
|
Pattanamaruthur |
- |
- |
|
|
Spic Nagar |
- |
- |
|
|
Thirespuram |
- |
- |
|
|
Karsewar Island |
- |
- |
|
|
Saltwater springs |
P |
A. par |
|
|
Harbour |
- |
- |
|
|
India Kanyakumari |
Thamaraikulam |
P |
A. par |
|
Iraq |
Abu-Graib, Baghdad |
P |
A. par |
|
Basra |
- |
- |
|
|
Dayala |
- |
- |
|
|
Mahmoodia |
- |
- |
|
|
Iran |
Urmia Lake |
B |
A. urm |
|
Schor-Gol |
- |
- |
|
|
Shurabil |
- |
- |
|
|
Athlit |
- |
- |
|
|
Israel |
Eilat North |
P |
A. par |
|
Eilat South |
- |
- |
|
|
Japan |
Chang Dao |
- |
- |
|
Tamano |
- |
- |
|
|
Yamaguchi |
P |
A. par |
|
|
Kuwait |
|
- |
- |
|
Korea |
Pusan |
- |
- |
|
Pakistan |
Karachi saltworks |
P |
A. par |
|
Sri Lanka |
Bundala |
- |
- |
|
Hambantota |
- |
- |
|
|
Palavi |
- |
- |
|
|
Putallam |
P |
A. par |
|
|
Taiwan |
Peinan Salina |
- |
- |
|
Beimen |
B |
A. sp |
|
|
Turkey |
Balikesir, Aivalik |
- |
- |
|
Camalti, Izmir |
P |
A. par |
|
|
Tuz Golii |
- |
- |
|
|
Ankara Salt Lake |
- |
- |
|
|
Konya Karapinar-Meke Salt Lake |
- |
- |
|
|
Imbros |
- |
- |
|
|
Artemia sites in Europe |
|||
|
Bulgaria |
Burgas |
P |
A. par |
|
Pomorye |
- |
- |
|
|
Croatia |
Secovlje, Portoroz |
P(4n) |
A. par |
|
Strunjan |
P |
A. par |
|
|
Ulcinj |
P |
A. par |
|
|
Cyprus |
Akrotiri Lake |
- |
- |
|
Larnaca Lake |
B |
A. sal |
|
|
France |
Aigues Mortes |
P |
- |
|
Carnac-Trinité sur Mer |
- |
- |
|
|
Guérande-le Croisic |
P |
A. par |
|
|
La Palme |
- |
- |
|
|
Lavalduc |
P |
A. par |
|
|
Mesquer-Assérac |
- |
- |
|
|
Porte La Nouvelle |
- |
- |
|
|
Salin de Berre |
P |
A. par |
|
|
Salin de Fos |
- |
- |
|
|
Salin de Giraud |
P |
A. par |
|
|
Salins d’Hyères |
- |
- |
|
|
Salin des Pesquiers |
- |
- |
|
|
Sète |
P |
A. par |
|
|
Greece |
Citros, Pieria |
P(4n) |
A. par |
|
Megalon Embolon, Thessaloniki |
P(4n) |
A. par |
|
|
Kalloni, Lesbos |
P(4n) |
A. par |
|
|
Polychnitos, Lesbos |
P(4n) |
A. par |
|
|
Mesolongi |
P |
A. par |
|
|
Milos Island |
P |
A. par |
|
|
Italy |
Quartu or salina di Poetto, Cagliary |
B |
A. sal |
|
Carloforte, Sardinia |
B |
A. sal |
|
|
Cervia, Ravenna |
P(4n) |
A. par |
|
|
Commachio, Ferrara |
P(4n) |
A. par |
|
|
Margherita di Savoia, Foggia |
P(2,4n) |
A. par |
|
|
Sant’ Antioco, Sardinia |
B |
A. sal |
|
|
Santa Gilla, Sardinia |
P(2n) |
A. par |
|
|
Siracuse, Sicily |
|
|
|
|
Tarquinia, Viterbo |
B |
A. sal |
|
|
Trapani, Sicily |
B |
A. sal |
|
|
Portugal |
Alcochete |
P |
A. par |
|
Tejo estuary |
- |
- |
|
|
Sado estuary |
- |
- |
|
|
Ria de Aveiro |
- |
- |
|
|
Ria de Farc |
- |
- |
|
|
Romania |
Lake Techirghiol |
P |
A. par |
|
Lacul Sârat Brâila |
P |
A. par |
|
|
Movila Miresii |
- |
- |
|
|
Romania Slâric Prahova |
Baia Baciului |
P |
A. par |
|
Romania Slâric Prahova |
Baia Neagrâ, SP |
P |
A. par |
|
Baia Verde I, SP |
P |
A. par |
|
|
Baia Verde II, SP |
P |
A. par |
|
|
Baia Verde III, SP |
P |
A. par |
|
|
Baia Rosie, SP |
P |
A. par |
|
|
Romania Telega |
Telega Bâi |
P |
A. par |
|
Telega II |
P |
A. par |
|
|
Telega III |
P |
A. par |
|
|
Ocra Sibiului |
P? |
|
|
|
Sovata |
P? |
|
|
|
Spain Alava |
Añana |
P(4n) |
A. par |
|
Spain Albacete |
Petrola |
P(4n) |
A. par |
|
Pinilla |
P(4n) |
A. par |
|
|
Spain Alicante |
Bonmati, S.Pola |
B, P(2, 4n) |
mixed |
|
Bras de Port, S.Pola |
B |
m |
|
|
Calpe |
P(2n) |
A. par |
|
|
La Mata |
P(2n) |
A. par |
|
|
Molina del Segura |
B |
|
|
|
Salinera Espanola, S. Pola |
B |
|
|
|
Villena |
B |
|
|
|
Spain Burgos |
Poza de la Sal |
B |
A. sp |
|
Spain Cadiz |
Sanlucar de Barrameda |
P |
A. par |
|
Dos hermanos |
B, P(2n) |
mixed |
|
|
San Eugenio |
B, P(2n) |
mixed |
|
|
San Felix |
B |
A. sal |
|
|
San Fernando |
B |
A. sal |
|
|
San Juan |
B, P |
mixed |
|
|
San Pablo |
B, P |
mixed |
|
|
Santa Leocadia |
B, P |
mixed |
|
|
Barbanera |
B |
A. sal |
|
|
Spain Canary islands |
Janubio, Lanzarote |
P(2n) |
A. par |
|
Spain Cordoba |
Encarnacion |
P(4n) |
A. par |
|
Puente Montilla |
P(4n) |
A. par |
|
|
Spain Formentera |
Salinera Espanola, |
B |
A. sal |
|
Spain Guadalajara |
Armalla |
P(4n) |
A. par |
|
Imon |
P(4n) |
A. par |
|
|
Olmeda |
P(4n) |
A. par |
|
|
Rienda |
P(4n) |
A. par |
|
|
Spain Huelva |
Ayamonte |
P(2n) |
A. par |
|
Lepe |
P(2n) |
A. par |
|
|
Isla Cristina |
P(2n) |
A. par |
|
|
San Juan del Puerto |
B |
A. sal |
|
|
Spain Huesca |
Rolda |
P |
A. par |
|
Peralta de la Sal |
P |
A. par |
|
|
Spain Ibiza island |
Salinera Espanola |
|
|
|
Spain Jaen |
San Carlos |
|
|
|
Don Benito |
|
|
|
|
Spain Malaga |
Fuente de Piedra |
|
|
|
Spain Mallorca |
Campos del Puerto |
B |
A. sal |
|
Spain Murcia |
San Pedro del Pinatar |
B |
A. sal |
|
Jumilla |
B |
A. sal |
|
|
sal. Punta Galera |
B |
A. sal |
|
|
sal. Catalana |
B |
A. sal |
|
|
Spain Soria |
Medinaceli |
P(4n) |
A. par |
|
Spain Tarragona |
Delta del Ebro |
P(4n) |
A. par |
|
Spain Teruel |
Arcos de las Salinas |
P(4n) |
A. par |
|
Spain Zaragoza |
Chiprana |
P(4n) |
A. par |
|
Bujaralo |
P(4n) |
A. par |
|
|
Artemia sites in former USSR |
|||
|
Russia |
Bolshoe Otar Mojnaksho |
|
|
|
Bolshoe Yarovoe |
P |
|
|
|
Maloe Yarovoe/Mojnakshoe/Dscharylgach |
|
|
|
|
Ghenicheskoe |
|
|
|
|
Karachi Lake |
|
|
|
|
Kujalnic liman |
P |
A. par |
|
|
Mangyshlak peninsula |
|
|
|
|
Schekulduk |
P |
|
|
|
Tanatar |
B |
A. par |
|
|
Kulundinskoe |
P |
|
|
|
Soljonoe |
P |
|
|
|
Mirabilit |
P |
|
|
|
Bolshoe Shklo |
P |
A. par |
|
|
Kurichye |
P |
|
|
|
Buazonsor |
P |
|
|
|
Mormishanskoe A |
P |
|
|
|
Mormishanskoe B |
P |
|
|
|
Kutchukskoe |
P |
|
|
|
|
P |
|
|
|
Kazakhstan |
Maraldi |
P |
|
|
Sejten |
P |
|
|
|
Turkmenistan |
|
P |
|
|
Ukraina |
Popovskoe (=Ojburgskoe) |
P |
|
|
Tchokrakskoe |
B |
|
|
|
Tobetchikskoe |
P |
|
|
|
Shtormovoe |
B |
|
|
|
Sakskoe |
|
|
|
|
Sasyk |
|
|
|
P = parthenogenetic strain
B = bisexual strain
A. par = Artemia parthenogenetica
A. sal = Artemia salina (= A. tunisiana)
A. fr = Artemia franciscana
A. fr. mon = Artemia franciscana monica
A. per = Artemia persimilis
A. urm = Artemia urmiana
A. sin = Artemia sinica
A. sp = Artemia species (unknown)
The genus Artemia is a complex of sibling species and superspecies, defined by the criterion of reproductive isolation. Early taxonomists assigned species names to populations with different morphologies, collected at different temperatures and salinities. Later on, the profusion of names was abandoned and all brine shrimp was referred to as Artemia salina Linnaeus 1758. Some authors continue this practice today. Generally, different names are assigned to reproductively isolated populations or clusters of populations:
· A. salina Linnaeus 1758: Lymington, England (now extinct), Mediterranean area;· A. tunisiana Bowen and Sterling 1978 synonym of A. salina;
· A. parthenogenetica Barigozzi 1974, Bowen and Sterling 1978: Europe, Africa, Asia, Australia;
· A. urmiana Gunther 1900: Iran;
· A. sinica Yaneng 1989: Central and Eastern Asia;
· A. persimilis Piccinelli and Prosdocimi 1968: Argentina;
· A. franciscana superspecies: Americas, Carribean and Pacific islands, including populations reproductively isolated in nature like A.(franciscana) franciscana Kellogg 1906 and A.(franciscana) monica Verrill 1869 (Mono Lake, California);
· Artemia sp. Pilla and Beardmore 1994: Kazakhstan.
The coexistence of two species in the same saline habitat is possible: mixtures of parthenogenetic and zygogenetic populations have been reported in Mediterranean salterns. In addition, commercial aquaculture ventures have seeded salterns with imported cysts on many occasions; A. Franciscana being introduced throughout Asia, Australia, and South America over the last 20 years. Because new populations are constantly being characterised, scientists are urged to use the denomination Artemia sp. unless they have sufficient biochemical, cytogenetic or morphological evidence to identify the species name.
The worldwide distribution of brine shrimp in a variety of isolated habitats, each one characterised by its own ecological conditions, has furthermore resulted in the existence of numerous geographical strains, or genetically different populations within the same sibling species; in particular the parthenogenetic Artemia with its di-, tri-, tetra- and pentaploid populations display a wide genotypic variation. Among these strains a high degree of genetic variability as well as a unique diversity in various quantitative characteristics have been observed. Some of these characteristics (i.e. the nutritional value of freshly-hatched nauplii) are phenotypical, and change from year to year or season to season. Others, however (i.e. cyst diameter, growth rate, resistance to high temperature) are strain specific and remain relatively constant, (i.e. they have become genotypical as a result of long-term adaptations of the strain to the local conditions; see chapter 4.1.2.4).
INTRODUCTION
While the nutritional value can be manipulated, other qualities favourable for aquaculture use can be obtained by selection of strains and/or their cross breeds. Although until recently over 90% of all marketed cysts originated from the Great Salt Lake, Artemia cysts are commercially available from various production sources in America, Asia, Australia and Europe. A knowledge of the characteristics (both genotypic and phenotypic) of a particular batch of cysts can greatly increase the effectiveness of its usage in a fish or shrimp hatchery.
SIZE AND ENERGY CONTENT
The nutritional effectiveness of a food organism is primarily determined by its ingestibility and, as a consequence, by its size and form (see further: chapter 4.3.3.). Data on biometrics of nauplii from various Artemia strains are given in Table 4.1.2.
Table 4.1.2. Size, individual dry weight and energy content of Artemia instar I nauplii from different cyst sources hatched in standard conditions (35g.l-1, 25°C)
|
cyst source |
length |
dry weight |
energy content |
|
San Francisco Bay, CA-USA |
428 |
1.63 |
366 |
|
Macau, Brazil |
447 |
1.74 |
392 |
|
Great Salt Lake, UT-USA |
486 |
2.42 |
541 |
|
Shark Bay, Australia |
458 |
2.47 |
576 |
|
Chaplin Lake, Canada |
475 |
2.04 |
448 |
|
Tanggu, Bohai Bay, PR China |
515 |
3.09 |
681 |
|
Aibi Lake, PR China |
515 |
4.55 |
- |
|
Yuncheng, PR China |
460 |
2.03 |
- |
|
Lake Urmiah, Iran |
497 |
- |
- |
Many strains can be differentiated on the basis of their biometrical characteristics. In spite of small variations between batches of the same strain, possibly caused by environmental and/or processing factors, generally the cyst diameter of different production batches of the same strain remains rather constant. Other biometrical characteristics such as cyst volume, cyst dry weight, instar I-naupliar length, individual naupliar weight and naupliar volume, energy content etc., show a high correlation with the cyst diameter. As a consequence, biometrical parameters, in particular cyst diameter, are good tools to characterize Artemia strains, and to help to define the origin of unknown or even mixed cyst samples.
Some general correlations can also be made between sibling species and size: parthenogenetic Artemia produce large cysts, A. tunisiana large cysts with a thick chorion, A. franciscana and A. persimilis small or intermediate cysts with a thin chorion.
HATCHING QUALITY
Comparative studies of the hatching behaviour of cysts of different origin show a considerable variation in hatching percentage, rate and efficiency. However, none of these parameters is strain specific as they are influenced by a wide array of factors like harvesting, processing, storage and hatching techniques, as well as production conditions affecting the parental generation. For optimal use of Artemia in aquaculture the hatching characteristics of any batch of cysts being used should be known. More information in this respect is given in chapter 4.2.5.2.
GROWTH RATE OF NAUPLII
Standard culture tests with brine shrimp from different geographical origin show important differences in growth rate even within the same sibling species, but not among batches of the same strain (Table 4.1.3.). Although in the field the population growth of Artemia (i.e. after inoculation) is determined by lots of factors, selection of a strain with a high potential growth rate will have a positive impact on maximal production output.
TEMPERATURE AND SALINITY TOLERANCE
Both temperature and salinity significantly affect survival and growth, the effect of temperature being more pronounced. A broad range of temperatures and salinities meets the requirements for >90% survival. Strains from thalassohaline biotopes share a common temperature area of preference in the range 20-25°C where mortalities are <10%. Interaction between temperature and salinity is limited; substantial differences in tolerance have been recorded at low salinities (around 5 g.l-1) and high temperatures (30-34°C). At elevated temperatures the survival of the GSL strain is significantly higher than for other strains.
Table 4.1.3. Growth of Artemia nauplii from different cyst sources under standard culture conditions (7 days, darkness, 10 instar I nauplii/25 ml, 35g.l-1, 25°C, fed with Dunaliella viridis)
|
cyst source |
growth expressed as % recorded for reference
strain |
|
San Francisco Bay, CA-USA, batch 1 |
96 |
|
San Francisco Bay, CA-USA, batch 2 |
96 |
|
Macau, Brazil, batch 1 |
98 |
|
Macau, Brazil, batch 2 |
103 |
|
Great Salt Lake, UT-USA, batch 1 |
125 |
|
Great Salt Lake, UT-USA, batch 2 |
127 |
|
Shark Bay, Australia |
95 |
|
Chaplin Lake, Canada |
130 |
|
Tanggu, Bohai Bay, PR China |
110 |
|
Aibi Lake, PR China |
105 |
|
Yuncheng, PR China |
109 |
LIFE HISTORY TRAITS AND REPRODUCTIVE CAPACITY
Life history and reproductive characteristics of Artemia strains are important factors when an introduction of brine shrimp in a new habitat is considered, especially when competition with a local strain is to be expected. These competitive abilities are related to factors like the length of reproductive, pre- and post-reproductive period, total lifespan, number of offspring per brood, broods per female, time in-between broods etc. In general New World (bisexual) populations have a very large number of offspring per brood, a large number of offspring/day/female and a fast development time to sexual maturity, which favours this group to Old World bisexual and parthenogenetic Artemia.
Age at first reproduction is a key factor determining the population growth rate, and the rate of colonisation of new environments with limited nutrient resources. Consequently, if environmental preferences and nutritional factors don’t interfere, New World bisexuals generally outcompete parthenogenetic strains, the latter in their turn predominating over Old World bisexuals. Inoculation experiments in natural habitats therefore require prior screening of candidate strains and of eventual local populations, as well as the study of prevailing environmental conditions. Uncontrolled introduction of Artemia may thus lead to a decrease of natural variability. Therefore, before inoculation of Artemia in a habitat with a local strain is undertaken, sufficient cyst material of the local population must be collected and stored in order to safeguard its gene-pool.
NUTRITIONAL VALUE
In the late seventies, when many fish and shrimp hatcheries started to go commercial, switching from one source of Artemia to another provoked unexpected problems (Table 4.1.4.). Very significant differences in production yields were even obtained with distinct Artemia batches of the same geographical origin. Especially the pattern of total lipids and fatty acid composition, as well as the metabolization of fatty acids in the Artemia, seemed to differ widely from strain to strain, and even from batch to batch, as a consequence of the fluctuations in biochemical composition of the primary producers (mainly unicellular algae) available to the adult population. Cyst products from inland resources are more constant in composition, be it however at suboptimal low levels. Appropriate techniques have thus been developed to improve the lipid profile of deficient Artemia strains, taking advantage of the indiscriminate filter-feeding behaviour of Artemia.
Applying simple methods lipophilic compounds can be easily incorporated into the Artemia before being offered as a prey (see chapter 4.4.).
A number of other compounds also appear to be variable from strain to strain: nutritional components such as total amount of free amino acids, pigments (canthaxanthin), vitamin C, minerals and trace elements, as well as contamination with chemicals such as pesticides and heavy metals. In most cases these variations are not strain specific, but just correspond to different production conditions. Despite this, their effects on larviculture success are usually far less significant.
Table 4.1.4. Growth and survival of fish and crustacean larvae fed with Artemia nauplii from different cyst sources
|
cyst source |
species tested |
survival |
growth |
|
San Francisco Bay, CA-USA, various batches |
Mysidopsis bahia |
+ |
+ |
|
Palaemon serratus |
+ |
+ |
|
|
Cyprinus caprio |
+ |
+ |
|
|
Solea solea |
+ |
+ |
|
|
Scophthalmus maximus |
+/±/- |
+/±/- |
|
|
Dicentrarchus labrax |
+ |
+ |
|
|
Macau, Brazil, various batches |
Mysidopsis bahia |
+ |
+ |
|
Cancer irroratus |
+ |
+ |
|
|
Menidia menidia |
+ |
± |
|
|
Cyprinus carpio |
+ |
± |
|
|
Scophthalmus maximus |
+ |
+ |
|
|
Pseudopleuronectes americanus |
+ |
± |
|
|
Great Salt Lake, UT-USA, various batches |
Mysidopsis bahia |
+/- |
+/- |
|
Cancer irroratus |
- |
- |
|
|
Palaemon serratus |
- |
- |
|
|
Penaeus vannamei |
- |
- |
|
|
Menidia menidia |
+ |
+ |
|
|
Cyprinus carpio |
+ |
+ |
|
|
Dicentrarchus labrax |
- |
- |
|
|
Shark Bay, Australia |
Mysidopsis bahia |
+ |
+ |
|
Cancer irroratus |
+ |
+ |
|
|
Menidia menidia |
+ |
+ |
|
|
Cyprinus carpio |
+ |
+ |
|
|
Pseudopleuronectes americanus |
+ |
+ |
|
|
Chaplin Lake, Canada |
Mysidopsis bahia |
± |
± |
|
Menidia menidia |
± |
- |
|
|
Cyprinus carpio |
+ |
- |
|
|
Pseudopleuronectes americanus |
+ |
± |
|
|
Pagrus major |
+ |
± |
|
|
Tianjin, Bohai Bay, PR China |
Mysidopsis bahia |
+ |
+ |
|
Menidia menidia |
+ |
+ |
|
|
Cyprinus carpio |
+ |
+ |
|
|
Pseudopleuronectes americanus |
+ |
+ |
Beardmore, J.A., Pilla, E.J., and Thomas, K.M. 1994. Genetic variation in Artemia: speciation, reproductive mode and potential for exploitation. In: Book of abstracts of contributions presented at the International Conference “Improvement of the commercial production of marine aquaculture species”. G. Gajardo and P. Coutteau (Eds), Puerto Montt, Chile, 5-9 December 1994.
Browne, R.A. and Bowen, S.T. 1991. Taxonomy and population genetics of Artemia. In: Artemia Biology. Browne, R.A., P. Sorgeloos and C.N.A. Trotman (Eds), CRC Press, Boca Raton Ann Arbor Boston, USA, pp 221-235.
Browne, R.A. and Halanych, K.M. 1989. Competition between sexual and parthenogenetic Artemia: a re-evaluation (Branchiopoda, Anostraca). Crustaceana, 57(1): 57-71.
Browne, R.A., Li, M., Wanigasekera, G., Simonek, S., Brownlee, D., Eiband, G. and Cowan, J. 1993. Ecological, physiological and genetic divergence of sexual and asexual (diploid and polyploid) brine shrimp (Artemia). Trends in Ecology: 1-14.
Browne, R.A., Sallee, S.E., Grosch, D.S., Segreti, W.O. and Purser, S.M. 1984. Partitioning genetic and environmental components of reproduction and lifespan in Artemia. Ecology, 65(3): 949-960.
Lavens, P. and Sorgeloos, P. 1987. The cryptobiotic state of Artemia cysts, its diapause deactivation and hatching, a review. In: Artemia Research and its Applications, Vol. 3. Sorgeloos, P., D.A. Bengtson, W. Decleir and E. Jaspers (Eds), Universa Press, Wetteren, Belgium, pp 27-63.
Léger, Ph., Bengtson, D.A., Sorgeloos, P., Simpson, K.L. and Beck, A.D. 1987. The nutritional value of Artemia, a review. In: Artemia Research and its Applications, Vol. 3. Sorgeloos, P., D.A. Bengtson, W. Decleir and E. Jaspers (Eds), Universa Press, Wetteren, Belgium, pp 357-372.
Persoone, G. and Sorgeloos, P. 1980. General aspects of the ecology and biogeography of Artemia. In: The brine shrimp Artemia. Vol. 3. Ecology, culturing, use in aquaculture. Persoone, G., P. Sorgeloos, O. Roels and E. Jaspers (Eds), Universa Press, Wetteren, Belgium, pp 3-24.
Pilla, E.J.S and Beardmore, J.A. 1994. Genetic and morphometric differentiation in Old World bisexual species of the brine shrimp (Artemia). Heredity, 72: 47-56.
Tackaert, W. and Sorgeloos, P. 1991. Semi-intensive culturing in fertilized ponds. In: Artemia Biology. Browne, R.A, P. Sorgeloos and C.N.A. Trotman (Eds), CRC Press, Boca Raton Ann Arbor Boston, USA, pp 287-315.
Triantaphyllidis, G.V., Zhang, B., Zhu, L. and Sorgeloos, P. 1994. International Study on Artemia. L. Review of the literature on Artemia from salt lakes in the People’s Republic of China. International Journal of Salt Lake Research, 3:1-12.
Vanhaecke, P., Tackaert, W. and Sorgeloos, P. 1987. The biogeography of Artemia: an updated review. In: Artemia research and its applications. Vol. 1. Morphology, genetics, strain characterisation, toxicology. Sorgeloos, P., D.A. Bengtson, W. Decleir and E. Jaspers (Eds), Universa Press, Wetteren, Belgium, pp 129-155.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}