![]()
![]()
![]()
Role in human metabolic processes
Overview of significant scientific information
Definition of population at risk
Dietary sources of vitamin C and limitations to vitamin C
Information used to derive dietary requirement of vitamin C
Future research
References
Vitamin C (chemical names: ascorbic acid and ascorbate) is a six-carbon lactone which is synthesised from glucose by many animals. Vitamin C is synthesised in the liver in some mammals and in the kidney in birds and reptiles. However, several species - including humans, non-human primates, guinea pigs, Indian fruit bats, and Nepalese red-vented bulbuls - are unable to synthesise vitamin C. When there is insufficient vitamin C in the diet, humans suffer from the potentially lethal deficiency disease scurvy (1). Humans and primates lack the terminal enzyme in the biosynthetic pathway of ascorbic acid, l-gulonolactone oxidase, because the gene encoding for the enzyme has undergone substantial mutation so that no protein is produced (2).
Background biochemistry
Vitamin C is an electron donor (reducing agent or antioxidant), and probably all of its biochemical and molecular functions can be accounted for by this function. The potentially protective role of vitamin C as an antioxidant is discussed in the antioxidants chapter of this report.
Enzymatic functions
Vitamin C acts as an electron donor for 11 enzymes (3, 4). Three of those enzymes are found in fungi but not in humans or other mammals (5, 6). They are involved in reutilisation pathways for pyrimidines and the deoxyribose moiety of deoxynucleosides. Of the 8 remaining human enzymes, three participate in collagen hydroxylation (7-9) and two in carnitine biosynthesis (10, 11); of the three enzymes which participate in collagen hydroxylation, one is necessary for biosynthesis of the catecholamine norepinephrine (12, 13), one is necessary for amidation of peptide hormones (14, 15), and one is involved in tyrosine metabolism (4, 16).
Ascorbate interacts with enzymes having either monooxygenase or dioxygenase activity. The monooxygenases dopamine b-monooxygenase and peptidyl-glycine a-monooxygenase incorporate a single oxygen atom into a substrate, either a dopamine or a glycine-terminating peptide. The remaining enzymes are dioxygenases which incorporate two oxygen atoms in two different ways. The enzyme 4-hydroxyphenylpyruvate dioxygenase incorporates two oxygen atoms into one product. The other dioxygenase incorporates one oxygen atom into succinate and one into the enzyme-specific substrate.
Miscellaneous functions
The concentrations of vitamin C in gastric juice were several fold higher (median, 249 mmol/l; range, 43-909 mmol/l) than those found in the plasma of the same normal subjects (39 mmol/l, 14-101 mmol/l) (17). Gastric juice vitamin C may prevent the formation of N-nitroso compounds, which are potentially mutagenic (18). High intakes of vitamin C correlate with reduced gastric cancer risk (19), but a cause-and-effect relationship has not been established. Vitamin C protects low-density lipoproteins ex vivo against oxidation and may function similarly in the blood (20; see Chapter 17).
A common feature of vitamin C deficiency is anaemia. The antioxidant properties of vitamin C may stabilise folate in food and in plasma, and increased excretion of oxidized folate derivatives in human scurvy was reported (21). Vitamin C promotes absorption of soluble non-haem iron possibly by chelation or simply by maintaining the iron in the reduced (ferrous, Fe2+) form (22, 23). The effect can be achieved with the amounts of vitamin C obtained in foods. However, the amount of dietary vitamin C required to increase iron absorption ranges from 25 mg upwards and depends largely on the amount of inhibitors, such as phytates and polyphenols, present in the meal (24). (See Chapter 13 on iron for further discussion.)
From the 15th century, scurvy was dreaded by seamen and explorers forced to subsist for months on diets of dried beef and biscuits. Scurvy was described by the Crusaders, during the sieges of numerous European cities, and as a result of the famine in 19th century Ireland. Three important manifestations of scurvy - gingival changes, pain in the extremities, and haemorrhagic manifestations - preceded oedema, ulcerations, and ultimately death. Skeletal and vascular lesions in scurvy probably arise from a failure of osteoid formation. In infantile scurvy the changes are mainly at the sites of most active bone growth; characteristic signs are a pseudoparalysis of the limbs caused by extreme pain on movement and caused by haemorrhages under the periosteum, as well as swelling and haemorrhages in areas of the gums surrounding erupting teeth (25). In adults one of the early, principle adverse effects of the collagen-related pathology may be impaired wound healing (26). Vitamin C deficiency can be detected from early signs of clinical deficiency, such as the follicular hyperkeratosis, petechial haemorrhages, swollen or bleeding gums, and joint pain, or from the very low concentrations of ascorbate in plasma, blood, or leukocytes. The Sheffield studies (26, 27) and later studies in Iowa (28, 29) were the first major attempts made to quantify vitamin C requirements. The studies indicated that the amount of vitamin C required to prevent or cure early signs of deficiency was between 6.5 and 10 mg/day. This range represents the lowest physiologic requirement. The Iowa studies (28, 29) and Kallner et al (30) established that at tissue saturation, whole body vitamin C content is approximately 20 mg/kg, or 1500 mg, and that during depletion vitamin C is lost at 3 percent of whole body content per day.
Clinical signs of scurvy appear in men at intakes lower than 10 mg/day (27) or when the whole body content falls below 300 mg (28). Such intakes are associated with plasma ascorbate concentrations below 11 mmol/l or leukocyte levels less than 2 nmol/108 cells. However, the plasma concentrations fall to around 11 mmol/l when dietary vitamin C is between 10 and 20 mg/day. At intakes greater than 25-35 mg/day, plasma concentrations start to rise steeply, indicating a greater availability of vitamin C for metabolic needs. In general, plasma ascorbate closely reflects the dietary intake and ranges between 20 and 80 mmol/l. Note that during infection or physical trauma, an increase in the number of circulating leukocytes occurs and these take up vitamin C from the plasma (31, 32). Therefore, both plasma and leukocyte levels may not be very precise indicators of body content or status at such times. However, leukocyte ascorbate remains a better indicator of vitamin C status than plasma ascorbate most of the time and only in the period immediately after the onset of an infection are both values unreliable.
Intestinal absorption of vitamin C is by an active, sodium-dependent, energy-requiring, carrier-mediated transport mechanism (33) and as intakes increase, the tissues progressively become more saturated. The physiologically efficient, renal-tubular reabsorption mechanism retains vitamin C in the tissues up to a whole body content of ascorbate of about 20 mg/kg body weight (30). However, under steady state conditions, as intakes rise from around 100 mg/day there is an increase in urinary output in so that at 1000 mg/day almost all absorbed vitamin C is excreted (34, 35).
The populations at risk of vitamin C deficiency are those for whom the fruit and vegetable supply is minimal. Epidemics of scurvy are associated with famine and war, when people are forced to become refugees and food supply is small and irregular. Persons in whom the total body vitamin C content is saturated can subsist without vitamin C for approximately 2 months before the appearance of clinical signs, and as little as 6.5-10 mg/day vitamin C will prevent the appearance of scurvy. In general, vitamin C status will reflect the regularity of fruit and vegetable consumption but also socio-economic conditions, because intake is determined not just by availability, but by cultural preferences and cost.
In Europe and the United States an adequate intake of vitamin C is indicated by the results of various national surveys (36-38). In the United Kingdom and Germany, the mean dietary intakes of vitamin C in adult men and women were 87 and 76 (37) and 75 and 72 mg/day (36), respectively. In addition, a recent survey of elderly men and women in the United Kingdom reported vitamin C intakes of 72 (SD 61) and 68 (SD 60) mg/day, respectively (39). In the United States, in the National Health and Nutrition Examination Survey (38), the median consumption of vitamin C from foods during the years 1988-91 was 73 and 84 mg/day in men and women, respectively. In all these studies there was a wide variation in vitamin C intake and 25-30 percent of the US population consumed less than 2.5 servings of fruit and vegetables daily. Likewise a survey of Latin American children in the United States suggested that less than 15 percent consumed the recommended intake of fruits and vegetables (40). It is not possible to relate servings of fruits and vegetables to an exact amount of vitamin C, but the World Health Organization (WHO) dietary goal of 400 g (41) aimed to provide sufficient vitamin C to meet the 1970 Food and Agriculture Organization of the United Nations (FAO)/WHO guidelines - that is, approximately 20-30 mg/day - and lower the risk of chronic disease. The WHO goal has been roughly translated into the recommendation of five portions per day (42).
Reports from India show that the available supply of vitamin C is 43 mg/capita/day, and in the different states of India it ranges from 27 to 66 mg/day. In one study, low-income children consumed as little as 8.2 mg/day of vitamin C in contrast to a well-to-do group of children where the intake was 35.4 mg/day (43). Other studies done in developing countries found plasma vitamin C concentrations lower than those reported for developed countries, for example, 20-27 mmol/l for apparently healthy adolescent boys and girls in China and 3-54 mmol/l (median 14 mmol/l) for similarly aged Gambian nurses (44, 45), although values obtained in a group of adults from a rural district in Northern Thailand were quite acceptable (17-118 mmol/l, median 44 mmol/l) (46). However, it is difficult to assess the extent to which sub-clinical infections are lowering the plasma vitamin C concentrations seen in such countries.
Data describing a positive association between vitamin C consumption and health status are frequently reported, but intervention studies do not support the observations. Low plasma concentrations are reported in patients with diabetes (47) and infections (48) and in smokers (49), but the relative contribution of diet and stress to these situations is uncertain (see Chapter 17). Epidemiologic studies indicate that diets with a high vitamin C content have been associated with lower cancer risk, especially for cancers of the oral cavity, oesophagus, stomach, colon, and lung (39, 50-52). However, there appears to be no effect of consumption of vitamin C supplements on the development of colorectal adenoma and stomach cancer (52-54), and data on the effect of vitamin C supplementation on coronary heart disease and cataract development are conflicting (55-74). Currently there is no consistent evidence from population studies that heart disease, cancers, or cataract development are specifically associated with vitamin C status. This of course does not preclude the possibility that other components in vitamin C - rich fruits and vegetables provide health benefits, but it is not yet possible to separate such an effect from other factors such as lifestyle patterns of people who have a high vitamin C intake.
Ascorbate is found in many fruits and vegetables (75). Citrus fruits and juices are particularly rich sources of vitamin C but other fruits including cantaloupe, honeydew melon, cherries, kiwi fruits, mangoes, papaya, strawberries, tangelo, watermelon, and tomatoes also contain variable amounts of vitamin C. Vegetables such as cabbage, broccoli, Brussels sprouts, bean sprouts, cauliflower, kale, mustard greens, red and green peppers, peas, tomatoes, and potatoes may be more important sources of vitamin C than fruits. This is particularly true because the vegetable supply often extends for longer periods during the year than does the fruit supply.
In many developing countries, limitations in the supply of vitamin C are often determined by seasonal factors (i.e., the availability of water, time, and labour for the management of household gardens and the short harvesting season of many fruits). For example, mean monthly ascorbate intakes ranged from 0 to 115 mg/day in one Gambian community in which peak intakes coincided with the seasonal duration of the mango crop and to a lesser extent with orange and grapefruit harvests. These fluctuations in dietary ascorbate intake were closely reflected by corresponding variations in plasma ascorbate (11.4-68.4 mmol/L) and human milk ascorbate (143-342 mmol/L) (76).
Vitamin C is also very labile, and the loss of vitamin C on boiling milk provides one dramatic example of a cause of infantile scurvy. The vitamin C content of food is strongly influenced by season, transport to market, shelf life, time of storage, cooking practices, and chlorination of water. Cutting or bruising of produce releases ascorbate oxidase. Blanching techniques inactivate the oxidase enzyme and help to preserve ascorbate as also will low pH, as in the preparation of sauerkraut (pickled cabbage). In contrast, heating and exposure to copper or iron or to mildly alkaline conditions destroys the vitamin, and too much water can leach it from the tissues during cooking.
The use of citrus fruits by the British navy in the 18th century gave rise to the term ‘limey’, a colloquial term for British sailors. However, it is important to realise that the amount of vitamin C in a food is usually not the major determinant of a food’s importance for supply, but rather regularity of intake. For example, in countries where the potato is an important staple food and refrigeration facilities are limited, seasonal variations in plasma ascorbate are due to the considerable deterioration in the potato’s vitamin C content during storage; the content can decrease from 30 to 8 mg/100 g over 8-9 months (77). Such data can indicate the important contribution the potato can make to human vitamin C requirements even though the potato vitamin C concentration is low.
An extensive study has been made of losses of vitamin C during the packaging, storage, and cooking of blended foods (maize and soya-based relief foods). Data from a US Agency for Internation Development programme show that vitamin C losses from packaging and storage in polythene bags of such relief foods are much less significant than the 52-82 percent losses attributable to conventional cooking procedures (78).
Calculating the dietary intake from the physiologic requirements
Adults
At saturation the whole body content of ascorbate in adult males is approximately 20 mg/kg, or 1500 mg. Clinical signs of scurvy appear when the whole body content falls below 300-400 mg, and the last signs disappear when the body content reaches about 1000 mg (28, 30). In these experiments, ascorbate in the whole body was catabolised at an approximate rate of 3 percent/day (2.9 percent/day, SD 0.6) (29).
There is a sigmoidal relationship between intake and plasma concentrations of vitamin C (79). Below 30 mg/day, plasma concentrations are around 11 mmol/l. Above this intake, plasma concentrations increase steeply to 60 mmol/l and plateau at around 80 mmol/l, which represents the renal threshold. Under near steady state conditions, plateau concentrations of vitamin C are achieved by intakes in excess of 200 mg/day (Figure 8) (34). At low doses dietary vitamin C is almost completely absorbed, but over the range of usual dietary intakes (30-180 mg/day), absorption may decrease to 75 percent because of competing factors in the food (35, 80).
Figure 8. Relationship between intake and plasma concentrations of vitamin C
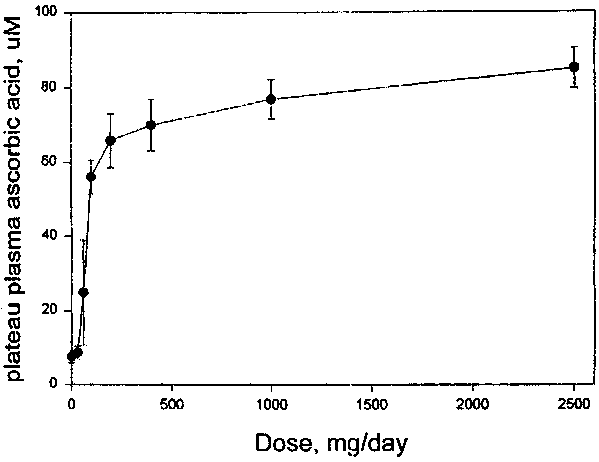
A body content of 900 mg falls half way between tissue saturation and the point at which clinical signs of scurvy appear. Assuming an absorption efficiency of 85 percent, and a catabolic rate of 2.9, the average intake of vitamin C can be calculated as:
900 x 2.9/100 x 100/85 = 30.7 mg/day, which can be rounded off to 30 mg/day.The recommended nutrient intake (RNI) would therefore be:
900 x (2.9 + 1.2)/100 x 100/85 = 43.4 mg/day, which can be rounded off to 45 mg/day.An RNI of 45 mg would achieve 50 percent saturation in the tissues in 97.5 percent of adult males. An intake of 45 mg vitamin C will produce a plasma ascorbate concentrations near the base of the steep slope of the diet-plasma dose response curve (Figure 8). No turnover studies have been done in women, but from the smaller body size and whole body content of women, requirements might be expected to be lower. However, in depletion studies plasma concentrations fell more rapidly in women than in men (81). It would seem prudent, therefore, to make the same recommendation for non-pregnant, non-lactating women as for men. Thus, an intake of 45 mg/day will ensure that measurable amounts of ascorbate will be present in the plasma of most people and will be available to supply tissue requirements for metabolism or repair at sites of depletion or damage. A whole body content of around 900 mg of vitamin C would provide at least 1 month’s safety interval, even for a zero intake, before the body content falls to 300 mg (82).
The Sheffield (27) and Iowa studies (28) indicated that the minimum amount of vitamin C needed to cure scurvy in men was less than 10 mg/day. This level however, is not sufficient to provide measurable amounts of ascorbate in plasma and leukocyte cells, which will remain low. As indicated above, no studies have been done on women and minimum requirements to protect non-pregnant and non-lactating women against scurvy might be slightly lower than in men. Although 10 mg/day will protect against scurvy, this amount provides no safety margin against further losses. The mean requirement is therefore calculated by interpolation between 10 and 45 mg/day, at an intake of 25-30 mg/day.
Pregnancy and lactation
During pregnancy there is a moderate extra drain on vitamin C, particularly during the last trimester, and 8 mg/day of vitamin C is reported to be sufficient to prevent scorbutic signs in infants aged 4-17 months (83). Therefore, the additional needs during pregnancy are unlikely to be more, particularly during the last trimester. An extra 10 mg/day throughout pregnancy should enable reserves to accumulate to meet the extra needs of the growing foetus in the last trimester.
During lactation, however, 20 mg/day of vitamin C is secreted in milk. For an assumed absorption efficiency of 85 percent, an extra 25 mg will be needed by the mother. It is therefore recommended that the RNI should be set at 70 mg to fulfil the needs of both the mother and infant during lactation.
Children
As mentioned earlier, 8 mg/day of vitamin C is sufficient to prevent scorbutic signs in infants (83). The vitamin C concentration in mature human milk is estimated to be 40 mg/l (mean, SD 10) (84), but the amount of vitamin C in human milk appears to reflect maternal dietary intake and not the infants needs (82, 83, 85). RNIs for infants aged 0-6 months are therefore set, somewhat arbitrarily, at 25 mg/day, and the RNI is gradually increased as children got older.
Elderly
Elderly people frequently have low plasma ascorbate values and intakes lower than those in younger people, often because of problems of poor dentition or mobility (86). Elderly people are also more likely to have underlying sub-clinical diseases, which can also influence plasma ascorbate concentrations (see Chapter 17). It has been suggested, however, that the requirements of elderly people do not differ substantially from those of younger people in the absence of pathology, which may influence absorption or renal functioning (82). The RNIs for the elderly are therefore the same as those for adults (45 mg/day).
Smokers
Kallner et al. (87) reported that the turnover of vitamin C in smokers was 50 percent greater than that in non-smokers. However, there is no evidence that the health of smokers would be influenced in any way by increasing their RNI. The panel therefore found no justification in making a separate RNI for smokers.
Recommended nutrient intakes for vitamin C
Table 15 presents a summary of the above discussed RNIs for vitamin C by age groups.
Table 15
Recommended nutrient intakes (RNIs) for vitamin C
|
Group |
RNI mg/day a |
|
|
Infants and children |
|
|
|
|
0-6 months |
25 |
|
|
7-12 months |
30b |
|
|
1-3 years 4-6 years |
30 b 30 b |
|
|
7-9 years |
35 b |
|
Adolescents, 10-18 years |
40 b |
|
|
Adults |
|
|
|
|
19-65 years |
45 |
|
|
65+ years |
45 |
|
Pregnancy |
55 |
|
|
Lactation |
70 |
|
Vitamin C toxicity
The potential toxicity of excessive doses of supplemental vitamin C relates to intra-intestinal events and to the effects of metabolites in the urinary system. Intakes of 2-3 g/day of vitamin C produce unpleasant diarrhoea from the osmotic effects of the unabsorbed vitamin in the intestinal lumen in most people (88). Gastrointestinal disturbances can occur after ingestion of as little as 1 g because approximately half of the amount would not be absorbed at this dose (35).
Oxalate is an end product of ascorbate catabolism and plays an important role in kidney stone formation. Excessive daily amounts of vitamin C produce hyperoxaluria. In four volunteers who received vitamin C in the range of 5-10 g/day, this amounted to approximately a doubling of urinary oxalate excretion, from 50 to 87 mg/day (range 60-126 mg/day) (89). However, the risk of oxalate stones formation may become significant at high intakes of vitamin C (>1 g) (90), particularly in subjects with high amounts of urinary calcium (89).
Vitamin C may precipitate haemolysis in some people, including those with glucose-6-phosphate dehydrogenase deficiency (91), paroxysmal nocturnal haemaglobinuria (92), or other conditions where increased risk of red cell haemolysis may occur or where protection against the removal of the products of iron metabolism may be impaired, as in people with the haptoglobin Hp2-2 phenotype (93). Of these conditions, only the haptoglobin Hp 2-2 condition was associated with abnormal vitamin C metabolism (lower plasma ascorbate than expected) under conditions where intake of vitamin C was provided mainly from dietary sources. Therefore, 1 g vitamin C appears to be the advisable upper limit of dietary intake.
Research is needed to gain a better understanding of the following:
1. Stewart, C.P. & Guthrie, D. [Editors] (1953). Lind’s treatise on scurvy. Edinburgh, University Press.
2. Nishikimi, M., Fukuyama, R., Minoshima, S., Shimizu, N. & Yagi, K. 1994. Cloning and chromosomal mapping of the Human nonfunctional gene for L-gulono-gamma-lactone oxidase, the enzyme for L-ascorbic acid biosynthesis missing in man. J. Biol. Chem., 269: 13685-13688.
3. Levine, M. 1986. New concepts in the biology and biochemistry of ascorbic acid. N. Engl. J. Med., 314:892-902.
4. Englard, S. & Seifter, S. 1986. The biochemical functions of ascorbic acid. Annu. Rev. Nutr., 6: 365-406: 365-406.
5. Wondrack, L.M., Hsu, C.A. & Abbott, M.T. 1978. Thymine 7-hydroxylase and pyrimidine deoxyribonucleoside 2'-hydroxylase activities in Rhodotorula glutinis. J. Biol. Chem., 253: 6511-6515.
6. Stubbe, J.A. 1985. Identification of two alpha keto glutarate dependent dioxygenases in extracts of Rhodotorula glutinis catalysing deoxyuridine hydroxylation. J. Biol. Chem., 260: 9972-9975.
7. Prockop, D.J. & Kivirikko, K.I. 1995. Collagens: molecular biology, diseases, and potential for therapy. Annu. Rev. Biochem., 64: 403-434.
8. Peterkofsky, B. 1991. Ascorbate requirement for hydroxylation and secretion of procollagen: relationship to inhibition of collagen synthesis in scurvy. Am. J. Clin. Nutr., 54:1135S-1140S.
9. Kivirikko, K.I. & Myllyla, R. 1985. Post-translational processing of procollagens. Ann. NY Acad. Sci., 460: 187-201.
10. Rebouche, C.J. 1991. Ascorbic acid and carnitine biosynthesis. Am. J. Clin. Nutr., 54: 1147S-1152S.
11. Dunn, W.A., Rettura, G., Seifter, E. & Englard, S. 1984. Carnitine biosynthesis from gamma-butyrobetaine and from exogenous protein-bound 6-N-trimethyl-L-lysine by the perfused guinea pig liver. Effect of ascorbate deficiency on the in situ activity of gamma-butyrobetaine hydroxylase. J. Biol. Chem., 259: 10764-10770.
12. Levine, M., Dhariwal, K.R., Washko, P.W., Butlesr, J.D., Wang, Y.H. & Bergsten, P. 1991. Ascorbic acid and in situ kinetics: a new approach to vitamin requirements. Am. J. Clin. Nutr., 54: 1157S-1162S.
13. Kaufman, S. 1974. Dopamine-beta-hydroxylase. J. Psychiatr. Res., 11: 303-316.
14. Eipper, B., Milgram, S.L., Husten, E.J., Yun, H. & Mains, R.E. 1993. Peptidylglycine alpha amidating monooxygenase: a multifunctional protein with catalytic, processing, and routing domains. Prot Sci. 2: 489-497.
15. Eipper, B., Stoffers, D.A. & Mains, R.E. 1992. The biosynthesis of neuropeptides: peptide alpha amidation. Annu. Rev. Neurosci., 15: 57-85.
16. Lindblad, B., Lindstedt, G. & Lindstedt, S. 1970. The mechanism of enzymic formation of homogentisate from p-hydroxyphenyl pyruvate. J. Am. Chem. Soc., 92: 7446-7449.
17. Schorah, C.J., Sobala, G.M., Sanderson, M., Collis, N. & Primrose, J.M. 1991. Gastric juice ascorbic acid: effects of disease and implications for gastric carcinogenesis. Am. J. Clin. Nutr., 53: 287S-93S.
18. Correa, P. 1992. Human gastric carcinogenesis: a multistep and multifactorial process-First American Cancer Society Award Lecture on Cancer Epidemiology and Prevention. Cancer Res., 52: 6735-6740.
19. Byers, T. & Guerrero, N. 1995. Epidemiologic Evidence for vitamin C and vitamin E in cancer prevention. Am. J. Clin. Nutr., 62: 1385S-1392S.
20. Jialal, I. & Grundy, S.M. 1991. Preservation of the endogenous antioxidants in low density lipoprotein by ascorbate but not probucol during oxidative modification. J. Clin. Inv., 87: 597-601.
21. Stokes, P.L., Melikian, V., Leeming, R.L., Portman-Graham, H. Blair, J.A. & Cooke, W.T. 1975. Folate metabolism in scurvy. Am. J. Clin. Nutr., 28: 126-9.
22. Hallberg, D., Brune, M. & Rossander-Hulthen, L. 1987. Is there a physiological role of vitamin C in iron absorption. Ann. NY Acad. Sci., 498: 324-332
23. Hallberg, L., Rossander, L., Persson, H. & Svahn, E. 1982. Deleterious effects of prolonged warming of meals on ascorbic acid content and iron absorption. Am. J. Clin. Nutr., 36: 846-850
24. Hallberg, L. 1987. Wheat fiber, phytates and iron absorption. Scand. J. Gastroenterol. Suppl., 129: 73-9: 73-79.
25. McLaren, D.S. 1992. A colour atlas of nutritional disorders. London, Wolfe Medical Publications.
26. Bartley, W., Krebs, H.A. & O’Brien, J.R.P. 1953. Vitamin C requirements of Human adults. Medical Research Council Special Report Series No. 280, London, HMSO.
27. Krebs, H.A. & Vitamin C Subcommittee of the Accessory Food Factors Committee M.R.C. 1948. Vitamin C Requirement of Human Adults: Experimental Study of Vitamin C Deprivation in Man. Lancet, 254: 853-858.
28. Baker, E.M., Hodges, R.E., Hood, J., Sauberlich, H.E. & March, S.C. 1969. Metabolism of ascorbic-1-14C acid in experimental Human scurvy. Am. J. Clin. Nutr., 22: 549-558.
29. Baker, E.M., Hodges, R.E., Hood, J., Sauberlich, H.E. March, S.C. & Canham, J.E. 1971. Metabolism of 14C- and 3H-labeled L-ascorbic acid in Human scurvy. Am. J. Clin Nut., 24: 444-454.
30. Kallner, A., Hartmann, D. & Hornig, D. 1979. Steady-state turnover and body pool of ascorbic acid in man. Am. J. Clin. Nutr., 32: 530-539.
31. Moser, U. & Weber, F. 1984. Uptake of ascorbic acid by Human granulocytes. Int. J. Vit. Nutr. Res., 54: 47-53
32. Lee, W., Davis, K.A., Rettmer, R.L. & Labbe, R.F. 1988. Ascorbic acid status: biochemical and clinical considerations. Am. J. Clin. Nutr., 48:286-290.
33. McCormick, D.B. & Zhang, Z. 1993. Cellular assimilation of water-soluble vitamins in the mammal: riboflavin, B6, biotin and C. Proc. Soc. Exp. Biol. Med., 202: 265-270.
34. Levine, M., Conry-Cantilena, C., Wang, Y., Welch R.W., Washko, P.W., Dhariwal K.R., Park, J.B., Lazarev, A. & Graumlich, J.K. 1996. Vitamin C pharmacokinetics in healthy volunteers: evidence for a Recommended Dietary Allowance. Proc. Natl. Acad. Sci., 93: 3704-3709.
35. Graumlich, J., Ludden, T.M., Conry-Cantilena, C., Cantilena, L.R., Wang, Y. & Levine M. 1997. Pharmacokinetic model of ascorbic acid in Humans during depletion and repletion. Pharmaceut. Res., 14: 1133-1139.
36. Arab, L., Schellenberg, B. & Schlierf, G. 1982. Nutrition and health. A survey of young men and women in Heidelberg. Ann. Nutr. Metab., 26: 1-77.
37. Gregory, J.R., Foster, K., Tyler, H. & Wiseman, M. 1990. The dietary and nutritional survey of British adults. London: HMSO.
38. Life Sciences Research Office, Interagency Board for Nutrition Monitoring and Related Research. 1995. Third Report on Nutrition Monitoring in the United States. Washington, D.C. U. S. Government Printing Office.
39. Finch, S., Doyle, W., Bates, C.J. Prentice, A., Smithers, G. & Clarke, P.C. 1998. National diet and nutrition survey: people aged 65 years and over. Volume 1: Report of the diet and nutrition survey. London: The Stationery Office.
40. Basch, C.E., Syber, P. & Shea, S. 5-a-day:dietary behavior and the fruit and vegetable intake of Latino children. Am. J. Public Health 1994; 84:814-818.
41. WHO. 1990. Diet, Nutrition and the Prevention of Chronic Diseases. WHO Technical Report Series 797. Geneva: World Health Organization.
42. Williams, C. 1995. Healthy eating: clarifying advice about fruit and vegetables. BMJ, 310: 1453-55.
43. Narasinga Rao, B.S. 1995. Dietary intake of antioxidants in relation to nutrition profiles of Indian population groups. Nutrition, lipids, health and disease, p.343-353 [A.S.H. Ong, E. Niki, and L. Packer editors]. Champaign Illinois: AOCS Press.
44. Chang-Claude, J.C. 1990. Epidemiologic study of precancerous lesions of the oesophagus in young persons in a high-incidence area for the oesophageal cancer in China. Heidelberg: Heidelberg University.
45. Knowles, J., Thurnham, D.I., Hill, A.V.S., Tang, C. & Greenwood, B.M. 1991. Plasma ascorbate concentrations in Human malaria. Proc. Nutr. Soc., 50: 66A.
46. Thurnham, D.I., Singkamani, R., Kaewichit, R. & Wongworapat, K. 1990. Influence of malaria infection on peroxyl-radical trapping capacity in plasma from rural and urban Thai adults. Br. J. Nutr., 64: 257-271.
47. Jennings, P.E., Chiroco, S., Jones, A.F., Lunec, J. & Barnett, A.H. 1987. Vitamin C metabolites and microangiography in diabetes mellitis. Diab. Res., 6: 151-154
48. Thurnham, D.I. 1994. b-Carotene, are we misreading the signals in risk groups? Some analogies with vitamin C. Proc. Nutr. Soc., 53: 557-569
49. Faruque, O., Rahman Khan, M., Rahman M. & Ahmed F. 1995. Relationship between smoking and antioxidant status. Br. J. Nutr., 73: 625-632.
50. Yong, L., Brown, C.C., Schatzkin, A., Dresser, C.M., Slesinski, M.J., Cox, C.S., & Taylor P.R. 1997. Intake of vitamins E, C, and A and risk of lung cancer. Am. J. Epidemiol.,146: 231-243.
51. Byers, T. & Mouchawar, J. 1998. Antioxidants and cancer prevention in 1997. In: Paoletti R, Sies H, Bug J., Grossi E, Poli A, eds. Vitamin C: the state of the art in disease prevention 60 years after the Nobel Prize. ,p. 29-40. Milan: Springer.
52. Schorah, C.J. 1998. Vitamin C and gastric cancer prevention. In: Paoletti R, Sies H, Bug J., Grossi E, Poli A, eds. Vitamin C: the state of the art in disease prevention sixty years after the Nobel Prize. p. 41-49. Milan: Springer,
53. Blot ,W.J., Li, J., Taylor, P.R., Guo, W., Dawsey, S., Wang, G.Q., Yang, C.S., Zheng, S.F., Gail, M. & Li G.Y. 1993. Nutrition intervention trials in Linxian, China: supplementation with specific vitamin/mineral combinations, cancer incidence, and disease-specific mortality in the general population. J. Natl. Cancer. Instit., 85: 1483-1492.
54. Greenberg, E.R., Baron, J.A. Tosteson, T.D., et al. 1994. A clinical trial of antioxidant vitamins to prevent colorectal adenoma. Polyp Prevention Study Group. N. Engl. J. Med., 331: 141-147.
55. Rimm, E.B., Stampfer, M.J., Ascherio, A., Giovannucci, E., Colditz, G.A. & Willett, W.C. 1993. Vitamin E consumption and the risk of coronary heart disease in men. N. Engl. J. Med., 328: 1450-1456.
56. Sahyoun, N.R., Jacques, P.F. & Russell, R.M. 1994. Carotenoids, vitamins C and E, and mortality in an elderly population. Am. J. Epidemiol., 144: 501-511.
57. Jha P., Flather, M., Lonn, E., Farkouh, M. & Yusuf, S. 1995. The antioxidant vitamins and cardiovascular disease: a critical review of the epidemiologic and clinical trial data. Ann. Intern. Med., 123: 860-872.
58. Losonczy, K.G., Harris, T.B. & Havlik, RJ. 1996. Vitamin E and vitamin C supplement use and risk of all cause and coronary heart disease mortality in older persons: the established populations for epidemiologic studies of the elderly. Am. J. Clin. Nutr., 64: 190-196.
59. Enstrom, J.E., Kanim, L.E. & Klein, M.A. 1992. Vitamin C intake and mortality among a sample of the United States population [see comments]. Epidemiology, 3: 194-202.
60. Enstrom, J.E., Kanim, L.E., & Breslow, L. 1986. The relationship between vitamin C intake, general health practices, and mortality in Alameda County, California. Am. J. Public Health, 76: 1124-1130.
61. Seddon, J.M., Ajani, U.A., Sperduto, R.D., et al. 1995. Dietary carotenoids, vitamins A, C, and E, and advanced age-related macular degeneration. Eye Disease Case-Control Study Group [see comments] [published erratum appears in JAMA 1995 Feb 22;273(8):622]. JAMA, 272: 1413-1420.
62. Riemersma, R.A., Wood, D.A., Macintyre, C.C., Elton, R.A., Gey, K.F. & Oliver M.F. 1991. Risk of angina pectoris and plasma concentrations of vitamins A, C, and E and carotene [see comments]. Lancet, 337:1-5.
63. Gey, K.F., Moser, U.K., Jordan, P., Stahelin, H.B., Eichholzer, M. & Ludin, E. 1993. Increased risk of cardiovascular disease at suboptimal plasma concentrations of essential antioxidants: an epidemiological update with special attention to carotene and vitamin C. Am. J. Clin. Nutr., 57:787S-797S.
64. Kushi, L.H., Folsom, A.R., Princas R.J., Mink, P.J., Wu, Y. & Bostick, R.M. 1996. Dietary antioxidant vitamins and death from coronary heart disease in postmenopausal women. N. Engl. J. Med., 334: 1156-1162.
65. Simon, J.A., Hudes, E.S. & Browner, W.S. 1998. Serum ascorbic acid and cardiovascular disease prevalence in US adults. Epidemiology, 9: 316-321.
66. Jacques, P.F., Chylack, L.T., Jr., McGandy, R.B. & Hartz, S.C. 1988. Antioxidant status in persons with and without senile cataract. Arch. Ophthalmol., 106: 337-340.
67. Robertson, J.M., Donner, A.P. & Trevithick, J.R. 1991. A possible role for vitamins C and E in cataract prevention. Am. J. Clin. Nutr., 53:346S-351S.
68. Leske, M.C., Chylack, L.T. & Wu, S. 1991. The lens opacities case/control study: risk factors for cataract. Arch. Opthalmol., 109: 144-251.
69. Italian-American Cataract Study Group. 1991. Risk factors for age-related cortical, nuclear, and posterior sub-capsular cataracts. Am. J. Epidemiol., 133: 541-553.
70. Goldberg, J., Flowerdew, G., Smith, E., Brody, J.A. & Tso, M.O. 1988. Factors associated with age-related macular degeneration. An analysis of data from the first National Health and Nutrition Examination Survey. Am. J. Epidemiol.,128: 700-710.
71. Vitale, S., West, S., Hallfrisch, J., et al. 1993. Plasma antioxidants and risk of cortical and nuclear cataract. Epidemiology, 4: 195-203.
72. Hankinson, S.E., Stampfer, M.J., Seddon, J.M., et al. 1992. Nutrient intake and cataract extraction in women: a prospective study. BMJ, 305: 335-339.
73. Mares-Perlman, J.A. 1997. Contribution of epidemiology to understanding relationships of diet to age-related cataract. Am. J. Clin. Nutr., 66: 739-740.
74. Jacques, P.F., Taylor, A., Hankinson, S.E., Willett, W.C., Mahnken B., Lee, Y., Vaid, k. & Lahav, M. 1997. Long-term vitamin C supplement use and prevalence of early age-related lens opacities. Am. J. Clin. Nutr., 66: 911-916.
75. Haytowitz, D. 1995. Information from USDA's Nutrient Data Book. J. Nutr., 125: 1952-1955.
76. Bates, C.J., Prentice, A.M. & Paul, A.A. 1994. Seasonal variations in vitamins A, C, riboflavin and folate intakes and status of pregnant of lactating women in a rural Gambian community: some possible implications. Eur. J. Clin. Nutr., 48: 660-668.
77. Paul, A.A. & Southgate, D.A.T. 1979. McCance and Widdowson’s, The Composition of Foods. London: HMSO.
78. N.A.S. Institute of Medicine. 1997. Vitamin C fortification of food aid commodities: Final Report, Washington D.C., National Academy Press.
79. Newton, H.M.V., Morgan, D.B., Schorah, C.J. & Hullin, R.P. 1983. Relation between intake and plasma concentration of vitamin C in elderly women. BMJ, 287: 1429
80. Melethil, S.L., Mason, W.E. & Chiang, C. 1986. Dose dependent absorption and excretion of vitamin C in Humans. Int. J. Pharm., 31:83-89.
81. Blanchard, J. 1991. Depletion and repletion kinetics of vitamin C in Humans. J. Nutr., 121: 170-176.
82. Olson, J.A. & Hodges, R.E. 1987. Recommended dietary intakes (RDI) of vitamin C in Humans. Am. J. Clin. Nutr., 45: 693-703.
83. Irwin, M.I. & Hutchins, B.K. 1976. A conspectus of research on vitamin C requirements in man. J. Nutr., 106: 821-879.
84. WHO. 1998. Complementary feeding of young children in developing countries: a review of current scientific knowledge. Geneva: World Health Organization.
85. Van Zoeren-Grobben, D., Schrijver, J., Van den Berg, G.J. & Berger, H.M. 1987. Human milk vitamin content after pasteurisation, storage, or tube feeding. Arch. Dis. Child., 62:161-165
86. Department of Health and Social Security. 1979. Nutrition and health in old age. Report on Health and Social Subjects. No. 16. London: H.M.S.O.
87. Kallner, A.B., Hartmann, D. & Hornig, D.H. 1981. On the requirements of ascorbic acid in man: steady state turnover and body pool in smokers. Am. J. Clin. Nutr., 34: 1347-1355.
88. Kubler, W. & Gehler, J. 1970. On the kinetics of the intestinal absorption of ascorbic acid: a contribution to the calculation of an absorption process that is not proportional to the dose. Int. J. Vit. Nutr. Res., 40: 442-453
89. Schmidt K.-H., Hagmaier, V., Hornig, D.H., Vuilleumier, J. & Rutishauser, G. 1981. Urinary oxalate excretion after large intakes of ascorbic acid in man. Am. J. Clin. Nutr., 34: 305-311
90. Urivetzky, M., Kessaris D. & Smith, A.D. 1992. Ascorbic acid overdosing: a risk factor for calcium oxalate nephrolithiasis. J. Urol., 147: 1215-1218.
91. Mehta J.B., Singhal S.B. & Mehta B.C. 1990. Ascorbic acid induced haemolysis in G-6-PD deficiency. Lancet, 336: 944.
92. Iwamoto, N., Kawaguchi, T., Horikawa, K., Nagakura, S., Hidaka M., Kagimoto T., Takatsuki, K. & Nakakuma H. 1994. Haemolysis induced by ascorbic acid in paroxysmal nocturnal haemoglobinuria. Lancet, 343: 357.
93. Langlois, M.R., Delanghe, J.R., De Buyzere, M.L., Bernard, D.R. & Ouyang, J. 1997. Effect of haptoglobin on the metabolism of vitamin C. Am. J. Clin. Nutr., 66: 606-610.
![]()
![]()
![]()