![]()
![]()
![]()
The role of selenium in human metabolic processes
Selenium deficiency
The influence of diet on selenium status
Absorption and bio-availability
Criteria for assessing selenium requirements
Recommended selenium intakes
Future research
References
Our understanding of the significance of selenium in the nutrition of human subjects has grown rapidly during the past 20 years (1, 2). Demonstrations of its essentiality to rats and farm animals were followed by appreciation that the development of selenium-responsive diseases often reflected the distribution of geochemical variables which restricted the entry of the element from soils into food chains. Such findings were the stimulus to in-depth investigations of the regional relevance of selenium in human nutrition (3). These studies have now yielded an increased understanding of the complex metabolic role of this trace nutrient. Selenium has been implicated in the protection of body tissues against oxidative stress, maintenance of defences against infection, and modulation of growth and development.
The selenium content of normal adult humans can vary widely. Values from 3 mg in New Zealanders to 14 mg in some Americans reflect the profound influence of the natural environment on the selenium contents of soils, crops, and human tissues. Approximately 30 percent of tissue selenium is contained in the liver, 15 percent in kidney, 30 percent in muscle, and 10 percent in blood plasma. Much of tissue selenium is found in proteins as seleno-analogues of sulphur amino acids; other metabolically active forms include selenotrisulphides and other acid-labile selenium compounds. At least 15 selenoproteins have now been characterised. Examples are given in Table 47.
Table 47
A selection of characterised selenoproteins
|
Protein |
Selenocysteine residues |
Tissue distribution |
|
Cytosolic glutathione peroxidase (GSHPx) |
1 |
All, including thyroid |
|
Phospholipid hydroperoxide GSHPx |
1 |
All, including thyroid |
|
Gastrointestinal GSHPx |
1 |
Gastrointestinal tract |
|
Extracellular GSHPx |
1 |
Plasma, thyroid |
|
Thioredoxin reductase |
1 or 2 |
All including thyroid |
|
Iodothyronine-deiodinase (type 1) |
1 |
Liver, kidneys, and thyroid |
|
Iodothyronine-deiodinase (type 2) |
1 |
Central nervous system (CNS), and pituitary |
|
Iodothyronine-deiodinase (type 3) |
1 |
Brown adipose tissue, CNS, and placenta |
|
Selenoprotein P |
10 |
Plasma |
|
Selenoprotein W |
1 |
Muscle |
|
Sperm capsule selenoprotein |
3 |
Sperm tail |
H2O2 + 2GSH àà
2H2O + GSSG
ROOH + 2GSH àà ROH + H2O + GSSG
where GSH is glutathione and GSSG is its oxidized form. At least four forms of GSHPx exist; they differ both in their tissue distribution and in their sensitivity to selenium depletion (4). The GSHPx enzymes of liver and blood plasma fall in activity rapidly at early stages of selenium deficiency. In contrast, a form of GSHPx associated specifically with phospholipid-rich tissue membranes is preserved against selenium deficiency and is believed to have broader metabolic roles (e.g., in prostaglandin synthesis) (6). In concert with vitamin E, selenium is also involved in the protection of cell membranes against oxidative damage (see Chapter 6, Chapter 9, and Chapter 17).
The selenoenzyme thioredoxin reductase is involved in disposal of the products of oxidative metabolism (5). It contains two selenocysteine groups per molecule and is a major component of a redox system with a multiplicity of functions, among which is the capacity to degrade locally excessive and potentially toxic concentrations of peroxide and hydroperoxides likely to induce cell death and tissue atrophy (6).
Another group of selenoproteins is essential in the conversion of thyroxin, or tetraiodothyronine (T4), to its physiologically active form, triiodothyronine (T3) (7). Three types of these iodothyronine deiodinases, differing both in tissue distribution and sensitivity to selenium deficiency, have been characterised. The consequences of a low selenium status on physiologic responses to a shortage of iodine are complex. The influence of a loss of selenium-dependent iodothyronine deiodinase differs in its severity depending on whether a target tissue needs a preformed supply of T3 (e.g., via plasma) or whether, as with the brain, pituitary gland, and placenta, it can rely upon local synthesis of T3 from T4. Despite this, marked changes in the T3-T4 ratio as a consequence of a reduced selenium status (when iodine supplies are also marginal) indicate the modifying influence of selenium on thyroid hormone balance in both animal models and human subjects. Their possible significance can be anticipated from the fact that whereas thyroid weights increase typically by 50 percent in rats offered an iodine-deficient diet, thyroid weight is increased 154 percent by diets concurrently deficient in both selenium and iodine.
Between 60 percent and 80 percent of selenium in human plasma is accounted for by a well-characterised fraction designated selenoprotein P, the function of which has yet to be determined. It is thought to be a selenium storage protein because there is limited evidence that it also has an antioxidant role. At least 10 other selenoproteins exist, including one which is a component of the mitochondrial capsule of sperm cells, damage to which may account for the development of sperm abnormalities during selenium deficiency. Other aspects of the function and metabolism of selenium are reviewed elsewhere (8, 9).
Non-endemic deficiencies of selenium
Biochemical evidence of selenium depletion (e.g., a decline in blood GSHPx activity) is not uncommon in subjects maintained on parenteral or enteral feeding for long periods. Blood selenium values declining to one-tenth of normal values have been reported when the selenium content of such preparations has not been maintained by fortification (10, 11). Low selenium contents of some commercial formulas for infants resulting in a fall in daily selenium intake to approximately 0.5 µg/day have been shown to strongly exacerbate the fall in serum selenium and GSHPx activity normally experienced from 2 to 8 months of age even in human-milk-fed infants typically receiving threefold higher selenium intakes (12, 13). The importance of maintaining trace element levels in such preparations was reviewed elsewhere (14).
Clinical manifestations of deficiency arising from such situations are uncommon and poorly defined. They include muscular weakness and myalgia with, in several instances, the development of congestive heart failure. In at least one instance such pathologic signs have developed as a consequence of a generally inadequate diet providing selenium at less than 10 µg/day. The 2-year-old subject recovered rapidly after selenium administration (15). With this last exception, virtually all of the above reports describe observations with subjects under close medical supervision. This may well be relevant to the scarcity of consistent pathologic findings (16).
Keshan disease
Keshan disease was first described in Chinese medical literature more than 100 years ago, but not until 40 years after its widespread occurrence in 1935 was it discovered that selenium deficiency was an important factor in its aetiology (3). Endemic in children aged 2-10 years and in women of childbearing age, this disease has a geographic distribution covering localities from northeast to southwest China. Typical manifestations are fatigue after even mild exercise, cardiac arrhythmia and palpitations, loss of appetite, cardiac insufficiency, cardiomegaly, and congestive heart failure. Pathologic changes include a multifocal myocardial necrosis and fibrosis. The coronary arteries are essentially unaffected. Ultrastructural studies show that membranous organelles, such as mitochondria or sarcolemma, are affected earliest. The disease has a marked seasonal fluctuation in incidence (3) and may appear after only 3 months exposure to conditions in localities known to be associated with a high risk of myocarditis (3, 8). Once the disease is established, selenium is of little or no therapeutic value. Prophylaxis consisting of oral administration of selenium 3 months before the periods of highest anticipated risk is highly effective.
Although geographic similarities in the distribution of Keshan disease and the selenium and vitamin E-responsive white muscle disease in animals first prompted successful investigation of the relevance of a low selenium status, evidence has grown steadily that the disease is multifactorial in origin. The strongest suspicions have fallen on the development of a viral myocarditis probably attributable to enhancement of the virulence of a coxsackievirus during its passage through selenium-deficient host tissues (17). Although other nutritional variables such as a marginal vitamin E status may also be involved, the finding of extremely low selenium contents in staple crops of affected areas and convincing demonstrations of the prophylactic effectiveness of selenium administration leave no doubt that selenium deficiency is the primary factor (3, 18).
Recent studies indicate that geochemical variables have an important influence on the distribution of Keshan disease. Acid soils high in organic matter and iron oxide content appear to be responsible for fixing selenium in forms which are poorly absorbed by staple crops which, in the instance of cereal grains, typically have a selenium content of less than 0.01 µg/g (19). A similar geochemical background is believed to be associated with reports of selenium-responsive disorders resembling Keshan disease in the Transbaikalia region of south Siberia. Dietary intakes of selenium are inadequate to maintain blood GSHPx activity; biochemical indicators of tissue peroxidative damage are elevated until selenium therapy is initiated (8).
Kaschin-beck disease
A selenium-responsive bone and joint disease (osteoarthropathy) has been detected in children aged 5-13 years in China and less extensively in south-east Siberia. The disease is characterised by joint necrosis - epiphyseal degeneration of the arm and leg joints resulting in structural shortening of the fingers and long bones with consequent growth retardation and stunting (3, 20). Although not identical to Keshan disease, Kaschin-Beck disease also occurs in areas where the availability of soil selenium for crop growth is low. The selenium contents of hair and of whole blood are abnormally low and the blood content of GSHPx is reduced. Although it is ameliorated by selenium therapy, other factors such as the frequent presence of mycotoxins in cereal grains grown in the area may be involved. A spontaneous decrease in incidence from 1970 (44 percent) to 1980 (14 percent) to 1986 (1 percent) has been attributed to general improvements in the nutritional status of Chinese rural communities (20).
Selenium status and susceptibility to infection
As stated earlier, the expressions of the cardiac lesions of Keshan disease probably involve not only the development of selenium deficiency but also the presence of a Coxsackie virus (BA) infection. Animal studies have confirmed that selenium-deficient mice infected with Coxsackie virus (CVB/0) were particularly susceptible to the virus. These studies also illustrated that passage of the virus through selenium-deficient subjects enhanced its virulence (17). Myocarditic virulence developed even in strains such as CVB/0 which normally were not myopathogenic. The enhancement of virulence in this RNA virus involves modifications to the nucleotide sequence of the phenotype. These modifications were maintained and expressed even during subsequent passage through animals with normal selenium status (21).
Enhancing the virulence of a virus with a selenium deficiency (resulting either from a nutritional challenge or an increased metabolic demand on tissue selenium depots) appears not to be unique to the Coxsackie viruses. The early pre-clinical stages of development of human immunodeficiency virus (HIV) infection are accompanied by a very marked decline in plasma selenium. Sub-clinical malnutrition assumes increased significance during the development of acquired immune deficiency syndrome (AIDS). However, for the nutrients affected, there are strong indications that only the extent of the decline in selenium status has predictive value with respect to both the rate of development of AIDS and its resulting mortality (22-25). The virulence of other RNA viruses such as hepatitis B and those associated with the development of haemolytic anaemias are enhanced similarly by a decline in selenium status. The mechanisms underlying these effects are not yet resolved. There are indications that the loss of protective antioxidant functions dependent on selenium and vitamin E are both involved and that the resulting structural changes in viral nucleotide sequences are reproducible and appear to provoke additional selenoprotein synthesis (26). It is suspected that this further depletes previously diminished pools of physiologically available selenium and accelerates pathologic responses (27-29).
Whatever mechanisms are involved, further understanding is needed of the influence of selenium status on susceptibility to viral diseases ranging from cardiomyopathies to haemolytic anaemias. The relationship already illustrates the difficulty of defining nutritional essentiality for nutrients which may primarily maintain defences against infection. Studies of the effects of selenium deficiency in several experimental animal species have shown that the microbicidal activity of blood neutrophils is severely impaired even though phagocytic activity remains unimpaired (30, 31). The complexity of species differences in the influence of selenium status on the effectiveness of cell-mediated immune processes is summarised elsewhere (8).
The possibility that increased intakes of selenium might protect against the development of cancer in humans has generated great interest (32). However, a number of epidemiologic studies have now been reported which show no relationship between selenium and cancer risk (33). Moreover, an analysis of the relationship between selenium and cancer suggests that “the question of whether selenium protects against cancer is still wide open” (34). An increased intake of selenium appears to stimulate tumorigenesis in some animal models of pancreatic and skin cancer. In contrast, the protective effect of higher exposures to selenium observed in several animal studies, together with small but statistically significant differences in selenium blood plasma levels detected in some retrospective-prospective studies of subgroups of people developing cancer, explains the continuing interest in the anticarcinogenic potential of selenium. However, the results of prospective-retrospective studies had no predictive value for individuals and could have reflected non-specific influences on groups. The association between low selenium intake and high cancer risk, although clearly of some interest, is in need of further investigation before a conclusion can be reached.
Although a biochemical mechanism can be postulated whereby selenium could protect against heart disease by influencing platelet aggregation (through an effect on the prostacyclin-thromboxane ratio), the epidemiologic evidence linking selenium status and risk of cardiovascular disease is still equivocal (33).
Selenium and thyroid hormones
The importance of selenium for thyroid hormone metabolism (35, 36) is evident from changes in the T3-T4 ratio which develop after relatively mild selenium depletion in infants and elderly (65+ years) subjects. Decreases in the T3-T4 ratio indicative of decreased thyroid hormone balance have been detected when serum selenium falls below 0.9 µmol/l (37). In a recent Scottish study these decreases were correlated with a decline in dietary and plasma selenium after the replacement of selenium-rich wheat from Canada and the United States with selenium-deficient wheat from European sources (38).
Communities noted for a high incidence of myxedematous cretinism have been found to have low plasma selenium status and GSHPx activity in addition to having low iodine status (39) and being exposed to high thiocyanate intakes from cassava. Restoration of iodine supply, particularly if excessive, tends to induce a high peroxidative stress through the action of iodide peroxidase, the first step in iodine utilisation by the thyroid. It is postulated that necrosis and thyroid fibrosis leading to irreversible hypothyroidism result if a concurrent deficiency of selenium limits peroxide destruction by the protective action of the selenium-dependent enzymes, GSHPx and, more probably, thioredoxin reductase (40). In areas where myxedematous cretinism is endemic and characterised by persistent hypothyroidism, dwarfism, and stunting, it has been recommended that attempts to introduce iodine therapy for mildly affected individuals should be preceded by an assessment of selenium status and rectification of any observed deficit (39). Although this suggestion is compatible with pathologic observations on hypothyroid rats differing in selenium status, its validity has yet to be assessed adequately in humans (41, 42).
Environmental conditions and agricultural practices have a profound influence on the selenium content of many foods. Tables 48, 49 and 50 illustrate the wide range of selenium content of the principal food groups and the variability in the selenium content of individual foods. This variability is exceeded only by that found in the iodine content of foods. Geographic differences in the content and availability of selenium from soils to food crops and animal products have a marked effect on the selenium status of entire communities. For example, the distribution of Keshan disease and Kaschin-Beck disease in China reflects the distribution of soils from which selenium is poorly available to rice, maize, wheat, and pasture grasses (Table 48). Cereal crop selenium contents of 3-7 ng/g are not uncommon (3) and it has been suggested that <10 ng/g for grain selenium and <3 ng/g for water-soluble soil selenium could be used as indexes to define deficient areas (19). Fluctuations in the selenium status of many communities in northern Europe reflect the intrinsically low selenium content of its glacial soils and the extent to which selenium supplementation of fertilisers has been successful in increasing the selenium content of cereal grains, milk, and other animal products. Deliberate importation of cereals from areas with relatively high available selenium in soil has also occurred or been recommended in some areas of Finland, New Zealand, and the United Kingdom after steady declines in the selenium status of some communities were noted. Conversely, low-selenium grains are being selected in China, India, and Venezuela to reduce the risks of selenosis.
Comprehensive data summarising the selenium contents of staple foods are available elsewhere (e.g., 45). Reports from the Food and Agriculture Organization of the United Nations (FAO) and the International Atomic Energy Agency (IAEA) provide representative data on daily total selenium intakes for more than 40 countries (see reference 8, p 215-217). The great influence of dietary and geographic variables on selenium status is evident from recent summaries of data describing national and regional differences for the selenium content of human and formula milks, diets, and human serum (Tables 48, 49, 50 and 51).
Selenium compounds are generally very efficiently absorbed by humans, and selenium absorption does not appear to be under homeostatic control (80). For example, absorption of the selenite form of selenium is greater than 80 percent whereas that of selenium as selenomethionine or as selenate may be greater than 90 percent (80, 81). Therefore, the rate-limiting step determining the overall availability of dietary selenium is not likely to be its absorption but rather its conversion within tissues to its metabolically active forms (e.g., its incorporation into GSHPx or 5'-deiodinase) (40). A number of depletion-repletion experiments have been carried out on animals to estimate the bio-availability of selenium in human foods (82). Based on the restoration of GSHPx activity in depleted rats, the bio-availability of selenium in wheat is quite good, usually 80 percent or better. The selenium in Brazil nuts and beef kidney also appears readily available (90 percent or more by most criteria). The selenium in tuna seems of lesser availability (perhaps only 20-60 percent of that from selenite) whereas the availability of selenium from certain other seafoods (shrimp, crab, and Baltic herring) is high. The selenium in a variety of mushrooms appears to be of uniformly low availability to rats.
Data on the nutritional bio-availability of selenium to humans are sparse. A supplementation study carried out on Finnish men of relatively low selenium status showed that selenate selenium was as effective as the selenium in seleniferous wheat in increasing platelet GSHPx activity (83). The wheat selenium, however, increased plasma selenium levels more than did selenate selenium and once the supplements were withdrawn, platelet GSHPx activity declined less in the group given wheat. This study showed the importance of estimating not only short-term availability but also long-term retention and the convertibility of tissue selenium stores into biologically active forms.
Table 48
The selenium contents of foods and diets
A. Typical ranges of selenium concentrations (ng/g fresh wt) in food groups.
|
Food group |
India (43) |
United States (44) |
International compilation (8) |
|
Cereals and cereal products |
5-95 |
10-370 |
10-550 |
|
Meat, meat products, and eggs |
40-120 |
100-810 |
10-360 |
|
Fish and marine |
280-1080 |
400-1500 |
110-970 |
|
Fish and freshwater |
- |
- |
180-680 |
|
Pulses |
10-138 |
- |
- |
|
Dairy products |
5-15 |
10-130 |
1-170 |
|
Fruits and vegetables |
1-7 |
1-60 |
1-20 |
|
|
China |
China |
India |
India |
Finland |
United Kingdom |
|
|
Keshan disease area (18) |
Disease-free area (18) |
Low-income vegetarian diets (43) |
Low-income conventional diets (43) |
(45) |
(46) |
|
Total diet |
7.7 |
16.4 |
27.4 |
52.5 |
30 |
31 |
|
Food group |
|
|
|
|
|
|
|
Cereals and cereal products |
5.4 |
11.6 |
15.7 |
21.1 |
2.8 |
7 |
|
Pulses |
- |
- |
3.9 |
3.6 |
1.1 |
- |
|
Meat and eggs |
>0.6 |
> 2.2 |
- |
3.7 |
9.2 |
10 |
|
Fish |
|
|
- |
18.4 |
9.5 |
4 |
|
Dairy products |
|
|
6.9 |
4.8 |
6.5 |
3 |
|
Fruits and vegetables |
1.7 |
2.6 |
0.9 |
0.9 |
0.5 |
6 |
|
Other |
- |
- |
- |
- |
1.1 |
3 |
Geographic differences in the selenium intakes of infants
|
Region or country |
Selenium intake (µg/day)a |
Reference |
|
Human milk |
|
|
|
China: Keshan disease area |
2.0 |
18 |
|
Burundi |
4.7 ± 0.8 |
48 |
|
New Zealand: south island |
5.3 |
49 |
|
Former Yugoslavia |
6.0 ± 1.3 |
50 |
|
Finland |
4.0-7.6 |
51 |
|
New Zealand: north island |
8.1-10.2 |
52 |
|
Belgium |
8.4 |
53 |
|
Austria |
8.8-9.8 |
13 |
|
Australia |
9.4 ± 3.6 |
54 |
|
Hungary |
9.6 ± 3.7 |
50 |
|
Sweden |
10.6 ± 2.3 |
50 |
|
United States, east coast |
8.8-11.4 |
55 |
|
United States, unspecified |
12.3 |
56 |
|
Zaire |
12.3 ± 3.6 |
50 |
|
Chile |
14.1 ± 2.6 |
50 |
|
India |
14.1 ± 3.6 |
50 |
|
Germany |
19.3 |
57 |
|
Philippines |
22.9 ± 4.1 |
50 |
|
China (seleniferous area) |
199 |
18 |
|
International reference value |
13.0 ± 18.2 |
58 |
|
Infant formula |
|
|
|
Belgium |
2.0 |
53 |
|
New Zealand |
3.3 |
59 |
|
Austria |
3.6 |
13 |
|
United Kingdom |
4.9 (2.3-8.2) |
53 |
|
Spain |
6.6 |
53 |
|
United States, 1982 |
5.9 (4.2-8.1) |
60 |
|
Germany |
6.5-6.8 |
57 |
|
New Zealand (selenium fortified) |
11.3 |
59 |
|
United States, 1997 |
11.7-18.3 |
61 |
Table 50
Geographic differences in the selenium intakes (µg/day) of adults
|
Region or country |
Selenium intake a |
Reference |
|
China, Keshan disease area |
3-11 |
62, 63 |
|
China, Kaschin -Beck disease area |
2.6-5.0 |
20 |
|
Sweden, vegans |
10 |
64 |
|
New Zealand, low-selenium area |
11 ± 3 |
64, 65 |
|
China, disease-free area |
13.3 ± 3.1 |
18 |
|
South Sweden, conventional diets |
40 ± 4 |
66 |
|
India,vegan low income |
27 |
43 |
|
Finland, before selenium fertilization |
26 |
67-69 |
|
Finland, after selenium fertilization |
56 |
67-69 |
|
Slovakia |
27 ± 8 |
70 |
|
United Kingdom, 1974 |
60 |
38 |
|
United Kingdom, 1985 |
43 |
38 |
|
United Kingdom, 1994 |
32 |
38 |
|
United Kingdom, 1995 |
33 |
46 |
|
Italy |
41 |
64 |
|
Germany |
38-48 |
71 |
|
France |
47 |
72 |
|
India, conventional diet |
48 |
43 |
|
United States, all |
80 ± 37 |
73 |
|
Males |
90 ± 14 |
73 |
|
Females |
74 ± 12 |
73 |
|
Canada |
98-224 |
74 |
|
United States, seleniferous area |
216 |
64 |
|
Venezuela |
80-500 |
75 |
|
China, seleniferous area |
1338 |
64 |
Table 51
Representative mean serum selenium concentration (µmol/l) in specific studiesa
|
Pathologic investigations |
Sample Serum Selenium
Concentrationsb |
|
Keshan disease (China) |
0.15-0.25 |
|
Kaschin-Beck disease (China) |
0.22 ± 0.03 |
|
Myxoedematous cretins (Zaire) |
0.26 ± 0.12 |
|
HIV and AIDSc |
0.36-0.54 |
|
Data for normal subjects |
|
|
Serbia and Croatia |
0.63-0.85 |
|
Bulgaria |
0.66-0.72 |
|
New Zealand |
0.69 |
|
Hungary |
0.71 ± 0.13 |
|
Norway |
1.52-1.69 |
|
United States, Maryland |
1.69-2.15 |
|
United States, South Dakota |
2.17-2.50 |
|
Proposed reference ranges for healthy subjects |
0.5-2.5 |
|
|
0.67- 2.04 |
Levander (84) convincingly illustrated the impracticability of assessing selenium requirements from input-output balance data because the history of selenium nutrition influences the proportion of dietary selenium absorbed, retained, or excreted. The changing equilibria when selenium intake is varied experimentally yield data which are of limited value for estimating minimal requirements. Examples are cited of estimates of selenium requirement for adults of 7.4 and 80 µg/day derived from Chinese and United States studies, respectively. Such discrepancies reflect differences in the usual daily selenium intakes of the experimental subjects and the extent to which this was changed experimentally. This situation, not unique to selenium, emphasises the importance of basing requirement estimates on functional criteria derived from evidence describing the minimum levels of intake which, directly or indirectly, reflect the normality of selenium-dependent processes.
New opportunities for the development of biochemical indexes of selenium adequacy such as those listed in Table 47 have yet to be exploited. Until this is done, the most suitable alternative is to monitor changes in the relationship of serum selenium to dietary selenium supply, taking advantage of its relatively constant proportionality to the fraction of serum selenium in functionally significant GSHPx (85).
A detailed review of 36 reports describing serum selenium values in healthy subjects indicated that they ranged from a low of 0.52 µmol/l in Serbia to a high of 2.5 µmol/l in Wyoming and South Dakota in the United States (76). It was suggested that mean values within this range derived from 7502 apparently healthy individuals should be regarded tentatively as a standard for normal reference. This survey clearly illustrated the influence of crop management on serum selenium level; in Finland and New Zealand, selenium fortification of fertilisers for cereals increased serum selenium from 0.6 to 1.5 µmol/l. A summary of these data in Table 51 also includes representative mean serum selenium values within the range of 0.15-0.55 µmol/l reported for specific diseases known to be associated with disturbances in selenium nutrition or metabolism. These include reports from studies of Keshan disease, Kaschin-Beck disease, and specific studies of cretinism, hypothyroidism, and HIV and AIDS where clinical outcome or prognosis has been related to selenium status.
This report and the report by the World Health Organization
(WHO), FAO, and IAEA (86) use virtually identical approaches to derive
their estimates of basal requirements for selenium
(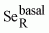 ). As yet there are no published
reports suggesting that these basal estimates using Se or GSHPx activity as
criteria of adequacy are invalid. Some modification is necessary however to
estimate population minimum intakes with adequate allowance for the variability
(CV) associated with estimates of the average selenium intakes from the typical
diets of many communities. In the WHO-FAO-IAEA report (86) a CV of 16
percent was assumed for the selenium conventional diets and 12.5 percent for the
milk-based diets of infants to limit the risks of inadequacy arising from
unexpectedly low selenium contents. More recent studies suggest that the
variability of selenium intake from diets for which the selenium content has
been predicted rather than measured may be substantially greater than estimated
previously (Table 49 [47] and Table
50).
). As yet there are no published
reports suggesting that these basal estimates using Se or GSHPx activity as
criteria of adequacy are invalid. Some modification is necessary however to
estimate population minimum intakes with adequate allowance for the variability
(CV) associated with estimates of the average selenium intakes from the typical
diets of many communities. In the WHO-FAO-IAEA report (86) a CV of 16
percent was assumed for the selenium conventional diets and 12.5 percent for the
milk-based diets of infants to limit the risks of inadequacy arising from
unexpectedly low selenium contents. More recent studies suggest that the
variability of selenium intake from diets for which the selenium content has
been predicted rather than measured may be substantially greater than estimated
previously (Table 49 [47] and Table
50).
Because balance techniques were shown to be inappropriate for determining selenium requirements, the WHO-FAO-IAEA report (86) presented requirement estimates based on of epidemiologic evidence derived from areas of China endemic or non-endemic for Keshan disease (18). These comprehensive biochemical and clinical studies showed that Keshan disease did not occur in regions where the mean intake of selenium by adult males or females was greater than 19.1 or greater than 13.3 µg/day, respectively. Although these intakes were sufficient to eliminate clinical evidence of myocarditis and other signs of Keshan disease, other studies showed that they were inadequate to restore erythrocyte or plasma selenium concentrations or GSHPx activities to levels indicative of reserves.
Studies with adult male subjects initially of low selenium
status given a carefully monitored diet providing selenium at 11 µg/day
together with supplements of selenomethionine given orally which provided 0, 10,
30, 60, or 90 µg/day. Starting at frankly deficient levels, total daily
selenium intakes of above 41 µg/day were found sufficient increase plasma
GSHPx substantially and to saturate plasma activity in 60-kg male subjects
within 5-8 months. It was estimated that satisfactory levels of plasma selenium
(>80 µmol/l) and of GSHPx (>0.3 mmol NADPH oxidized/min/l;
approximately two-thirds of plasma saturation activity) indicative of adequate
selenium reserves would be attained after intakes of approximately 27
µg/day by 65-kg male subjects (86). Such criteria satisfying the
definition of average normative requirements for selenium
( ) have been used as the basis for
calculating recommended nutrient intake (RNI) values in this report after
interpolating estimates of average requirements by allowing for differences in
weight and basal metabolic rate of age groups to up to 65 years and adding a 25
percent increase (2 x assumed SD) to allow for individual variability in the
estimates of RNI (Table 52).
) have been used as the basis for
calculating recommended nutrient intake (RNI) values in this report after
interpolating estimates of average requirements by allowing for differences in
weight and basal metabolic rate of age groups to up to 65 years and adding a 25
percent increase (2 x assumed SD) to allow for individual variability in the
estimates of RNI (Table 52).
Table 52
Recommended nutrient intakes of selenium (µg/day)
|
|
|
Average normative requirement |
|
|
|
Age Group |
Assumed Weight |
|
|
RNIc, µg/day |
|
Infants and children |
|
|
|
|
|
0-6 months |
6 |
0.85 |
5.1 |
6 |
|
7-12 months |
9 |
0.91 |
8.2 |
10 |
|
1-3 years |
12 |
1.13 |
13.6 |
17 |
|
4-6 years |
19 |
0.92 |
17.5 |
22 |
|
7-9 years |
25 |
0.68 |
17.0 |
21 |
|
Adolescents |
|
|
|
|
|
Female,10-18 years |
49 |
0.42 |
20.6 |
26 |
|
Male, 10-18 years |
51 |
0.50 |
22.5 |
32 |
|
Adults |
|
|
|
|
|
Female,19-65 years |
55 |
0.37 |
20.4 |
26 |
|
Male, 19-65 years |
65 |
0.42 |
27.3 |
34 |
|
Female, 65+ years |
54 |
0.37 |
20.2 |
25 |
|
Male, 65+ years |
64 |
0.41 |
26.2 |
33 |
|
Pregnancy |
|
|||
|
2nd trimester |
28 |
|||
|
3rd trimester |
30 |
|||
|
Lactation |
|
|||
|
0-6 months post-partum |
35 |
|||
|
7-12 months post-partum |
42 |
|||
+ 2 x assumed standard error
(of 12.5 percent)The estimates of RNI for infants (Table 52) are compatible with estimates of the international reference range of the selenium content of breast milk (18.5 µg /l; Table 49), with data from an extensive international survey of breast milk selenium (WHO-IAEA [50]) and with WHO data (47) on the milk consumption of exclusively human-milk-fed infants in developed and developing countries. Data from the WHO-IAEA (50) survey from six countries suggest that the human milk from all countries met the RNI for infants aged 0-6 months. In two of six countries, Hungary and Sweden, the human milk selenium was marginal with respect to the RNI for infants aged 7-12 months.
Data from Germany (13, 88), Austria (12), the United States (89), and elsewhere suggest that infant formula may contain selenium in amounts insufficient to meet the RNI or recommended dietary allowance for infants. Lombeck et al. (13) in an extensive study showed that cow-milk-based formula could provide less than one-third of the selenium of human milk. Estimates of selenium intake by 2-month-old infants were 7.8 µg/day from formula compared with 22.4 µg/day from human milk. Levander (89) has suggested that infant formulas should provide a minimum of 10 µg/day and not more than 45 µg/day. This recommendation may well have been implemented judging from recent increases in the selenium content of infant formulas (61).
Selenium requirements during pregnancy and lactation
Data from balance experiments are not sufficiently consistent for defining the increase of selenium needed to support foetal growth and development during pregnancy. For this reason the European Union Scientific Committee for Food (90), the UK Committee on Medical Aspects of Food Policy (91), and the Netherlands Food and Nutrition Council (92) have suggested that the component of selenium needed for human pregnancy is obtained by an adaptive increase in the efficiency of absorption of dietary selenium rather than by an increased dietary demand.
Others, contesting this view, have attempted to predict the increase of dietary selenium needed for pregnancy by factorial estimation of the likely quantity of selenium incorporated into the tissues of the foetus (47, 86). Such estimates have assumed that the total products of conception amount to 4.6-6 kg lean tissue with a protein content of approximately 18.5-20 percent. If, as appears to be a reasonable assumption, the selenium content of this protein resembles that of a skeletal muscle, growth of these tissues could account for between 1.0 and 4.5 µg/day of selenium depending on whether the analyses reflect consumption of diets from a low-selenium (but non-pathogenic) environment (e.g., New Zealand [49, 52]) or from a region with relatively high selenium intakes (e.g., United States, Table 50 [73]). Typically such estimates have assumed an 80 percent absorption and utilisation of dietary selenium from which it would appear reasonable to estimate that allowing for a variability of estimates (CV 12.5 percent) an increase of 2 µg/day would be appropriate for the second trimester and 4 µg/day would be appropriate for the third trimester of pregnancy (Table 52).
As is evident from Table 49 the selenium content
of human milk is sensitive to changes in maternal dietary selenium. The increase
of maternal dietary selenium needed to meet requirements for lactation has been
estimated from the estimated RNI for infants aged 0-6 months and 7-12 months. It
is assumed that the selenium of maternal milk is used with an efficiency of 80
percent, and a SD of 12.5 percent is assumed. For the period 0-6 months it is
estimated that the infant must receive 6 µg/day from human milk. The
increase of maternal dietary selenium required to produce this will be 6 x
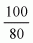 + (2 x SD) = 9 µg/day. The
corresponding increase needed to meet the infant RNI of 10 µg/day for ages
7-12 months will be 16 µg/day. Added to the non-pregnancy maternal RNI of
26 µg/day, the total RNI for lactation during the first 6 months
post-partum will be 35 µg/day and for months 7-12 will be 42 µg/day
(Table 52).
+ (2 x SD) = 9 µg/day. The
corresponding increase needed to meet the infant RNI of 10 µg/day for ages
7-12 months will be 16 µg/day. Added to the non-pregnancy maternal RNI of
26 µg/day, the total RNI for lactation during the first 6 months
post-partum will be 35 µg/day and for months 7-12 will be 42 µg/day
(Table 52).
As implied by the data in Tables 48, 49 and 50, agricultural growing practices, geologic factors, and social deprivation enforcing the use of an abnormally wide range of dietary constituents may significantly modify the variability of dietary selenium intakes. If accumulated experience suggests that the CV of selenium intake may be 40 percent or more and tabulated rather than analysed data are used to predict the dietary content of selenium, the selenium allowance may have to be increased accordingly or assessed by using the WHO-FAO-IAEA technique (86).
Upper tolerable nutrient level for selenium
A comprehensive account of the clinically significant biochemical manifestations of chronic and acute intoxication from selenium arising from high concentrations in food, drinking water, and the environment were published jointly by WHO and the United Nations Environment Programme and the International Labour Organisation (80). This report (44) stresses that the signs and symptoms of human overexposure to selenium are not well defined. Common clinical features are hair loss and structural changes in the keratin of hair and of nails, the development of icteroid skin, and gastrointestinal disturbances (93, 94). An increased incidence of nail dystrophy has been associated with consumption of high-selenium foods supplying more than 900 µg/day. These foods were grown in selenium-rich (seleniferous) soil from specific areas in China (95). A positive association between dental caries and urinary selenium output under similar circumstances was reported (96, 97).
Sensitive biochemical markers of impending selenium intoxication have yet to be developed. In their absence it is suggested that the upper tolerable nutrient intake level (UL) for selenium should be set, provisionally, at 400 µg/day for adults. It is noteworthy that a maximum tolerable dietary concentration of 2 mg/kg dry diet was suggested for all classes of domesticated livestock and has proved satisfactory in use (98). This suggests that the proposed UL of 400 µg/day for human subjects provides a fully adequate margin of safety. The UL for children and for pregnant or lactating women has yet to be determined.
Comparison with previous estimates
Compared with WHO-FAO-IAEA (86), US (87), UK (91), and European Union (90) recommendations, the present proposals represent a significant decrease in the suggested need for selenium. Reasons for this are:
the need to derive recommendations which are applicable for a proportionally lower weight range than for most Western and developed communities;The reduced estimates presented in this report are physiologically justifiable and will only give rise to concern if there are grounds for serious uncertainty as to the predictability of dietary selenium intake.the decision, also accepted by WHO, FAO, and IAEA (86), that it is neither essential nor desirable to maintain selenium status at a level which fully saturates blood GSHPx activity when, on current evidence, this is not an advantage for health; and
the decision to present estimates as RNIs which, although including an allowance for individual variability, do not provide for the possibility that foods may often differ widely in selenium content according to their geographic sources.
Food commodity inputs are changing rapidly and in some instances, unpredictably. Under most circumstances it will be unreasonable to expect that the often marked influence of geographic variability on the supply of selenium from cereals and meats can be taken into account. Changes in trade patterns with respect to the sources of cereals and meats are already having significant influences on the selenium nutrition of consumer communities (38, 66). Such evidence fully justifies the warning to allow for a high intrinsic variability of dietary selenium content when estimating selenium requirements of populations for which the principal sources of this microelement are unknown.
Relationships between selenium status and pathologically relevant biochemical indexes of deficiency merit much closer study with the object of providing more reliable and earlier means of detecting a suboptimal status.
Indications that a suboptimal selenium status may have much wider significance in influencing disease susceptibility must be pursued. Such studies must cover both the impact of selenium deficiency on protection against oxidative damage during tissue trauma and its genetic implication for viral virulence.
We lack knowledge of the influence of soil composition on the selenium content of cereals and animal tissues. Chinese experience with respect to the dramatic influence of soil iron and low pH on selenium availability may well be relevant to extensive tracts of lateritic soils in Africa and elsewhere. There are grounds for the belief that factors in common for selenium and iodine may influence their supply and availability from soils into the human food chain. FAO should be encouraged to develop studies relevant to the influence of soil conditions on the supply of these two metabolically interdependent elements which affect human health.
The early detection of selenium toxicity (selenosis) is hindered by a lack of suitable biochemical indicators. Effective detection and control of selenosis in many developing countries awaits the development of improved specific diagnostic techniques.
1. Levander, O.A. 1986. Selenium. In: Trace elements in human and animal nutrition 5th edn. Mertz, W. ed. p 209-279. Orlando, Florida 209-279. Academic Press Inc.
2. Arthur, J.R. & Beckett, G.J. 1994. Neometabolic roles for selenium. Proc. Nutr. Soc. 53: 615-624.
3. Ge, K. & Yang, G. 1993. The epidemiology of selenium deficiency in the etiological study of endemic diseases in China. Am. J. Clin. Nutr., Supplement 57: 259S-263S.
4. Arthur, J.R., Bermano, G., Mitchell, J.H. & Hesketh, J.E. 1996. Regulation of selenoprotein gene expression and thyroid hormone metabolism. Biochem. Soc. Trans., 24: 384-388.
5. Howie, A.F., Arthur, J.R., Nicol, T., Walker, S.W., Beech, S.G. & Beckett, G.J. 1998. Identification of a 57-kilodalton selenoprotein in human thyrocytes as thioredoxin reductase. J. Clin. Endocrino.l Metab., 83: 2052-2058.
6. Mairrino, M., Thomas, J.P., Girotti, A.W. & Ursini, F. 1991. Reactivity of phospholipd hydroperoxide glutathione peroxidase with membrane and lipoprotein lipid hydroperoxides. Free. Radic. Res. Commun. 12: 131-135.
7. Arthur, J. 1997. Selenium biochemistry and function. In: Trace Elements in Man and Animals - 9. Proceedings of the Ninth International Symposium on Trace Elements in Man and Animals. Fischer, P.W.F., L'Abbe, M.R., Cockell, K.A., Gibson, R.S. eds. p. 1-5.Ottawa, Canada, NRC Research Press.
8. Reilly, C. 1996. Selenium in food and health. London, Blackie Academic and Professional.
9. Anikina, L.V. 1992. Selenium-deficient cardiomyopathy (Keshan disease). In: Fifth International Symposium on Selenium in Biology and Medicine. Burk, R.F. ed. Vanderbilt University, Nashville, TN p 122.
11. Brennan, M.F. & Horwitz, G.D. 1984. Total parenteral nutrition in surgical patients. Advances in Surgery, 17: 1-7. van RiJ., A.M., Thompson, D., McKenzie, J.M. & Robinson, M.F. 1979. Selenium deficiency in total parenteral nutrition. Am. J. Clin. Nutr., 32: 2076-2085.
12. Rossipal, E. & Tiran, B. 1995. Selenium and glutathione peroxidase levels in healthy infants and children in Austria and the influence of nutrition regimens on these levels. Nutrition, 11 (5 suppl): 573-575.
13. Lombeck, I., Kasperek, K., Bonnermann, B. et al. 1975. Selenium content of human milk, cows milk and cows milk infant formulas. Eur. J. Pediatr., 139-145.
14. Okada, A., Takagi, Y., Nezu, R., Sando, K. & Shenkin, A. 1995. Trace element metabolism in parenteral and enteral nutrition. Nutrition, 11; 106-113.
15. Collip, P.J. & Chen, S.Y. 1981. Cardiomyopathy and selenium deficiency in a two year old girl. N. Engl. J. Med., 304: 1304-1305.
16. Lombeck, I., Ebert, K.H., Kasparek, K. et al. 1984. Selenium intake of infants and young children, healthy children and dietetically treated patients with phenylketonnria. Eur. J. Pediatr., 143: 91-102.
17. Levander, O.A. & Beck, M.A. 1997. Interacting nutritional and infectious ecologies of Keshan Disease. Biol. Trace Elem. Res., 56: 5-21.
18. Yang, G-Q., Zhu, L-Z., Liu, S-J., Gu, L-Z., Qian, P-C., Huang, J-H. & Lu, M-D. 1984. Human selenium requirements in China. In: Selenium in Biology and Medicine. Combs, G.R., Spallholz, J.E., Levander, O.A., Oldfield, J.E. eds. p.589-607. New York, AVI Van Nostrand.
19. Johnson, C.C., Ge, X., Green, K.A. & Liu, X. 1996. Studies of selenium distribution in soil, grain, drinking water and human hair samples from the Keshan Disease belt of Zhangjiakou district, Henei Province, China. Technical Report WC/96/52. Nottingham, UK, Overseas Geology Series, British Geological Survey.
20. Li, J-Y., Ren, S-X., Cheng, D-Z., Wan, H-J., Liang, S-T., Zhang, F-J. & Gao, F-M. 1984. Distribution of selenium in the microenvironment related to Kaschin-Beck disease. In: Selenium in Biology and Medicine. Combs, G.F., Spallholz, J.E., Levander, O.E., Oldfield, J.E. eds. p.911-925. New York, AVI Van Nostrand.
21. Beck, M.A. 1998. The influence of antioxidant nutrients on viral infection. Nutr. Revs., 56: S140-S146.
22. Baum, M.K. & Shor-Posner, G. 1998. Micronutrient status in relationship to mortality in HIV-1. Nutr. Revs., 56: S135-S139.
23. Baum, M.K., Shor-Posner, G., Lai, S.H., Zhang, G.Y., Fletcher, M.A., Sanberlich, H. & Page, J.B. 1997. High risk of HIV-related mortality is associated with selenium deficiency. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol., 15: 370-374.
24. Cirelli, A., Ciardi, M. & DeSimone, C. 1991. Serum selenium concentration and disease progress in patients with HIV infection. Clin. Biochem., 24: 211-214.
25. Dworkin, B.M. 1994. Selenium deficiency in HIV infection and the acquired immunodeficiency syndrome (AIDS). Chem. Bio. Interact., 91: 181-186.
26. Taylor, E.W., Nadimpalli, R.G. & Ramanthan, C.S. 1997. Genomic structures of viral agents in relation to the synthesis of selenoproteins. Biol. Trace Elem. Res., 56: 63-91.
27. Zazzo, J.F., Chalas, A. & LaFont. 1988. Is monobstructive cardiomyopathy in AIDS a selenium deficiency-related disease. J. Parenteral Enteral Nutr., 12: 537-538.
28. Kavanaugh-McHugh, A.L., Ruff, A. & Pearlman, A. 1991. Selenium deficiency and cardiomyopathy in acquired immunodeficiency syndrome. J. Parenteral Enteral Nutr., 15: 347-349.
29. Ramanathan, C.S. & Taylor, E.W. 1997. Computational genomic analysis of Hemorrhagic viruses; viral selenoproteins as a potential factors in pathogenesis. Biol. Trace Elem. Res., 56: 93-106.
30. Serfass, R.E. & Ganther, H.E. 1975. Defective microbial activity in glutathione peroxidase deficient neutrophils of selenium deficient rats. Nature, 225: 640-641.
31. Boyne, R. & Arthur, J.R. 1986. The response of selenium deficient mice to Candida albicans infection. J. Nutr., 116: 816-822.
32. Ip, C. & Sinha, D.K. 1981. Enhancement of mammary tumorigenesis by dietary selenium deficiency in rats with a high polyunsaturated fat intake. Cancer Res., 41: 31-34.
33. Levander, O.A. 1987. A global view of human selenium nutrition. Annu. Rev. Nutr., 7: 227-250.
34. Birt, D.F., Pour, P.M. & Pelling, J.C. 1989. The influence of dietary selenium on colon, pancreas, and skin tumorigenesis. In: Wendel A., ed. Selenium in biology and medicine. p. 297-304. Berlin, Springer-Verlag.
35. Arthur, J.R., Nicol, F. & Beckett, G.J. 1993. Selenium deficiency thyroid hormone metabolism and thyroid hormone deiodinases. Am. J. Clin. Nutr., Supplement, 57: 236S-239S.
36. Corrilain, B., Contempre, B., Longombe, A.O., Goyens, P., Gervy-Decoster, C., Lamy, F., Vanderpas, J.B. & Dumont, J.E. 1993. Selenium and the thyroid: how the relationship was established. Am. J. Clin. Nutr., Supplement 57: 244S-248S.
37. Olivieri, O., Girelli, D., Stanzial, A.M., Rossi, L., Bassi, A. & Corrocher, R. 1996. Selenium, zinc and thyroid hormones in healthy subjects. Low T3/T4 ratio in the elderly is related to impaired selenium status. Biol. Trace Elem. Res., 51: 31-41.
38. MacPherson, A., Barclay, M.N.J., Scotts, R. & Yates, R.W.S. 1997. Loss of Canadian wheat imports lowers selenium intake and status of the Scottish population. In: Trace Elements in Man and Animals -Proceedings of 9th International Symposium on Trace Elements in Man and Animals. Fischer, P.W.F., L'Abbe, M.R., Cockell, K.A., Gibson, R.S. p. 203-205. eds. Ottawa, Canada, NRC Research Press.
39. Vanderpas, J.B., Contempre, B., Duale, N.L. & Deckx, H. 1993. Selenium deficiency mitigates hypothyroxinimia in iodine deficient subjects. Am. J. Clin. Nutr., 57 Suppl: 271S-275S.
40. Contempre, B., Le Moine, O., Dumont, J.E., Denef, J-F. & Many, M.C. 1996. Selenium deficiency and thyroid fibrosis. A key role for macrophages and TGF-á. Mol. Cell. Enyzmol., 124: 7-15.
41. Ma, T., Guo, J. & Wang, F. 1993. The epidemiology of iodine deficiency diseases in China. Am. J. Clin. Nutr., Supplement, 57: 264S-266S.
42. Contempre, B., Many, M.C., Duale, G.L., Denef, J.F. & Dumont, J.E. 1996. Selenium and iodine in thyroid function: the combined deficiency in the etiology of the involution of the thyroid leading to myxoedematous cretinism. In: Thyroid and Trace Elments. 6th Thyroid Symposium. p. 35-39. Eds. Browerman, L.E., Kohsle, J., Eber, O., Langsteger, W. Graj-Eggenberg: Barmhersige Brudes.
43. Mahalingam, T.R., Vijayalakshni, S., Krishna, & Prabhu, R. 1997. Studies on some trace and minor elements in blood. A survey of the Kalpakkan (India) population. Part III: Studies on dietary intake and its correlation to blood levels. Biol. Trace Elem. Res., 57: 223-238.
44. Levander, O.A. 1987. A global view of human selenium nutrition. Annu. Rev. Nutr., 7: 227-250.
45. Varo, P. & Koivistoinen, P. 1980. Mineral element composition of Finnish foods. XII General discussion and nutritional evaluation. Acta Agric. Scand., Supplement No. 22; 165-171.
46. MAFF. 1997. UK Dietary Intake of Selenium. MAFF Food Surveillance Information Sheet: No. 126. London, MAFF/HMSO.
47. World Health Organization. 1998. Complementary Feeding of Young Children in Developing Countries \\WHO/NUT/98.1. Geneva, WHO.
48. Robberecht, H., Benemariya, H. & Dellstra, H. 1995. Daily dietary intake of copper, zinc and selenium of exclusively breast fed infants of middle-class women in Burundi, Africa. Biol. Trace Elem. Res., 49: 151-159.
49. Williams, M.M.F. 1983. Selenium and glutathione peroxidase in mature human milk. Proceedings of the University of Otago Medical School, Dunedin, 61: 20-21.
50. World Health Organisation/International Atomic Energy Agency. 1989. Minor and Trace Elements in Milk. Geneva, WHO.
51. Kumpulainen, J., Vuori, E. & Kuitunen, P. 1983. Longetudinal study on the dietary selenium intake of exclusively breast fed infants and their mothers in Finland. Int. J. Vit. Nutr. Res., 53: 420-426.
52. Millar, K.R. & Sheppard, A.D. 1972. a-Tocopherol and selenium levels in human and cow's milk. NZ J. Sci., 15: 3-15.
53. Sumar, S., Kondza, B. & Foster, L.H. 1997. Selenium levels in infant formulae and breast milk from the United Kingdom: a study of estimated intakes. In: Trace Elements in Man and Animals - 9. Proceedings of the Ninth International Symposium on Trace Elements in Man and Animals. Fischer, P.W.F., L'Abbe, K.A., Cockell, K.A. Gibson, R.S. p.282-283.Ottawa, Canada, NRC Research Press.
54. Cumming, F.J., Fardy, J.J. & Woodward, D.R. 1992. Selenium and human lactation in Australia: milk and blood selenium levels in lactating women and selenium intake of their breast-fed infants. Acta Paediatrica, 81: 1058-1061.
55. Levander, O.A., Moser, P.B. & Morris, V.C. 1987. Dietary selenium intake and selenium concentrations of plasma, erythrocytes, and breast milk in pregnant and postpartum lactating and nonlactating women. Am. J. Clin. Nutr., 46: 694-698.
56. Shearer, T.R. & Hadjimarkos, D.M. 1975. Geographic distribution of selenium in human milk. Arch. Environ. Health, 30: 230-233.
57. Lombeck, I., Kasparek, & Bonnerman, B. 1975. Selenium content of human milk, cow's milk and cow's milk infant formulaes. Eur. J. Paediatr.. 129: 139-145.
58. Iyengar, V. & Wooittiez, J. 1988. Trace elements in human clinical specimens: evaluation of literature to identify reference values. Clin. Chem., 34: 474-481.
59. Darlow, B.A., Inder, T.E., Sluis, K.B., Nuthall, G. et al. 1995. Selenium status of New Zealand infants fed either a selenium supplemented or a standard formula. J. Paediatr. Child. Health, 31: 339-344.
60. Smith, A., Picciano, M.F. & Milner, J.A. 1982. Selenium intakes and status of human milk formula fed infants. Am. J. Clin. Nutr., 35: 521-526.
61. Lonnerdal, B. 1997. Effects of milk and milk components on calcium, magnesium, and trace element absorption during infancy. Physiol. Revs., 77: 643-669.
62. Yang, G., Wang, S., Zhou, R. & Sun, S. 1983. Endemic sleenium intoxication of Humans in China. Am. J. Clin. Nutr., 37: 872-881.
63. Luo, X.M., Yang, C.L., Wei, H.J., Liu, X. & Qixo, C.H. 1984. Selenium intake and metabolic balance in 10 men consuming self-selected diets in a selenium-deficient area of Hebei Provence, PR China. FASEB J., 43: Abstr. 1097.
64. Parr, R.M., Crawley, H., Aldulla, M., Iyengar, G.V. & Kumpulainen, J. 1992. Human dietary itnakes of trace elements: A global literature survey mainly for the period 1970-1991. I Data listings and sources of information. NAHRES 12, Vienna, International Atomic Energy Agency.
65. Robinson, M.T. & Thomason, C.D. 1984. Status of the food supply and residents of New Zealand. In: Combs GF (ed) Selenium in Biology and Medicine. p. 631-644. New York, AVI Van Nostrand.
66. Abdulla, M.A., Behbehani, A. & Dashti, H. 1989. Dietary intake and bio-availability of trace elements. Biol. Trace Elem. Res., 21: 173-178.
67. Koivistoinen, P. & Varo, P. 1987. Selenium in Finnish food. In: Selenium in Biology and Medicine. Cambs. J.F., Spallholz, J.E., Lavander, O.A., p. 645-651. New York, Oldfield. Van Nostrand.
68. Mutanen, M. 1985. Comparison of chemical analysis and calculation emthod in estimating selenium content of Finnish diets. Nutr. Res., 5: 693-697.
69. Mutanen, M. 1984. Dietary intake and sources of selenium in young Finnish women. Hum. Nutr: Appl. Nutr., 38A: 265-269.
70. Kadrabova, J., Madaric, A. & Ginter, E. 1998. Determination of the daily selenium intake in Slovakia. Biol. Trace Elem. Res., 61: 277-286.
71. Oster, O. & Prellwitz, W. 1989. The daily dietary selenium intake of West German adults. Biol. Trace Elem. Res., 20: 1-14.
72. Simonoff, M. & Simonoff, G. 1991. Le Selenium et la Vie. Paris, Masson.
73. Levander, O.A. & Morris, V.C. 1984. Dietary selenium levels needed to maintain balance in North American adults consuming self-selected diets. Am. J. Clin. Nutr., 39: 809-815.
74. Thomson, J.N., Erdody, P. & Smith, D.C. 1975. Selenium in Canadian Foods and diets. J. Nutr., 105: 274-279.
75. Bratter, P, Bratter, N. & Gwlik, D. 1993. Selenium in Human monitors related to the regional dietary intake levels in Venezuela. J. Trace Elem. Electrolyte Health Dis., 7: 111-112.
76. Alfthan, G. & Neve, J. 1996. Reference values for serum selenium in various areas evaluated according to the TRACY protocol. J. Trace Elem. Med. Biol., 10: 77-87.
77. Diplock, A.T., Contempre, B., Dumont, J., Bebe, N. & Vanderpas, J. 1997. Interaction of selenium and iodine deficiency diseases. In: Trace Elements in Man and Animals - 9. Proceedings of the ninth International Symposium on Trace Elements in Man and Animals. p. 63-68. Fischer, P.W.F. L'Abbe, M.R., Cockell, K.A., Gibson, R.S. Ottawa, ON, NRC Research Press.
78. Diplock, A.T. 1993. Indexes of selenium status in Human populations. Am. J. Clin. Nutr., Supplement 57: 256S-258S.
79. Versieck, J. & Cornelis, R. 1989. Trace elements in Human plasma or serum. Boca Raton, CRC Press.
80. WHO. 1987. Selenium, Geneva, World Health Organization. (Environmental Health Criteria, 58).
81. Patterson, B.H., Zech, L.A., Swanson, C.A. & Levander, O.A. 1993. Kinetic modelling of selenium in Humans using stable isotope tracers. J. Trace Elem. Electrolyte Health Dis. 7: 117-120.
82. Mutanen, M. Bio-availability of selenium. Ann. Clin. Res.,1986; 18: 48-54.
83. Levander, O.A. 1983.Bio-availability of selenium to Finnish men as assessed by platelet glutathione peroxidase activity and other blood parameters. Am. J. Clin. Nutr., 37: 887-897.
84. Levander, O.A. 1988. The global selenium agenda. In: Trace Elements in man and animals - 6. Proceeding of the 6th International Symposium on Trace Elements in Man and Animals. Hurley, L.S., Keen, C.L., Lonnerdal, B., Rucker, R.B. eds. New York Plenum Press Inc.
85. Gu, Q-P., Xia, Y-M., Ha, P-C., Butler, J.A. & Whanger, P.D. 1998.Distribution of selenium between plasma fractions in guinea pigs and Humans with various intakes of selenium. J. Trace Elem. Med. Biol., 12: 8-15.
86. WHO/FAO/IAEA. 1996. Trace elements in Human nutrition and health. Geneva. World Health Organization.
87. National Research Council, Food and Nutirtion Board. 1988. Recommended Dietary Allowances, 10 edition. Washington, D.C. US National Academy Press.
88. Lombeck, I., Kaspereck, K., Harbisch, H.D., Feinendegen, L.E. & Bremer, H.J. 1997. The selenium status of healthy children. I. Serum selenium concentration at different ages; activity of glutathione peroxidase of erythrocytes at different ages; selenium content of food of infants. Eur. J. Paediatr., 125: 81-88.
89. Levander, O.A. 19893 Upper limit of selenium in infant formulas. J. Nutr., 119: 1869-1871.
90. Scientific Committee for Food. 1993. Nutrient and Energy Intakes for the European Community. Report of the Scientific Committee for Food, Thirty First Series. Office for Official Publications of the European Communities, Brussels.
91. Department of Health. 1991. Dietary Reference Values for Food Energy and Nutrient Intakes for the United Kingdom. Report on Health and Social Subjects No.41.London, HMSO.
92. Netherlands Food and Nutrition Council. 1989. Recommended Dietary Allowances. The Netherlands. The Hague.
93. Smith, M.I., Franke, K.W. & Westfall, B.B. 1936. The selenium problem in relation to publish health. US Public Health Report., 51: 1496-1505.
94. Smith, M.I. & Westfall, B.B. 1937. Further field studies on the selenium problem in relation to public health. US Public Health Report, 52: 1375-1384.
95. Yang, G., Wang, S., Zhou, R. & Sun, S. 1983. Endemic selenium intoxication of Humans in China. Am. J. Clin. Nutr., 37: 872-881.
96. Hadjimarkos, D.M. 1973. Selenium in relation to dental caries. Food Cosmetic Toxicol., 11: 1083-1095.
97. Hadjimarkos, D.M., Storveik, C.A. & Renmert, L.T. 1952. Selenium and dental casies. An investigation among school children of Oregon. J. Paediatr., 40: 451-455.
98. National Research Council. 1980. Mineral Tolerance of Domestic Animals. Commission on Natural Resources., Washinton, D.C. National Academy of Sciences.1.
![]()
![]()
![]()