![]()
![]()
![]()
Il convient de souligner ici que l'emploi des données de la pêche commerciale pouvant être effectées de biais a divers niveaux. Ceci aura une incidence sur les estimations des paramètres de croissance et de mortalité. Dans notre cas, une partie seulement de la population est représentée dans les captures commerciales, probablement en raison de phénomènes liés au comportement, (distribution, réaction des poissons à la lumière et a l'engin de pêche variables avec l'age) et a la technique de pêche.
Le Limnothrissa miodon du lac Kivu est un espèce à durée de vie courte, deux années a peu près, caractérisée par un taux de croissance élevé (FIGURE 3). Diverses estimations de croissance obtenues dans plusieurs études pour des années différentes, ont conduit à des valeurs semblables (TABLEAU 1). La croissance semble différente pour les males et les femelles qui atteignent. une taille plus grande et probablement un age plus éleve (FIGURE 9). La croissance de cette espèce dans le lac Kivu est semblable à celle observée dans le lac Tanganyika, son environnement d'origine, mais très différente de celle observée dans les lacs artificiels où elle a été introduite (Kariba et Cahora Bassa). La complexité des facteurs limitant la croissance n'est pas bien connue, mais il est évident que L. miodon présente une bonne capacité d'adaptation à divers milieux lacustres (MARSHALL, 1987, MANNINI, 1990).
TABLEAU 1 : Paramètres de croissance de Limnothrissa miodon dans la période 1980–1981
| Loo (mm) | K (annuel) | Année | Sources |
| 145 | 1.20 | 1980 | SPLIETHOFF and DE IONGH (1983) |
| 149 | 1.30 | 1980 | MANNINÏ (1990) |
| 154 | 1 .31 | 1983 | " " |
| 138 | 1 .32 | 1986 | " " |
| 144 | 1 .28 | 1989 | " " |
| 14S | 1.20 | 1990 | Cette étude |
La valeur élevée de mortalité naturelle obtenue avec l'expression de Pauly est en accord avec ce que l'on attend d'un espèce pélagique tropicale a croissance rapide. La prédation sur le L. miodon du lac Kivu due au cannibalisme, à Clarias garipenus (DECEUNINK, 1990) et aux oiseaux aquatiques est probablement bien inférieure à celle observee dans la plupart des autres stocks. L'expression de PAULY qui ne prend en compte que L∞K et la température de surface ne permet pas de tenir compte d'une telle situation où la prédation est faible.
L'estimation de mortalité totale “Z” des population de L. miodon des lacs Kivu. Tanganyika et Kariba en utilisant la courbe de capture convertie, donnent généralement des valeurs élevées (MARSHALL, 1987 MARSHALL in MACHENA, 19S8, MUNYANDONERO, 1989, MANNINI, 1990). Les estimation de “F” qui en découlent sont aussi surestimées car F = Z-M. Les distributions de fréquences de tailles du lac Kivu obtenues a partir des données commerciales ne sont pas totalement représentatives de la distribution réelle de la population et l'analyse de la courbe de capture ne peut être employée. La pente prononcee sur la partie droite de la courbe de distribution de fréquence conduit inévitablement à des estimations élevées de “Z” qui ne sont pas necessairement dues à une mortalité drastique mais à l'absence de poissons âgés dans les captures. Ceci pouvant être dû a une émigration des poissons âgés hors de la zone de pêche ou a un évitement des poissons les plus âgés à l'engin de capture. Des taux de mortalité relativement élevés sont cependant normaux chez L. miodon comme chez tous les petits pélagiques.
L'analyse de cohorte de JONES montre clairement qu'en 1990, seuls les poissons de taille comprise entre 90 et 110 mm âgés de 10 à 14 mois (age relatif) font l'objet de la mortalité par pêche, le reste du stock étant. relativement peu exposé (FIGURE 4). Cette situation pourrait être le résultat du comportement des poissons (migrations) et des limitations de la technique de pêche. Les trimarans de pêches propulsés à la pagaie présentent des limitations évidentes, car ils ne peuvent s'éloigner trop de la côte pour des raisons de sécurité et les zones de pêche sont restreintes aux régions côtières.
On a invoqué la migration comme l'un des facteurs possibles, mais les phénomènes ne sont pas très bien connus. Il existe des preuves que les juvéniles passent les premiers deux a trois mois de leur vie dans les eaux côtières peu profondes jusqu'à 20 mètres (SPLIETHOFF et al., 1983, REUSSENS, 1988), ils occupent ensuite tout le lac et on pense que les grands individus reviennent vers la côte, ils trouvent la une nourriture plus abondante et deviennent les prédateurs de leurs propres larves.
La sélection due au maillage ne semble pas devoir être invoquée car on a pu remarquer dans des distributions de fréquences certaines années des poissons de petite taille a certaines périodes (MANNINI, 1990).
Le choix de la mortalité par pêche finale (Ft = mortalité par pêche du groupe le plus âgé) est avec le choix de “M” l'une des éléments critiques dans l'analyse des populations virtuelles (Analyse de cohortes de Jones) et on a dû avoir recours à une estimation arbitraire. La valeur très faible de “Ft” de 0,001 utilisée rend compte du fait que les individus les plus âgés ne sont pas soumis à la mortalité par pêche. Les captures commerciales ou ces poissons sont sous-representés en apportent la preuve. Le rapport M/K est de 1,5 soit compris entre les valeurs 0,8 et 2,5 considérées comme les plus probables (BEVERTON .et HOLÏ, in CSIRKE, 1988).
Les estimations de la mortalité totale peuvent être obtenues en utilisant les valeurs de “F” par classes d'age de l'analyse des cohortes de Jones et en considérant “M” constant pour tous les âges (TABLEAU 2). Il est bien, évident que “Z” n'est pas constant et qu'il peut varier de 1,80 à 2,75 au cours de la vie du poisson. Le fait d'utiliser une valeur de “M” constante dans cette méthode est critiquable (GULLAND, 1988) car elle affecte les estimations de “Z”.
TABLEAU 2: Mortalité par pêche (F) et totale (Z = M + F) par classe de taille.
| Longueur | F | Z | |
| 27,5 | 0,0004 | 1,8004 | |
| 32,5 | 0,0046 | 1,8045 | |
| 37,5 | 0,0171 | 1,8171 | |
| 42,5 | 0,0350 | 1,8350 | |
| 47,5 | 0,0431 | 1,8431 | |
| 52,2 | 0,0337 | 1,8337 | |
| 57,5 | 0,0323 | 1,8323 | |
| 62,5 | 0,0600 | 1,8600 | |
| 67,5 | 0,0917 | 1,8917 | |
| 72,5 | 0,1554 | 1,9554 | |
| 77,5 | 0,2626 | 2,0626 | |
| 82,5 | 0,2650 | 2,0650 | |
| 87,5 | 0,2040 | 2,0040 | |
| 92,5 | 0,4783 | 2,2783 | |
| 97,5 | 0,9482 | 2,7482 | |
| 102,5 | 0,6320 | 2,4320 | |
| 107,5 | 0,1053 | 1,9053 | |
| 112,5 | 0,0444 | 1,8444 | |
| 117,5 | 0,0167 | 1,8167 | |
| 122,5 | 0,0085 | 1,8085 | |
| 127,5 | 0,0019 | 1,8019 | |
| 132,5 | 0,0028 | 1,8028 | |
| 137,5 | 0,0010 | Ft* | 1,8010 |
Le rapport de la production sur la biomasse (P:B) est fonction du taux de mortalité quand la croissance des poissons suit l'équation de Von Bertallanfy et que la mortalité est de type exponentielle multiple (ALLEN, 1971). Le L. miodon du lac Kivu a une mortalité totale élevée, ce qui signifie que la population doit avoir un fort taux de renouvellement. Cette caractéristique semble commune aux petits pélagiques d'eaux douces tels que Stolothrissa tanganyicae du lac Tanganyika (COULTER, 1981). Selon CHAPMAN et VAN WELL (1978), un taux de mortalité élevé tel que celui de S. tanganyicae indique des possibilités d'exploitations élevées.
Le modèle de prédiction a long terme de Thompson et Bell suggère que le stocK de L. miodon est actuellement sous-exploité et que la situation présente est loin du rendement maximum soutenu MSY (FIGURE 5). Il est possible d'augmenter l'effort sans effets négatifs sur le stock, même si avec le schéma actuel de la pèche seule une partie du stock est exploitée, cette fraction exploitée est constituée d'individus d'un an (age relatif) qui, s'ils ne sont pas capturés, mourront de mort naturelle. A titre d'exemple, une augmentation de l'exploitation correspondant a un accroissement de l'effort actuel de 30% conduirait à une augmentation de 25%, des captures en induisant une diminution de 5% des PUE. Simplement par le fait que la pêcherie n'exploite qu'une partie de la biomasse exploitable, on peut supposer qu'un accroissement de l'effort conduirait à un accroissement des captures inférieur au niveau qui correspondrait a une mise en exploitation de la totalité de la biomasse. Le schéma d'exploitation actuel est représenté sur la FIGURE 10 qui montre la composante de biomasse qui contribue à la majorité des captures.
Ce scénario à long terme ne signifie pas bien sûr que le stock pourrait être pêché de façon extensive sans problèmes, mai:-simplement qu'au niveau actuel, le stock est sous-exploité et que ses caractéristiques biologiques permettent un niveau d'exploitation plus important.
Fig. 10. Tailles exploitées fit structure de ta biomasse de L. miodon
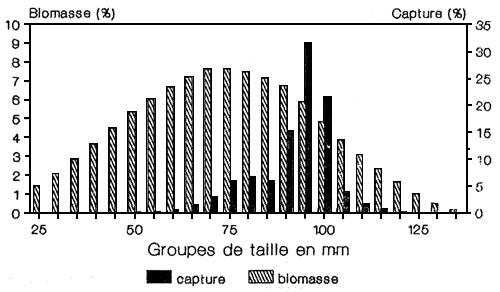
Il est important de noter que les chiffres de captures des modèles de prédiction n'ont qu'une valeur toute relative et ne sont donnés que pour indiquer les possibilités de développement. Les variations temporelles des paramètres de l'espèce, liées a celles de l'environnement pourront conduire a des modifications des potentialités de capture, en particulier compte tenu du fait que le modèle n'intègre qu'une seule variable; la mortalité par pêche. L'influence du phénomène “El nino” (STEELE et al., 1984) sur la pêcherie pélagique du Pérou montre bien la difficulté de prédire les cycles des clupéidé.
L'estimation actuelle du rendement maximum soutenu MSY de 9200 tonnes de Limnothrissa miodon pour la totalité du lac Kivu, correspond a 33 kg/Ha/an. On peut comparer avec les estimations de REYNTJENS (1981) qui a utilisé diverses méthodes (TABLEAU 3). La variabilité des estimations de REYNTJENS est due aux méthodes employées et au fait qu'elles sont ou non bien adaptées aux particularités du lac Kivu, l'auteur indique en conclusion une production de 35 a 55 kg/Ha/an. SPLIETHOFF et al. (1983) ont évalué la production a 42 kg/Ha/an correspondant à une rendement total de 11300 tonnes/an.
TABLEAU 3: Estimation du potentiel de production du lac Kivu
| Méthodes | Estimation en kg/ha/an | Production totale en t/an |
| IME | 34 – 55 | 9313 – 14800 |
| Production primaire | ||
| Méthode de Melack | 248 | 67000 |
| Méthode d'Oglesby | 11 | 3040 |
| Standing crop zooplancton | 55 | 14800 |
L'activité de reproduction de L. miodon est plus ou moins continue avec une saison de reproduction assez étendue. Chaque mois plus de 50% des individus échantillonnes sont en reproduction ou près de l'être. On note cependant deux pics d'activité maximum de mars a juin avec un maximum en avril et en septembre (FIGURE 7). Ces deux pics d'activité reproductrice ont été décrits de la même manière par MAHY (1982), SPLIETHOFF et al. (1983) et MUTAMBA et MULINDABIGWI (1990).
Aux lacs Tanganyika et Kivu, on pense que la saison de reproduction dépend de la disponibilité de nourriture (ELLIS, 1971, SPLIETHOFF et al., 1983). Au lac Kivu le pic de reproduction de septembre pourrait faire suite au maximum de Production planctonique de la fin de la Salson seche en août/septembre (VERBEKE, 1957), l'autre pic de reproduction en avril doit avoir une origine différente si l'on considère que la production planctonique est faible a cette période de l'année.
Le schéma de recrutement, en échelle ce temps relative, montre un maximum qui dure quatre mois et un deuxième pic moins important d'une durée de deux mois (FIGURE 11). 11 semble qu'il soit possible de lier les quatre mois du recrutement principal aux quatre mois (mars-juin) du pic de reproduction; de la même manière, le pic de reproduction de septembre peut être mis en relation avec le deuxième pic de recrutement. Dans les deux cas les pics de reproduction et de recrutement son séparés par trois mois .
Fig. 11. Schéma de recrutement (1990)
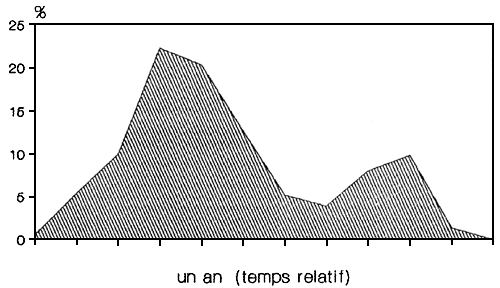
La dynamique de la reproduction et du recrutement sont des indicateurs de la stratégie de l'espèce et de sa capacité a réagir aux conditions de l'environnement. Il a aussi été observé que l'abondance des clupéides marins est complètement indépendante de la taille du stock des reproducteurs (MURPHY, 1977). On peut ainsi penser que la capture annuelle de L. miodon an niveau actuel de son intensité n'a pas grand effet sur la capture de l'année suivante. Les facteurs de l'environnement qui conduisent à des défauts de recrutements semblent avoir été les facteurs prépondérants dans les cas d'effondrement de stocks de clupéidés marins intensément exploités.
![]()
![]()
![]()