C.D. Allen
Craig D. Allen is with the United States Geological Survey, Fort Collins Science Center, Jemez Mountains Field Station, Los Alamos, New Mexico, United States.
An introduction to emerging global patterns of climate-induced forest mortality.
Forests, which today cover 30 percent of the world’s land surface (FAO, 2006), are being rapidly and directly transformed in many areas by the impacts of expanding human populations and economies. Less evident are the pervasive effects of ongoing climatic changes on the condition and status of forests around the world. Recent examples of drought and heat-related forest stress and dieback (defined here as tree mortality noticeably above usual mortality levels) are being documented from all forested continents, making it possible to begin to see global patterns. This article introduces these patterns and considers the possibility that many forests and woodlands today are at increasing risk of climate-induced dieback. A more comprehensive article (Allen et al., 2009) addresses this topic in considerably greater detail.
While climate events can damage forests in many ways ranging from ice storms to tornadoes and hurricanes, the emphasis here is on climatic water stress, driven by drought and warm temperatures.
The Earth’s climate is recognized to be undergoing significant human-caused changes, with global mean temperatures now outside the historic range of at least the past 1 300 years (IPCC, 2007). Markedly greater shifts in climatic patterns are projected for the coming decades in many regions, including much warmer temperatures and altered precipitation patterns that drive the availability of water to plants.
Since most of the world’s forests are found in areas where temperature, light or nutrients limit tree growth and productivity, recent global warming, changes in atmospheric composition (i.e. increased concentrations of nitrogen compounds and CO2 from massive societal emissions) and local increases in sunlight and precipitation have benefited the growth of many forests in recent decades, when and where water has not been limiting (Boisvenue and Running, 2006).
On the other hand, about one-third of the Earth’s land is currently too dry to support tree growth, and significant areas of forest and woodland grow in marginal climate zones where net primary vegetation productivity is strongly water limited (Boisvenue and Running, 2006). Forests in such semi-arid regions may display substantial growth declines or increases in mortality in response to droughts or warming temperatures (e.g. Peñuelas, Lloret and Montoya, 2001), as do tree species at the drier edges of their range of distribution (e.g. Jump, Hunt and Peñuelas, 2006).
Growth and mortality in wetter forests throughout the globe, however, from tropical moist forests to boreal systems, are also highly sensitive to drought (Clark, 2004; Nepstad et al., 2007; Soja et al., 2007). Temperate forests growing on productive sites may exhibit major growth declines, high levels of mortality and delayed multi-year effects from extreme drought and heat stress, as observed throughout Europe from the 2003 drought and heat wave (Ciais et al., 2005; Breda et al., 2006). Warmer temperatures alone can increase forest water stress independent of precipitation amount (Barber, Juday and Finney, 2000; Angert et al., 2005). As such, it is not apparent that any forests globally are safe from the impacts of drought.
Tree mortality commonly involves multiple, interacting factors, ranging from drought to insect pests and diseases, often making the determination of a single cause unrealistic. Abiotic stress factors, however, commonly underlie forest health problems, with climate stresses thought to be a primary factor triggering many extensive forest insect and disease outbreaks (Desprez-Loustau et al., 2006; Raffa et al., 2008).
Climate-induced water stress may directly cause tree mortality through short-term acute effects such as irreversible disruption of water columns within tree stems and leaves (cavitation). Tree species vary widely in their resistance and vulnerability to cavitation, a key determinant of drought resistance. When subject to water stress, trees minimize the risk of cavitation through stomatal closure, which reduces water loss and subsequent tension within the xylem. Stomatal closure comes at a cost, however, as it prevents CO2 diffusion into the foliage, thereby reducing photosynthesis. Chronic water stress over long periods will weaken and ultimately kill trees, either directly through carbon starvation or indirectly through the attacks of pests such as bark beetles which overwhelm the diminished defences of such chronically starved trees (McDowell et al., 2008). Climatic conditions also directly affect the population dynamics of forest insects and fungal pathogens (e.g. Hicke et al., 2006). Thus, some massive outbreaks of tree-killing forest insects may be attributed to climate drivers (Raffa et al., 2008). Regardless of the exact mechanism, dieback is often a non-linear process; it can emerge abruptly at a regional scale when climatic conditions exceed a tree species’ physiological thresholds of tolerance or trigger outbreaks of insect pests (Allen, 2007).
Many reports link increased forest mortality to various combinations of notable dry and/or hot conditions, such as drought in the tropics from severe El Niño events in 1988 and 1997–1998, the persistent warming and widespread drought over much of western North America since the 1990s, and the extreme heat wave and drought of summer 2003 in western Europe.
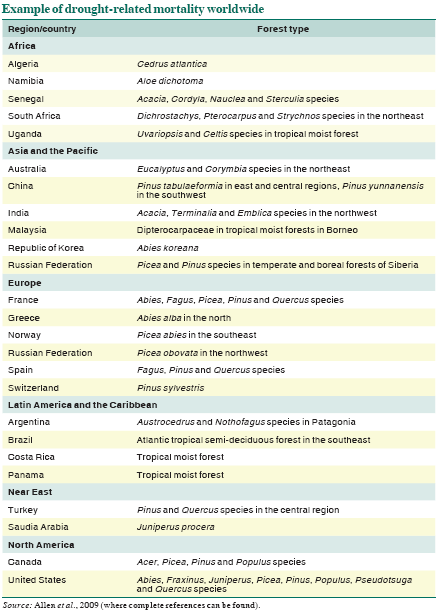
Forest mortality associated with drought has been documented recently from all wooded continents (Figure, p. 46) and from diverse forest types and climatic zones. Forest dieback is commonly reported near the geographic or elevational margins of a forest type or tree species (Jump, Hunt and Peñuelas, 2006), presumably near its historic thresholds of climatic suitability, where the most sensitive response to climate fluctuations would be expected.
Some examples of forest mortality driven by climatic water and heat stress since 1970, based on a global review of more than 120 documented examples (Allen et al., 2009), are presented in the Table. While forest dieback is commonly noted in semi-arid regions where trees are near the physiological limits of dryness for woody plant growth (e.g. Fensham,
Fairfax and Ward, 2009), it is clear that climate-induced drought and heat stress have the potential to cause forest dieback across a broad range of forest and woodland types around the world. Examples are particularly well documented from southerly parts of Europe (Peñuelas, Lloret and Montoya, 2001; Breda et al., 2006) and in temperate and boreal forests of western North America, where background mortality rates have increased rapidly in recent decades (van Mantgem et al., 2009) and widespread death of many tree species in multiple forest types has affected well over 10 million hectares since 1997 (Breshears et al., 2005; Raffa et al., 2008).

 |
Assessing the potential for, and consequences of, extensive climate-induced forest dieback is fundamentally important because trees grow relatively slowly but can die quickly. A 100-year-old tree may be killed by severe drought within a few months to a few years. As a result, drought-triggered forest mortality can result in rapid ecosystem changes over huge areas, far more quickly than the gradual transitions that occur from tree regeneration and growth. Land-use impacts such as anthropogenic burns and forest fragmentation, interacting with climate-induced forest stress, are likely to amplify forest dieback in some regions, for example the Amazon Basin (Nepstad et al., 2008). If current forest ecosystems are forced to adjust abruptly to new climate conditions through massive forest dieback, many pervasive and persistent ecological and social effects will result from the loss of forest products and ecosystem services – including sequestration of atmospheric carbon.
One consequence of substantial forest dieback is redistribution of withinecosystem carbon pools and rapid losses of carbon back to the atmosphere. For instance, climate-driven effects of forest dieback, insect and disease mortality and fire impacts have recently turned Canada’s temperate and boreal forests from a net carbon sink into a net carbon source (Kurz et al., 2008). Similarly, it is possible that “widespread forest collapse via drought” could transform the world’s tropical moist forests from a net carbon sink into a large net source during this century (Lewis, 2005).
Given the potential risks of climate-induced forest dieback, increased management attention to adaptation options for enhancing forest resistance and resilience to projected climate stress can be expected, for example thinning stand densities to reduce competition, selection for different genotypes (e.g. drought resistance) or translocation of species to match expected climate changes.
Foresters and ecologists have long known that climate stress has major effects on forest health. Awareness of, and interest in, climate-induced forest dieback is not new (Auclair, 1993; Ciesla and Donaubauer, 1994). It is known that natural climate variation historically triggered episodes of widespread forest mortality (Swetnam and Betancourt, 1998). So, one might ask, is anything new or different occurring now? Certainly the Earth is currently experiencing substantial, rapid, directional global climate change driven by major and pervasive human alterations of the Earth’s atmosphere, land surface and waters (IPCC, 2007). Concurrent with these changes, climate-related forest mortality is apparently increasing in many parts of the world. While the available evidence is not yet conclusive, it is possible that the increasing reports of dieback represent just the beginning of globally significant increases in problems associated with forest health and dieback. Given the dieback problems already reported under relatively modest recent increases in global mean temperature (about 0.5ºC since 1970) and drying climate in some areas (e.g. Seager et al., 2007), far greater chronic forest stress and mortality risk could be expected because much greater increases in mean temperature (about 2º to 4ºC globally, and more in some areas) and significant long-term regional drying in some places are projected to occur by 2100 (IPCC, 2007). Beyond changes in mean climate conditions, other climate changes such as extreme droughts, elevated maximum temperatures and longer-duration heat waves, which are projected to increase in frequency and severity (IPCC, 2007), might be expected to exacerbate forest dieback.
A number of information gaps and scientific uncertainties currently limit the conclusions that can be drawn about trends in forest mortality and the predictions that can be made about future climate-induced forest dieback. First, despite many national and even regional forest monitoring efforts, there is an absence of adequate global data on forest health status (FAO, 2006). Reliable long-term, global-scale forest health monitoring, combining remote-sensing and ground-based measurements, is needed to determine the status and trends of forest stress and mortality on the planet accurately, as well as to understand ecosystem responses after dieback events.
Second, adequate quantitative knowledge of the physiological thresholds of individual tree mortality from chronic or acute water stress is available for only a few tree species (McDowell et al., 2008), and associated temperature sensitivities are largely unknown. Further, there is little detailed understanding about the place-specific sequences and ranges of mean and extreme climatic conditions that can trigger species-specific tree mortality in forests on real landscapes and potentially lead to extensive forest dieback.
Third, scientists lack adequate knowledge of the feedback and non-linear interactions between climate-induced forest stress and other climate-related disturbance processes, such as insect outbreaks and fire, that can cause widespread forest mortality (Allen, 2007).
These scientific uncertainties about fundamental tree mortality processes represent a key limitation to more accurate quantitative modelling of future climate-induced forest dieback (e.g. Huntingford et al., 2008). Accordingly they also limit the ability to predict the implications of dieback for the potential of global forests to sequester excess atmospheric carbon or, alternatively, to become carbon sources and thereby contribute to amplified climate change (Lucht et al., 2006).
Overall, additional monitoring of global forest health and new research are needed to improve scientific certainty regarding risks of future climate-induced forest dieback for more accurate input to policy decisions and forest management worldwide.
![]() Bibliography
Bibliography
Allen, C.D. 2007. Cross-scale interactions among forest dieback, fire, and erosion in northern New Mexico landscapes. Ecosystems, 10: 797–808.
Allen, C.D., Macalady, A., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., Gonzales, P., Hogg, T,. Rigling, A., Breshears, D.D., Fensham, R., Zhang, Z., Kitzberger, T., Lim, J.-H., Castro, J., Running, S.W., Allard, G., Semerci, A. & Cobb, N. 2009. Drought-induced forest mortality: a global overview reveals emerging climate change risks. (Submitted for publication)
Angert, A., Biraud, S., Bonfils, C., Henning, C.C., Buermann, W., Pinzon, J., Tucker, C.J. & Fung. I. 2005. Drier summers cancel out the CO2 uptake enhancement induced by warmer springs. Proceedings of the NationalAcademy of Sciences of the United States of America, 102(31): 10823–10827.
Auclair, A.N.D. 1993. Extreme climatic fluctuations as a cause of forest dieback in the Pacific Rim. Water, Air and Soil Pollution, 66(3–4): 207–229.
Barber, V.A., Juday, G.P. & Finney, B.P. 2000. Reduced growth of Alaskan white spruce in the twentieth century from temperature-induced drought stress. Nature, 405: 668–673.
Boisvenue, C. & Running, S.W. 2006. Impacts of climate change on natural forest productivity – evidence since the middle of the 20th century. Global Change Biology, 12(5): 862–882.
Breda, N., Huc, R., Granier, A. & Dreyer, E. 2006. Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Annals of Forest Science, 63: 625–644.
Breshears, D.D., Cobb, N.S., Rich, P.M., Price, K.P., Allen, C.D., Balice, R.G., Romme, W.H., Kastens, J.H., Floyd, M.L., Belnap, J., Anderson, J.J., Myers, O.B. & Meyer, C.W. 2005. Regional vegetation die-off in response to global-change-type drought. Proceedings of the NationalAcademy of Sciences of the United States of America, 102(42): 15144–15148.
Ciais, P., Reichstein, M., Viovy, N., Granier, A., Ogee, J., Allard, V., Aubinet, M., Buchmann, N., Bernhofer, C., Carrara, A., Chevallier, F., De Noblet, N., Friend, A.D., Friedlingstein, P., Grunwald, T., Heinesch, B., Keronen, P., Knohl, A., Krinner, G., Loustau, D., Manca, G., Matteucci, G., Miglietta, F., Ourcival, J.M., Papale, D., Pilegaard, K., Rambal, S., Seufert, G., Soussana, J.F., Sanz, M.J., Schulze, E.D., Vesala, T. & Valentini, R. 2005. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature, 437(7058): 529–533.
Ciesla, W.M. & Donaubauer, M.E. 1994. Decline and dieback of trees and forests: a global overview. FAO Forestry Paper No. 120. Rome, FAO.
Clark, D.A. 2004. Sources or sinks? The responses of tropical forests to current and future climate and atmospheric composition. Philosophical Transcripts of the Royal Society of London, Series B, 359: 477–491.
Desprez-Loustau, M.-L., Marcais, B., Nageleisen, L.-M., Piou, D. & Vannini, A. 2006. Interactive effects of drought and pathogens in forest trees. Annals of Forest Science, 63: 597–612.
FAO. 2006. Global Forest Resources Assessment 2005 – Progress towards sustainable forest management. FAO Forestry Paper No. 147. Rome.
Fensham, R.J., Fairfax, R.J. & Ward, D.P. 2009. Drought-induced tree death in savanna. Global Change Biology. (In press)
Hicke, J.A., Logan, J.A., Powell, J. & Ojima, D.S. 2006. Changing temperatures influence suitability for modeled mountain pine beetle (Dendroctonus ponderosae) outbreaks in the western United States. Journal of Geophysical Research, 111: G02019.
Huntingford, C., Fisher, R.A., Mercado, L., Booth, B.B.B., Sitch, S., Harris, P.P., Cox, P.M., Jones, C.D., Betts, R.A., Malhi, Y., Harris, G., Collins, M. & Moorcroft, P. 2008. Towards quantifying uncertainty in predictions of Amazon “die-back”. Philosophical Transcripts of the Royal Society of London, Series B, 363(1498): 1857–1864.
Intergovernmental Panel on Climate Change (IPCC). 2007. Climate change 2007 –the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the IPCC. Cambridge, UK, Cambridge University Press.
Jump, A., Hunt, J.M. & Peñuelas, J. 2006. Rapid climate change-related growth decline at the southern range edge of Fagus sylvatica. Global Change Biology,12: 2163–2174.
Kurz, W.A., Stinson, G., Rampley, G.J., Dymond, C.C. & Neilson, E.T. 2008. Risk of natural disturbances makes future contribution of Canada’s forests to the global carbon cycle highly uncertain. Proceedings of the NationalAcademy of Sciences of the United States of America, 105: 1551–1555.
Lewis, S.L. 2005. Tropical forests and the changing earth system. Philosophical Transcripts of the Royal Society of London, Series B, 361: 195–210.
Lucht, W., Schaphoff, S., Erbrecht, T., Heyder, U. & Cramer, W. 2006. Terrestrial vegetation redistribution and carbon balance under climate change. Carbon Balance and Management, 1: 6.
McDowell, N., Pockman, W.T., Allen, C.D., Breshears, D.D., Cobb, N., Kolb, T., Sperry, J., West, A., Williams, D. & Yepez, E.A. 2008. Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? Tansley Review. New Phytologist, 178: 719–739.
Nepstad, D.C., Stickler, C.M., Soares-Filho, B. & Merry, F. 2008. Interactions among Amazon land use, forests and climate: prospects for a near-term forest tipping point. Philosophical Transcripts of the Royal Society of London, Series B, 363(1498): 1737–1746.
Nepstad, D.C., Tohver, I.M., Ray, D., Moutinho, P. & Cardinot, G. 2007. Mortality of large trees and lianas following experimental drought in an Amazon forest. Ecology,88(9): 2259–2269.
Peñuelas, J., Lloret, F. & Montoya, R. 2001. Severe drought effects on Mediterranean woody flora in Spain. Forest Science, 47: 214–218.
Raffa, K.F., Aukema, B.H., Bentz, B.J., Carroll, A.L., Hicke, J.A., Turner, M.G. & Romme, W.H. 2008. Cross-scale drivers of natural disturbances prone to anthropogenic amplification: the dynamics of bark beetle eruptions. Bioscience, 58(6): 501–517.
Seager, R., Ting, M., Held, I., Kushnir, Y., Lu, J., Vecchi, G., Huang, H.-P., Harnik, N., Leetmaa, A., Lau, N.-C., Li, C., Velez, J. & Naik, N. 2007. Model projections of an imminent transition to a more arid climate in southwestern North America. Science, 316: 1181–1184.
Soja, A.J., Tchebakova, N.M., French, N.H.F., Flannigan, M.D., Shugart, H.H., Stocks, B.J., Sukhinin, A.I., Varfenova, E.I., Chapin F.S. & Stackhouse, P.W. Jr. 2007. Climate-induced boreal forest change: Predictions versus current observations. Global and Planetary Change,56(3-4): 274–296.
Swetnam, T.W. & Betancourt, J.L. 1998. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. Journal of Climate, 11: 3128–3147.
van Mantgem, P.J., Stephenson, N.L., Byrne, J.C., Daniels, L.D., Franklin, J.F., Fulé, P.Z., Harmon, M.E., Larson, A.J., Smith, J.M., Taylor, A.H. & Veblen, T.T. 2009. Widespread increase of tree mortality rates in the western United States. Science, 323: 521–524.