![]()
![]()
![]()
Constantin MAXIM
Institut Roumain de Recherches Marines, Constantza, Roumanie
MATERIEL ET METHODE
Les prises de 1972-1981 sont extraites de COPACE/PACE SERIES 83/27, et les prises totales pour 1982 de CECAF Statistical Bulletin 1984 N°4. Les prises de 1983, 1984 et 1985 sont estimées à un niveau respectif de 280, 300 et 320 milliers de tonnes.
Le coefficient de mortalité naturelle est fixé à 0,5 comme dans les précédentes estimations.
Le taux de recrutement partiel est estimé à:
0,07 pour la classe 0+
0,06 pour la classe 1
1,00 pour la classe 2 et plus.
Nous avons considéré qu'a 2 ans les deux espèces (Trachurus trachurus et Trachurus trecae) sont pleinement recrutées.
Le vecteur F(1985) de départ est calculé à partir de la relation:
F = 0,01539.f + 0,2239
Pour une PUE de 13,92 tonnes/jour en 1985, on trouve un effort total f=22 985 jours de pêche.
| AGE | CAPTURE TOTALE (1000 t) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0+ | 1 | 2 | 3 | 4 | 5 | 6 | 7+ | ||
| 1972 | 0,00 | 1492,00 | 798,10 | 403,90 | 188,00 | 106,40 | 52,00 | 18,50 | 390,123 |
| 1973 | 0,00 | 2457,00 | 680,90 | 250,90 | 114,50 | 60,30 | 43,20 | 48,50 | 383,052 |
| 1974 | 982,00 | 3273,20 | 947,30 | 240,40 | 44,90 | 26,20 | 12,30 | 11,80 | 414,632 |
| 1975 | 964,30 | 2078,10 | 493,60 | 152,30 | 48,90 | 7,70 | 4,80 | 2,60 | 302,317 |
| 1976 | 3153,20 | 3202,30 | 685,60 | 97,40 | 35,60 | 16,30 | 4,60 | 1,10 | 394,145 |
| 1977 | 36,20 | 2927,20 | 1391,20 | 380,30 | 52,10 | 12,00 | 6,30 | 1,40 | 392,572 |
| 1978 | 60,00 | 1650,40 | 1162,80 | 338,70 | 78,20 | 16,50 | 3,00 | 0,50 | 294,151 |
| 1979 | 0,00 | 1323,30 | 534,70 | 231,00 | 99,00 | 29,60 | 10,00 | 0,80 | 223,165 |
| 1980 | 0,00 | 150,20 | 676,60 | 582,10 | 799,60 | 306,60 | 86,00 | 11,40 | 503,081 |
| 1981 | 0,00 | 230,40 | 415,20 | 462,20 | 357,90 | 207,20 | 72,60 | 14,10 | 357,935 |
| 1982 | 0,00 | 310,50 | 840,40 | 542,40 | 254,10 | 71,50 | 8,10 | 1,10 | 310,464 |
| 1983 | 0,00 | 591,10 | 727,30 | 262,90 | 135,90 | 88,40 | 62,50 | 31,80 | 280,000* |
| 1984 | 807,70 | 424,60 | 621,40 | 274,90 | 167,10 | 105,70 | 68,40 | 41,90 | 300,000* |
| 1985 | 0,00 | 145,60 | 800,10 | 360,30 | 109,80 | 126,40 | 48,70 | 2,50 | 320,000** |
| AGE | F PONDERE 1-5 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0+ | 1 | 2 | 3 | 4 | 5 | 6 | 7+ | ||
| 1972 | 0,000000 | 0,511725 | 0,788882 | 0,841723 | 0,767820 | 0,616278 | 0,322343 | 0,705246 | 0,705287 |
| 1973 | 0,000000 | 0,710480 | 0,691405 | 0,962460 | 0,953076 | 0,933614 | 0,833445 | 0,850099 | 0,850218 |
| 1974 | 0,082420 | 0,911205 | 1,462620 | 0,858620 | 0,660721 | 0,921245 | 0,740458 | 0,879356 | 0,879611 |
| 1975 | 0,057783 | 0,351619 | 0,472105 | 0,686220 | 0,617037 | 0,312602 | 0,620453 | 0,488036 | 0,487918 |
| 1976 | 0,139428 | 0,387599 | 0,261425 | 0,221046 | 0,483193 | 0,632487 | 0,445506 | 0,397193 | 0,397150 |
| 1977 | 0,002260 | 0,258610 | 0,413621 | 0,318220 | 0,245832 | 0,424763 | 0,816728 | 0,332598 | 0,332224 |
| 1978 | 0,005763 | 0,184893 | 0,215220 | 0,232136 | 0,136975 | 0,167873 | 0,248101 | 0,185781 | 0,185410 |
| 1979 | 0,000000 | 0,233446 | 0,115179 | 0,082385 | 0,135158 | 0,096253 | 0,187148 | 0,132838 | 0,132484 |
| 1980 | 0,000000 | 0,030647 | 0,250393 | 0,245816 | 0,651817 | 1,229370 | 0,641243 | 0,481579 | 0,481609 |
| 1981 | 0,000000 | 0,045911 | 0,152152 | 0,383773 | 0,330540 | 0,502452 | 2,385422 | 0,283060 | 0,282977 |
| 1982 | 0,000000 | 0,098016 | 0,327470 | 0,430687 | 0,546851 | 0,139337 | 0,043822 | 0,308936 | 0,308472 |
| 1983 | 0,000000 | 0,168887 | 0,496638 | 0,224000 | 0,253589 | 0,540531 | 0,241525 | 0,337494 | 0,336677 |
| 1984 | 0,186031 | 0,134262 | 0,380006 | 0,511330 | 0,341390 | 0,457392 | 0,010983 | 0,357427 | 0,357427 |
| 1985 | 0,040390 | 0,063021 | 0,577001 | 0,577000 | 0,577000 | 0,577000 | 0,577000 | 0,530840 | 0,530840 |
| M | 0,50 | 0,50 | 0,50 | 0,50 | 0,50 | 0,50 | 0,50 | 0,50 | |
| pig | 19,00 | 65,00 | 99,00 | 175,00 | 274,00 | 401,00 | 483,00 | 563,00 | |
| AGE | BIOMASSE 0-7+ | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0+ | 1 | 2 | 3 | 4 | 5 | 6 | 7+ | ||
| 1972 | 9836,60 | 4635,10 | 1800,00 | 871,70 | 432,00 | 286,60 | 236,60 | 45,10 | 1191,900 |
| 1973 | 11053,10 | 5966,20 | 1685,30 | 496,00 | 227,90 | 121,60 | 93,90 | 104,00 | 1066,500 |
| 1974 | 15718,70 | 674,10 | 1779,00 | 512,00 | 114,90 | 53,30 | 29,00 | 24,70 | 1080,900 |
| 1975 | 21772,00 | 8779,60 | 1634,80 | 379,00 | 131,60 | 36,00 | 12,90 | 8,40 | 1273,900 |
| 1976 | 30610,60 | 12463,90 | 3746,50 | 618,40 | 115,70 | 43,10 | 16,00 | 4,20 | 1929,900 |
| 1977 | 20373,30 | 1614,90 | 5130,70 | 1749,60 | 300,70 | 43,30 | 13,90 | 6,20 | 2360,900 |
| 1978 | 13264,60 | 12329,10 | 7563,30 | 2057,80 | 771,90 | 142,60 | 17,20 | 3,70 | 2441,400 |
| 1979 | 10411,70 | 7999,20 | 6215,70 | 3699,10 | 989,50 | 408,20 | 73,90 | 8,10 | 2455,600 |
| 1980 | 10737,70 | 6315,00 | 3841,60 | 3359,80 | 2066,20 | 524,30 | 224,90 | 37,20 | 2488,700 |
| 1981 | 6939,10 | 6512,70 | 3714,70 | 1813,90 | 1593,70 | 653,00 | 93,00 | 71,80 | 2024,300 |
| 1982 | 7913,90 | 4208,80 | 3772,90 | 1935,00 | 749,60 | 694,50 | 239,70 | 5,20 | 1738,600 |
| 1983 | 7041,20 | 4800,00 | 2314,40 | 1649,30 | 763,00 | 263,10 | 366,50 | 139,10 | 1533,400 |
| 1984 | 6000,00 | 4270,70 | 2458,90 | 854,50 | 799,60 | 359,10 | 93,00 | 174,60 | 1290,900 |
| 1985 | 7300,00 | 3021,40 | 2264,90 | 1019,90 | 310,80 | 357,80 | 137,90 | 7,50 | 1037,300 |
| AGE | ||||||||
|---|---|---|---|---|---|---|---|---|
| 0+ | 1 | 2 | 3 | 4 | 5 | 6 | 7+ | |
| 1972 | 186,89 | 301,28 | 178,20 | 152,54 | 118,37 | 114,92 | 114,30 | 25,41 |
| 1973 | 210,01 | 387,80 | 166,84 | 86,81 | 62,43 | 48,76 | 45,33 | 58,54 |
| 1974 | 298,66 | 435,76 | 176,12 | 89,60 | 31,47 | 21,37 | 14,00 | 13,93 |
| 1975 | 413,67 | 570,68 | 161,84 | 66,33 | 36,05 | 14,43 | 6,21 | 4,72 |
| 1976 | 581,60 | 810,16 | 370,90 | 108,22 | 31,71 | 17,27 | 7,71 | 2,36 |
| 1977 | 387,09 | 1049,75 | 507,94 | 306,18 | 82,39 | 17,36 | 6,70 | 3,49 |
| 1978 | 252,03 | 801,39 | 748,77 | 360,11 | 211,50 | 57,20 | 8,29 | 2,09 |
| 1979 | 197,82 | 519,95 | 615,35 | 647,34 | 271,13 | 163,71 | 35,68 | 4,58 |
| 1980 | 204,02 | 410,48 | 380,32 | 587,97 | 566,13 | 210,25 | 108,62 | 20,92 |
| 1981 | 131,84 | 423,33 | 367,75 | 317,44 | 436,68 | 261,87 | 44,92 | 40,44 |
| 1982 | 150,36 | 273,57 | 373,52 | 338,63 | 205,38 | 278,51 | 115,75 | 2,92 |
| 1983 | 133,78 | 312,00 | 229,13 | 288,63 | 209,05 | 105,51 | 177,00 | 78,33 |
| 1984 | 114,00 | 277,60 | 243,43 | 149,54 | 219,09 | 144,00 | 44,90 | 98,29 |
| 1985 | 138,70 | 196,39 | 224,22 | 178,49 | 85,16 | 143,48 | 66,59 | 4,25 |
| AGE | CAPTURE* F pond. BIOMASSE* | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 1 | 2 | 3 | 4 | 5 | 6 | 7 | ||
| CAPTURE | 0,000 | 145,600 | 800,100 | 360,300 | 109,800 | 126,400 | 48,700 | 2,500 | 257,428 |
| MORTALITE N | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | |
| MORTALITE P | 0,040 | 0,063 | 0,577 | 0,577 | 0,577 | 0,577 | 0,577 | 0,531 | |
| STOCK INCEPUT | 7300,000 | 3021,400 | 2264,900 | 1019,900 | 310,800 | 357,800 | 137,900 | 7,500 | 1037,280 |
| STOCK SFIRSIT | 7300,000 | 4252,400 | 1720,700 | 771,500 | 347,400 | 105,900 | 121,900 | 47,000 | 943,399 |
| P moyen (g) | 19,000 | 65,000 | 99,000 | 175,000 | 274,000 | 401,000 | 483,000 | 563,000 | |
| Variante 1 | |||||||||
| CAPTURE | 158,800 | 721,100 | 453,800 | 203,500 | 91,600 | 27,900 | 32,100 | 12,400 | 189,227 |
| MORTALITE N | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | |
| MORTALITE P | 0,028 | 0,240 | 0,400 | 0,400 | 0,400 | 0,400 | 0,400 | 0,400 | 0,171 |
| STOCK | 7300,000 | 4305,400 | 2028,900 | 699,600 | 313,600 | 141,200 | 43,000 | 49,600 | 933,101 |
| P moyen (g) | 19,000 | 65,000 | 99,000 | 175,000 | 274,000 | 401,000 | 483,000 | 563,000 | |
| Variante 2 | |||||||||
| CAPTURE | 178,400 | 800,700 | 499,800 | 224,100 | 100,900 | 30,800 | 35,400 | 13,600 | 208,899 |
| MORTALITE N | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | |
| MORTALITE P | 0,032 | 0,270 | 0,450 | 0,450 | 0,450 | 0,450 | 0,450 | 0,450 | 0,190 |
| STOCK | 7300,000 | 4290,400 | 1968,900 | 665,400 | 298,400 | 134,400 | 40,900 | 47,100 | 910,888 |
| P moyen (g) | 19,000 | 65,000 | 99,000 | 175,000 | 274,000 | 401,000 | 483,000 | 563,000 | |
| Variante 3 | |||||||||
| CAPTURE | 197,900 | 878,100 | 543,800 | 243,800 | 109,800 | 33,500 | 38,500 | 14,800 | 227,810 |
| MORTALITE N | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | |
| MORTALITE P | 0,035 | 0,300 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,209 |
| STOCK | 7300,000 | 4275,400 | 1910,700 | 633,000 | 283,800 | 127,800 | 38,900 | 44,800 | 889,600 |
| P moyen (g) | 19,000 | 65,000 | 99,000 | 175,000 | 274,000 | 401,000 | 483,000 | 563,000 | |
| Variante 4 | |||||||||
| CAPTURE | 227,800 | 993,300 | 607,800 | 272,500 | 122,700 | 37,400 | 43,100 | 16,600 | 255,521 |
| MORTALITE N | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | 0,500 | |
| MORTALITE P | 0,040 | 0,346 | 0,577 | 0,577 | 0,577 | 0,577 | 0,577 | 0,577 | 0,237 |
| STOCK | 7300,000 | 4252,400 | 1824,500 | 586,100 | 262,800 | 118,300 | 36,100 | 41,500 | 858,530 |
| P moyen (g) | 19,000 | 65,000 | 99,000 | 175,000 | 274,000 | 401,000 | 483,000 | 563,000 | |
(* : dans cette colonne, les captures et biomasses sont exprimées en milliers de tonnes)
C. MAXIM, I. PANAIT, I.STAICU
Institut Roumain de Recherches Marines
Constantza
INTRODUCTION
La pêche dans les années 80 sur les côtes mauritaniennes a conduit à une continuelle croissance des captures de cométe coussut, son taux dans la categorie des chinchards augmentant, selon la statistique roumaine, depuis environ 4% en 1980 à plus de 40% en 1983, pour atteindre en 1985 un niveau aussi élevé, dépassant 36% (tableau 1).
La tendance du cométe coussut à devenir un objet principal des captures, à côté de l'existence d'un échantillonnage biologique et statistique multi-annuel, ainsi que la présence dans le secteur sénégalo-mauritanien (11° -24° lat. N) d'un stock distinct (COPACE - 1983), constituent les principaux éléments qui ont rendu nécessaire et opportune l'évaluation du stock de cette espéce.
Une hypothése, non prouvée jusqu'à présent mais généralement acceptée, est que l'abondance du stock du cométe coussut a eu de grandes variations dans le temps. Ces fluctuations numériques peuvent étre associées aux fortes différences existant dans la structure annuelle par longueur et par àge observées dans les captures industrielles, ainsi qu'aux facteurs du milieu, tels la température, la variation de la direction et de l'intensité des vents dominants, l'abondance de la nourriture ou les maladies. D'autre part, ainsi qu'on l'a prouvé pour d'autres espéces (LETT et al., 1975), le recrutement, l'abondance du stock et la capture peuvent grandement varier dans les conditions d'une mortalité par la pêche diminuée, comme c'était sans doute le cas du cométe coussut à la fin des années 60.
Le but de cet article est donc d'établir l'abondance du stock du cométe coussut sénégalo-mauritanien pendant la période 1972-1985, intervalle durant lequel on peut parler d'une véritable pêche industrielle de l'espéce.
MATERIEL ET METHODE
La pêche du comète coussut le long des côtes nord-ouest africaines a une longue et riche tradition pour les pays riverains ainsi que pour les flottilles expéditionnaires européennes. Les quantités relativement réduites que l'on pouvait y pêcher, donc le taux faible dans la pêche spécialisée des chinchards et de la sardine lui ont conféré au cours des années un fort caractére d'espéce auxiliaire. Mais, a partir des années 70, on peut parler d'une participation augmentée du cométe coussut à la pêche effectuée sur le plateau continental mauritanien (34.1.3 + 34.3.1), les captures oscillant entre 15 et 38 mille tonnes pendant la période 1972-1985 (tableau 2) (CECAF - Stat.Bul. n°4 - 1984).
Les éléments nécessaires à l'évaluation provenaient du calcul du nombre d'exemplaires capturés par classes d'âge, du poids moyen correspondant, de la determination des mortalités naturelle et par pêche pour la dernière année, de l'âge maximum, des recrutements partiels, du niveau de recrutement en 1985 et de toute une série de données biologiques telles que l'âge de la première maturation, l'âge de la maturation générale, les constantes de l'équation de croissance en longueur et en poids, etc.
On a déterminé les données fondamentales comme suit:
Le nombre d'individus par classes d'âge a été calculé, pour chaque année de l'analyse, selon la fréquence par classes de longueur et les poids moyens correspondants. Des que l'on a trouve le nombre total d'individus pêchés chaque année, on l'a divisé par classes d'âge conformément à la structure résultant de l'echantillonnage roumain (tableau 3);
Les poids moyens par classes d'âge ont été déterminés conformément aux relations suivantes (MAXIM, STAICU - 1976):
lt = 55,7 × (1 - e-0,136(t+1,295))
P = 0,0799 × L2,5043
d'ou résultent les valeurs comprises dans le tableau 4;
La mortalité naturelle M a été déterminée par cinq méthodes différentes, en considérant l'âge de la première maturation (2 ans), l'âge de la maturation générale (3 ans), l'âge maximum observé dans l'échantillonnage (13 ans), K=0,448, to=-1,295, Loo=55,7 cm, la moyenne annuelle de la température de la surface de l'eau (20,6°C) dans le secteur peuplé par le stock analysé, et l'indice des gonades (0,17).
La valeur moyenne résultante est M = 0,45;
Le coefficient de mortalité instantanée par la pêche pour la dernière année et l'âge maximal dans la dernière évaluation, à été de F = 0,40. Cette valeur a résulté d'un premier roulement en V.P.A. avec F initial en 1984 de 0,45. On a pu ainsi calculer les valeurs pondérées de F pour les principaux groupes d'âge des captures, de 3 à 6 ans compris.
L'équation linéaire qui relie les valeurs pondérées de F3-6 à l'effort total exprime en jours (calculé comme le rapport entre la capture totale du cométe coussut dans les divisions 34.1.3 et 34.3.1 et la PUE roumaine dans la pêche de l'espèce (tableau 2) a la forme suivante:
F = 0,000227 × f - 0,62976
De la relation ci-dessus, avec un effort total f1985 = 4537 jours, on a obtenu une valeur de F correspondant à l'année de base de 0,398. Dans notre évaluation nous avons utilisé une mortalité instantanée par la pêche F1985 = 0,40.
Les recrutements partiels, calculés à partir des rapports entre les mortalités par la pêche à 1 et 2 ans et F3-6, sont de 0,0 à l'âge de 1 an, 0,6 à 2 ans et 1 à 3 ans et plus.
Etant donné le manque d'autres voies d'estimation du niveau de recrutement, on a considéré pour les années 1984 et 1985 la valeur 415,6 millions d'individus, qui représente la moyenne de la période 1971-1983. Ce niveau peut être considéré faible si on regarde la tendance évidente de croissance du recrutement durant les années 1982 et 1983, mais pour une première évaluation de l'espèce on peut l'estimer satisfaisant, étant donné que le principal objectif poursuivi est l'évolution de l'ordre de grandeur du stock au cours du temps.
RESULTATS OBTENUS
Conformément aux estimations obtenues par l'analyse de la population virtuelle en ce qui concerne la structure en nombre d'individus par classes d'âge (tableau 5), l'abondance du comète coussut de la région sénégalo-mauritanienne a enregistré d'importantes variations annuelles en raison de l'apparition de générations riches au cours des années 1976, 1977 et 1980–1982 inclues.
L'abondance maximale observée en 1983 diminue ensuite peu à peu mais en se maintenant à des niveaux supérieurs à 1100 millions d'individus (tableau 7). Le grand recrutement des années 1980, 1981 et 1982 apparaft comme une conséquence directe de la diminution de l'abondance numérique de deux autres espèces fondamentales de la pêche sur le plateau continental mauritanien, le chinchard d'Europe (Trachurus trachurus) et surtout le chinchard cunène (Trachurus trecae), ce qui a créé une disponibilité de la nourriture du comète coussut (IRCM - 1985, 1986).
On n'est pas encore sûr du crédit à apporter à la relation directe entre la variation de la PUE (tableau 2) et la valeur de la biomasse totale (tableau 7), mais dans l'ensemble de la période, leur évolution est relativement identique sauf pendant les années 1977 et 1981 (figure 1).
Une meilleure corrélation apparaft dans le cas de la PUE et de la mortalitépar la pêche (tableau 6), qui ont des variations annuelles beaucoup plus proches (figure 2).
La capture par unité d'effort (tonnes/jour), des chalutiers qui pêchent sur le plateau continental mauritanien et qui obtiennent la plupart des captures des dernières années, peut donc être appréciée comme une mesure acceptable de l'abondance du stock.
L'évaluation de la biomasse totale (tableau 7) est directement liée au taux de mortalité par la pêche (F) que nous avons choisi comme valeur de départ dans la denière année de base (1985). Le calcul de l'abondance des différentes générations, ayant ce F terminal, peut devenir une source d'erreurs dans le cas d'une surestimation ou sous-estimation de cette valeur. La valeur de F (0,40) utilisée en V.P.A. semble pourtant être acceptable (un peu surestimée dans notre évaluation), de mainère à éviter les écarts majeurs de la vraie abondance par classes d'âge.
Une valeur diminuée (F = 0,3 par exemple) aurait impliqué une augmentation de la biomasse totale également en 1985 (croissance d'environ 18% par rapport à 1984), aspect que l'on accepte qu'avec réserves tant que l'on ne connait pas la valeur du recrutement.
Sans tenir compte de la valeur de F utilisée, la tendance d'augmentation de la biomasse totale est evidente. Son accroissement depuis 1980 jusqu'en 1985 dépasse 66% (tableau 7). Notre analyse indique une variabilité relativement faible de la grandeur du stock ( iii 000 à 206 000 t de 1972 à 1985), pouvant constituer un indicateur de la dynamique survenue au niveau des abondances annuelles.
Si on regarde les avantages qu'offrent l'utilisation d'un F terminal le plus proche possible de la réalité, ainsi que la connaissance correcte de la valeur du recrutement, on peut apprécier que ces deux éléments constituent toujours les fondements essentiels pour obtenir une image nette de l'évolution de l'abondance du comète coussut sénégalo-mauritanien.
BIBLIOGRAPHIE
COPACE - 1983 : Rapport du groupe de travail ad-hoc sur les chinchards et les maquereaux de la zone nord du COPACE.COPACE/PACE Séries 83/27:1–45
LETT P.F., W.T.STOBO and W.G.DOUBLEDAY - 1975 : A system simulation of the Atlantic mackerel fishery in ICNAF Subareas 3, 4 and 5 and Statistical Area 6; with special reference to stock management. Ann. Meet.Int.Commm.Northw.Atlant.Fish. 1975, Res.Doc.32, Serial N°3511 (mimeographed).
MAXIM C.et I.STAICU - 1976 : Observations sur la biologie et la dynamique du stock de saurel (Caranx rhonchus Geof.St.Hil.) dans l'Atlantique est-central. IRCM, Cerc.Mar.nr.9:215-226
IRCM - 1985 : Raport asupra rezultatelor cercetärilor românesti efectuate in zonele de pescuit din Oceanul Mondial in anul 1984. IRCCM - MAIA/DIA; 233 pp.
IRCM - 1986 : Rapport asupra rezultatelor cercetärilor românesti efectuate in zonele de pescuit ale Oceanul Atlantic in anul 1985 si evaluarea resurselor exploatabile..IRCCM - MAIAAPA;210 pp.
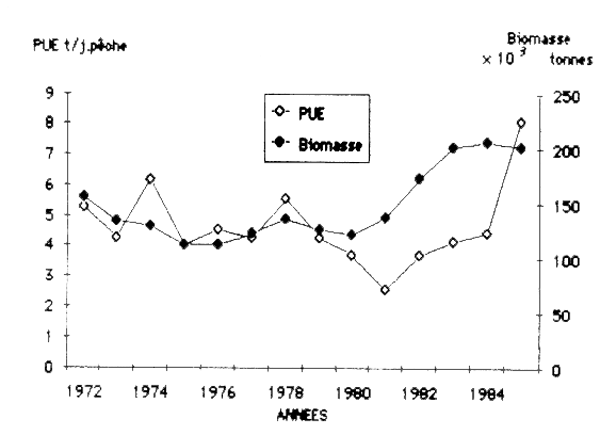
figure 1 : Evolution des PUE et des Biomasses
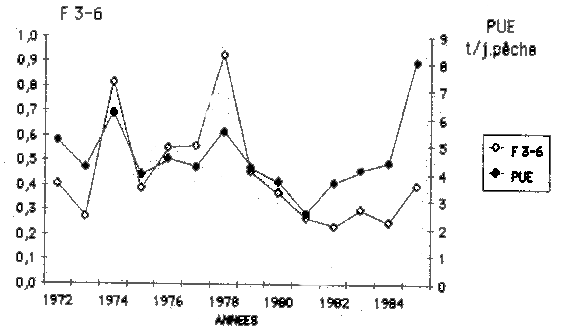
figure 2 : Evolution des mortalités par pêche et des PUE.
| Espèce/année | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 |
|---|---|---|---|---|---|---|
| Total chinchards | 64043 | 64099 | 44527 | 20239 | 17739 | 45409 |
| Trachurus t | 50015 | 18878 | 14101 | 1928 | 4090 | 12416 |
| trachurus % | 78,1 | 29,5 | 31,7 | 9,5 | 23,1 | 27,3 |
| Trachurus t | 11648 | 39203 | 20572 | 10105 | 7846 | 16326 |
| trecae % | 18,2 | 61,2 | 46,2 | 49,9 | 44,2 | 36,0 |
| Decapterus t | 2380 | 6018 | 9854 | 8206 | 5803 | 16667 |
| rhonchus % | 3,7 | 9,3 | 22,1 | 40,6 | 32,7 | 36,7 |
| Capture Roumaine (t) | Effort Roumaine (jours) | PUE Roumaine (t/jour) | Capture totale (t) | Effort total (jours) | |
|---|---|---|---|---|---|
| 1972 | 5832 | 1114 | 5,24 | 28645 | 5477 |
| 1973 | 1373 | 323 | 4,25 | 20850 | 4906 |
| 1974 | 4727 | 764 | 6,19 | 38156 | 6164 |
| 1975 | 3079 | 768 | 4,01 | 19164 | 4779 |
| 1976 | 668 | 147 | 4,54 | 22931 | 5062 |
| 1977 | 486 | 114 | 4,26 | 21449 | 4954 |
| 1978 | 61 | 11 | 5,55 | 30051 | 5575 |
| 1979 | 111 | 26 | 4,27 | 20633 | 4889 |
| 1980 | 2380 | 642 | 3,71 | 17210 | 4639 |
| 1981 | 6018 | 2357 | 2,55 | 14981 | 5921 |
| 1982 | 9854 | 2678 | 3,68 | 17941 | 5040 |
| 1983 | 8206 | 1987 | 4,13* | 20000 | 4843 |
| 1984 | 5803 | 1316 | 4,41* | 22000 | 4989 |
| 1985 | 16667 | 2065 | 8,07* | 35000 | 4537 |
| AGE | CAPTURE TOTALE (t) | |||||||
|---|---|---|---|---|---|---|---|---|
| 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
| 1972 | 2,20 | 26,00 | 47,80 | 16,10 | 5,10 | 2,80 | 28645 | |
| 1973 | 1,20 | 45,80 | 38,60 | 10,20 | 3,40 | 0,80 | 20850 | |
| 1974 | 4,80 | 52,20 | 35,20 | 5,80 | 1,60 | 0,40 | 38156 | |
| 1975 | 18,00 | 34,70 | 38,80 | 3,00 | 4,00 | 1,50 | 19164 | |
| 1976 | 4,00 | 29,00 | 29,50 | 11,50 | 23,50 | 2,50 | 22931 | |
| 1977 | 32,20 | 45,30 | 14,40 | 2,70 | 2,00 | 3,40 | 21449 | |
| 1978 | 12,10 | 26,20 | 39,30 | 14,30 | 2,70 | 3,10 | 2,30 | 30051 |
| 1979 | 27,10 | 70,20 | 2,60 | 0,10 | 0,10 | 0,10 | 20633 | |
| 1980 | 2,00 | 12,90 | 32,50 | 31,20 | 18,30 | 3,10 | 17210 | |
| 1981 | 0,30 | 18,80 | 39,80 | 32,50 | 8,60 | 0,10 | 1,10 | 14981 |
| 1982 | 16,90 | 12,60 | 25,80 | 25,50 | 17,40 | 1,70 | 0,10 | 17941 |
| 1983 | 0,80 | 17,50 | 37,00 | 18,20 | 16,70 | 9,00 | 0,80 | *20000 |
| 1984 | 17,40 | 42,10 | 18,20 | 11,60 | 10,40 | 0,30 | *22000 | |
| 1985 | 2,00 | 41,10 | 36,00 | 14,40 | 5,20 | 1,20 | 0,10 | *35000 |
| AGE | ||||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| Lm(cm) | 14,90 | 20,10 | 23,80 | 27,70 | 31,10 | 34,00 | 26,80 | 39,00 |
| P moy(g) | 66,00 | 142,00 | 228,00 | 306,00 | 416,00 | 542,00 | 687,00 | 812,00 |
| AGE | CAPTURE TOTALE (t) | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8+ | ||
| 1972 | 0,0 | 0,00 | 2,8 | 24,3 | 32,9 | 8,5 | 2,1 | 1,0 | 28645 |
| 1973 | 0,0 | 0,00 | 1,4 | 31,2 | 19,3 | 3,9 | 1,0 | 0,2 | 20850 |
| 1974 | 0,0 | 0,00 | 8,0 | 65,0 | 32,3 | 4,1 | 0,9 | 0,2 | 38156 |
| 1975 | 0,0 | 23,3 | 25,6 | 18,2 | 1,2 | 1,5 | 0,5 | 0,00 | 19164 |
| 1976 | 0,0 | 8,3 | 30,9 | 20,6 | 6,3 | 10,6 | 1,0 | 0,00 | 22931 |
| 1977 | 0,0 | 0,00 | 30,2 | 31,8 | 7,4 | 1,1 | 0,6 | 0,9 | 21449 |
| 1978 | 0,0 | 34,2 | 34,5 | 38,6 | 10,3 | 1,5 | 1,4 | 0,8 | 30051 |
| 1979 | 0,0 | 53,8 | 72,4 | 1,8 | 0,1 | 0,0 | 0,0 | 0,00 | 20633 |
| 1980 | 0,0 | 2,8 | 10,6 | 18,3 | 13,2 | 6,4 | 0,9 | 0,00 | 17210 |
| 1981 | 0,0 | 0,8 | 14,0 | 17,8 | 9,6 | 2,4 | 0,2 | 0,2 | 14981 |
| 1982 | 0,0 | 17,9 | 9,6 | 14,8 | 11,1 | 5,8 | 0,5 | 0,2 | 17941 |
| 1983 | 0,0 | 1,8 | 20,0 | 29,4 | 9,6 | 7,4 | 3,3 | 0,2 | *20000 |
| 1984 | 0,0 | 0,00 | 24,1 | 33,9 | 10,0 | 5,3 | 4,0 | 0,1 | *22000 |
| 1985 | 0,0 | 6,0 | 69,2 | 43,1 | 12,6 | 3,4 | 0,7 | 0,1 | *35000 |
| AGE | F 3 – 6 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8+ | ||
| 1972 | 0,000 | 0,000 | 0,018 | 0,463 | 1,479 | 1,526 | 1,427 | 0,872 | 0,402 |
| 1973 | 0,000 | 0,000 | 0,009 | 0,384 | 1,248 | 1,020 | 1,120 | 0,665 | 0,274 |
| 1974 | 0,000 | 0,000 | 0,120 | 1,052 | 1,328 | 1,689 | 0,001 | 1,047 | 0,813 |
| 1975 | 0,000 | 0,128 | 0,387 | 0,594 | 0,058 | 0,236 | 0,003 | 0,319 | 0,388 |
| 1976 | 0,000 | 0,047 | 0,332 | 0,880 | 0,582 | 1,514 | 0,324 | 0,827 | 0,553 |
| 1977 | 0,000 | 0,000 | 0,314 | 0,968 | 1,505 | 0,247 | 0,396 | 0,759 | 0,559 |
| 1978 | 0,000 | 0,123 | 0,511 | 1,253 | 1,664 | 7,921 | 0,000 | 2,837 | 0,923 |
| 1979 | 0,000 | 0,256 | 0,561 | 0,057 | 0,011 | 0,003 | 0,000 | 0,157 | 0,454 |
| 1980 | 0,000 | 0,019 | 0,095 | 0,357 | 1,053 | 3,490 | 2,323 | 1,249 | 0,366 |
| 1981 | 0,000 | 0,004 | 0,163 | 0,303 | 0,434 | 0,772 | 0,189 | 0,418 | 0,264 |
| 1982 | 0,000 | 0,067 | 0,084 | 0,345 | 0,422 | 0,714 | 0,485 | 0,391 | 0,230 |
| 1983 | 0,000 | 0,005 | 0,129 | 0,538 | 0,539 | 0,779 | 2,172 | 0,496 | 0,300 |
| 1984 | 0,000 | 0,000 | 0,112 | 0,448 | 0,480 | 0,940 | 2,808 | 0,495 | 0,247 |
| 1985 | 0,000 | 0,028 | 0,400 | 0,400 | 0,400 | 0,400 | 0,400 | 0,400 | 0,400 |
* : valeurs estimées
| AGE | Abondance totale | Biomasse totale (* 1000t) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8+ | |||
| 1972 | 214,6 | 296,3 | 191,4 | 80,1 | 50,2 | 12,8 | 3,3 | 2,1 | 850,8 | 156,1 |
| 1973 | 240,4 | 136,8 | 188,9 | 119,8 | 32,1 | 7,3 | 1,8 | 0,5 | 727,6 | 134,0 |
| 1974 | 375,5 | 153,3 | 87,2 | 119,4 | 52,0 | 5,9 | 1,7 | 0,4 | 795,4 | 129,3 |
| 1975 | 354,7 | 239,4 | 97,7 | 49,3 | 26,6 | 8,8 | 0,7 | 0,0 | 777,2 | 111,1 |
| 1976 | 258,6 | 226,2 | 134,3 | 42,3 | 17,3 | 16,0 | 4,4 | 0,0 | 699,1 | 111,7 |
| 1977 | 570,9 | 164,9 | 137,7 | 61,5 | 11,2 | 6,2 | 2,2 | 2,0 | 956,6 | 122,5 |
| 1978 | 459,0 | 364,0 | 105,1 | 64,1 | 14,9 | 1,6 | 3,1 | 1,0 | 1012,8 | 135,5 |
| 1979 | 287,0 | 292,7 | 205,1 | 40,2 | 11,7 | 1,8 | 0,0 | 0,0 | 838,5 | 125,5 |
| 1980 | 362,4 | 183,4 | 144,4 | 74,7 | 24,2 | 7,4 | 1,1 | 0,0 | 797,6 | 120,6 |
| 1981 | 538,2 | 231,1 | 114,7 | 83,7 | 33,3 | 5,4 | 0,1 | 0,1 | 1006,6 | 137,0 |
| 1982 | 693,8 | 343,2 | 146,7 | 62,1 | 39,4 | 13,8 | 1,6 | 0,1 | 1300,7 | 172,0 |
| 1983 | 631,7 | 442,4 | 204,7 | 86,0 | 28,1 | 16,5 | 4,3 | 0,6 | 1414,3 | 201,6 |
| 1984 | 415,6 | 402,8 | 280,7 | 114,7 | 32,0 | 10,4 | 4,8 | 0,3 | 1261,3 | 206,3 |
| 1985 | 415,6 | 265,0 | 256,8 | 160,0 | 46,8 | 12,6 | 2,6 | 0,2 | 1159,6 | 200,8 |
Domanevsky L.N., Krivospichenko S.G., Mylnikov N.I.
AtlantNIRO, Kaliningrad, URSS
1. INTRODUCTION
Au cours de la période 1982–1985 les chercheurs de l'AtlantNIRO ont prêté une attention particulière à l'état des stocks du maquereau d'Espagne. En outre, ils ont continué à recueillir les données bio-statiques sur le chinchard d'Europe et le chinchard ouest-africain, nécessaires à l'étude de la dynamique de leur abondance dans les années à venir. Une grande attention à été prêtée à la détermination de la grandeur du recrutement (premier groupe d'âge) à la base des prospections quantitatives.
2. MAQUEREAU D'ESPAGNE (Scomber japonicus)
Au cours de la période passée depuis le moment de la préparation des matériaux pour le second groupe de travail du COPACE sur les chinchards et les maquereaux, on a continué à déterminer l'abondance et la biomasse du maquereau d'Espagne, la dépendance empirique entre l'abondance du stock des parents et la postérité est mise en évidence.
2.1. Méthodes
Les informations relatives à la structure de taille du maquereau d'Espagne de la division 34.1.3 sont obtenues par les bateaux soviétiques à grand tonnage; le nombre d'observations estillustré dans les tableaux.
Les données sur les proportions de différents groupes d'âge du maquereau d'Espagne sont calculées pour chaque année à partir de la composition par taille à l'aide des clés taille-âge.
Les calculs de l'analyse de cohorte sont basés sur les données statistiques du COPACE (Bulletin statistique, 1984) et les données des bulletins STATLANT 34a, élaborés par le VNIRO pour le COPACE. Dans certains cas on utilise les estimations préliminaires données dans les annexes.
Les calculs de l'analyse de cohorte sont réalisés d'après la formule Pope (Pope, d'après Ricker, 1979):
N1 = N2 × eM + C1 × eM/2
où N1 : abondance du groupe d'âgeâge cadet pour une année n-1
N2 : abondance du groupe d'âge ainé pour une année n
C1 : nombre d'individus capturés en une année n-1
e : base du degré de logarithmes naturels
M : coefficient de mortalité naturelle momentanée du maquereau
d'Espagne
Le coefficient de mortalité naturelle momentanée du maquereau d'Espagne est pris à 0,6 (Krivospitchenko, Domanevsky, 1984)
L'abondance du maquereau pour la première année de la cohorte a été calculée d'après la formule:
N1 = C1/Fst
où N1 : abondance du maquereau pour une année n
C1 : abondance d'individus capturés en une année
Fst : coefficient de départ de mortalité momentanée par pêche.
Le coefficient de départ de mortalité momentanée par pêche pour les générations entièrement capturées est pris egal à 0,6 (Krivospitchenko, Domanevsky, 1984). Les coefficients de départ de mortalité momentanée par pêche pour les générations non entièrement capturées sont calculés à l'aide de dépendances empiriques par rapport au niveau de l'effort de péche total standardisé (type BMRT).
Les donnees relatives aux variations de mortalité par pêche sont obtenues à pantin des calculs de l'analyse de cohorte des générations entièrement capturées.
Sont obtenues les dépendances suivantes:
F 2–3 = 0,27 + (0,006 × f)
F 3–4 = 0,42 + (0,003 × f)
F 4–5 = -0,14 + (0,043 × f)
F 5–6 = 0,42 + (0,019 × f)
Le niveau d'effort standardisé étant 12 900 – 19 500 jours de pêche, les coefficients de départ de mortalité momentanée sont :
F 2–3 = 0,4
F 3–4 = 0,5
F 4–5 = 0,6
F 5–6 = 0,7
Le coefficient de mortalité momentané par pêche pour la première année de vie est pris pour 0,2.
Les calculs de la biomasse sont réalisés à partir de l'abondance des cohortes et la masse moyenne de chaque groupe d'âge (Krivospitchenko, Domanevsky, 1984).
Les calculs des relations entre l'abondance du stock de parents et sa postérité (individus àgés d'un an) sont basés sur la “courbe de recrutement” de Ricker (1979). Le calcul de la dépendance linéaire de la différence entre les logarithmes naturels de l'abondance de la postérité et du stock de parents et l'abondance du stock de parents est effectué par moindres carrés (Plokhinsky, 1970). Les coefficients obtenus de l'équation de régression sont utilisés pour la “courbe de recrutement”.
2.2. Composition par taille
La composition par taille des captures de maquereau d'Espagne dans la division 34.1.3 pour les années 1972–1979 a été examinée dans le travail précédent (Krivospitchenko, Domanevsky, 1984) et n'est pas illustrée ici. Les données précisées pour 1980–1985 sont montrées dans le tableau 2.1.
2.3. Composition par âge
La composition par âge du maquereau d'Espagne dans la division 34.1.3 a subi de considérables variations en 1972–1985 (tab. 2.2). En 1977, 1982 et 1985 la part d'individus âgés d'un an a augmenté. En 1973, 1975, 1979 et 1981 la base des captures a été constituée des poissons àgés de 2–3 ans.
2.4. Statistiques de pêche
La prise du maquereau d'Espagne dans la division 34.1.3 variait entre 128 000 et 22 000 tonnes (tab. 2.3), grâce aux captures de l'URSS en général. La diminution des captures soviétiques en 1978 est conditionnée par la baisse de l'effort de pêche. La capture soviétique pour 1985 correspond conventionnellement au niveau des années 1981–1982.
2.5. Biomasse et abondance
La prise du maquereau d'Espagne dans la division 34.1.3 variait entre 200 et 500 millions d'individus; elle a été très basse - 80,7 millions d'individus - en 1978 et très élevée - 748,0 millions d'individus - en 1977 (tab. 2.4).
L'abondance du maquereau en 1972 et 1977 a dépassé 2 milliards d'individus; en 1975, 1980 et 1983 elle a été inférieure à 1,5 milliards d'individus (tab. 2.5). Ces variations sont dues à la croissance et la diminution de la part du premier groupe d'âge dans l'abondance totale.
La biomasse du maquereau d'Espagne a subi de moindres fluctuations (tab. 2.6). Cependant, dans certaines années (1977, 1984), la moitié de la biomasse a été constituée des individus du premier groupe d'âge.
2.6. Dépendance entre l'abondance du stock de parents et la postérité
Dans la division 34.1.3 le maquereau d'Espagne (50 %) atteint sa maturité mesurant 25,7 cm, ce qui correspond à un âge un peu inférieur à 2 ans (Krivospitchenko, Domanevsky, 1984). Donc, l'abondance des groupes d'âge II–VI d'une année peut étre considérée comme l'abondance du stock de parents, et l'abondance du groupe d'âge I de l'année suivante comme l'abondance de la postérité. L'analyse des variations communes de caractéristiques données a permis de mettre en évidence leur dépendance mutuelle et d'appliquer l'approche de Ricker (Ricker, 1979), pour la période 1972–1981 dont les informations sont assez solides. La corrélation entre la différence des logarithmes naturels de l'abondance de la postérité et du stock de parents (Log(Nr) - Log(Np) ) et l'abondance du stock de parents a été négative et incontestable du point de vue des statistiques (au niveau de signification de 0,05). Sa valeur est la suivante:
n = 10 ; r = -0,895
L'équation de corrélation est la suivante:
Nr = 11,48 × Np × exp (-2,80 × 10–3 ×Np)
où : Nr - abondance de la postérité en millions d'individus
Np - abondance du stock de parents en millions d'individus
La corrélation entre les valeurs réelles de l'abondance de la postérité et les valeurs théoriques calculées à la base de l'équation donnée est incontestable ( au niveau de signification de 0,05).
Sa valeur:
n = 10 ; r = 0,763
3. CHINCHARDS (Trachurus trachurus L.- chinchard d'Europe; Trachurus trecae C. - chinchard ouest-africain)
3.1. Matériel et méthode
Les matériaux publiés dans ce travail représentent la suite de matériaux basés sur les résultats de recherches soviétiques publiés après la réunion du groupe de travail tenu en janvier-février 1983 à Nouadibhou (1984). Les données sur la structure de taille sont obtenues par les bateaux de pêche soviétiques à grand tonnage. Les rangs de taille ont été calculés en fonction de la grandeur de la prise par heure de chalutage; puis ils ont été généralisés par mois et pour toute une année.
La structure de l'âge des chinchards est calculée par les clés taille-âge pour l'année à partir de la structure de taille.
Les statistiques de pêche pour quelques dernières années sont prises du Bulletin Statistique (1984) et des Bulletins Statland 34A pour 1983 et 1984 préparés par le VNIRO pour le COPACE. Les proportions de chinchards ont été calculées en fonction de leurs poids dans les prises. Ces propertion ont été calculées par mois, trimestres, années.
3.2. Composition par taille
Dans la division 34.1.3 la structure de taille du chinchard d'Europe subit de considérables variations inter-annuelles. Comme nous avons noté lors du Groupe de travail précédent (1984), la composition par taille est influencée par le rendement des générations.
Les travaux d'inventaire, que nous avons accomplis en 1981–1985, ont montré la présence riche du chinchard d'Europe. En raison de cela, la part de chinchards dont la taille est inférieure à 20 cm (tab. 3.1) est importante dans les prises des bateaux de pêche pour les années 1982–1985. En 1984, de grands individus du chinchard d'Europe mesurant 32–33 cm dominent dans la structure de taille.
Les chinchards ouest-africains appartenant au stock sénégalo-mauritanien des divisions 34.1.3 et 34.3.1 durant les années 1979–1983 et 1985 sont représentés en général par de grands individus dont la taille est supérieure à 20 cm (tab. 3.2). En 1984 la base du stock est constituée des poissons mesurant 19–22 cm appartenant probablement à la génération riche de 1982.
3.3 Composition par âge
Les individus âgés de 1–2 à 3 ans, appartenant aux générations riches de 1980–1984, sont dominants dans la composition par âge du chinchard d'Europe dans la période 1982–1985 (tab. 3.3). Dans la période 1984 la base du stock est constituée de chinchards d'Europe àgés de 5–6 ans appartenant aux générations riches de 1980–1981.
La composition par âge du chinchard ouest-africain est variable durant ces quelques dennières années; cependant, on peut distinguer une part considérable de jeunes individus appartenant aux générations de 1981–1982 et 1984 de naissance (tab. 3.4). L'augmentation de la part des jeunes dans les prises est conditionnée par de riches générations de ces années. La génération de 1984 par exemple est distinguée lors des travaux d'inventaire effectués en 1985. Une grande importance dans les prises appartient aux chinchards âés de plus de 2 ans.
3.4. La part du chinchard dans les captures (tab. 3.5)
Les proportions de chinchards dans les captures dans les divisions 34.1.3 et 34.3.1 reflètent leur répartition dans la région Centre-Est Atlantique. Le chinchard d'Europe domine dans la division 34.1.3. Le chinchard ouest-africain n'apparaît au large du Sahara (34.1.3) que dans les saisons et les années chaudes.
Dans la division 34.3.1, les proportions de ces deux espèces de chinchards sont presque égales, la part de chinchard ouest-africain étant un peu plus élevée. En règle générale, le chinchard d'Europe y domine dans les saisons et les années froides, et seulement dans la partie nord de la zone. L'importance du chinchard ouest-africain est très grande dans la partie sud de la zone.
3.5. Statistiques de pêche (tab. 3.6)
La capture totale des chinchards du genne Trachurus dans la division 34.1.3 se situe au diapason de 150–250 000 tonnes. L'absence de statistiques pour 1983–1984 ne permet pas d'évaluer précisément la capture totale des chinchards dans ces années; cependant, si l'on juge les pêches et les tendances des captures de chinchards au cours de ces dernières années, leur capture totale sera de 200 000 tonnes et pas moins.
L'absence de statistiques de pêches dans la division 34.3.1 pour la période 1983–1984 rend difficile l'analyse de statistiques totales et l'application de méthodes mathématiques de l'évaluation des stocks de chinchards.
4. REFERENCES BIBLIOGRAPHIQUES
KRIVOSPITCHENKO S.G. et DOMANEVSKY L.N., 1984. Le maquereau (Scomber japonicus). Recherches soviétiques sur les chinchards et les maquereaux de la zone Nord du COPACE. COPACE/TECH/84/59 (Fr.), Dakar, pp. 2–9, 22–24, 36–60.
MYLNIKOV N.I., OVERKO S.M. et DOMANEVSKY L.N., 1984. Le chinchard cunène (Tranchurus trecae Cadenat). Recherches soviétiques sur les chinchards et les maquereaux de la zone nord du COPACE. COPACE/TECH/85/59 (Fr.), Dakar, pp. 14–18, 78–86.
MYLNIKOV N.L., OVERKO S.M. et DOMANEVSKY L.N., 1984. Le chinchard d'Europe (Tranchurus trachurus). Recherches soviétiques sur les chinchards et les maquereaux de la zone Nord du COPACE. COPACE/TECH/85/59 (Fr.), Dakar, pp. 9–14, 61–77.
PLOKHINSKY N.A., 1970. Biométrie. Edition de l'Université de Moscou.
RICKER W.E., 1979. Méthodes de l'estimation et interprétation d'indices biologiques des populations de poissons, M., “pitcshevaya promychlennost”, 408 p.
| Taille cm | ANNEES | |||||
|---|---|---|---|---|---|---|
| 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | |
| 10 | 0,2 | |||||
| 11 | 2,1 | 0,1 | ||||
| 12 | 2,5 | 0,6 | ||||
| 13 | 4,3 | 0,0 | 0,0 | 0,6 | ||
| 14 | 5,9 | 0,0 | 2,1 | 0,6 | 1,1 | |
| 15 | 1,0 | 0,1 | 9,5 | 4,0 | 3,4 | |
| 16 | 0,1 | 0,2 | 8,3 | 0,3 | 6,2 | 3,6 |
| 17 | 0,2 | 0,5 | 7,1 | 1,4 | 5,7 | 4,1 |
| 18 | 0,6 | 1,7 | 6,9 | 5,0 | 8,7 | 8,0 |
| 19 | 1,5 | 4,0 | 4,4 | 11,1 | 10,6 | 4,8 |
| 20 | 5,1 | 5,0 | 4,9 | 15,0 | 6,9 | 8,4 |
| 21 | 7,5 | 4,5 | 4,4 | 15,0 | 4,7 | 8,2 |
| 22 | 7,8 | 4,4 | 5,0 | 13,3 | 1,5 | 11,3 |
| 23 | 5,8 | 3,5 | 5,6 | 10,9 | 1,4 | 12,0 |
| 24 | 2,7 | 2,9 | 6,7 | 7,0 | 1,5 | 8,5 |
| 25 | 3,2 | 3,5 | 13,1 | 5,0 | 2,6 | 5,6 |
| 26 | 4,7 | 3,6 | 7,7 | 3,6 | 2,6 | 3,8 |
| 27 | 6,2 | 4,6 | 3,1 | 2,8 | 3,2 | 2,2 |
| 28 | 5,8 | 5,2 | 2,0 | 1,9 | 4,4 | 1,8 |
| 29 | 5,6 | 4,7 | 1,1 | 0,9 | 5,2 | 1,1 |
| 30 | 6,0 | 6,7 | 0,8 | 0,4 | 6,0 | 2,0 |
| 31 | 4,9 | 7,8 | 0,6 | 0,3 | 6,4 | 2,6 |
| 32 | 4,7 | 7,9 | 1,2 | 0,5 | 5,4 | 2,0 |
| 33 | 4,1 | 6,5 | 0,8 | 0,7 | 4,8 | 1,2 |
| 34 | 3,4 | 4,7 | 1,4 | 1,1 | 3,2 | 1,2 |
| 35 | 1,9 | 5,2 | 0,7 | 0,9 | 2,1 | 0,8 |
| 36 | 1,2 | 4,4 | 0,7 | 0,7 | 0,9 | 0,4 |
| 37 | 0,4 | 2,9 | 1,0 | 0,6 | 0,6 | 0,2 |
| 38 | 0,2 | 2,5 | 0,3 | 0,4 | 0,4 | 0,2 |
| 39 | 0,2 | 1,4 | 0,3 | 0,4 | 0,2 | 0,1 |
| 40 | 0,1 | 0,8 | 0,1 | 0,3 | 0,1 | 0,0 |
| 41 | 0,1 | 0,5 | 0,1 | 0,3 | 0,1 | 0,1 |
| 42 | 0,0 | 0,2 | 0,1 | 0,1 | ||
| 43 | 0,1 | 0,0 | 0,1 | |||
| 44 | 0,0 | 0,0 | ||||
| total% | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 |
| nombre | ||||||
| échantil | 18978 | 27910 | 76493 | 19708 | 12067 | 7336 |
N.B. : Les données de 1985 sont préliminaires (Y compris le mois d'Aoot)
| AGE | ||||||
|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |
| 1972 | 26,90 | 33,20 | 25,60 | 12,10 | 1,90 | 0,30 |
| 1973 | 0,20 | 74,80 | 16,80 | 6,90 | 1,00 | 0,30 |
| 1974 | 36,60 | 47,80 | 13,50 | 1,40 | 0,50 | 0,10 |
| 1975 | 15,90 | 19,80 | 35,60 | 21,00 | 5,70 | 2,00 |
| 1976 | 37,60 | 26,70 | 24,20 | 7,40 | 2,30 | 1,80 |
| 1977 | 80,60 | 17,70 | 1,40 | 0,10 | 0,10 | 0,10 |
| 1978 | 26,00 | 37,60 | 24,80 | 8,60 | 2,20 | 0,80 |
| 1979 | 5,00 | 55,60 | 29,60 | 7,60 | 1,90 | 0,30 |
| 1980 | 37,50 | 28,70 | 27,90 | 4,90 | 0,80 | 0,20 |
| 1981 | 21,40 | 19,70 | 26,90 | 19,00 | 7,80 | 5,20 |
| 1982 | 59,00 | 31,60 | 4,90 | 2,70 | 1,40 | 0,40 |
| 1983 | 35,70 | 47,50 | 10,30 | 2,50 | 2,40 | 1,60 |
| 1984 | 47,30 | 18,00 | 15,30 | 12,70 | 5,80 | 0,90 |
| 1985 | 61,70 | 26,80 | 7,90 | 2,70 | 0,60 | 0,30 |
| Captures du maquereau | |||
|---|---|---|---|
| URSS | Autres pays | Total | |
| 1972 | 111765 | 16038 | 127803 |
| 1973 | 73110 | 9115 | 82225 |
| 1974 | 72429 | 6865 | 79294 |
| 1975 | 66689 | 13394 | 80083 |
| 1976 | 99768 | 3891 | 103659 |
| 1977 | 90089 | 8745 | 98834 |
| 1978 | 18211 | 4678 | 22889 |
| 1979 | 60073 | 8612 | 68685 |
| 1980 | 51766 | 5164 | 56930 |
| 1981 | 82588 | 3330 | 85918 |
| 1982 | 83917 | 3523 | 87440 |
| 1983 | 70060 | 3500* | 73560 |
| 1984 | 76749 | 3500* | 80249 |
| 1985 | 80000** | 3500* | 83500 |
** : au niveau des années 1981–1982
| Capture millers tonnes | AGE | captures millions individus | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| 1972 | 128,076 | 120,60 | 148,40 | 114,40 | 54,30 | 8,60 | 1,40 | 447,70 |
| 1973 | 82,225 | 0,60 | 213,70 | 47,90 | 19,60 | 2,90 | 0,80 | 285,50 |
| 1974 | 79,285 | 134,40 | 175,50 | 49,60 | 5,20 | 1,80 | 0,40 | 366,90 |
| 1975 | 80,584 | 34,50 | 43,00 | 77,40 | 45,50 | 12,30 | 4,40 | 217,10 |
| 1976 | 103,659 | 145,00 | 103,00 | 93,40 | 28,50 | 8,90 | 7,00 | 385,80 |
| 1977 | 98,843 | 604,90 | 130,70 | 10,40 | 0,70 | 0,70 | 0,70 | 748,10 |
| 1978 | 22,889 | 21,20 | 29,90 | 20,20 | 7,00 | 1,80 | 0,60 | 80,70 |
| 1979 | 68,685 | 11,10 | 123,80 | 65,90 | 16,90 | 4,20 | 0,70 | 222,60 |
| 1980 | 59,930 | 71,80 | 54,90 | 53,40 | 9,40 | 1,50 | 0,40 | 191,40 |
| 1981 | 85,918 | 48,20 | 44,40 | 60,60 | 42,80 | 17,60 | 11,70 | 225,30 |
| 1982 | 87,440 | 283,80 | 152,00 | 23,60 | 13,00 | 6,70 | 1,90 | 481,00 |
| 1983 | 73,560 | 134,60 | 179,30 | 38,80 | 9,40 | 9,10 | 6,00 | 377,20 |
| 1984 | 80,249 | 140,70 | 53,50 | 45,50 | 37,80 | 17,20 | 2,70 | 297,40 |
| 1985 | 83,500 | 288,70 | 125,30 | 36,90 | 12,60 | 2,80 | 1,40 | 467,70 |
| AGE | 2-6 | 1-6 | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| 1972 | 1438,10 | 570,47 | 255,07 | 82,65 | 14,04 | 2,33 | 924,56 | 2362,66 |
| 1973 | 1042,57 | 699,94 | 203,14 | 55,23 | 5,13 | 1,33 | 964,77 | 2007,34 |
| 1974 | 761,71 | 571,77 | 225,82 | 76,00 | 15,79 | 0,67 | 890,05 | 1651,76 |
| 1975 | 479,56 | 318,48 | 183,78 | 87,19 | 37,86 | 7,33 | 634,64 | 1114,20 |
| 1976 | 1159,61 | 237,64 | 142,94 | 43,52 | 14,14 | 11,67 | 449,91 | 1609,52 |
| 1977 | 1587,68 | 529,01 | 54,11 | 9,24 | 2,77 | 1,17 | 596,30 | 2183,98 |
| 1978 | 1222,40 | 423,20 | 193,51 | 21,99 | 4,56 | 1,00 | 644,26 | 1866,66 |
| 1979 | 753,50 | 655,20 | 210,12 | 91,24 | 6,88 | 1,17 | 964,61 | 1718,11 |
| 1980 | 691,40 | 405,33 | 267,88 | 66,49 | 37,55 | 0,67 | 777,92 | 1469,32 |
| 1981 | 965,56 | 326,28 | 181,79 | 107,46 | 29,53 | 19,50 | 664,56 | 1630,12 |
| 1982 | 1155,08 | 494,23 | 146,18 | 54,87 | 27,26 | 3,17 | 725,71 | 1880,79 |
| 1983 | 557,80 | 423,68 | 158,64 | 62,74 | 20,48 | 10,00 | 675,54 | 1233,34 |
| 1984 | 760,69 | 206,42 | 99,69 | 58,32 | 27,47 | 4,50 | 396,40 | 1157,09 |
| 1985 | 1443,50 | 313,25 | 73,80 | 21,00 | 4,00 | 2,33 | 414,38 | 1857,88 |
| AGE | 2–6 | 1–6 | ||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | |||
| 1972 | 149,56 | 131,78 | 98,71 | 45,79 | 10,08 | 2,03 | 288,39 | 437,95 |
| 1973 | 108,43 | 161,69 | 78,62 | 30,60 | 3,68 | 1,16 | 275,75 | 384,18 |
| 1974 | 79,22 | 132,08 | 87,39 | 42,10 | 11,34 | 0,58 | 273,49 | 352,71 |
| 1975 | 49,87 | 73,57 | 71,12 | 48,30 | 27,18 | 6,39 | 226,56 | 276,43 |
| 1976 | 120,60 | 54,89 | 55,32 | 24,11 | 10,15 | 10,18 | 154,65 | 275,25 |
| 1977 | 165,12 | 122,20 | 20,94 | 5,12 | 1,99 | 1,02 | 151,27 | 316,39 |
| 1978 | 127,13 | 97,76 | 74,89 | 12,18 | 3,27 | 0,87 | 188,97 | 316,10 |
| 1979 | 78,36 | 151,35 | 81,32 | 50,55 | 4,94 | 1,02 | 289,18 | 367,54 |
| 1980 | 71,91 | 93,63 | 103,67 | 36,84 | 26,96 | 0,58 | 264,68 | 336,59 |
| 1981 | 100,42 | 75,37 | 70,35 | 59,53 | 21,20 | 17,00 | 243,45 | 343,87 |
| 1982 | 120,13 | 114,17 | 56,57 | 30,40 | 19,57 | 2,76 | 223,47 | 343,60 |
| 1983 | 58,01 | 97,87 | 61,39 | 34,76 | 14,70 | 8,72 | 217,44 | 275,45 |
| 1984 | 79,11 | 47,68 | 38,58 | 32,31 | 19,72 | 3,92 | 142,21 | 221,32 |
| 1985 | 150,12 | 72,36 | 28,56 | 11,63 | 2,87 | 2,03 | 117,45 | 267,57 |
| Taille cm | ANNEES | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1972 | 1973 | 1974 | 1975 | 1976 | 1977 | 1978 | 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985* | |
| 9 | 0,2 | + | ||||||||||||
| 10 | 0,1 | 0,2 | 1,2 | 0,3 | 0,1 | 0,1 | 0,6 | |||||||
| 11 | 0,7 | 0,4 | 3,3 | 0,1 | 1,3 | 1,5 | 0,9 | 3,9 | ||||||
| 12 | 2,7 | 0,1 | 0,3 | 1,0 | 6,6 | 0,1 | 1,3 | 3,8 | 0,9 | 5,2 | ||||
| 13 | 5,1 | 0,1 | 2,0 | 5,3 | 9,1 | 0,9 | 0,3 | 2,8 | 3,8 | 0,1 | 2,6 | |||
| 14 | 10,5 | 0,6 | 2,8 | 14,7 | 7,3 | 0,1 | 0,1 | 0,4 | 1,3 | 2,7 | 7,8 | 0,1 | 0,8 | |
| 15 | 9,2 | 1,6 | 6,2 | 18,1 | 4,7 | 0,8 | 0,1 | 0,6 | 1,7 | 2,4 | 8,7 | 0,2 | 1,3 | |
| 16 | 7,8 | 1,8 | 9,2 | 12,5 | 9,1 | 2,0 | 0,1 | 0,8 | 1,8 | 1,8 | 11,1 | 0,4 | 2,4 | |
| 17 | 7,0 | 1,8 | 5,6 | 11,0 | 16,7 | 4,2 | 0,8 | 0,4 | 0,5 | 4,0 | 1,2 | 13,2 | 1,6 | 3,7 |
| 18 | 5,9 | 3,6 | 6,3 | 7,8 | 13,9 | 6,6 | 0,7 | 0,8 | 0,9 | 7,8 | 1,3 | 12,9 | 3,2 | 6,8 |
| 19 | 6,1 | 7,3 | 7,2 | 6,2 | 11,0 | 11,2 | 2,7 | 1,8 | 3,9 | 11,3 | 4,1 | 14,4 | 4,0 | 14,1 |
| 20 | 7,6 | 13,6 | 8,3 | 4,2 | 7,0 | 13,8 | 23,5 | 2,5 | 7,5 | 10,4 | 5,3 | 12,0 | 2,8 | 16,8 |
| 21 | 6,4 | 13,2 | 9,4 | 3,2 | 4,5 | 11,3 | 39,2 | 3,5 | 8,4 | 8,4 | 10,6 | 6,2 | 3,3 | 11,7 |
| 22 | 4,9 | 8,7 | 12,4 | 2,8 | 2,2 | 6,2 | 23,6 | 3,0 | 4,9 | 7,7 | 14,3 | 2,8 | 3,1 | 8,2 |
| 23 | 3,6 | 10,5 | 11,1 | 1,9 | 1,3 | 14,6 | 7,6 | 4,8 | 4,1 | 6,5 | 11,6 | 1,6 | 4,6 | 4,2 |
| 24 | 2,6 | 7,7 | 5,7 | 1,7 | 0,9 | 16,5 | 1,0 | 9,2 | 6,5 | 5,2 | 11,2 | 1,0 | 3,6 | 3,9 |
| 25 | 1,8 | 5,9 | 3,0 | 1,2 | 0,4 | 8,7 | 0,7 | 14,0 | 15,0 | 5,3 | 5,2 | 0,5 | 6,7 | 4,0 |
| 26 | 1,5 | 4,8 | 1,8 | 1,4 | 0,2 | 2,4 | 0,1 | 32,8 | 19,0 | 3,7 | 2,7 | 0,4 | 4,0 | 2,9 |
| 27 | 1,6 | 3,2 | 1,4 | 1,1 | 0,1 | 0,8 | 20,8 | 9,5 | 3,4 | 1,6 | 0,4 | 3,3 | 2,3 | |
| 28 | 2,0 | 3,3 | 1,0 | 0,5 | 0,1 | 0,2 | 5,0 | 4,3 | 6,4 | 1,9 | 0,3 | 2,5 | 1,7 | |
| 29 | 2,1 | 2,7 | 1,1 | 0,8 | 0,2 | 0,7 | 1,9 | 4,9 | 1,9 | 0,2 | 2,1 | 1,1 | ||
| 30 | 4,1 | 3,2 | 1,2 | 1,2 | 0,2 | 0,1 | 1,5 | 4,5 | 1,8 | 0,2 | 4,8 | 0,6 | ||
| 31 | 3,8 | 3,1 | 1,2 | 1,0 | 0,2 | 0,1 | 1,0 | 2,5 | 1,6 | 0,2 | 7,2 | 0,4 | ||
| 32 | 2,4 | 2,6 | 1,1 | 0,8 | 0,1 | 0,1 | 0,1 | 1,5 | 1,9 | 1,7 | 0,2 | 17,5 | 0,3 | |
| 33 | 0,9 | 0,5 | 1,1 | 0,3 | 0,1 | 0,1 | 1,6 | 0,6 | 1,6 | 0,1 | 11,0 | 0,3 | ||
| 34 | 0,2 | 0,2 | 0,5 | 0,3 | 0,1 | 1,5 | 0,3 | 1,7 | 0,1 | 9,8 | 0,1 | |||
| 35 | 0,1 | 0,1 | 0,1 | 0,8 | 0,1 | 1,4 | 3,0 | 0,1 | ||||||
| 36 | 0,1 | 0,1 | 1,1 | 1,1 | + | |||||||||
| 37 | 0,1 | 0,6 | + | |||||||||||
| 38 | 0,3 | + | ||||||||||||
| 39 | 0,2 | |||||||||||||
| total | 100,6 | 100,2 | 100,0 | 99,9 | 99,8 | 100,3 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 | 100,0 |
| nombre | ||||||||||||||
| échant | 74077 | 61959 | 22361 | 63931 | 27201 | 18683 | 6501 | 9649 | 19172 | 24401 | 32366 | 67322 | 46579 | 41301 |
| Taille cm | ANNEES | ||||||
|---|---|---|---|---|---|---|---|
| 1979 | 1980 | 1981 | 1982 | 1983 | 1984 | 1985 | |
| 10 | |||||||
| 11 | 0,4 | ||||||
| 12 | 0,1 | 0,1 | 1,2 | ||||
| 13 | 0,2 | 0,3 | 0,5 | 3,8 | 0,1 | ||
| 14 | 1,7 | 1,5 | 0,1 | 5,9 | 0,5 | 0,2 | |
| 15 | 2,0 | 1,9 | 0,1 | 4,0 | 0,9 | 1,0 | |
| 16 | 0,5 | 2,0 | 0,8 | 2,4 | 2,0 | 0,2 | 1,9 |
| 17 | 0,4 | 2,3 | 3,9 | 1,5 | 3,5 | 0,7 | 3,2 |
| 18 | 1,3 | 1,1 | 3,5 | 1,0 | 3,8 | 4,9 | 6,1 |
| 19 | 2,8 | 0,5 | 4,3 | 1,1 | 3,9 | 21,9 | 6,5 |
| 20 | 5,8 | 0,5 | 4,7 | 2,9 | 3,3 | 29,9 | 5,7 |
| 21 | 4,5 | 1,9 | 5,4 | 2,8 | 2,1 | 11,5 | 6,3 |
| 22 | 2,5 | 3,9 | 6,7 | 0,7 | 2,1 | 9,8 | 6,6 |
| 23 | 1,6 | 7,3 | 9,8 | 2,1 | 3,1 | 6,0 | 8,8 |
| 24 | 2,5 | 10,8 | 8,9 | 3,2 | 3,3 | 5,8 | 10,3 |
| 25 | 5,1 | 9,7 | 5,7 | 4,6 | 3,4 | 2,4 | 10,3 |
| 26 | 11,9 | 7,1 | 3,9 | 5,1 | 2,5 | 1,3 | 7,1 |
| 27 | 10,6 | 6,9 | 3,1 | 2,7 | 3,2 | 1,3 | 3,9 |
| 28 | 10,4 | 11,2 | 3,0 | 4,2 | 5,8 | 2,7 | 3,1 |
| 29 | 12,3 | 10,7 | 4,9 | 5,6 | 9,7 | 0,9 | 4,4 |
| 30 | 14,4 | 11,4 | 8,0 | 11,4 | 11,3 | 0,4 | 5,5 |
| 31 | 5,9 | 4,7 | 8,5 | 14,5 | 19,6 | 0,6 | 3,2 |
| 32 | 2,0 | 2,0 | 6,4 | 12,1 | 8,8 | 2,3 | |
| 33 | 1,0 | 1,0 | 3,9 | 4,5 | 6,6 | 1,6 | |
| 34 | 0,4 | 0,7 | 2,1 | 1,5 | 5,0 | 1,1 | |
| 35 | 0,2 | 0,3 | 1,0 | 0,6 | 2,9 | 0,6 | |
| 36 | 0,1 | 0,5 | 0,2 | 1,3 | 0,1 | ||
| 37 | 0,1 | 0,1 | 0,8 | ||||
| 38 | 0,1 | 0,4 | 0,1 | ||||
| 39 | 0,2 | ||||||
| total % | 100,0 | 100,0 | 100,0 | 100,0 | 110,0 | 100,3 | 100,0 |
| ?? | |||||||
| nombre échantil | 16529 | 25419 | 23667 | 13899 | 29079 | 12476 | 18301 |
N.B. : En 1981, les données roumaines sont aussi utilisées; le nombre d'échantillons mesurés est inconnu.
En 1985,les données ne concernent que le premier semestre.
En 1983, les donnees sont sujettes à caution (total %=110)
| AGE | ||||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 1972 | 49,40 | 26,60 | 7,30 | 5,60 | 6,10 | 3,90 | 1,00 | 0,10 |
| 1973 | 13,30 | 47,50 | 19,60 | 9,30 | 5,90 | 3,60 | 0,80 | 0,10 |
| 1974 | 33,90 | 43,20 | 13,80 | 3,80 | 2,20 | 1,80 | 1,10 | 0,20 |
| 1975 | 70,40 | 18,50 | 4,70 | 2,40 | 1,80 | 1,20 | 0,60 | 0,30 |
| 1976 | 71,30 | 26,10 | 1,90 | 0,40 | 0,10 | 0,20 | 0,00 | 0,00 |
| 1977 | 9,10 | 63,60 | 26,30 | 0,50 | 0,30 | 0,20 | 0,00 | 0,00 |
| 1978 | 7,00 | 84,20 | 8,70 | 0,10 | 0,00 | 0,00 | 0,00 | 0,00 |
| 1979 | 1,60 | 16,30 | 50,50 | 29,00 | 2,50 | 0,10 | 0,00 | 0,00 |
| 1980 | 11,30 | 36,10 | 26,60 | 17,40 | 5,40 | 1,80 | 1,20 | 0,20 |
| 1981 | 11,50 | 54,50 | 12,90 | 9,30 | 8,90 | 2,80 | 0,10 | 0,00 |
| 1982 | 19,80 | 36,50 | 27,10 | 4,30 | 4,60 | 2,10 | 1,50 | 4,10 |
| 1983 | 62,50 | 33,90 | 2,30 | 0,60 | 0,40 | 0,20 | 0,10 | 0,00 |
| 1984 | 6,70 | 14,80 | 17,70 | 7,30 | 21,10 | 16,30 | 8,80 | 7,30 |
| 1985 | 33,90 | 43,00 | 17,30 | 3,40 | 1,60 | 0,50 | 0,20 | 0,10 |
| AGE | ||||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
| 1979 | 6,10 | 17,20 | 19,50 | 33,30 | 17,30 | 5,00 | 1,40 | 0,20 |
| 1980 | 9,20 | 14,10 | 27,60 | 28,80 | 13,70 | 4,40 | 1,70 | 0,50 |
| 1981 | 9,00 | 30,90 | 18,50 | 11,00 | 12,20 | 10,70 | 6,00 | 1,70 |
| 1982 | 20,20 | 9,60 | 12,90 | 12,50 | 18,60 | 19,40 | 6,00 | 0,80 |
| 1983 | 10,70 | 14,50 | 9,20 | 18,70 | 16,10 | 13,60 | 11,60 | 5,60 |
| 1984 | 5,80 | 79,10 | 9,50 | 4,90 | 0,50 | 0,20 | 0,00 | 0,00 |
| 1985 | 12,50 | 33,90 | 27,70 | 11,40 | 7,10 | 3,90 | 2,70 | 0,80 |
N.B. Pour 1985 les données ne concernent que le premier semestre
| ANNEES | |||||||
|---|---|---|---|---|---|---|---|
| Région | Espèce | 1979 | 1980 | 1981 | 1982 | 1983 | 1984 |
| Sahara | T.trachurus | 56 | 89 | 52 | 82 | 97 | 91 |
| 34.1.3 | T.trecae | 38 | 10 | 44 | 16 | 2 | 8 |
| Cap Vert | T.trachurus | 30 | 30 | 30 | 30 | 39 | 48 |
| 34.3.1 | T.trecae | 60 | 60 | 60 | 60 | 48 | 35 |
N.B. : En l'absence d'observations de 1979 à 1982 dans la division 34.3.1 la répartition spécifique dans la région du Cap Vert est arbitraire
| ANNEES | |||||||
|---|---|---|---|---|---|---|---|
| Région | Pays | 1979 | 1980 | 1981 | 1982 | 1983 | 1984 |
| Sahara | URSS | 104730 | 141376 | 148208 | 157614 | 163614 | 168118 |
| 34.1.3 | autres | 49518 | 107843 | 70598 | 44883 | * | * |
| Total | 154248 | 249219 | 218806 | 202497 | * | * | |
| Cap Vert | URSS | 25783 | 90154 | 33858 | 23493 | 4148 | 703 |
| 34.3.1 | autres | 39991 | 75160 | 89355 | 84474 | * | * |
| Total | 65774 | 165314 | 123213 | 107967 | * | * | |
| Total | Total | 220022 | 414533 | 342019 | 310464 | * | * |
La base des recherches de la situation et du développement des stocks de chinchards et maquereaux dans la zone du COPACE ont été les travaux systématiques et l'échantillonnage permanent à bord des bateaux industriels de pêche qui ont opéré dans les eaux mauritaniennes. L'échantillonnage a compris des travaux relatifs à la composition des tailles , du poids, de la composition des sexes et de la maturité des gonades,ainsi que du contenu de l'estomac et des intestins.
Normalement, les missions biologiques à bord des chalutiers-usine (type super-trawler 3) ont été réalisées au printemps, en été et en automne.
Les résultats scientifiques ont étépubliés dans différentes publications.
Il faut mentionner que depuisle printemps 1982 en coopération entre le CNROP de la République Islamique de Mauritanie et l'Institut de la Pêche Maritime (ITP) de la RDA à Rostock ont été réalisées des recherches hydroacoustiques et des prospections par chaluts au moyen de navires océanographiques. Les campagnes ont commencé par la mission conjointe RIM/RDA par le N/O "Ernst Haeckel" et le chalutier “Walter Barth” en mars et avril 1982 et continué au printemps 1983 par les N/O “Eisbar”,“Alexander von Humbolt” (Académie des sciences de la RDA) et “Walter Barth”. Pendant la deuxième campagne une prospection hydroacoustique par intégrateur de tout le plateau continental jusqu'à la zone océanique a été conduite. Simultanément des recherches sur le milieu océanique ont été réalisées. Une attention spéciale durant cette mission a été donnée aux juvéniles de chinchards et maquereaux.
Pendant une autre mission pour la recherche des crevettes dans les eaux mauritaniennes faite en novembre1983 (chalutier "Karl Wolff") ont été pris des échantillons de chinchards.
En mars et avril 1984 avec la participation de l'Institut d'Océanographie (IFM), de l'ITP et du CNROP de Nouadibhou, le N/O “Alexander von Humbolt” et des chalutiers industriels ont réalisé un programme de “l'investigation des fosses sous-marines” dans la région du cap Timiris, qui s'occupait de l'influence de la fosse de Timiris sur l'upwelling local et par conséquence sur la nourriture et les poissons, dont un des plus importants a été T.trachurus (L.). Les résultats seront publiés dans un article de “Beiträge für Meereskunde 1987”.
En complément aux travaux de recherches dans la zone économique de la Mauritanie nous avons pu analyser quelques échantillons de T.trachurus et de S.japonicus provenant de la pêche des bateaux qui opèrent dans les eaux marocaines sur la base d'une société mixte.
by
S.Holzlohner
Institute of Deep Sea Fishery and Fish Processing,
Rostock,GDR
In the years 1981–1985 the GDR performed a licence fishery in the EEZ of Mauritania (CECAF 34.1.3 and 34.3.1). Taking part were 2–4 FVS III (3 880 HP), and additionally, until the first half year 1983, Zubringertrawler (1 750 HP) together with a mothership.
The total catch as well as the catch of horsemackerel and chubmackerel is shown in table 1. The official landing statistics are based on the recalculation from the production figures and only give data for horsemackerels together. But for the years 1980–1983 estimations by species analyses for the three dominant horsemackerels Trachurus trachurus,Trachurus trecae and Decapterus rhonchus are available. The decline of total catch in 1981–1985 is mainly due to reduction of the fishing effort.
Table 2 shows the total catches as well as the catches of horsemackerels and chubmackerels of Mauritania by CECAF divisions 34.1.3 and 34.3.1. The majority of the horsemackerel catches was taken in all reporting years in division 34.3.1. The catches of chubmackerels were in the years 1981,1983,1984 within the division 34.3.1 higher than in division 34.1.3, but were equal in the years 1982 and 1985.
In table 3 and figure 1 the catch per fishing day for all species of horsemackerel and chubmackerel is presented on the basis of FVS III. The data show for the sum of all species as well as for the horsemackerel a strong drop in 1981–1983 and a strong ascent in 1983–1984. In 1985 the data given increased slightly in comparison with 1984. The catch per fishing day for chubmackerel shows,with the exception of the decline of 1985 in comparison to 1984, the tendencies given for horsemackerel. The catch per fishing day data given in figure 1 for the second important group of Sardinella spp. demonstrate a progressive ascent in 1981–1984 and in 1985 nearly at the level of 1984.
The catch per fishing hour and the sum of fishing hours for FVS III given in table 4 show analogous tendencies as the catch per fishing day in 1981–1985.
The separation of the horsemackerel into species in the years 1981–1983 is given in table 5. For the two Trachurus species the decline of the catch per unit effort results from the same tendency already mentioned for all horsemackerels. Obviously there is a strong drop of Trachurus trecae 1982 in comparison with 1981. The value for Decapterus rhonchus fluctuated without a perceptible tendency.
Tables 6,7 and 9 as well as figure 2 give a common view of the catch per fishing day for the total of all species, for horsemackerels and for chubmackerel in the annual cycle of the years 1981–1985. In figure 2 is added the catch per unit effort for Sardinella species, the second important fish group.
In the average of the years 1981–1985 there results a curve with two peaks for the total of all species, for horsemackerels and chubmackerel. There is a main peak for the total of all species, for horsemackerels together and for chubmackerel in June. A second peak is for the total of all species and for horsemackerels in January and for chubmackerel in December.
Table 8 and figure 3 show the separation of horsemackerels into species in the years 1981–1983. The theoretic main peak for horsemackerels splits into a May peak for Trachurus trecae, in an April peak for Trachurus trachurus and in a July peak for Decapterus rhonchus.
The annual cycle with two peaks obviously is true among the horsemackerel species only for Trachurus trecae, as for Scomber japonicus. The spring peak is interpreted for both species mainly as migration through the EEZ of Mauritania from south to north and the late autumn peak as migration in the opposite direction.
Trachurus trachurus as a low temperature-adapted species obviously concentrates off Mauritania mainly from January to April/May and in the other months stays mostly outside the northern borders of EEZ of Mauritania. Compared to Trachurus trachurus, the species Decapterus rhonchus as a ward temperatureadapted species obviously migrates in the EEZ of Mauritania in the warmest months July to October from the south. An analogous annual cycle shows groups of Sardinella species, given for comparison.
Table 10 shows the latitude of the fishery off Mauritania 1981–1984 in the annual cycle. It results that in nearly all months of all years the ranges extend through division 34.1.3 as well as division 34.3.1 (border 19°N). Therefore a statistical separation of the catches in the Mauritanian EEZ in the two CECAF divisions is difficult. In the average the fishery in the first half year was more southerly than in the second half year. But in all months of all years, with exception of June, the fleet fishes also north of 20°N.
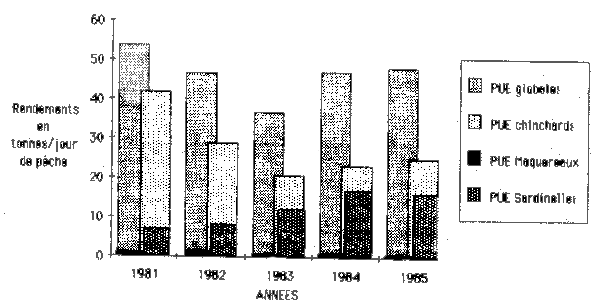
figure 1 : Evolution des rendements en tonnes/jour de mer des FVS III
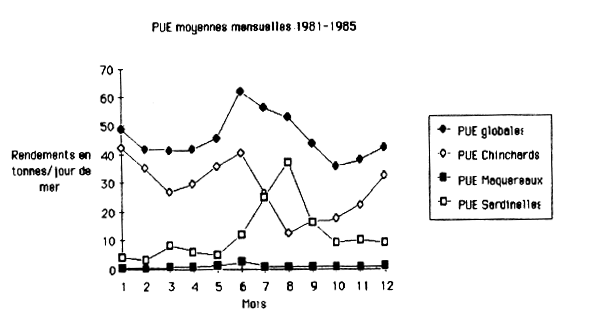
figure 2: PUE moyennes mensuelles de 1981 à 1985 en tonnes/jour de mer pour les FVS III
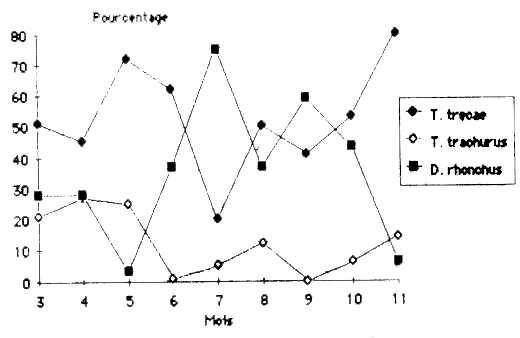
figure 3: Répartition specifique moyenne en 1981 – 1983 des espèces de chinchards
| ANNEES | |||||
|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | 1984 | 1985 | |
| Total catch | 103,9 | 95,1 | 50,8 | 29,2 | 20,9 |
| Horsemackerels from it : | 89,1 | 71,3 | 29,9 | 14,5 | 10,9 |
| Trachurus trachurus | 10,7 | 10,7 | 2,4 | ||
| Trachurus trecae | 65,0 | 38,5 | 22,3 | ||
| Decapterus rhonchus | 13,4 | 22,1 | 5,3 | ||
| chubmackerel | 1,2 | 1,8 | 0,9 | 0,7 | 0,2 |
| ANNEES | |||||
|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | 1984 | 1985 | |
| Total catch | |||||
| 34.1.3. | 42,4 | 39,1 | 14,0 | 8,9 | 4,2 |
| 34.3.1. | 61,5 | 55,8 | 36,8 | 20,3 | 16,6 |
| Horsemackerels | |||||
| 34.1.3. | 34,7 | 25,5 | 7,3 | 4,2 | 1,6 |
| 34.3.1. | 54,4 | 45,9 | 22,6 | 10,3 | 9,3 |
| chubmackerel | |||||
| 34.1.3. | 0,4 | 0,9 | 0,2 | 0,2 | 0,1 |
| 34.3.1. | 0,8 | 0,9 | 0,7 | 0,3 | 0,1 |
| ANNEES | |||||
|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | 1984 | 1985 | |
| Catch/fishing day | |||||
| alltogether | 53,5 | 46,5 | 36,5 | 47,0 | 48,1 |
| Horsemackerels | 41,6 | 28,6 | 20,4 | 23,3 | 25,1 |
| Chubmackerel | 1,0 | 1,3 | 0,7 | 0,8 | 0,4 |
| Number of fishing days | 1224 | 1296 | 996 | 620 | 432 |
| ANNEES | |||||
|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | 1984 | 1985 | |
| Catch/fishing hour | |||||
| alltogether | 4,8 | 4,7 | 3,4 | 4,5 | 4,7 |
| Horsemackerels | 3,6 | 2,8 | 1,9 | 2,3 | 2,4 |
| Chubmackerel | 0,1 | 0,1 | 0,1 | 0,1 | + |
| Number of fishing hours | 14076 | 13330 | 10588 | 6409 | 4464 |
| ANNEESS | |||||
|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | X * | squ. dev. | |
| Trachurus trachurus | 30,4 | 15,4 | 15,2 | 20,3 | 7,1 |
| Trachurus trecae | 5,0 | 4,3 | 1,6 | 3,6 | 1,5 |
| Decapterus rhonchus | 6,2 | 8,9 | 3,6 | 6,2 | 2,2 |
| Catch per fishing day (t) | Number of fishing days | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | 1984 | 1985 | mean * | squ. dev. | 1981 | 1982 | 1983 | 1984 | 1985 | |
| Jan. | 57,1 | 55,4 | 35,7 | 45,0 | 50,3 | 48,7 | 7,8 | 114,0 | 107,0 | 126,0 | 49,0 | 51,0 |
| Febr. | 46,4 | 38,8 | 27,2 | 52,1 | 43,5 | 41,6 | 8,4 | 81,0 | 87,0 | 87,0 | 44,0 | 46,0 |
| March | 48,9 | 40,5 | 29,8 | 46,4 | (19,4) | ** 41,4 | 7,4 | 111,0 | 105,0 | 108,0 | 51,0 | 11,0 |
| April | 50,9 | 37,5 | 28,1 | 46,4 | 45,3 | 41,6 | 8,0 | 102,0 | 108,0 | 65,0 | 52,0 | 26,0 |
| May | 60,6 | 51,2 | 40,4 | 39,1 | 37,0 | 45,7 | 8,9 | 101,0 | 110,0 | 71,0 | 60,0 | 26,0 |
| June | 73,6 | 68,7 | 51,0 | 63,8 | 52,7 | 62,0 | 8,8 | 92,0 | 107,0 | 81,0 | 44,0 | 42,0 |
| July | 70,8 | 53,4 | 39,7 | 59,3 | 59,0 | 56,4 | 10,1 | 103,0 | 99,0 | 56,0 | 55,0 | 54,0 |
| August | 53,4 | 54,1 | 47,7 | 50,0 | 58,7 | 52,8 | 3,8 | 110,0 | 113,0 | 84,0 | 56,0 | 53,0 |
| Sept. | 46,7 | 48,2 | 40,0 | 36,7 | 47,2 | 43,8 | 4,6 | 94,0 | 108,0 | 74,0 | 54,0 | 45,0 |
| Oct | 43,4 | 38,1 | 28,2 | 33,4 | 31,2 | 35,8 | 5,6 | 100,0 | 104,0 | 81,0 | 57,0 | 27,0 |
| Nov | 35,1 | 40,0 | 33,0 | 46,2 | 35,5 | 38,0 | 4,7 | 102,0 | 109,0 | 80,0 | 47,0 | 24,0 |
| Dec. | 54,8 | 32,1 | 37,7 | 51,3 | 56,6 | 42,2 | 8,8 | 108,0 | 139,0 | 83,0 | 51,0 | 27,0 |
** without consideration of the value of 1985
| Catch per fishing day (t) | |||||||
|---|---|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | 1984 | 1985 | mean * | squ.dev. | |
| Jan. | 55,7 | 50,1 | 27,6 | 42,8 | 34,9 | 42,2 | 10,1 |
| Febr. | 42,3 | 30,4 | 19,0 | 50,7 | 32,4 | 35,1 | 10,8 |
| March | 39,8 | 29,0 | 15,7 | 22,3 | (16,8) | ** 26,7 | 8,9 |
| April | 48,4 | 28,5 | 14,9 | 28,7 | 27,2 | 29,5 | 10,7 |
| May | 57,2 | 43,4 | 28,0 | 26,2 | 23,6 | 35,7 | 12,8 |
| June | 60,5 | 57,3 | 26,4 | 24,0 | 34,4 | 40,5 | 15,4 |
| July | 51,3 | 31,9 | 8,0 | 15,7 | 25,6 | 26,5 | 14,9 |
| August | 16,8 | 7,8 | 14,4 | 10,5 | 11,7 | 12,2 | 3,1 |
| Sept. | 26,0 | 7,6 | 26,6 | 10,2 | 12,5 | 16,6 | 8,1 |
| Oct | 29,5 | 15,9 | 13,0 | 10,6 | 18,6 | 17,5 | 6,6 |
| Nov | 26,5 | 25,3 | 24,6 | 20,3 | 14,4 | 22,2 | 4,4 |
| Dec. | 45,4 | 16,5 | 27,0 | 26,0 | 47,2 | 32,4 | 11,9 |
** without consideration of the value of 1985
| ANNEE | Espèce/Mois | III | IV | V | VI | VII | VIII | XI | X | XI |
|---|---|---|---|---|---|---|---|---|---|---|
| 1981 | Trachurus trecae | 46 | 50 | 72 | 85 | 30 | 17 | 3 | 75 | 82 |
| Trachurus trachurus | 15 | 21 | 25 | 3 | 11 | 26 | 0 | 13 | 4 | |
| Decapterus rhonchus | 39 | 29 | 3 | 12 | 59 | 57 | 97 | 12 | 14 | |
| 1982 | Trachurus trecae | 29 | 21 | 39 | 30 | 90 | 79 | 32 | 63 | |
| Trachurus trachurus | 39 | 51 | 0 | 4 | 0 | 0 | 4 | 37 | ||
| Decapterus rhonchus | 32 | 28 | 61 | 66 | 10 | 21 | 64 | 0 | ||
| 1983 | Trachurus trecae | 79 | 63 | 0 | 44 | 48 | 97 | |||
| Trachurus trachurus | 9 | 8 | 0 | 11 | 0 | 0 | ||||
| Decapterus rhonchus | 12 | 29 | 100 | 45 | 52 | 3 | ||||
| mean | Trachurus trecae | 51 | 45 | (72) | 62 | 20 | 50 | 41 | 53 | 80 |
| Trachurus trachurus | 21 | 27 | (25) | 1 | 5 | 12 | 0 | 6 | 14 | |
| Decapterus rhonchus | 28 | 28 | (3) | 37 | 75 | 37 | 59 | 43 | 6 | |
| sq.dev | Trachurus trecae | 21 | 18 | 23 | 14 | 30 | 38 | 18 | 14 | |
| Trachurus trachurus | 13 | 18 | 15 | 5 | 11 | 0 | 2 | 17 | ||
| Decapterus rhonchus | 11 | 13 | 25 | 18 | 20 | 38 | 22 | 6 |
| Catch per fishing day (t) | |||||||
|---|---|---|---|---|---|---|---|
| 1981 | 1982 | 1983 | 1984 | 1985 | mean* | squ.dev. | |
| Jan. | 0,5 | 0,6 | 0,2 | 0,4 | 0,1 | 0,4 | 0,2 |
| Febr. | 0,1 | 0,7 | 0,3 | 0,1 | 0,2 | 0,3 | 0,2 |
| March | 0,4 | 1,6 | 0,2 | 0,4 | 0,2 | **0,7 | 0,6 |
| April | 0,3 | 0,4 | 0,4 | 1,3 | 0,9 | 0,7 | 0,4 |
| May | 1,4 | 0,9 | 0,8 | 3,0 | 0,8 | 1,4 | 0,8 |
| June | 5,0 | 2,3 | 1,8 | 2,5 | 1,1 | 2,5 | 1,3 |
| July | 1,7 | 1,1 | 0,4 | 0,3 | 0,3 | 0,8 | 0,6 |
| August | 0,2 | 1,8 | 0,9 | 0,1 | - | 0,6 | 0,7 |
| Sept. | 0,1 | 2,6 | 0,2 | - | - | 0,9 | 1,2 |
| Oct | 0,6 | 1,9 | 0,3 | 0,1 | - | 0,7 | 0,7 |
| Nov | 0,6 | 1,4 | 1,0 | 0,1 | 1,2 | 0,9 | 0,5 |
| Dec. | 1,1 | 1,1 | 1,5 | 1,4 | 1,4 | 1,1 | 0,3 |
** without consideration of the value of 1985
| 1981 | 1982 | 1983 | 1984 | |
|---|---|---|---|---|
| Jan | 16,01–19,05 | 17,30–20,10 | 16,12–19,40 | 16,10–20,35 |
| Febr. | 17,37–20,12 | 16,40–20,36 | 16,10–19,34 | 17,45–19,01 |
| March | 16,14–19,40 | 16,17–20,01 | 16,12–19,03 | 16,40–20,35 |
| April | 16,15–20,30 | 16,44–19,40 | 16,28–19,40 | 18,13–20,35 |
| May | 16,29–19,39 | 16,10–20,36 | 16,05–18,00 | 16,50–19,10 |
| June | 17,25–19,57 | 16,18–19,40 | 16,22–19,24 | 16,25–19,40 |
| July | 18,13–20,39 | 18,00–20,30 | 16,25–20,30 | 18,00–19,40 |
| August | 18,23–20,40 | 18,20–20,30 | 18,07–20,35 | 18,14–20,09 |
| Sept. | 17,50–20,46 | 18,45–20,30 | 19,00–19,35 | 17,32–20,10 |
| Oct | 18,50–20,35 | 17,20–20,30 | 16,45–20,30 | 16,35–20,15 |
| Nov | 18,28–20,36 | 18,13–20,35 | 18,15–20,35 | 16,35–20,11 |
| Dec | 18,40–20,31 | 16,22–21,14 | 16,35–19,36 | 16,46–19,42 |
![]()
![]()
![]()