![]()
![]()
![]()
En la América Latina actualmente sólo se encuentran en funcionamiento facilidades de pasaje de peces de dos tipos: escalas en escalones-tanque y esclusa para peces tipo Borland. Del primer tipo se dispone de información sobre la eficiencia de la de Cachoeira de Emas en Brasil (Godoy, 1975) y del segundo tipo se dispone de algunos resultados en Salto Grande (Delfino et al., 1986).
Entre 1942 y 1966 funcionó con un desnivel de casi 5 m. Se la reformó en 1984 y actualmente funciona con un desnivel de 3 m (Godoy, 1985). El río Mogi Guassu a esta altura tiene un caudal medio anual menor a los 100 m3/s.
Hasta el año 1977 el caudal efectivo en el escalón-tanque de entrada de peces a la escala varió entre 0.5 y 2.5 m3/s. Un caudal menor que 0.5 m3/s no resultaba adecuado para la atracción de los peces en el ascenso (Castello, 1982). La velocidad en el primer escalón-tanque aguas abajo variaba entre 1.2 y 3.6 m/s según la altura hidrométrica del río. Entre 2.3 y 3.6 m/s el flujo turbulento de las aguas al pie de la escala y de la presa hacia que los peces vieran dificultada su entrada al sistema.
Las principales especies que utilizaron la escala en sus migraciones reproductivas hacia las cabeceras del Mogi Guassu son también las de mayor importancia comercial (Fig. 10). En observaciones realizadas durante el año 1961, Godoy (1975) menciona a Prochilodus scrofa, Leporinus copelandii, Leporinus octofasciatus, Leporinus elongatus, Salminus maxillosus, Salminus hilarii, Triurobrycon lundii y Pimelodus clarias entre otras. En una ocasión llegaron a pasar de 80 a 100 peces/min de 30 a 70 cm de largo total y con las gónadas en proceso de maduración (Godoy, 1975). En conteos de peces realizados entre 1954 y 1963 se determinó el ascenso de cardúmenes mixtos de 100,000 peces durante 4 o 5 días (Godoy, 1985).

Figura 10. Río Mogi Guassu, Cachoeira de Emas (Pirassununga), Estado de Sao Paulo, Brasil. Escala de peces (desnivel de 5 m en 1959) en la represa de Cachoeira de Emas (Cascada de Emas). Migración reproductiva del “curimbatá” (Prochilodus scrofa Steindacher, 1881) el día 26.10.59. Los dos “curimbatás” saltaban hacia el penúltimo escalóntanque de la escala. Para vencer el último escalón los peces debían saltar una altura de 2.0 m. (Foto: M.P. de Godoy)
La central hidroeléctrica de la represa de Salto Grande entró en funcionamiento en 1979 y el sistema de esclusas para peces lo hizo en 1984. Durante este período fue frecuente la presencia de altas densidades de peces en los cuencos de acumulación aguas abajo (Fig. 6). Su entrada era inducida sólo por el agua auxiliar proveniente de la diferencia de presión entre la zona de descarga de turbinas y el cuenco adyacente a la misma. En el período 1980–85 las mayores acumulaciones aguas abajo de la represa se produjeron hacia fines de marzo comienzos de abril, en coincidencia con el aumento del nivel hidrológico del río, y en los meses de septiembre y octubre en aguas altas. En general se observaron acumulaciones de peces en el período septiembre–abril y nunca en invierno. Durante 2 o 3 días se observaban altas densidades y luego las densidades eran muy bajas. A los tres o cuatro días se repetía el ciclo (Delfino et al., 1986). Los períodos en los cuales se observó mayor densidad aguas abajo coincidieron en general con migraciones ascendentes hacia aguas arriba del embalse. Las especies que mostraron tendencias migratorias ascendentes definidas fueron Salminus maxillosus y Leporinus obtusidens (Delfino y Baigún, 1985). Durante muestreos realizados a lo largo de tres años consecutivos los individuos de estas especies capturados aguas abajo de la represa tenían un factor de condición mayor que los capturados en el embalse. Exactamente lo contrario ocurrió con Prochilodus platensis.
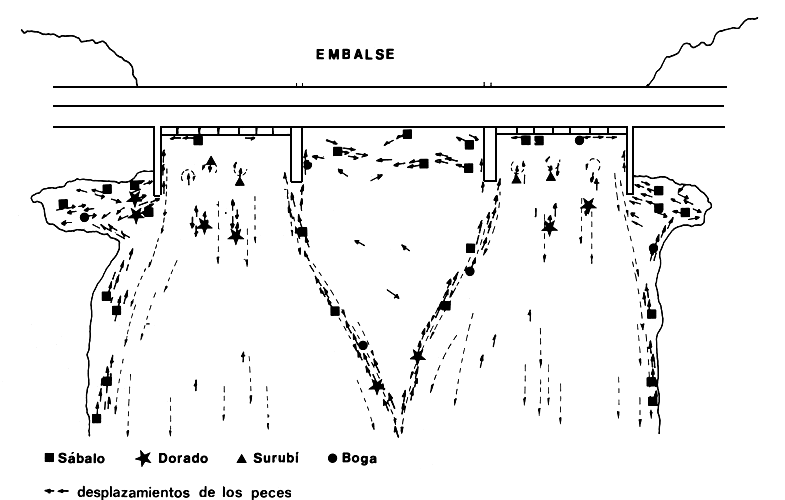
Figura 11. Aproximación de los peces a la represa de Salto Grande (no a escala). Sábalo: Prochilodus platensis; Dorado: Salminus maxillosus; Surubí: Pseudoplatystoma coruscans; Boga: Leporinus obtusidens, (Delfino et al., 1986)
Las acumulaciones de peces aguas abajo se produce en una zona de aproximadamente 200–300 m a todo lo ancho del río sobre el frente de la represa. Las principales especies que se han registrado han sido: Prochilodus platensis, Leporinus obtusidens, Salminus maxillosus, Pseudoplatystoma coruscans, Luciopimelodus pati y Serrasalmus sp. Siempre que está presente una de las especies conocidas como migradoras las otras también lo están. Por otro lado parece que los ejemplares de cada especie se desplazan por la zona con independencia de los de las otras. En los lances pesqueros realizados aguas abajo las capturas frecuentemente fueron uniespecíficas (Delfino et al., 1986).
Se ha observado que los cardúmenes de Prochilodus platensis a unos 1,500 m aguas abajo de la represa, se dirigen hacia esta nadando cerca de la costa. Con el vertedero cerrado, a unos 300–400 m aguas abajo, se aproximan a la represa siguiendo las líneas de velocidad del agua proveniente de las turbinas (Fig. 11). Se observa a Salminus, Leporinus y Prochilodus aproximarse nadando en las zonas de flujo menos turbulento cercanas al agua calma, frente a los vertederos. Esta sería la vía principal de entrada a los cuencos de acumulación. Las otras dos zonas de aproximación corresponden a las cercanas a las respectivas costas, acumulándose en dos zonas de aguas quietas sobre las costas y cercanas a la presa (Fig. 11). Desde esta zona se observa pasar cardúmenes de aproximadamente 200–300 ejemplares de Prochilodus saltando por encima del espigón y del límite entre aguas quietas y turbulentes. Posteriormente se acercan a la represa nadando muy próximos a la pared del espigón (Fig. 12). Otra zona de acumulación de Prochilodus es aquella de aguas quietas frente a los vertederos.
Los individuos de Pseudoplatystoma coruscans, que se desplazan siguiendo los relieves del fondo (Poddubnyi et al., 1981), parecen dirigirse a las salidas de turbinas en el fondo del río. Es común observar ejemplares de esta especie “revolcados” en las corrientes ascendentes de agua de las salidas de turbina. Unos 20 o 30 m aguas abajo de dichas salidas, donde el flujo turbulento ya tiene dirección definida aguas abajo, es común pescar ejemplares de Salminus (Fig. 11).
En general en todas las zonas cercanas a la pared de la represa se encuentran ejemplares de Prochilodus y Leporinus nadando en dirección contraria a las múltiples y débiles corrientes secundarias que allí se producen (Delfino et al., 1986).
Los peces que siguen las líneas de corriente en el límite con las aguas quietas, frente al vertedero, se dirigen hacia los cuencos de acumulación de peces (Fig. 11). Una vez frente a estos se acumulan y luego parte penetra a los cuencos y la mayor parte se dirige hacia la zona de descarga de turbinas. Una vez dentro de los cuencos los peces se acumulan frente a la compuerta de entrada a la esclusa aguas abajo. Algunos permanecen nadando en dirección contraria pegados a las paredes guiados por corrientes secundarias. Estas se producen por interacción entre el agua que se descarga por la esclusa y el agua auxiliar de llamada, proveniente de la zona de turbinas. Las especies que se observaron en los cuencos son: Prochilodus, Leporinus y Salminus (Fig. 13).
La entrada de los peces, y en particular Prochilodus, a la esclusa a través de la compuerta inferior (C2, Fig. 9) parece ser del tipo “todo o nada”: cuando entra al pez que está a la cabeza del cardumen lo sigue el resto. En general se observa que permanecen nadando frente a la compuerta períodos prolongados de tiempo sin decidirse a entrar. Esto podría estar ligado a la falta de iluminación del sistema de esclusas y producirse en C2 una caída brusca en la intensidad de luz (Fig. 13).
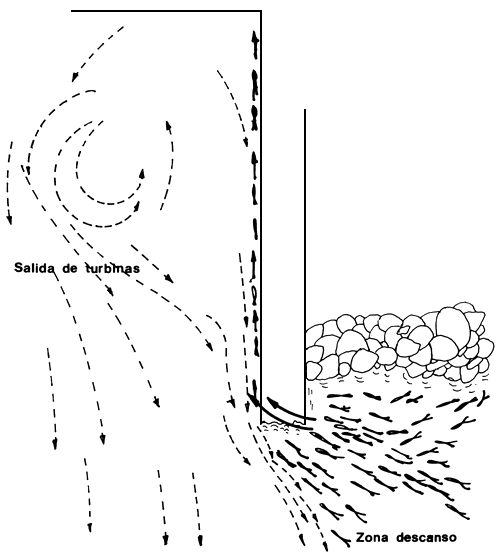
Figura 12. Zona de descanso de los peces sobre costa uruguaya. Saltan sobre el espigón y luego se dirigen, pegados al mismo, hacia los descargadores de fondo y pared de la represa (Delfino et al., 1986)
Durante 1984–85 se detectó que las especies que pasaron por el sistema son: Prochilodus platensis, Leporinus obtusidens, Salminus maxillosus y se observó el pasaje de un ejemplar de Pellona flavipinnis (Delfino et al., 1986). El personal que atiende el sistema menciona el pasaje de Pseudoplatystoma coruscans. A medida que las esclusas se van llenando se ve saltar a los peces fuera de la superficie del agua (Fig. 14) y al completarse el llenado y abrirse completamente la compuerta superior (C1, Fig. 9) la salida hacia el embalse se produce bruscamente con saltos fuera de la superficie del agua y en conjunto. Además de las especies mencionadas, y acompañando a las mismas, se detectan a la salida de las esclusas aguas arriba un número importante de ejemplares de especies de pequeño porte. Su salida de la cámara superior la realizan pegados a las paredes y evitando zonas de turbulencia. Un número importante de estos pequeños peces parece no abandonar la cámara superior hacia el embalse y posiblemente permanezcan dentro de la esclusa durante varios ciclos. No se puede asegurar que no hayan entrado a la esclusa con el agua de llenado proveniente del embalse. Su número en los cuencos de acumulación aguas abajo también es muy importante (Delfino et al., 1986).
En la etapa de llamada de los peces las velocidades en la sección de la compuerta inferior (C2, Fig. 9) se pueden variar entre menos de 0.1 m/s y hasta 1.8 m/s, dependiendo fundamentalmente de la apertura de la compuerta superior y del nivel del río aguas abajo. La velocidad del agua en las entradas a los cuencos sólo se la puede acotar con un límite inferior debido a que la cantidad de agua auxiliar varía en función del número de turbinas en funcionamiento. Para un caudal de 0.9 m3/s en la esclusa y una altura del río aguas abajo de 5.5 m, la velocidad del agua en las entradas a los cuencos es superior a 0.6 m/s (Delfino et al., 1986).
La interacción de los peces que se aproximan a la represa con la instalación para el pasaje, se puede resumir en: (a) una gran proporción de los mismos no penetra al sistema debido al patrón de velocidades aguas abajo, (b) los que penetran a los cuencos encuentran corrientes secundarias dentro de los mismos y una resistencia para entrar a la cámara inferior posiblemente ligada al cambio brusco de iluminación. En todos los ciclos de esclusaje se observan peces pequeños pero sólo en menos del 5 por ciento se produce el pasaje de cardúmenes de 100–200 individuos de las especies de mayor tamaño. Posiblemente se deba a las deficiencias arriba mencionadas y en menor medida a una falta de experiencia en el manejo de la instalación (Delfino et al., 1986).
La puesta en funcionamiento de las modificaciones recomendadas (ver punto 4.4) aumentará la eficiencia de la instalación en lo que respecta a su interacción con los stocks de peces que penetran a los cuencos de acumulación. Sin embargo el diseño original del sistema de esclusas no tomó adecuadamente en cuenta el tipo de río sobre el que se construiría la instalación, el tipo de embalse a formar y el funcionamiento de la central y vertederos. El embalse de Salto Grande es del tipo de canal principal con ramificaciones. El tiempo medio anual de permanencia del agua es de 11 días, y en las épocas en que los peces se acumulan aguas abajo de la represa es menor de 5 días (Quirós y Cuch, 1982). La consiguiente baja capacidad de regulación se ve reflejada en el funcionameinto de la instalación para el pasaje de peces.
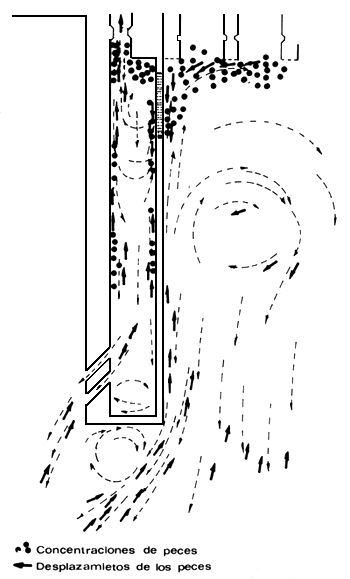
Figura 13. Entrada de los peces a los cuencos de acumulación. La mayoría de ellos se dirige hacia la zona de descarga de turbinas (Delfino et al., 1986)
En los meses de aguas altas (Fig. 15), el caudal de entrada al embalse (Qentr) es prácticamente igual al caudal de salida por vertederos y turbinas (Qsal). La regresión lineal entre el promedio mensual del nivel del río inmediatamente aguas abajo de la represa (L) y el caudal mensual promedio de entrada al embalse (Qentr) para el período 1982–86 es
L = 5.38 + 5.26 × 10-4 × Qentr' n = 72, r = 0.99
con L en m, y Q en m/s. El límite superior de funcionamiento de la esclusa es L = 9.20 m (Comisión Técnica Mixta de Salto Grande, 1983) que corresponde a un caudal de entrada de 7,280 m3/s. Este valor está sólo un 6 por ciento por arriba de la media histórica para el máximo caudal en el ciclo hidrológico anual (Fig. 15). Otro factor que limita el funcionamiento de las esclusas es la apertura de vertederos. Ello está relacionado con el nivel aguas abajo. La relación entre número de días por mes con vertedero cerrado (DC) versus caudal de entrada (Qentr) es
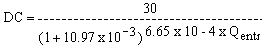
con n = 64 y r = -0.97.
Para el primer período de migraciones ascendentes (marzo-abril) predice que
las esclusas no funcionarían durante el 9 por ciento de tiempo total. Para el
segundo período (septiembre–octubre) no funcionarían un 46 por ciento del
tiempo disponible. Otra restricción al funcionamiento es el nivel del embalse
(Comisión Técnica Mixta de Salto Grande (1983). Para los años 1982–86 no
hubiera funcionado un 44 por ciento del tiempo en el primer período y un 62
por ciento en el segundo. Para el período 1984–86 efectivamente no funcionó
un 72 y 54 por ciento del tiempo durante el primer y segundo períodos de migraciones
respectivamente (Delfino et al., 1986).
Un cálculo aproximado nos dará una idea de las limitaciones de las esclusas Borland de Salto Grande para operar con vertedero abierto. Cada una de las 14 turbinas Kaplan de la represa requiere 600 m3/s a carga completa, y el ancho del vertedero es de 357 m, suponiendo que el caudal del río esté en la media histórica para el mes de octubre, de 6,900 m3/s, y las centrales operen al 50 por ciento de su capacidad máxima. El nivel aguas abajo que se predice con la regresión anterior es de 9 m. La profundidad neta media aguas abajo en la zona de vertedero sería de unos 6 m y el caudal vertido de 2,700 m3/s. Ello nos da una velocidad media mínima del agua por debajo de los disipadores del vertedero de 1.3 m/s. Esta es la velocidad máxima que podría estimarse para las aberturas de entrada de los peces a los cuencos de acumulación. El ancho de cada una de las cuatro entradas es de 1 m y el del vertedero de 357 m. En todo caso si algunos peces lograran encontrar las entradas y pasar al embalse, posiblemente a la salida de estas serían atrapados en el vertedero y volverían aguas abajo.
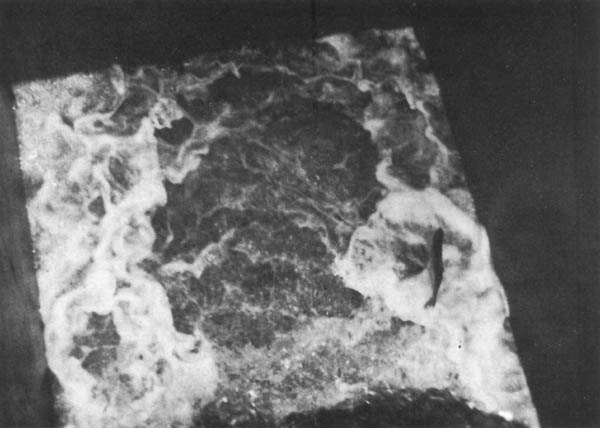
Figura 14. Etapa de llenado de la esclusa. Prochilodus saltando fuera de la superficie del agua. (Foto: R. Delfino)
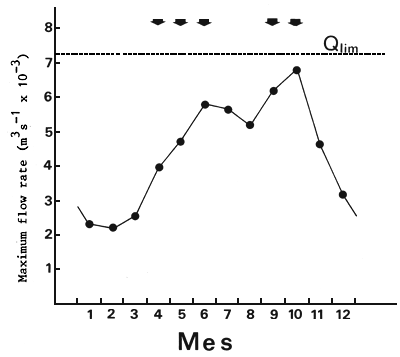
Figura 15. Ciclo hidrológico del río Uruguay. Caudales mensuales promedio. Qlim: caudal de entrada al embalse por encima del cual el sistema de esclusas no funciona. Las flechas indican los meses en los cuales se verificaron migraciones ascendentes en el río Uruguay (ver explicación en el texto)
Un manejo de los caudales de la turbina y del vertedero que tomara en cuenta estas limitaciones podría llegar a mejorar un poco el panorama anterior. Las mayores demandas de energía se producen durante el día, período durante el cual Prochilodus, Salminus y posiblemente Leporinus desarrollan sus actividades migratorias. Dentro de lo posible sería conveniente disminuir al mínimo el caudal por vertedero durante el día y descargar durante la noche cuando Pseudoplatystoma y Luciopimelodus se desplazan. Para estos la eficiencia del sistema de transferencia como un todo parece ser muchisimo menor (Delfino et al., 1986).
Resumiendo, el sistema presenta deficiencias de funcionamiento, con respecto a los peces, a dos niveles: en la aproximación de los peces a la represa para encontrar las entradas a los cuencos de acumulación, y en la interacción de los peces con el sistema de esclusas. Por encima de estos problemas, se encuentran los del diseño con respecto al caudal y al ciclo hidrológico del río.
![]()
![]()
![]()