![]()
![]()
![]()
Qualitative phenotypes are usually less important than quantitative phenotypes for food fish farmers, but an understanding of how they can be controlled and exploited by selective breeding programmes is important for two reasons: First, and most important, is the fact that qualitative phenotypes can affect the value of the crop or the cost of production. If a farmer can produce a more attractive product, consumers are often willing to pay more for it, which increases his profits. On the other hand, some qualitative phenotypes, such as deformities, can decrease the value of the population, either by increasing the cost of production or by producing fish so unattractive that consumers will not buy them. In both cases, farmers can use selection to improve their populations. Secondly, an understanding of how selective breeding programmes can be used to fix (frequency = 100%) the desired qualitative phenotypes and eliminate (frequency = 0%) the undesired ones and thus produce true-breeding populations may make it easier to understand how selection can be used to improve quantitative phenotypes.
Those who already have a good grasp of Mendelian genetics and understand how breeding programmes can be used to fix or to eliminate qualitative phenotypes, or those who only wish to work with quantitative phenotypes, may skip this chapter and go directly to Chapter 4.
Before conducting a selective breeding programme, a farmer must realize that he can use selection to accomplish only those goals which are achievable biologically. Selection is a breeding programme that exploits heritable phenotypic variance; consequently, the two prerequisites for a successful selective breeding programme are: one, the phenotype that the farmer wants to change must exhibit variance; two, the variance must be heritable. Breeders are able to accomplish only what the fish and the fish's genes allow them to accomplish. Breeders cannot custom-design fish by removing undesirable phenotypes, such as sharp pectoral spines, unless individuals without these spines are discovered, and even if such individuals are discovered, the phenotype must be heritable or selection will be an expensive, futile, and frustrating waste of effort. If the phenotypes in question are heritable, the genetics should be known before a breeding programme is conducted. This information will enable the breeder to use the most appropriate breeding programme. Finally, the farmer must know the relative costs of the traits (production costs) and what they are worth (market value) before he initiates a selective breeding programme.
Qualitative phenotypes can be described as “either/or” phenotypes. A fish either has one phenotype or it has another; in other words, the fish in a population fall into discrete, non-overlapping phenotypic categories. Because selection exploits phenotypic variance, at least two phenotypes must exist in the population, and one must be the phenotype that the farmer wants. If there is only one phenotype, there is no phenotypic variance, which means that selection cannot be used to alter the phenotype. For example, if all fish in a population are black, there is no phenotypic variance for body colour; consequently, selection cannot be used to change body colour.
Not only must variance exist, but the variance that is observed must be heritable, or selection will not be able to alter phenotypic frequencies. The distinction between heritable and non-heritable variance is important, because many qualitative phenotypes that are observed at fish farms and at fish hatcheries are non-heritable deformities. Some deformities have a genetic basis, but most are caused by nutritional deficiencies, environmental disturbances, toxins, disease, or injury or they are developmental mistakes. The deformities that are produced by non-genetic factors cannot be eliminated by selection. The only way to eliminate a non-heritable deformity is to discover the environmental factor that causes it and to eliminate the factor or modify the culture environment. It is often difficult, if not impossible, to identify the cause of a non-heritable deformity, so unless the frequency approaches 0.5% it probably should be ignored.
Before selection is used to fix or to eliminate qualitative phenotypes and produce true-breeding populations, the way the phenotypes are inherited should be determined. If a previous experiment has already determined the mode of inheritance, that information can be used to design the breeding programme. If the mode of inheritance is known, the correct breeding programme can be chosen, and this will enable the farmer to achieve his goal quickly and efficiently. If the mode of inheritance is not known prior to the breeding programme, the farmer may choose the wrong programme, which means he will not accomplish his goal and will waste effort, facilities, and money.
The research programmes that are needed to determine the modes of inheritance for qualitative phenotypes are not that complicated. In general, the inheritance is deciphered by conducting paired matings across two generations and by determining the phenotypic ratios produced by these matings (some of these matings and the ratios that are used are illustrated in Figures 3 and 5). Even though this type of research is not that complicated, it requires effort, facilities, and the ability to analyze the results statistically. Such research should be left to scientists at universities or governmental research stations.
Before conducting a selective breeding programme, a farmer should conduct an assessment to determine if a selective breeding programme is needed. The assessment can be conducted by a farmer (such studies are not cheap, and they require a high level of sophistication), or they can be conducted by fisheries cooperatives or a local fish farming research station. The farmer needs to determine if his customers want fish with a different body colour, or if they will pay extra for a prettier fish. His assessment will also tell him if the proposed breeding programme could open new markets. A farmer should only conduct a selective breeding programme when it is necessary or when it will increase his profits. Changing body colour or other phenotypes might produce a prettier fish, but if the assessment reveals that consumers do not want a different body colour, the breeding programme should not be conducted.
Finally, the farmer must know the costs of producing the alternate phenotypes before conducting a selective breeding programme. The values of some phenotypes are so obvious that no formal scientific study is needed. If a farmer wants to use selection to eliminate a heritable deformity which lowers growth rate or viability, formal studies to determine the relative values of the phenotypes are unnecessary, because he already knows that the deformity costs him money.
On the other hand, the relative production costs of some phenotypes are more difficult to determine and require evaluation in a scientific experiment. The relative production costs are determined by assessing the effects that the phenotypes have on growth rate, survival, egg production, etc. These secondary effects are called “pleiotropic effects,” and in agriculture and aquaculture they can be more important than the phenotype itself--especially if growth, survival, or fecundity are affected. Almost all mutant body colours that have been investigated in fish have exhibited negative pleiotropic effects, in that they adversely affected growth rate or survival. Figure 8 illustrates the negative pleiotropic effect that red body colour has on viability in Nile tilapia.
If there are no negative pleiotropic effects or if the extra costs of production (slower growth rate and/or lower viability) are offset by the increased market value of the crop (extra money consumers are willing to pay), selection is worthwhile. On the other hand, if the increased costs of production exceed the extra market value of the crop, it would be foolish to conduct a selective breeding programme because the farmer will earn less money.
When working with qualitative phenotypes, the goal of all selective breeding programmes is to produce a true-breeding population. A true-breeding population is one that contains only one allele for the locus in question because it was fixed; selection eliminated the other allele. This means that every gamete produced by the select brood fish will contain only the desired allele, which means that every offspring produced by the select brood fish will have the desired phenotype. The only way the undesired phenotype can be produced after selection has been used to create a true-breeding population is by the accidental (or intentional) stocking of fish from another population or by mutation.
The undesired phenotype and the allele that produces it are eliminated by a process called “culling.” Culled fish are not allowed to reproduce. They do not have to be killed; they can be grown and then sold as food, but they must not be allowed to spawn.
There are three basic rules that govern the selection of qualitative phenotypes:
If the desired phenotype is controlled only by a homozygous genotype, a single act of selection will create a true-breeding population.
If the desired phenotype is controlled by two or more genotypes, selection cannot create a true-breeding population. Progeny testing must be used to fix the desired allele and produce a true-breeding population.
If the desired phenotype is controlled by the heterozygous genotype, no breeding programme can create a true-breeding population. A population in which every fish has the desired phenotype can be produced by crossing the two homozygous phenotypes, but this procedure must be repeated every breeding season.
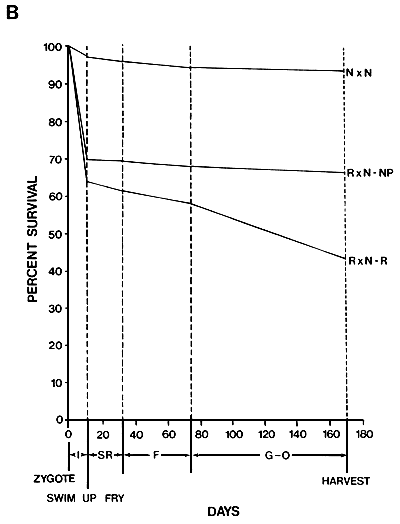
Figure 8. Effect of red body colour on viability in Nile tilapia. This is an example of a negative pleiotropic effect. Viability or red fish (R × N-R) is compared to that of their normally pigmented sibs (R × N-NP) and a control population which had no red fish (N × N). Viabilities during artificial incubation of eggs (I), sex reversal (SR), fingerling production (F), and grow-out (G-O) are shown. At harvest, fish averaged 250 g.
Source: After El Gamal, A.R.A.L. 1987. Reproductive performance, sex ratios, gonadal development, cold tolerance, viability and growth of red and normally pigmented hybrids of Tilapia aurea and T. nilotica. Doctoral dissertation, Auburn University, Alabama, USA.
When this type of selection is used, it is usually done because a farmer wants to fix the recessive phenotype and eliminate the dominant phenotype, and the phenotypes are produced by an autosomal gene that exhibits complete dominance. The term “recessive” does not imply inferiority; similarly, the term “dominant”does not imply superiority. The terms simply refer to the way the alleles express themselves and to the phenotypes that are produced by those alleles. Many recessive phenotypes have been shown to be quite valuable in various farmed plants and animals, and breeding programmes have been conducted to fix them. The terms “inferior” and “superior” should be applied to phenotypes only after biological and economic evaluations have been conducted.
If the desired trait is the recessive phenotype that is produced by a gene that exhibits complete dominance, the selective breeding programme that is needed to fix the trait and to produce a true-breeding population is simple, and this goal can be achieved by a single act of selection. To accomplish this, fish with the recessive phenotype will be saved, and fish with the dominant phenotype will be culled.
Genetically, fish with the undesired dominant phenotype are either homozygous dominant or heterozygous. Because every fish that has at least one dominant allele expresses the undesired dominant phenotype, a simple one-step culling of all fish with the dominant phenotype will eliminate all copies of the dominant allele. Conversely, every fish that expresses the desired recessive phenotype is homozygous recessive, which means that when these fish are selected, none will possess a copy of the undesired dominant allele. Thus, the recessive allele will be the only allele that exists in the select population, which means that it will breed true.
For example, say a tilapia farmer has both normally pigmented and pink Nile tilapia, and decides to grow only pink tilapia. He can create a true-breeding pink population by a simple one-step selective breeding programme during which he will cull all normally pigmented fish. Pink body colour is the recessive phenotype, and it is produced by the recessive b allele (bb genotype); normal body colour is the dominant phenotype, and it is produced by the dominant B allele (BB and Bb genotypes).
The culling of all normally pigmented tilapia will remove all B alleles from the population. The only fish that remain will be pink and, since they are homozygous recessive bb, the b allele will be the only allele that will remain in the population of select brood fish. Consequently, a single act of selection will create a true-breeding pink population (Figure 9).
This type of selective breeding programme can also be used to fix either the recessive or the dominant phenotypes when the mode of gene action is incomplete dominance, and it can also be used to fix either homozygous phenotype when the mode of gene action is additive. In both cases, there are two homozygous genotypes that can be identified and isolated. By culling the undesired phenotypes, true-breeding populations can be created by a single act of selection.
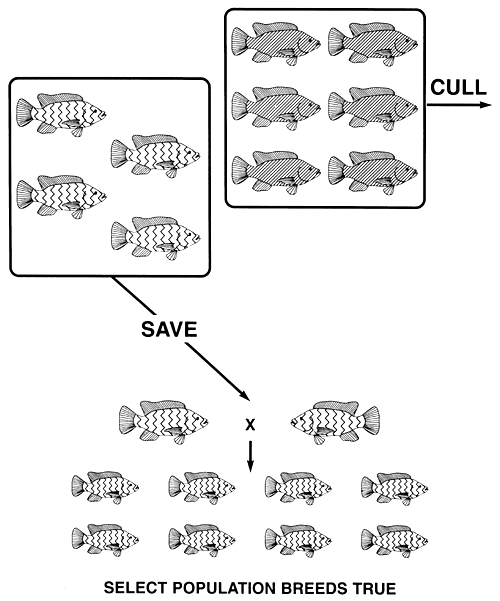
Figure 9. Selective breeding programme needed to produce a true-breeding population of pink Nile tilapia. If all normally pigmented (the dominant phenotype) fish are culled, all copies of the dominant B allele will be eliminated. The only fish that remain are pink (the recessive phenotype), and since they are homozygous recessive bb, the select population of pink brood fish will breed true and will produce only pink offspring. Pictorial representations of the phenotypes are the same as those used in Figure 2. Any recessive phenotype can be fixed this easily.
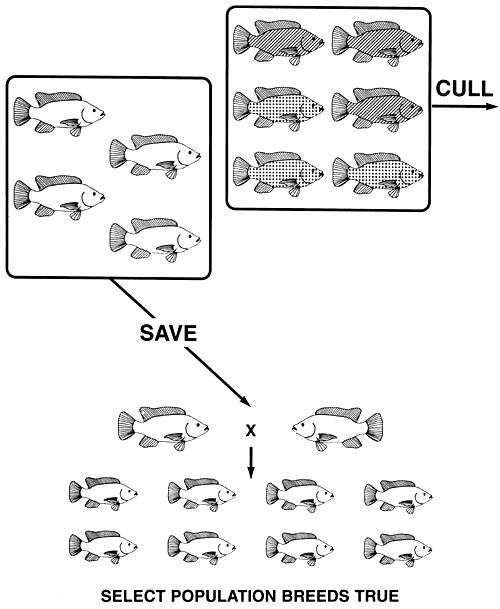
Figure 10. Selective breeding programme needed to produce a true-breeding population of gold Mozambique tilapia. If all melanistic (black [GG] and bronze [Ggl]) fish are culled, all copies of the dominant G. allele will be eliminated. The only fish that remain are gold (the recessive phenotype), and since they are homozygous recessive gg, the select population of gold brood fish will breed true and will produce only gold offspring. Pictorial representations of the phenotypes are the same as those used in Figure 4.
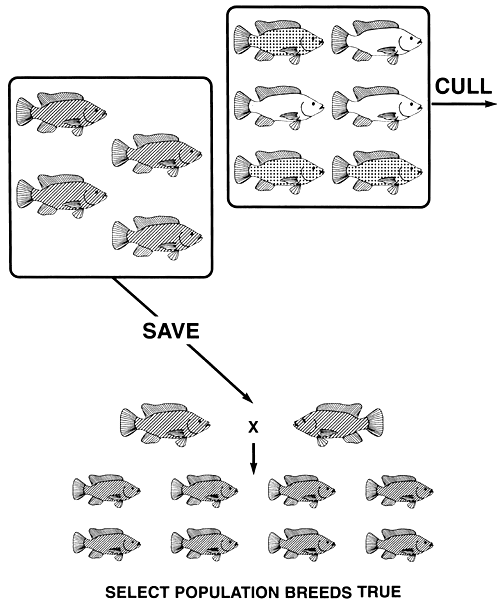
Figure 11. Selective breeding programme needed to produce a true-breeding population of black Mozambique tilapia. If all bronze (the heterozygous [Gg] phenotype) and all gold (the recessive [gg] phenotype) fish are culled, all copies of the recessive g. allele will be eliminated. The only fish that remain are black (the dominant phenotype), and since they are homozygous dominant GG, the select population of black brood fish will breed true and will produce only black offspring. Pictorial representations of the phenotypes are the same as those used in Figure 4.
For example, say a tilapia farmer has black, bronze, and gold body colours in his population of Mozambique tilapia. If the farmer wants to produce either a true-breeding population of gold or a true-breeding population of black fish, he can accomplish either goal by a single act of selection. Gold is the recessive phenotype, and it is produced by the homozygous recessive genotype (gg); black is the dominant phenotype, and it is produced by the homozygous dominant genotype (GG); bronze is the heterozygous (Gg) phenotype.
A true-breeding population of gold tilapia, can be produced by culling all black and all bronze fish (Figure 10). Conversely, a true-breeding population of black tilapia can be produced by culling all gold and all bronze fish (Figure 11).
This same concept can be extended to phenotypes that are controlled by two or more genes. If the desired phenotype is controlled only by a homozygous genotype, a true-breeding population can be created by a single act of selection.
For example, if a carp farmer has a population of common carp with all four scale phenotypes and wants to produce a true-breeding population that has a reduced scale pattern, all he has to do is save the fish with the mirror phenotype and cull all other fish. Three of the scale phenotypes have a reduced number of scales, but only mirror is produced by a homozygous genotype (ss, nn), so it is the only one that is capable of breeding true. Both the leather (ss, Nn) and line (SS, Nn and Ss, Nn) phenotypes are undesired, because they are heterozygous at the N locus, which means that when two leather and/or line fish mate, 25% of their offspring will die (those that are homozygous NN. regardless of the genotype at the S locus-see Figure 6). Leather and line phenotypes are also undesired, because fish with these phenotypes have several negative pleiotropic effects, among them lowered growth rates and viabilities.
If the farmer culls all leather, line, and scaled individuals, all S and N alleles will be eliminated. The only fish that will remain in the select population will be those with the mirror phenotype, and since they are homozygous recessive (ss, nn) the s and n alleles will be the only ones that remain in the population of select brood fish. Consequently, a single act of selection will create a true-breeding population of mirror common carp (Figure 12).
Selection cannot fix a phenotype that is controlled by more than one genotype. This means selection cannot fix a dominant phenotype and create a true-breeding population if it is produced by a gene with complete dominance. Conversely, this also means that selection cannot eliminate a recessive phenotype (and the recessive allele) if it is produced by a gene with complete dominance. Despite the fact that this type of selective breeding programme will not work, it is routinely used in a misguided effort to eliminate undesired phenotypes that are produced or that are assumed to be produced by recessive alleles.
This type of breeding programme will not achieve its goal for either of two reasons. The first is the fact that most undesired phenotypes are abnormalities that are created by non-genetic factors (environmental disturbances or developmental mistakes). Many assume that all abnormal phenotypes are mutant phenotypes which are produced by recessive alleles. Some are, but most are not. If the undesired phenotype is produced by a non-genetic factor, selection will not work, because no selective breeding programme can fix or eliminate non-heritable phenotypes.
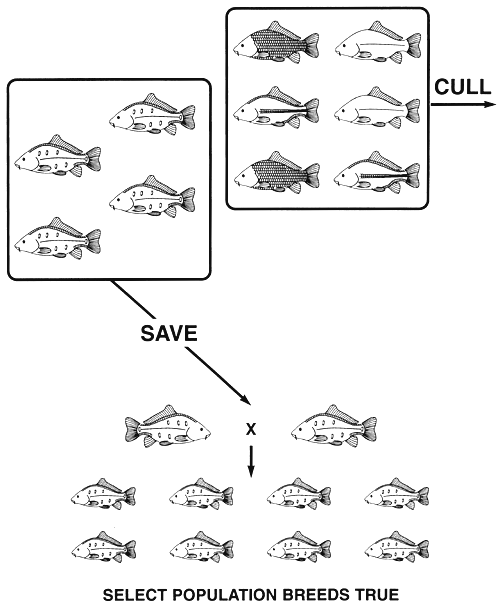
Figure 12. Selective breeding programme needed to produce a true-breeding population of mirror common carp. If all scaled, line, and leather fish are culled (phenotypes that are produced by genotypes with at least one dominant allele-see Figure 6), all copies of the S and N alleles will be eliminated. The only fish that remain are those with the mirror (the recessive) phenotype, and since they are homozygous recessive ss, nn the select population of mirror brood fish will breed true and will produce offspring with only the mirror phenotype. Even if a set of phenotypes are produced by two or more genes, if the desired phenotype is the recessive phenotype (all loci responsible for the production of the phenotype are homozygous recessive), selection can produce a true-breeding population this easily.
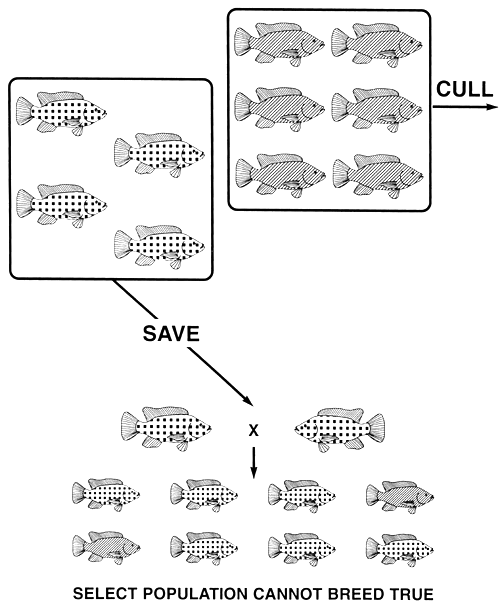
Figure 13. Selective breeding programme used in an unsuccessful attempt to produce a true-breeding population of red Nile tilapia. Red fish are represented by fish covered with squares, while normally pigmented fish are those covered with stripes (the same pictorial representations for body colour will be used in Figure 15). If all normally pigmented fish (the recessive phenotype) are culled, the select population will be composed of only red fish (the dominant phenotype). Since red body colour is produced by a dominant allele that exhibits complete dominance, red is produced by both the homozygous dominant (RR) and by the heterozygous (Rr) genotypes. Every heterozygous red brood fish (Rr) that is saved will carry a copy of the r allele to the select population; consequently, selection cannot eliminate it. When select red brood fish are spawned, the mating of two heterozygotes (Rr) will produce some normally pigmented offspring (the mating that is depicted). Recessive phenotypes cannot be eliminated by culling them; they will reappear when the select brood fish are spawned. Consequently, if the desired phenotype is the dominant phenotype and it can be produced by both the homozygous dominant and by the heterozygous genotype, selection cannot create a true-breeding population.
Even when a deformity is produced by a recessive allele, if it is an autosomal gene that exhibits complete dominance, selection will not create a deformity-free population. The reason why it will fail is quite simple: The dominant (normal) phenotype is produced by either of two genotypes, and it is impossible to differentiate normal fish that are homozygous dominant from those that are heterozygous.
Because of that, when a farmer culls the recessive (abnormal) fish, the select population will be composed of fish with one phenotype (normal) but two genotypes, and every heterozygous select normal brood fish will carry a copy of the undesired recessive allele. Because the select population contains both alleles, it cannot breed true. When two heterozygous select normal brood fish mate, they will produce offspring with the undesired abnormal (recessive) phenotype. This type of selection will reduce the frequency of an undesired recessive phenotype, but it cannot eliminate it and produce a true-breeding deformity-free population.
For example, say a tilapia farmer has a population of Nile tilapia that contains both normally pigmented and red individuals, and decides to produce only red tilapia. If he tries to produce a true-breeding population of red Nile tilapia by culling all normally pigmented fish, he will discover that he will be unable to produce a red population that breeds true. The select red brood fish will produce both red and normally pigmented offspring. This is because red is the dominant phenotype, and it is produced by the dominant R allele (RR and fir genotypes), while normal pigmentation is the recessive phenotype, and it is produced by the recessive r allele (rr genotype). Because the homozygous dominant and the heterozygous genotypes produce identical red phenotypes, some of the select red brood fish (the heterozygotes) will have a copy of the undesired r allele. When two heterozygous red brood fish mate, they will produce some normally pigmented offspring (Figure 13).
The same concept applies for phenotypes that are controlled by two or more genes. If the desired phenotype is produced by more than one genotype, and if at least one of the genes can be in either the homozygous or heterozygous state, selection cannot create a true-breeding population. As was the case for phenotypes produced by a single gene, some of the select brood fish will carry a copy of the undesired recessive allele(s) and, when two heterozygotes mate, they will produce offspring with the undesired phenotype(s).
For example, if a carp farmer has a population of common carp with all four scale phenotypes and decides to produce a true-breeding population of scaled common carp by selection, he will discover that he cannot create a true-breeding population. This is because the select scaled brood fish will have two genotypes--SS, nn or Ss, nn-and it is impossible to tell heterozygous scaled fish from homozygous scaled fish. The farmer can eliminate the dominant N allele by culling all fish with the line and leather phenotypes (a dominant allele can be culled by a single act of selection); those individuals with two copies of the N, allele were culled by nature and died. However, the farmer cannot eliminate the undesired s allele by culling the remaining phenotype-mirror. The select population of scaled brood fish will produce both mirror and scaled offspring when two select heterozygous (Ss, nn) scaled brood fish mate (Figure 14).
Progeny testing: Since selection against (culling) a recessive phenotype cannot fix a dominant allele and produce a true-breeding population (when the mode of gene action is complete dominance), another type of breeding programme must be used to achieve this goal. The only way an undesired recessive allele can be eliminated is by progeny testing. Progeny testing is a breeding programme that deciphers a parent's genotype by determining its offspring's phenotypes. Once the genotypes of the dominant parents are deciphered, selection is used to save the homozygotes and cull the heterozygotes, which will eliminate all copies of the recessive allele, fix the dominant allele, and produce a true-breeding population.
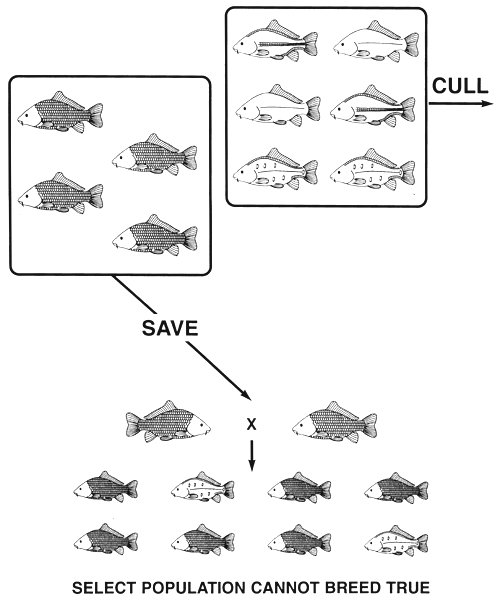
Figure 14. Selective breeding programme that is used in an unsuccessful attempt to produce a true-breeding population of scaled common carp. If all fish with the line, mirror, and leather phenotypes are culled, the select population will be composed of scaled fish. This act of selection will eliminate all copies of the undesired dominant N allele, because every heterozygote (line and leather phenotypes) will be culled (the homozygotes were culled by nature, since the NN genotype is lethal). Unfortunately, this selective breeding programme will not produce a true-breeding population because you cannot eliminate the undesired recessive s allele by culling the recessive phenotype (mirror). Because the scaled phenotype is produced by a dominant allele that exhibits complete dominance, both homozygous (SS, nn) and heterozygous (Ss, nn) scaled fish will be saved. Every heterozygous select scaled brood fish will carry a copy of the recessive s allele to the select population; consequently, selection cannot eliminate it. When select scaled brood fish are spawned, the mating of two heterozygotes will produce some offspring with the mirror phenotype (the mating that is depicted). As was the case with a phenotype produced by a single gene, if the desired phenotype is produced by more than one genotype and if one of the genotypes is a heterozygous genotype, selection cannot create a true-breeding population.
This is accomplished by pairing and mating fish with the dominant phenotype to test fish (a fish whose genotype is known). Test fish are usually fish with the recessive phenotype, because such fish are homozygous recessive (experiments are not needed to determine a recessive phenotype's genotype). Because a test fish can produce gametes with only a recessive allele, the dominant fish is the parent that determines its offspring's phenotypes (see Figure 3c and 3e). If the dominant parent is homozygous, all offspring will have the dominant phenotype (Figure 3c); if the dominant parent is a heterozygote, half the offspring will have the dominant phenotype and half will have the recessive phenotype (Figure 3e).
The production of a single offspring with the recessive phenotype is enough evidence to state with certainty that the dominant parent is a heterozygote and must be culled. If no offspring with the recessive phenotype is detected in a random sample of at least 20 offspring, the dominant parent can be declared to be homozygous and kept for breeding purposes. Once a sufficient number of males and females are progeny tested and declared to be homozygous dominant, all other fish (including all offspring produced in the progeny tests) are culled. The population will now breed true and produce only offspring with the dominant phenotype.
For example, a tilapia farmer that has a population of red and normally pigmented Nile tilapia can create a true-breeding red population by using progeny testing to identify and keep red fish that are homozygous (RR) and to identify and cull red fish that are heterozygous (Rr). Before initiating the progeny testing programme, the farmer will cull all normally pigmented fish except for the ones that he will use as test fish.
The farmer progeny tests the red fish by pairing a red fish with a test fish (in this case, normally pigmented tilapia are used as test fish since they are homozygous recessive rr). Red parents that are homozygotes (RR) will produce only red offspring and will be saved; red parents that are heterozygotes (Rr) will produce equal numbers of red and normally pigmented offspring and will be culled. After one generation of progeny testing, the farmer will have a select population of red brood fish that will breed true and produce only red offspring, because every select red brood fish is homozygous RR (Figure 15).
The same principle applies if the desired phenotype is controlled by two or more genotypes and the phenotype is controlled by more than one gene. For example, if a carp farmer has a population of common carp with all four scale patterns and decides to produce a true-breeding population of scaled common carp, he will have to use progeny testing to accomplish his goal. Scaled fish are produced by both the SS, nn and Ss, nn genotypes. In this case, fish with the mirror scale pattern are used as the test fish because they are homozygous recessive (ss, nn). Before initiating the progeny testing programme, the farmer will cull all fish with the line and leather phenotypes and most fish with the mirror phenotypes; he will keep a few fish with the mirror phenotype and use them as test fish.
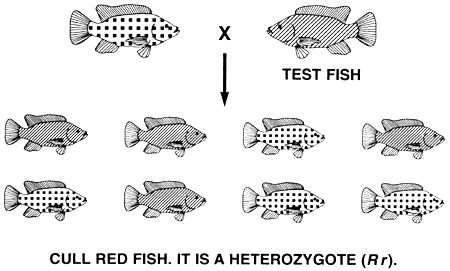
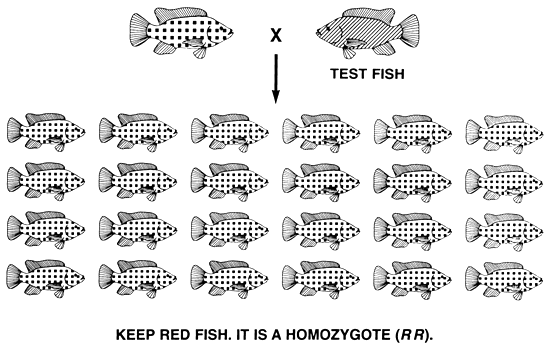
Figure 15. Schematic diagram of the progeny testing programme that must be used to create a true-breeding population of red Nile tilapia. Progeny testing is used to identify and cull the heterozygous (Rr) red fish and to identify and save the homozygous (RR) red fish. A red fish's genotype is deciphered by mating it to a test fish (a normally pigmented fish [rr]), and by then determining the phenotypes of its offspring. In the top mating, the red fish is identified as a heterozygote (Rr) and is culled, because half of its offspring are normally pigmented. In the bottom mating, the red fish is considered to be a homozygote (RR) and is saved, because no normally pigmented fish was found in a random sample of a least 20 of its offspring. Progeny testing will create a population of select red brood fish that are all certified to be homozygous BE, and such a population will breed true and produce only red offspring. Pictorial representations of the phenotypes are the same as those used in Figure 13.
Progeny testing is done to identify and save scaled fish that are homozygous at the S locus (SS,nn) and to identify and cull scaled fish that are heterozygous at the S locus (Ss.nn). If the scaled parent is homozygous at the S locus (SS, nn), all offspring will have the scaled phenotype; if the scaled parent is a heterozygote (Ss, nn), half the offspring will have the scaled phenotype and half will have the mirror phenotype. After one generation of progeny testing, the farmer will have a select population of scaled brood fish that are all SS, nn, and it will breed true and produce only scaled progeny (Figure 16).
When progeny testing, if the fish can be stripped, a single test fish can be used to progeny test a number of fish with the dominant phenotype. Additionally, it is not necessary to raise thousands of individuals from each family. A random sample of 100–200 fertilized eggs should be sufficient, if survival to the early fingerling stage is good. Consequently, progeny testing does not have to be expensive; it can be done in a small hatchery in aquaria, in small plastic swimming pools, or in hapas suspended in a single pond no larger than 0.04 ha. While the families are being raised, the dominant brood fish must be kept in isolated tanks or hapas so they can be saved or culled once the progeny are examined. Several fish can be kept in a tank or hapa if they are marked by fin clips. A single generation of progeny testing will create a true-breeding population, and it will not be needed again, unless fish carrying the undesired recessive allele are acquired from another farm or the allele arises by mutation.
If the heterozygous phenotype is desired, no breeding programme can be used produce a true-breeding population. This is because fish with the desired phenotype are unable to breed true. Heterozygotes produce two types of gametes and produce them in equal numbers. Consequently, when they mate, they produce offspring with three phenotypes (selection for the heterozygous phenotype can only be done for phenotypes controlled by autosomal genes with incomplete dominance or with additive gene action). A farmer who culls the two homozygous phenotypes and creates a select population containing only the heterozygous phenotype, will quickly discover that the selective breeding programme is a failure, because only half the offspring produced by the select brood fish will have the desired phenotype.
For example, if a tilapia farmer has black, bronze, and gold phenotypes in his population of Mozambique tilapia and decides to produce a true-breeding population of bronze fish, he will find that it is impossible. If he conducts a selective breeding programme, he will cull the gold (the recessive phenotype) and the black (the dominant phenotype) fish and will save the bronze ones (the heterozygotes). When he breeds the select population of bronze brood fish, they will produce offspring with all three phenotypes (Figure 17).
If the farmer wants to produce nothing but bronze fish, he must mate gold fish with black fish. This type of mating (the mating of the two homozygotes; it is illustrated in Figure 5c) will produce 100% heterozygous offspring (Figure 18).
The only requirement for this breeding programme is that the farmer must be able to accurately sex his fish. If the sex of a single fish is mis-identified, gold or black offspring will be produced. This type of mating scheme requires the use of at least two ponds (or tanks) for offspring production: one will be stocked with gold females and black males; the other will be stocked with black females and gold males.
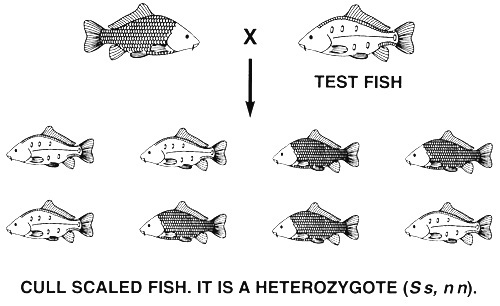
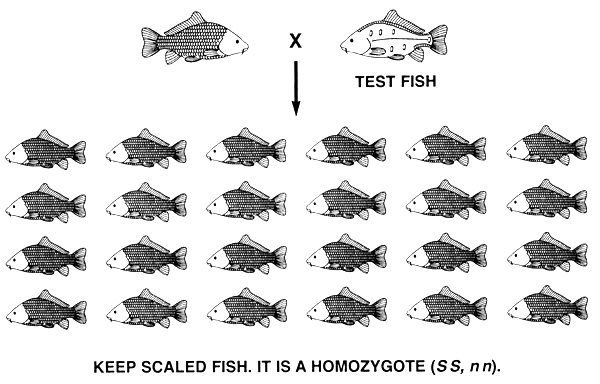
Figure 16. Schematic diagram of the progeny testing programme that must be used to create a true-breeding population of scaled common carp. Progeny testing is used to identify and cull the heterozygous (Ss, nn) scaled fish and to identify and save the homozygous (SS, nn) scaled fish. A scaled fish's genotype is deciphered by mating it to a test fish (a fish with the mirror phenotype [ss, nn]), and by then determining the phenotypes of its offspring. In the top mating, the scaled fish is identified as a heterozygote (Ss, nn) and is culled, because half of its progeny have the mirror phenotype. In the bottom mating, the scaled fish is considered to be a homozygote (SS, nn) and is saved, because no fish with the mirror phenotype was found in a random sample of at least 20 of its offspring. Progeny testing will create a select population of scaled brood fish that are all certified to be homozygous SS, nn, and such a population will breed true and produce only scaled offspring.
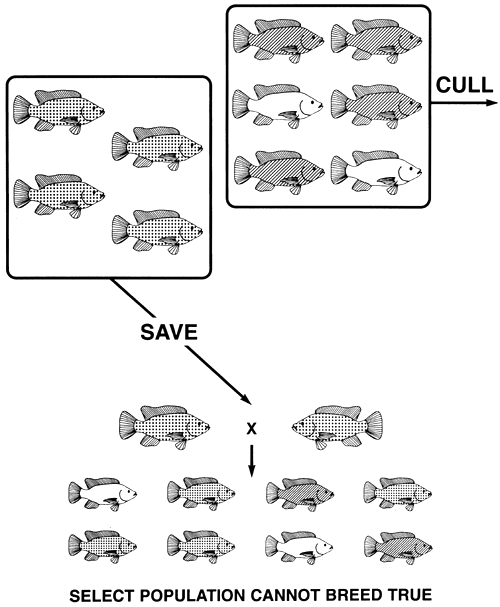
Figure 17. Selective breeding programme used in an unsuccessful attempt to produce a true-breeding population of bronze Mozambique tilapia. In this case, both homozygotes-black (GG) and gold (ggl)-are culled, and the bronze (Gg) fish are saved. Because bronze fish are heterozygotes, the select population cannot breed true. Only half the progeny produced by the select brood fish will be bronze. Pictorial representations of body colour are the same as those used in Figure 4.
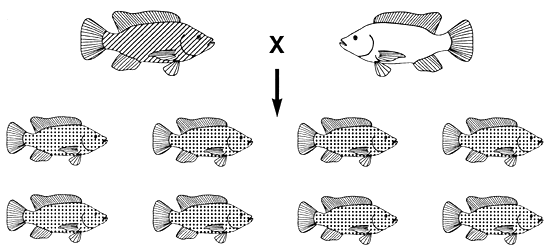
Figure 18. Breeding programme needed to produce 100% bronze Mozambique tilapia. A population of bronze (Gg) fish can be produced only by mating black (GG) fish with gold (gg) fish. If a farmer wants to grow only bronze fish, he must use this mating programme every breeding season. Pictorial representations of body colour are the same as those used in Figure 4.
When compared with the volume of records that must be kept when working with quantitative phenotypes, the types and amount of records that must be taken and maintained when working with qualitative phenotypes are minimal. One type of information that should be recorded is a description of abnormal or “mutant”phenotypes that are observed every year. Farmers' memories are not perfect, and they often feel that they saw more abnormal fish than really existed, simply because these fish are so unusual. A census of these phenotypes will tell a farmer what types of abnormalities have been produced and will also let him know if an abnormality is becoming a problem. These data will also provide a farmer with information about other qualitative phenotypes that exist in his population.
A farmer should have a data sheet on which he records the date, the name of the phenotype (a name can be invented if none exists), and a brief description of the phenotype. In addition, the number of fish that had this phenotype and the total number of fish that were produced should be recorded. This will let the farmer know the frequencies for each abnormality. If the frequencies begin to increase over time, the farmer will know that there is a potential problem. An example of this type of data sheet is presented in Table 7.
Table 7. Example of a data sheet that can be used to record abnormal and/or other qualitative phenotypes that are observed.
| Date: March 23, 1995 Species: Nile tilapia Number produced: 25,500 | |||
| Phenotype | Description | Number | Frequency |
| Tail less | No tail; no caudal peduncle | 3 | 0.000117 |
| Semi-operculum | Right operculum short; gills partially exposed | 5 | 0.000196 |
| Stumpbody | Dwarf; trunk abnormally short | 2 | 0.000078 |
If the farmer is going to conduct a selective breeding programme to fix a qualitative phenotype, he should record the date(s) on which selection occurred, the number of fish that were saved, the number of fish that were culled, and the pond(s) into which the select brood fish were stocked. When the select brood fish spawn, he should record whether they did or did not breed true. An example of this type of data sheet is presented in Table 8.
Table 8. Example of a data sheet that can be used to record data from a selective breeding programme. In this case, a selective breeding programme that was used to produce a true-breeding population of gold Mozambique tilapia.
| Date: June 1, 1995 | |
| Species: Mozambique tilapia | |
| Breeding programme: Create a true-breeding gold population | |
| Phenotype culled | Number culled |
| Black | 456 |
| Bronze | 935 |
| Phenotype saved | Number saved |
| Gold | 204 females and 196 males |
Did the population breed true?
Yes, on June 23, 1995, the select brood fish produced only gold offspring
Yes, on July 5, 1995, the select brood fish produced only gold offspring
Yes, on July 17, 1995, the select brood fish produced only gold offspring
If the farmer is going to conduct a progeny test, he should record the date on which each mating was made and the results of each mating. He should record whether he saved or culled a fish and should record where the select brood fish were stocked. When the select brood fish spawn, he should record whether they did or did not breed true. An example of this type of data sheet is presented in Table 9. The farmer also must maintain records on where each fish and each family was held until the fish was either saved or culled during the progeny test.
Finally, if a farmer is going to produce heterozygotes for grow-out, he should record the ponds which contain each homozygote, the number of fish in each pond, and when they were stocked; the ponds/tanks where the fish are spawned; the number of brood fish that were spawned; and the results of the matings. An example of this type of data sheet is presented in Table 10.
Table 9. Example of a data sheet that can be used to record data from a progeny test. In this case, a progeny test to cull heterozygous (Rr) red fish and to save homozygous (RR) red fish in order to produce a true-breeding population of red Nile tilapia. Data on individual matings are given only for fish 1–5 in this example.
| Dates: May 29–June 15, 1995 Species: Nile tilapia Progeny test: Progeny test red fish to cull heterozygotes and to keep homozygotes. Normally pigmented fish were used as test fish. | |||
| Date | Red brood fish | Progeny ratio (red:normal) | Cull/Save |
| June 1 | 1 | 156:0 | Save |
| June 1 | 2 | 73:70 | Cull |
| June 1 | 3 | did not spawn | Cull |
| June 2 | 4 | 75:0 | Save |
| June 2 | 5 | 56:61 | Cull |
| Total number of red fish saved: 75 females and 49 males Did the select fish breed true? Yes, on July 25, 1995, the select brood fish produced only red offspring Yes, on July 31, 1995, the select brood fish produced only red offspring | |||
Table 10. Example of a data sheet that can be used to record data about a breeding programme that is used to produce heterozygotes for grow-out. In this case, a breeding programme that mates gold with black fish to produce an all-bronze population of Mozambique tilapia.
| Date: July 25–August 1, 1995 | Species: Mozambique tilapia | |||
| Breeding programme: Mating gold × black to produce 100% bronze | ||||
| Phenotype | Date selected | Pond No. | No. of males | No.of females |
| Black | June 1, 1995 | 23 | 125 | 230 |
| Gold | June 1, 1995 | 32 | 119 | 198 |
| Production of bronze fish | ||||
| Date | Pond No. | Phenotype/sex | Phenotype/sex | Offspring |
| August 1 | 2 | 23 gold males | 45 black females | 8,300 bronze |
| August 1 | 3 | 50 gold females | 30 black males | 9,500 bronze |
| August 1 | 4 | 45 gold females | 32 black males | 10,300 bronze |
The selective breeding programmes that were outlined in this chapter are not technically difficult and they require minimal record keeping. The breeding programmes needed to produce true-breeding populations are simple and, in most cases, can be conducted in a single breeding season. In some cases, they can be conducted in a single day. Once a true-breeding population is created, selection is finished. Even if a true-breeding population cannot be produced, a simple breeding programme can be designed to produce only the desired phenotype.
The selective breeding programmes that were outlined in this chapter have been used with important aquacultured species to produce populations with only the desired phenotype. For example: The undesired saddleback phenotype (deformed dorsal fin) in the Auburn University strain of blue tilapia was eliminated by a single act of selection, because it was produced by a dominant allele. State fisheries agencies in West Virginia and Pennsylvania (U.S.A.) use both selection and the mating of homozygotes to produce both golden (selection) and palomino (mating of homozygotes) rainbow trout for stocking in public waters. Many common carp farmers in Europe fixed the mirror scale pattern in their populations by culling all other scale phenotypes. Finally, ornamental fish farmers routinely conduct these types of breeding programmes to produce more valuable fish.
Prior to selection, a farmer or farmer cooperative should conduct an economic analysis to determine if a selective breeding programme should be conducted. Two pieces of information are needed: The first is the market values of the phenotypes. The second is the cost of production of the phenotypes, which will be determined by the pleiotropic effects, such as growth rate, survival, etc. If the results indicate that a farmer can increase his profits by fixing a prettier body colour, etc. then he should conduct a selective breeding programme. On the other hand, if the results indicate that increased production costs would exceed the increased market value of the crop, then the farmer should not conduct a selective breeding programme.
A farmer also needs to know when selection is unnecessary because the problem does not need to be corrected. All populations contain deformed individuals. In the wild, fish with deformities are generally eaten by predators, so the percentage of deformed fish that are observed is quite small. Fish farmers exclude predators from their ponds, so fish with deformities are more numerous, even though the incidence is still usually rare. Farmers . often think that they have a problem because the odd-looking fish are very noticeable and they remember them. If the frequency of a deformity is less than one in 250 fish, the occurrence is probably not a problem that needs to be solved. On the other hand, if the deformity occurs in one out of every 100–200 fish, the problem should be investigated, and selection may be necessary if the deformity is heritable.
Finally, when a farmer conducts a selective breeding programme, he should save 100–200 select brood fish. The reason for saving this many fish is that a farmer should spawn at least 25 males and 25 females every breeding season. If this is done, he can prevent inbreeding from reaching levels that cause problems.
![]()
![]()
![]()