Section 5 - Introduction to general taxonomy and biology/ ecology of stored products insect pests
The classification of storase insects
The biology of some important primary, secondary
and associated species of stored products coleoptera
Moths of economic importance infesting stored
products: Selected notes on bionomics and identification
The classification of storase insects
INTRODUCTION
Insects would have to be considered the most successful group of animals in the world if the basis for this statement was on the immense number of species already described (1 million ea) and the population densities that can be and so often are attained. Approximately 85% of all known animal species presently existing on earth are insects and it is generally assumed that only one fifth of all insects have been classified and named, meaning overall numbers of species would be approaching 4 million!
Because insects have existed on earth ever since the Palaeozoic era (230-570 million years ago) and those resembling existing present day forms have been found in the rocks of the Permian period (230-280 million years ago), it is little wonder that due to their potential adaptation, diversification and immense numbers of different species, some have found an ecological niche in stored grain!
The advantages of small size and short generation intervals are important in their survival, when you consider that small animals need less food and oxygen to maintain them; small animals can find more room in a given area than larger ones and can find shelter where larger animals can find none.
If we exclude casual visitors to stores and warehouses (i.e. crickets, grasshoppers and other field related pests), we find that stored products range in size from approximately 1 mm-12 mm, while the majority do not exceed 5 mm. Therefore a gap in floorboards, or any cracks on crevices in which floor sweepings become lodged, may support an insect population sufficient to give rise to serious infestation of grain stocks being received into the warehouse.
Insects are members of the superclass Hexapoda, class Insecta of the Phylum Arthropoda (Latin meaning "jointed legs") which also contains the arachnids, crustaceans, myriapods, pycnogonids merostomes, symphylans and trilobites (extinct). Insects have the following morphological characteristics as being representative of the class;
(i) invertebrate animals that have a cuticle hardened to from
an external skeleton
(ii) body divided into obvious segments
(iii) three pairs of jointed segmented legs
(iv) most insects have bodies clearly divided into
Like all animals, insects are classified into various groupings or taxa, the basic unit being the species defined as, "a group of inter breeding organisms which is reproductively isolated from other related groups." Systematic taxonomy attempts to classify animals according to either their evolutionary relationships (phylogen) or by morphological divergence (phenetic affinity). A simplified representation of the class Insecta is given in Appendix I.
The following insect orders are most commonly encountered in stored products.
| a)COLEOPTERA: | Beetles-most of these are either primary or secondary pests, some are associated pests such as scavengers, fungus-feeders, structural pests or predators. |
| b)LEPIDOPTERA: | Moths-either primary or secondary pests, the larval stage inflicting losses. |
| c)HYMENOPTERA: | (i) wasps-beneficial parasites or parasitoids on beetles and moths. |
| (ii) ants-usually scavengers, but occasionally pests. | |
| d) PSOCOPTERA: | Dustlice-scavengers and pests, particularly in Southeast Asia where they sometimes occur in prolific numbers. |
| e) HEMIPTERA: | Bugs-some incidental pests but mostly beneficial predators on beetles and moths. |
| f) DIPTERA: | True flies-some invaders or incidental intruders, some are scavengers and a few are predators. |
| g) DERMAPTERA: | Earwigs-incidental intruders and scavengers. |
| h) ISOPTERA: | Termites-structural pests in warehouses of wooden construction. |
| i) THYSANURA: | Cockroaches-scavengers which may occasionally approach pest status. |
| k) ORTHOPTERA: | Grasshoppers, crickets and locusts-most are incidental field pests, but the "house cricket", Acheta domesticus Linnaeus, occasionally becomes a nuisance indoors, eating almost any kind of food or refuse. |
| l) SIPHONAPTERA: | Fleas-bloodsucking ectoparasites of mammals and birds. Wingless, with complete metamorphosis. 1900 species of which Pulex irritans Linnaeus and Xenopsylla cheopis Rothschild are sometimes found in granaries and warehouses. |
Nearly one thousand species of insects have been associated with stored products throughout the world, of which the majority belong to the Coleoptera (60%) and Lepidoptera (8-9%).
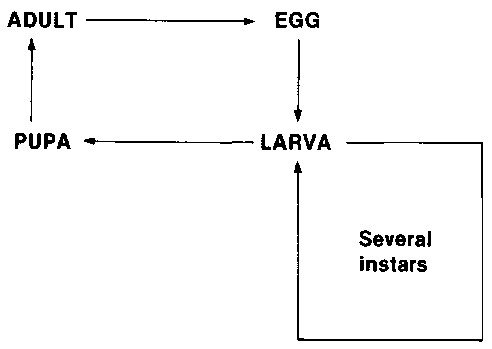
Beetles and moths are advanced, holometabolous types of insects which have a life-cycle we describe as COMPLETE METAMORPHOSIS. This life-cycle has two very important features:
(i) Because an insect has an external skeleton that cannot grow or stretch, a growing insect has to shed its skin (external skeleton) so that it can expand its size while the new skin is still soft. The development of an insect happens in several stages, and at the end of each stage the old skin is shed (the discarded old skins are called "exuvia").
(ii) The immature insects (larvae) that develop from the eggs have evolved as rather simple stages designed for feeding and growing. They are completely different in appearance from the mature adults. Therefore at the end of the period of larval growth, the larva becomes a pupa. This pupa does not feed or move, but inside its skin the body becomes completely changed into the form of the adult. At the end of the pupal stage, the mature adult emerges from the pupal skin. The four basic stages-egg, larva (consisting of several growth stages, known as "instars"), pupa and adult are completely different in appearance.
The life-cycle of beetles and moths is summarized in the following diagram. The larvae of all beetle and moth storage pests feed on the stored food. The adults lay eggs in large numbers during their lifetime. The adults of many important beetle pests also feed on the stored food, but the adults of some beetle pests and all moth pests are shortlived (laying many eggs very quickly) and do not feed.
Importance of the major pests in world cereal production:
Stored grain insects can be grouped according to their relative importance on a particular commodity in a particular geographic region. Species can be classified according to frequency and abundance on particular commodities, such that "major insect pests" are found frequently due to their almost cosmopolitan distribution and in large numbers. Examples are Sitophilus oryzae Linnaeus and Tribolium castaneum Herbst. "Minor insect pests" would then simply mean those species that are encountered infrequently and at relatively low population densities.
A more frequent and misunderstood classification is based on the insects capacity or ability to infest sound, undamaged kernels. On whole cereal grains, pulses and oilseeds we often distinguish between primary and secondary pests, which have the following distinguishing characteristies; primary pests are:
secondary pests are
It must be emphasized that primary pests does not necessarily mean more important, but simply the dynamic processes involved whereby secondary pests can cohabitate or follow primary pests and inflict serious and economic losses, especially under longterm storage. Controlling primary pests can reduce the potential damage caused by secondary pests. Some pests do not fall into either specific category, whereby some secondary pests can attack whole grain if the moisture content is high, some preferentially feed on the germ while others can breed on whole grains, but do so much more rapidly on damaged grains. A more convenient and meaningful classification of the relative importance of insect pests, can be best described in relation to both losses incurred on different commodities and the absolute need to implement control strategies to prevent economic loss.
| Pests of Major Importance - | Those that regularly cause significant losses of commodity or require applied control measures to prevent such losses; |
| Pests of Moderate Importance - | those that occasionally cause significant losses or need control measures; |
| Pests of Minor Importance - | those that rarely cause loss or need control but are often associated with a commodity. |
| Rare or Absent Pests - | those that are rarely found in or are absent from a commodity. |
The percentage of maize, paddy, rice, sorghum and millet at risk to damage by insect pests are presented graphically in Appendix II.
Paddy:
Unhusked rice (paddy) is usually considered more resistant to insect attack than most other cereals. Moisture content is particularly important and field infestations of S. cerealella are the main problem. This pest is of major importance in areas yielding over 80% of paddy production. Infestations occur particularly during after harvest but do not persist in storage. The initial attack, however, allows other species such as O. surinamensis and to a lesser extent T. castaneum to become established. These species may also be prevalent as a result of husk damage or breakage of grain during harvesting or threshing, for example from combine harvesting. Where infestations of S. cerealella do persist in storage, they occur primarily in the surface layers and particularly around the edges of bulk stored paddy. Bagged paddy is less liable to attack, especially if stored in close-woven bags.
S. oryzae and R. dominica may cause serious damage in parboiled paddy and in varieties less resistant to attack particularly at comparatively high moisture contents. R. dominica predominates in the drier batches when these are left undisturbed for long periods. There is considerable confusion as to the relative roles of S. oryzae and S. zeamis. S. oryzae appears to be the dominant species in the dry areas and S. zeamais in the warm moist areas. S. zeamais, for example, is not common in the Middle East or in India where 5. oryzea is predominant, whereas in Southeast Asia, S. zeamais is particularly abundant in milled rice and maize. The occurrene of field infestation, the greater mobility or S. zeamais, the rapid turnover a stocks in mills and the associated production of maize appear a probable combination of factors that, together with high local temperatures and humidities, would favour this. However, it has been demonstrated in Indonesia that S. oryzae appears the dominant species on paddy (Haines, 1981; Haines and Pranata, 1982).
Most other species are of little consequence except that occasional outbreaks of C. cephalonica and T. granarirm may occur.
Rice:
Milled rice is particularly susceptible to insect attack. It is used to store the grain as paddy and process or mill it as required, keeping the storage period of the processed product to a minimum.
S. oryzee is the perdominant species. It has been recorded as a major pest in areas that yield 80% of rice production. S. zeamais has not received a high rating as a pest but, as with paddy, its real importance is probably much greater than appreciated because of its being confused with 5. oryzae particularly in eastern Asia and in Central America and the Caribbean.
B. dominica has been recorded as a major pest in areas yielding approximately half of rice production but because of the limited storage periods usually involved, this species would not be expected to cause damage of the same order as 5. oryzae. S. oryzae. C. cephalonica and T. castaneum have frequencies of occurrence similar to R.. dominica, and would be of greater significance. O. surinamensis and E. cautella must also be taken into account as significant pests though they do not occur as frequently as the species listed earlier.
Rice bran is a commodity notoriously susceptible to storage pests. It provides a serious source of infestation in many rice mills- T. castaneum is the principal species involved.
Maize:
World production of maize is of the same order as rice (considered as dehusked paddy) and not substantially less than wheat. The export trade is second only to wheat but because maize has a greater association with subsistence farming and local animal feeding, a lower proportion enters international trade. The two grains together account for approximately 80% of the world export trade in cereals.
Maize, more than any other cereal, is subject to field infestation. The only other cereals in which field infestation is problem today are the sorghums and paddy where infestations primarily concern S. cerealella attacking unthreshed grain left drying in the field after harvest. Maize, in addition to being attacked by S. cerealella, is heavily attacked when standing in the field at early stage of ripening by Sitophllus zeamais. This species may complete one or even two life cycles before harvest. Such field infestations which are often accompanied by a complex of Nitidulidae, Lathridlidae, Mycetophagidae and sometimes in later stages species such as T. castaneum, reflect varietal varietal characteristics of the maize plant such as husk length, and their effect may be augmented, sometimes independently, by damage from birds and field crop insects such as Heliothis. The problem may be accentrated by crib drying and storing of maize cobs.
As with sorghum, varietal differences in hardness of the grain also can have a marked influence on severity attack.
Sitophilus oryzae is again the most important pest. S. Zeamais is considerably less important though its importance probably would have been somewhat not separating it from S. oryzae in some maize-producing countries where it is known to be a pest were taken into account. The pattern of occurrence of the two species conforms reasonably with what could be expected from climatic considerations-in the warmer often more humid areas, S. aramais is more important than S. oryzae (e.g. Australia, Philippines, Malaysia, Burma, Zambia, Nigeria and El Salvador), with S. zeamais becoming less important as the humidity decreases (e.g. Egypt, India and Botswana), or the temperature decreases (e.g. Argentine, Canada, Czechoslovakia), Both species appear absent in very hot areas such as in the Sudan but in most instances the lower itmperature limits for maize appear not to preclude either species being present at least as pests of minor importance. S. granarius can also be regarded as a significant pest of stored maize particularly in these cooler areas, and in storage of imported maize in countries which it cannot be grown.
S. cerealella is of comparable importance to the Sitophilus spp. As with sorghum stored on the head, maize stored on the cob is liable to reinfestation in storage increasing the potential of the species. The larger grain size and hence intergranular spaces of maize allow some movement of S. cerealella in shelled grain and thus increase the chance of reinfestation in grain bulks. This is not common with the smaller more tightly-packed grains but when it occurs infestations are usually confined to the surface and the periphery of bulks.
A list of the important primary and secondary pests for maize, as well as groundnuts, mungbeans and soyabeans is given in Table 2.
| COMMODITY | CLASSIFICATION | SCIENTIFIC NAME COMMON NAME |
| MAIZE | Primary Pests | Sitophilus zeamais Motschulsky - Curculionidae (COL) |
| "Maize Weevil" | ||
| Sitophilus oryzae (Linnaeus) - Curcu - lionidae (COL) | ||
| "Rice Weevil" | ||
| Rhyzopertha dominica (Fabricus) - Bostrychidae (COL) | ||
| "Lesser grain borer" | ||
| Sitotroga cerealella (Oliver) - Gele - chiidae (LEP) | ||
| "Angoumois grain moth" | ||
| Secondary Pests | Tribolium castaneum (Herbs") - Tene - brionidae (COL) | |
| "rust-red flour beetle" | ||
| Cryptolestes ferrugineus (Stephens) - Cucujidae (COL) | ||
| "rusty grain beetle" | ||
| Cryptolests pusillus (Schonherr) - Cucujidae (COL) | ||
| "flat grain beetle" | ||
| Oryzaephilus surinamensis (Linnaeus) - Silvanidae (COL) | ||
| "saw-toothed grain beetle" | ||
| MAIZE | Secondary Pests | Oryzaephilus mercator (Fauvel) - silva- nidae (COL) |
| "merchant grain beetle" | ||
| Latheticus oryzae Waterhouse - Tenebrioni - dae (COL) | ||
| "long-headed flour beetle" | ||
| Palorus spp. | ||
| P. ratzeburgi (Wissmann) - Tenebrionidae (COL) | ||
| "small eyed flour bettle" | ||
| P. subdopressus (Wollaston) - Tenebrioni dae (COL) ) | ||
| "depressed flour bettle" | ||
| Ahasverus advena (Waltl) - Silvanidae (COL) | ||
| "foreign grain bettle" | ||
| Carpophilus spp. - Nitidulidae (COL) "dried | ||
| fruit beetles" | ||
| Lasioderma serricome (Fabricus) - Ano - biidae (COL) | ||
| "cigarette beetle" | ||
| Lophocateres pusillus (Klug) - Trogositidae (COL) | ||
| "Siamese grain bettle" | ||
| Tenebroides mauritanicus (Linneaeus) - Trogositiae | ||
| (COL) "The cadelle" | ||
| Typhaea stercorea (Linnaeus) - Mycetopha- gidae (COL) | ||
| "hairy fungus beetle" | ||
| Corcyra cephalonica (Stainton) - Pyrali - dae (LEP) | ||
| "Rice moth" | ||
| Ephestia cautella (Walker)- Pyralidae (LEP) "Tropical | ||
| warehouse moth" or "almond moth" | ||
| Plodia interpunctella (Hubner)- Pyra Lidae (LEP) | ||
| "Indian mealmoth" | ||
| Several other minor secondary pests. | ||
| GROUNDNUTS | Primary Pests | Caryedon scrratus (Oliv) - Bruchidae (COL) |
| "ground nut borer" | ||
| T. castaneum (H)-"rust-red flour bettle" | ||
| GROUNDNUTS | Primary Pests | O. mercator (Fauv.) - "merchant grain beetle" |
| Secondary Pests | O. surinamensis (c)-"saw-toothed grain beetle" | |
| A. advena (Walt)-"foreign grain beetle" | ||
| Cryptotestes spp.-"flat grain beetle" | ||
| T. mauritanicus (L)-"cadelle" | ||
| Wechobia rufipes (Degeer) - Cleridae (COL) | ||
| "redlegged horn beetle" or "copra beetle" | ||
| C. cephalonica (Staint.)-"Rice moth" | ||
| E. cautella (Walker)-"Tropical warehouse moth" | ||
| some minor pest species | ||
| PULSES | Primary Pests | Callosobruchus analis (Fabricus) - Bru - chidae (COL) |
| "cowpea weevil" | ||
| Callosobruchus chinensis (Linnaeus)- Bruchidae (COL) | ||
| "cowpea weevil" | ||
| Callosobruchus maculatus (Fabricus)- Bruchidae (COL) | ||
| "cowpea weevil" | ||
| Acanthosselides obtectus (Say) - Bruchi - dae (COL) | ||
| "dried bean beetle" | ||
| Zabrotes subfasciatus (Boh)- Bruchidae (COL) | ||
| Sitophilus oryzae (L)- "rice weevil" | ||
| Secondary Pests L. Serricurne (Fab)-"cigarette betle" | ||
| Stegobium panecium (Linnaeus) - Ano - biidae (COL) | ||
| "Biscuit beetle" | ||
| T. castaneum | ||
| Various minor pest species. | ||
| SOYBEANS | Primary Pests | Callosobruchus spp.-"bean weevils" |
| Secondary Pests | T. castaneum ((H)-"rust-red flour beetle" | |
| P. interpunctella (Hub)-"lndian mealmoth" | ||
| A few minor pest species |
*Source: Dr. C. P. Haines, BIOTROP, Indonesia.
**Special pulse feeding strain-cultured on mungbeans.
The occurrence of Sitophilus oryzae which is capable of attacking pulses has also been recorded by Pemberton and Rodriguez (1980) on carob pods from Portugal. Carobs are normally very susceptible to storage pests which predominanty attack dried fruits, since the pod contains a high sugar content. Certain strains of S. oryzae are also konwn to breed on other pulaes and legumes, such as yellow split peas (Coombs, et al., 1977).
Distribution of stored products insects In Southeast Asia:
Systematic surveys of the presence and distribution of stored products insects has not been attempted on a regional basis in the ASEAN countries. Separate surveys however have been completed on a national basis in the Philippines (Morallo-Rejesus, 1978); Indonesia (Prevett, 1975; Morallo-Rejesus and Pranata, 1978; Haines and Pranata, 1982); Thailand (Chuvit, 1976) and Malaysia (Champ and Dyte, 1976; Lim and Tan, 1978), but no data is available for Singapore.
SOURCES OF INFESTATION
A. CROSS INFESTATION
This may be defined as the infestation of one commodity by movement of insects and mites from one commodity to another. This happens when sound grains are stored side by side with infested materials or by loading both products into the same hold of a ship or the same railroad car.
B. RESIDUAL INFESTATION
Residual infestation results from attack by insects which have remained in the structure of the store, vessel or vehicle after the removal of a previously infested commodity. Grains that have collected in corners, cracks and crevices in walls, floors and ceilings of a warehouse, or in the holds of a bulk tanker or railroad vehicle support insect populations that are capable of reproducing in large numbers and infesting clean grain.
Residual infestation and the potential it creates maybe of greater significance than insects within the bulk, which can be effectively treated or processed before insect numbers develop into large populations. Once insects become established in stores or within the fabric of storage structures, they maybe extremely difficult or impossible to eradicate unless the entire building can be effectively fumigated. This would be relatively expensive and may not be practicable if a sufficient level of gastightness cannot be maintained within the storage structure. The costeffectiveness of the treatment also may not be warranted if cross-infestation from adjacent storages (who are not prepared to carry out extensive pest control measures) cannot be prevented.
(i) Infestation arising from the use of infested containers
Bags and sacks and other repeatedly used containers are often infested by insects. Of these, jute bags are most often infested by flour beetles, warehouse moths, tropical warehouse moths or rice moths. The receival of infested stocks without inspection or the recycling used sacks that are not adequately cleaned and treated with either a fumigant or impregnated with a residual insecticide, further compound the problem and reduce the efficiency of control measures.
(ii) Intestation arising from mills
(iii) Combine harvesters, field bins, augers and transportation equipment
After use, this are usually left in the field for several months and pockets of infestation may occur inside the machinery.
(iv) Grain elevators and related conveyance machinery
C. INFESTATION BY FLIGHT AND CRAWLING OF INSECTS
Some stored grain insect pests are capable of flight and infestations are frequently initiated in this manner. All moths and many beetles move to stored products by flight. Some species also crawl into new uninfested stored products, but migration by crawling is confined within narrow limits.
Field infestations have been dramatically reduced by the adoption of combine harvesting in many Western countries. In the Southern states of the US, cereals especially corn is at risk in the field to such pests as Sitophilus zeamais, Sitotroga cerealella, Sathrobrota rileyi, Nemapogon granella, Araecerus fasciculatus and Carpophilus dimidiatus among others, the first two being the most dominant and serious.
Field activity of S. zeamais and S. cerealella has also been reported in the Southern parts of the USSR (Trisuyatkii, 1966) as well as in the Northeastern states of USA following mild winter conditions (Cotton, 1964). Both species commonly occur in maize grown in the coastal region of NSW, Australia (Greening, 1978), while Sitophilus oryzea infested 9-17% of sampled standing heads of wheat in Queensland during October and November (Rossiter, 1970).
Field infestations can be curtailed by growing corn varieties that possess tight husk characteristics, disposing of infested grain in farm storages before the corn reaches the silking stage in the field, early harvesting, application of recommended grain protectants, drying and shelling and storing corn in tight bins or containers adaptable for fumigation.
Both S. zeamais and Rhyzopertha dominica have been shown to infest sorghum in the field. Preharvest spraying with either Tetrachlorvinphos, Pirimiphosmethyl or Malathion (in order of effectiveness) during the dry or wet season harvest afforded protection is storage for 3 and 2 months, respectively, without any further post-harvest treatment (Caring and Morallo-Fejesus, 1976).
D. NATURAL SOURCE OF INFESTATION
Infestation may also arise from natural sources including nests of birds, rodents, spiders and insects.
FACTORS GOVERNING THE DEVELOPMENT OF INFESTATIONS
The likelihood of a pest infestation developing and the severity of the infestations are determined both by the location of the stored grain habitat and by the specific features of the habitat itself.
(a) Important characteristics of the stored grain 'habitat'
i) Size of the grain bulk is of critical importance. Small bulks are very much subject to the vagaries of their immediate surroundings. Other things being equal conditions tend to become more stable as the bulk grows larger-the effect of climate within a bulk is tempered and the relative importance of mass immigration and emigration declines due to the insulating properties of bulk grain. If it is not moved or aerated, conditions in bulk will be largely governed by the temperature and moisture content of the grain when it was placed in storage.
ii) Shelter offered -masses of grain kept in tightly sealed containers are less subject to the vagaries of climate and less susceptible to mass invasions by biological agents (or to mass exports of the same).
iii) Climate- local conditions of temperature and moisture have the profoundest influence on the susceptibility of grain to infestation and the course of events in these infestations. Generally pest control is much more difficult and grain deteriorates more rapidly in warmer-moisture areas than in drier cooler regions. But as pointed out in (i) and (ii) the effects of climate may be greatly altered both by the size of the bulk and the degree of shelter offered by the particular habitat.
iv) The duration of stay of the grain- if the grain only stays a very short time in the habitat, then organisms infesting the grain will be subject to considerable disturbance and the dynamics of infestation will be greatly altered. While ephemeral habitats may serve as sites for initiating infestation they are not places where pest numbers increase to any extent; rather these are often places which test the survival capacity of the pest. It is only in undisturbed sites, preferably sheltered and preferably with at least modest quantities of grain that infestations can develop to damaging levels.
v) Proximity of sources of infestation. Some habitats such as poorly-cleaned storages may have adjacent sites harbouring numerous pests which can quickly invade incoming grain. Other habitats may be far from any such reservoirs and hence, other things being equal, should present less difficulty from a control point of view.
(b) Important ecological characteristics of storage insects
i) In the protected environment of grain, insect mortality is low in the immature stage. As a result, their capacity for increase is great.
ii) As the temperature inside stores is usually higher and more stable than the external environment, the effective temperature is more conducive for the development of storage pests than for field pests. Furthermore, many of stored product pests have originated from tropical or sub-tropical regions, thus they are capable of reproducing continuously throughout the year.
iii) Stored grains are usually hard and are relatively dry (as food for pests grains are usually stored at between 9 and 14% m.c., depending on the type of grain, whereas growing plants are at about 90% m.c.). The important pests are adapted to feed on hard food and to survive on a very low water content in their diet. They are quite sensitive to grain moisture content and equilibrium relative humidity.
Both (ii) and (iii) are optimized in the humid tropics for the rapid development of stored grain pests, i.c. temperatures between 27 and 32°C and equilibrium relative humidities between 70 and 75%. As a result storage insects are able to develop at more-or-less the same rate throughout the year. Most of the important pests have generation times of between 3 and 6 weeks on most products in tropical stores, i.e. between 8 and 16 generations per year.
iv) Insects that are small in size are extremely suitable for living in stockpiles of grain. Light affects different species, and different stages of development, although they commonly prefer darkness. There are many of the adult species that have already lost their flying ability.
v) The movement of infested parcels of grain and grain products in international trade has provided an effective means of ensuring the widest possible geographic spread of storage pests since cereals were first harvested. The continued introduction of the major pest species into climates outside their normal distribution, ensures also that all these species may be found in all parts of the world.
Because of the bulk and relatively low value of cereals, international transport is almost exclusively by sea although rail and road transport may become significant in limited areas. The infestation which occur in ships originate either in the commodity before loading or from cross-infestation from residues of previous cargoes or other infested cargoes being carried. It is often difficult, however, to determine the precise origin of particular infestations.
DYNAMICS OF THE GRAIN COMMUNITY
Clearly the amount of damage done to a bulk of grain is dependent upon the number and kind of pests present in the grain and the time available for these organisms to do the damage. Accordingly it is not unreasonable to center one's understanding of the events in grain around the population dynamics of the pest.
An insect population in bulk grain experiences a certain rate of population growth (or decline as the case may be) and the population acquires a particular structure. Different stages of insect may be more damaging to grain than others. Growth rate and composition are products of factors influencing survivorship of adult and developmental stages, rate of development to maturity and the egg laying schedules to females. The insect population is influenced both by its environment-physical and biological conditions in its surroundings-and by its own size and composition. Depending on the species, insects may feed on whole grains, frass from other insects, fungi, grain particles and dust or other insects. In addition, the metabolic heat of insects and the water released by them encourages their own growth and that of other pests (mites, fungi and other insects). The chain of cause and effect is complete when insects influence the very factors which had affected their population growth.
The dynamic processes associated with the growth and development of stored grain insects is summarized in Table 3.
Influence of abiotic environmental factors on pests of stored grain
(a) Temperature
Below a certain minimum threshold temperature insects do not complete development from egg to maturity and the pest population cannot increase. At temperatures only slightly above their threshold, say within 4-5°C, mortality rates are extremely high for virtually all stages of development. Most species do not multiply fast enough to become a pest until temperature is somewhere between 3-6°C above the minimum threshold for development.
The highest rate of oviposition usually occurs at a temperature about or just below the developmental optimum. Temperature has a profound impact on insect and mite locomotion, the net effect being that insects tend to move out of areas where temperature is unfavourable and relocate in more favourable zones.
Temperature effects on a pest is not always direct. Different members of the ecosystem have different temperature optima, hence temperature influences processes such as competition, predation and ecological succession. Temperature is also the main variable determining the rate of population growth, insect activity, the liberation of heat and moisture and the production of frass and other waste products which lead to grain deterioration.
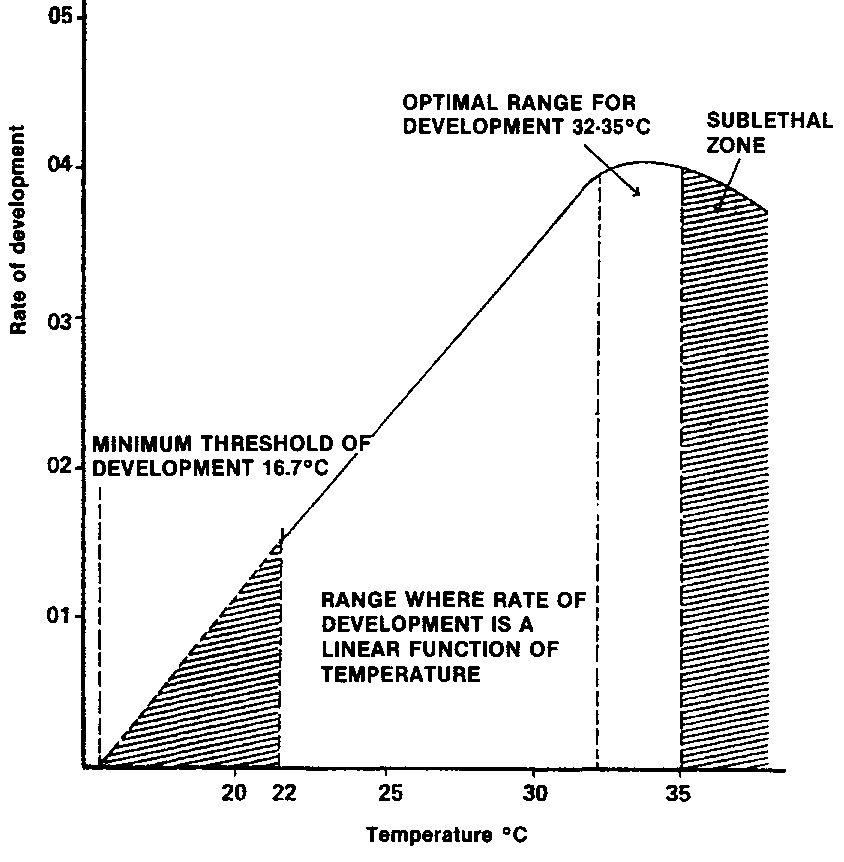
The effects of temperature are graphically illustrated in Fig. 1, using Tribolium castaneum (Herbst) as the example.
In general, the higher the temperature, the shorter the developmental period. The most favorable temperature for development and reproduction is about 28°C. Below 21°C and above 35°C reproduction almost ceases. Developmental period from egg to adult becomes longer and the number of eggs laid fewer, at lower temperatures. At higher temperature, the life cycle becomes shorter while the oviposition rate increases. Rice weevils, for insxance, complete their life cycle in 25 days at 30°C while they take about 94 days at 18°C. Most insects grow more quickly at 2932°C
At a temperature over 34°C, insects usually cannot develop. Lesser grain borers and Khapra beetles will however continue to develop at 34°C and 40°C, respectively.
The lower temperature limit for insect development depends on the species. It is 14-15°C for the rice weevil while it is 18-19°C for the rust-red flour beetle. These limits are the most important factor affecting the geographical distribution of stored insect species.
(b) Moisture
Different developmental stages of insects, mites and fungi have different susceptibilities to aridity or excessive moisture. Generally the dormant stages-eggs and pupae for insects, eggs and resting stages of mites, and spores of fungi can best resist desiccation while active feeding stages may die out if conditions are too dry. Tolerance depends on the species-some insects may increase even at a R.H. of 10-30%, while miters and other insects require at least 65% R.H. and fungi at least 70% R.H. (Ideal range for reproduction and development in storage insects in 65-80% R.H.).
Insects depend primarily on their food supply for moisture to carry on their life processes. Stored product insects can develop on food with moisture content as low as 2 to 14%. Moisture requirements vary with species. Generally, optimum grain moisture for development and reproduction is 14-18%. Most species do not develop below 10% m.c.; some exceptions are the Mediterranean flour moth and Khapra beetle can develop on food with negligible moisture content (down to 2%), weevils cannot live on grain containing moisture of less than 10% and lesser grain borers can subsist on grain with 9% moisture content while the confused flour beetles can live on 2% moisture grain.
Moisture and temperature interact to provide conditions favorable for multiplication and survival of stored product insects. As temperature is increased, insects are able to reproduce in grain of lower MC and when moisture is increased, they can multiply at lower temperatures.
(c) Gas concentrations
Metabolic activities of grain, but to a much greater extent of mites, insects and especially fungi may cause local reduction in the concentration of oxygen and increases in carbon dioxide. As grain settles, intergranular passageways become constricted or even blocked if dust or grain particles are present. Low levels of oxygen and high levels of carbon dioxide cause metabolic stress on insects, mites and aerobic microorganisms generally increasing mortality while lowering fecundity and rates of development. If oxygen should drop below 2.5% conditions become lethal for insects, but fungi can survive with levels lower than 1 % O2.
Source: ADAB International Training Course on the Preservation of Stored Cereals, Selected Reference Papers, Part I (1975).
Table 4. Lower limits of relative humidity, temperature, optimal range of temperatures and degree of hardiness of pests of stored grain, together with estimate of maximum rate of increase per lunar month
| Insect | Min. Rel. Humidity % | Min. Temp. °C | Opt. Temp. °C | Maximum Rate of Increase + |
| Acarus siro | 65 | 7 | 21-27 | 2,500 |
| Ephestia kuehniella | 1 | 10 | 24-27 | 50 |
| Ephestia elutella | 30 | 10 | 25 | 15 |
| Ptinus tectus | 50 | 10 | 23-25 | 4 |
| Endrosis sarcitrella | 80 | 10 | 24-26 | 30 |
| H.pseudospretella | 80 | 13 | 24-26 | 2 |
| Sitophilus granarius | 50 | 15 | 26-30 | 15 |
| Sitotroga cerealella | 30 | 16 | 26-30 | 50 |
| Gnathocerus cornutus | 40 | 16 | 24-30 | 15 |
| Stegobium paniceum | 60 | 14 | 25-28 | 7.5 |
| Sitophilus oryzae | 60 | 17 | 27-31 | 25 |
| Ephestia cautella | 25 | 17 | 28-32 | 50 |
| Plodia interpunctella | 40 | 18 | 28-32 | 30 |
| Corycra cephalonica | 30 | 18 | 28-32 | 10 |
| Dermestes masculatus | 30 | 20 | 30-25 | 30 |
| Oryzaephillus mercator | 10 | 20 | 31-34 | 20 |
| Cryptolestes turcicus | 50 | 21 | 30-33 | 50 |
| Oryzaephilus surinamensis | 10 | 21 | 31-34 | 50 |
| Tribolium confusum | 1 | 21 | 30-33 | 60 |
| Lasioderma serricorne | 30 | 22 | 32-35 | 20 |
| Necrobia rufipes | 50 | 22 | 30-34 | 25 |
| Tribolium castaneum | 1 | 22 | 32-35 | 70 |
| Cryptolestes ferrugineus | 10 | 23 | 32-35 | 60 |
| Trogoderma granarium | 1 | 24 | 33-37 | 12.5 |
* Minimum temperature at which epidemic
increases may occur, i.e.3-5°C above the development minimum.
+ Maximum rate of increase for stable age population per lunar
month.
Source: Howe (1965).
Influence of biotic environmental factors on pests of stored grain
(a) Food:
Grain itself is just one of several resources used by pests for nourishment-in addition they may feed on damaged grains, grain particles and dust, fragments of insects and mites, including frass and cast skins, dead and living arthropods and fungi. Thus, fungus beetles and certain moths show marked preference for mouldy grains or frass from other insects; certain mites make their living by feeding on other mites, and flour beetles supplement their diet by feeding on the dormant stages of their own and other species.
Stored grains are an ideal food source for stored product insect pests, providing the essential elements required for continued growth and development. Levels of carbohydrates, proteins, fats and vitamins required varies with the species concerned. The Indian meal moth, warehouse moth, Mediterranean flour moth, and saw-toothed grain beetle cannot live in artificial diet containing carbohydrates of less than 20%, while the confused flour beetle and cigarette beetle can live on foodstuff with less than 2% carbohydrates.
The saw-toothed grain beetle can grow with 2-3% protein of artificial diet and hide beetles need at least 20% protein. Insects need 10 kinds of essential amino acids: arginine, isoleucine, leucine, Iysine, methionine, phenylalanine, threonine, trytophan and saline.
Some Insects utilize fats in food materials as their source of energy, and some, such as the warehouse moths and tropical house moths, fats are indispensable. Their adults will partially or completely lack scales on their wings or fail to emerge from pupae if fats are not available.
Most of the vitamin B group such as thiamine (B.), riboflavin (B2), pyridoxine (B6), nicotinic acid, patothenic acid, biotin, folic acid, and choline are indispensable to all insects.
However, the saw-toothed grain beetle, drugstore beetle and cigarette beetle can live on food materials without some B group vitamins, not because they can dispense with these vitamins, but because they have in their bodies some intercellular symbiotic microorganisms which are able to synthesize and supply such vitamins. Vitamins A and C are not required by any of the stored products insects.
(b) Competition
At low and moderate population densities members of a given species will improve conditions for their fellows. Initial attacks on hard grains may make things easier for the next, while heat and moisture released by metabolic activity may push temperature and moisture into more favourable zones or at least encourage the growth of other species. Beyond optimal population densities, the scramble for food, for mates and for oviposition sites enhances mortality, lowers rates of development and leads to lower fecundity.
Members of various species residing in stored grain have considerable influence on the welfare of other consumer species. Thus as a larva Tribolium castaneum, the fulur beetle benefits from encounters with eggs of say Ephestia cautella the moth, which it feeds on, but as an adult the beetle may get injured by a larval moth. Insects and mites may lower the fecundity of the other species simply by mechanical disturbance-jostling-or by releasing noxious chemicals. Feeding and release heat, water, and waste by some consumers may pave the way for succession by other consumers, or species may engage in a scramble for resources causing such depletion that other consumers die out or are forced to leave the locality.
The competition for food is more severe between species which have the same nutritional and ecological requirements. Parasites (mostly hymenopterous) and predators have, to some extent, helped in reducing insect populations. Some insects such as Tribolium spp. or flour beetles exhibit cannibalism.
TYPES OF DAMAGE
A. Direct damage
1. Reduction or loss in weight:
The direct feeding of insect pest on stored grains results in food and weight losses. A rice weevil will eat 14 milligrams of a rice kernel weighing about 20 milligrams during its developmental period from egg to adult. However, this results in the commercial loss of the whole rice kernel.
Weevils, which feed mainly on the carbohydrate portion of a maize, remove a considerable amount of the calorie potential but little of the protein and vitamins which are mainly in the germ and bran. In legumes, in which the protein and vitamins are more evenly distributed throughout the grain, infestation of beetles, which can be responsible for consuming up to 50% of peas and beans of which some 25% of the dry matter is crude protein, can cause a loss of about 12% of the available protein.
2. Reduction or loss of seed viability:
A seed grain which has been attacked by a germ feeder will not germinate. Examples are Triblim spp., Cryptolestes spp., Tenebroides mauritanieus, Ephestia spp.
B. Indirect damage
1. Quality loss
a) Nutrient loss. When grains are attacked by insect species which feed selectively on the germ leaving the endosperm almost untouched, food loss is not apparent; weight loss is also small compared to loss of loss of vitamins, proteins, etc.
b) Heating and spotlage. Heating from insect infestation accelerates further infection by microflora and bacteria. This results in spoilage of grain which often causes more serious economic damage than loss in weight by insect consumption.
c) Contamination, tainting or discoloration. Contamination and tainting of foodstuffs with frass materials such as insect fragments, excrete, secretions, webbings and dusts also contributes to a deterioration in quality. Insect secretions or entomoxins, such as quinone, may also prove to be serious contaminants.
d) Production of off-flavors and odors. Cereal, after prolonged storage, but particularly when ground into a meal or flour or when infested by insects, show an increase in free-fatty acid content (FFA). If, prior to milling, the maize has been attacked by adults of Tribolium castaneum, the initial faa of the resultant meal is much higher than that of uninfested meal, and the FFA rises to a much higher level.
2. Monetary loss
Financial losses are not simply in terms of that lost in reduction in weight, but also the downgrading or absolute rejection through the presence of live insects or signs of their activity. The cost of any applied control measures, as well as all the agricultural inputs invested in growing the grain, become important "hidden'' costs if grain is consumed or destroyed by insects before reaching the consumer.
3. Loss of goodwill
A country loses its goodwill and earns a bad reputation if its produce are of low quality, insectinfested and contaminated by toxins, insect, excrete, etc.
STRATEGY FOR SURVIVAL
Successful storage pest insects have adopted strategies that can be summarized as being opportunist. By a variety of biological mechanisms, all these species are capable of very rapid population growth when they invade a batch of grain in a suitable condition. Most pest species have potential rates of increase of between 20 and 60 times per month under optimum conditions (See Table 4). We should consider what this means by thinking about a pest with a 40-times-permonth rate of increase: One gravid female invading the grain can lead to 40 individuals after one month, 1600 individuals after two months, 64,000 after three months, 2,560,000 after four months.
Fortunately, at some point in the development of the population, natural or artificial (pest control) factors operate to control the increase. If pest control is not applied, the commonest natural limiting factor is competition for available food while other forms of competition or disturbance, presence of fungal growth and mycotoxins, parasites, predators and pathogens, pheromones and defensive secretions from other species, and cannibalism become contributory factors in suppressing insect populations.
Levels of temperature and moisture liberated by the development of an insect population often becomes excessive for the survival of the pests, eventually forcing the insects to relocate themselves elsewhere, thus increasing the hotspot to a point where adults can only survive in the superficial layers or on the surface. The very success of a species or species complex within a population may very well form the basis for its own extinction.
As well as being opportunists in taking advantage of abundant food placed in a store, most major pest species also have a survival strategy for the time when the food is removed. in many cases, this involves the establishment of small residual populations which immediately have the potential to infest a new batch of grain entering the store. Another survival strategy found in many primary pests is their ability to disperse to ripening grain in the field. Finally, many species rely on human distribution and transportation to implant them in another storage locality. The use of second hand untreated sacks helps promulgate and distribute major storage pests, and is perhaps one of the major causes of cross-infestation.
As mentioned previously, storage insects have become adapted to surviving in a low moisture ensvironment relative to their field pest counterparts. Any significant changes in temperature and relative humidity (which are functions of the equilibrium moisture content of different grains) will have a critical effect on the rate and development of insects, as well as the composition or species complex constituting a pest population.
It must be emphasized that the accepted level or "safe moisture content" to which grain should be dried does not prevent attack by insect pests, and in fact a relative humidity of 70% is well within the tolerance of most major pest species. Oftentimes in the humid tropics, grain possesses a moisture content somewhere within the 70-75% e.r.h. range. At the higher levels, the development of storage fungi is delayed, but has the deleterious effect that many major insect pests develop much more rapidly and many minor insect pests that are inhibited at 70% r.h. attain pest status.
If grain is not fully dried to the safe moisture levels, then
i) the slightly softer grain allows attack by many pests that initially cannot develop on sound, undamaged kernels.
ii) higher humidity becomes more favourable to a wider range of pests, and
iii) presence of slight but significant fungal growth allows rapid increase of certain pests that require a certain proportion of fungus in their diet. Finally, it should be noted that drying must be uniform throughout a batch of grain. The presence of a few grains that are not fully dried can allow the rapid development of some insects consequently producing more moisture, and allowing the development of pests in grains that were originally dry but have absorbed the extra moisture. The infestation ''hotspot" can thus expand from a very small volume and become a serious problem throughout the entire batch of grain unless some measure of control is implemented to reduce the population and disperse increased moisture.
REFERENCES
CARINO, F. O. and B. MORALLO-REJESUS. (1976). Control of Sitophilus zeamais and Rhyzopertha dominica by pre-harvest sprays and sack treatment with three insecticides in three sorghum varieties. Phil. Agric. Vol. 60 (1976) 3 and 4, AugSept 81-95.
CHAMP, B. R. and C. B. DYTE. (1976). Report of the FAO global survey of pesticide susceptibility of stored grain pests. FAO Plant Protection Series, No. 5. Rome, 1976.
COOMBS, C. W., C. J. BILLINGS and J. E. PORTER. (1977). The effect of yellow split peas (Pisum sativum L.) and other pulses on the productivity of certain strains of Sitophilus oryzae (L) and the ability of other strains to breed thereon. J. Stored Prod. Res. 13, 53-58.
COTTON, R. T. (1964). Controlling the Angoumois grain moth. Northwest Miller 271, 19-23.
DAVEY, P. M. (1965). Insect pests of stored products in the tropics and the commodities and conditions in which they occur TSPI (N) 377-386.
GREENING, H. G. (1979). Observations on the occurrence of insect pests of stored grain in New South Wales, Australia, Cont. to the Symposium on Protection of Grain Against Insect Damage. Moscow, 1978. CSIRO Div. of Ent. by D. E. Evans.
HAINES, C. P. and R. l. PRANATA. (1982). Results of a survey of the insects and arachnids associated with stored products in some parts of Java. Paper presented at the Reg. Grains Post-Harvest Workshop Jan. 19-21, Chiang Mai, Thailand (Proc. in press).
HAINES, C. P. (1981). Paddy losses to insects: notes on selected problems of insect ecology, damage, inspection, loss assessment and varietal susceptibility. Paper presented at GASGA Seminar on "Paddy Deterioration in the Humid Tropics" Oct. 11-17,1981. Bagulo, Philippines (Proc. in press).
HOWE, R. W. (1965). A summary of estimates of optimal and mininal conditions for population increases of smoe stored products insects. J. Stored Prod. Res. 1:177-184.
KOHLMEYER, R. W. (1981). The agricultural market place of the 1980's.
MORALLO-REJESUS, B. (1978). Stored grain pest problems and research needs in Southeast Asia. SEARCA professorial lecture in Entomology, July 21, 1978. 41 pp.
PEMBERION, G. W. and A. DE RODRIGUEZ. (1980). The occurrence of a strain of Sitophilus oryzae (L.) (Coleoptera: Curculionidae) breeding in Portuguese kibbled carobs. J. Stored Prod. Res. Vol. 17, pp. 37-38.
ROSSITER, P. D. (1970). Field infestation of the rice weevil in wheat. QLD J. Agric. Anim. Sci. 27, 119-121.
TRISVYATSKII, L.A. (1966). English translation by D.M. Krane. Vol. 1-3 Nat. Lending Lib. Sci. Tech. England, 1969.
FURTHER READING
Setected Reference Papers (Part I) of the International Training Course in Preservation of Stored Cereals. Office of the Australian Development Assistance Bureau, Canberra, Australia.
"Stored Grain Insects", USDA Agricultural Research Service Handbook, No. 500.
"Common Insect Pests of Stored Food Products - A Guide to their Identification", Edited by Paul Freeman, Brit. Museum (Nat. Hist.) Econ. Series No. 15.
"Pests of Stored Products", by J. W. Monro. and "Pest Control in Buildings" (a guide to the meaning of terms) by P. B. Cornwell, Rentokil Library Series.
{kind=link}
{kind=link}
{kind=link}