![]()
![]()
![]()
Abstract
Introduction
Materials and methods
Results and discussion
Acknowledgements
References
H.M. Abdel Magid, A.M.A. El Tilib and E.O. Ahmed
Dept. of Biochemistry and Soil Science,
Faculty of Agriculture, University of Khartoum, Shambat, Sudan
The performance of leucaena (Leucaena leucocephala (Hawaii No. K8, cv. Brewbaker)) was investigated in pot culture experiments, using five arid-zone soils ranging in salinity from 0.75 to 16 ECe (mmhos/cm at 25° C), and sodicity (ESP) from 6.8 to 46. The soil samples were either treated with or not treated with gypsum. Inoculated (Tel No. 1145 strain) and uninoculated replicates were arranged in a randomised complete block design and placed under full sunlight in the greenhouse. Moisture was maintained at field capacity throughout the growth period. The parameters studied included nodulation potential, foliage appearance, fresh and dry weights of tops, shoot and root lengths, N levels in shoot and root, and Ca, Mg, Na, K and Cl contents of plants. The data show that leucaena can grow successfully on these soils.
Vast areas of the arid and semi-arid regions of Sudan consist of salt-affected soils that are poorly productive. The Soil Survey Administration of the Ministry of Agriculture and Natural Resources has surveyed about 400,000 hectares of such marginal lands in the Khartoum, Gezira, White Nile. Northern and Nile Provinces. However, very little research is being done on the reclamation and use of these soils.
The National Academy of Sciences (NAS, 1977; 1979) has recommended research on some legume species (e.g. Leucaena leucocephala) for use as forages and green-manure crops in the fragile ecosystems of the tropics, where vast areas remain barren because of their unsuitability for conventional crops due to poor soil conditions and lack of water. Moreover, Brewbaker et al (1981) emphasised the need for site adaptability studies on Leucaena leucocephala and other tree-legumes in tropical regions.
In recent years species of the genus Leucaena (subfamily Mimosoideae) have received considerable attention in the tropics and subtropics as an economically important plant group. Leucaena has desirable qualities such as high growth rate, drought resistance and tolerance to a wide range of soil and climatic conditions (Thoma, 1983; NAS, 1979; Brewbaker, 1978; Brewbaker et al, 1972).
According to our information there are no reports on site adaptability of leucaena in salt-affected soils. As a result, this study was started to examine the possibility of using such marginal soils as forage reserves, since there is an urgent need for good-quality fodder crops in Sudan. The objective of the work was to examine the performance of inoculated and uninoculated Leucaena leucocephala (Hawaii No. K 8 cv Brewbaker) grown in gypsum-amended and unamended salt-affected soils.
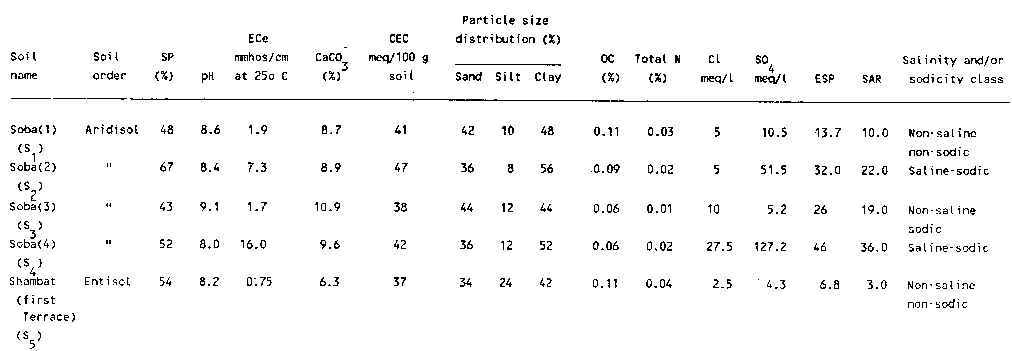
The five soil samples used in this study were selected to include some of the major salt-affected soil groups south of Khartoum (within the immediate vicinity of Soba Agricultural Research Sub-station). The properties of the soils are summarised in Table 1. Organic carbon content was determined by the method of Walkley-Black (1934). Other characteristics were determined according to the methods of USDA (1954).
Table 1. Properties of the soils used.
Composite bulk samples of each soil were brought to the laboratory. They were then ground, sieved through a 2 mm mesh, mixed with gypsum at the rate of 12.5 t/ha and potted in medium-sized plastic buckets, each with 6 drainage holes.
The treatments used were:
g0 = Unammended soil
g1 = Soil amended with gypsum
I0 = Uninoculated soil
I1 = Soil inoculated with rhizobium strain Tall No. 1145.
Treatment combinations were:
g0 I0 = unamended and uninoculated
g0 I0 = unamended and inoculated
g1I0 = amended and uninoculated
g1I1 = amended and inoculated.
Soil and treatments were combined in a completely randomised block design with 4 replicates.
The seed of Leucaena leucocephala (Hawaii No. K 8 cv Brewbaker) and the rhizobium strain (Tel No. 1145, an elite endophyte for Leucaena leucocephala) were supplied by Dr. Paul Singleton, Agronomist, NifTAL Project, University of Hawaii, Paia, Hawaii, USA.
The seeds of many tropical legumes germinate slowly and irregularly unless the testa is removed or made permeable. As such, the leucaena seeds used in this study were soaked in concentrated sulphuric acid for 15 minutes to break down the testa and to sterilise the surface of the seeds. The seeds were then washed with water until all traces of acid were removed.
The sterile seeds were planted in the plastic buckets, which contained 3.5 kg of soil (oven-dry basis) maintained at field capacity. Inoculum was applied to each hole by adding 1 ml of a turbid suspension of the test culture containing 109 cells/ml. Sterile dry gravel was spread over the surface of the soil to reduce the possibility of contamination through the top of the system. Macronutrients, mainly N (as a starter dose) and P, were added at sowing: 45 kg N/ha as urea and 250 kg P/ha as KH2PO4. A starter dose of N was considered to be necessary because the soils used are inherently deficient in nitrogen. A liquid micronutrient concentrate (Monterry Chemical Company, USA) was added at 0.5 ml per kg soil, which provided 7.5 mg Fe; 2.5 mg Zn; 2.3 mg Mn; 1.75 mg B; 0.75 mg Cu; 0.2 mg Mo; and 0.15 mg Co per kg of soil. The buckets were then placed in an unshaded space in the greenhouse during the winter. Weather data for the period of growth are given in Table 2. Moisture was maintained at field capacity by weighing and irrigating according to a consistent schedule.
The plants were cut at soil level 50 days after sowing, and the fresh weight and height of tops were determined. The plants were then dried at 70° C, weighed, ground and composite subsamples of each treatment were used for determination of total N. Ca, Mg, Na, X and Cl (USDA, 1954; Lavkulich, 1978; Chapman and Pratt, 1961). Immediately after the tops had been harvested, the roots were carefully removed from the soil for nodule observations and root total N determination.
Table 2. Weather data for the peroid of growth (12 December - 31 January 1985).
|
Attribute |
Range |
|
Daily minimum temperature (°C) |
10.3 - 21.5 |
|
Daily maximum temperature (°C) |
25.6 - 37.5 |
|
Mean daily temperature (°C) |
16.8 - 33.2 |
|
Daily sunshine (hours) |
6.3 - 10.9 |
|
|
(average 9.9) |
|
Solar radiation (MJ/m2 per day) |
15.4 - 22.2 |
|
|
(average 18.7) |
|
Pan evaporation (mm) |
5.5 - 12.5 |
|
|
(average 9.3) |
|
Wind velocity (miles/hour) |
2.1 - 7.9 |
|
|
(average 5.0) |
|
Rainfall (mm) |
Nil |
Seed germination
Germination counts were made on all the treatments under study. Rapid and uniform germination was evident one week after sowing, and 100% germination was observed in all treatments. The temperature (Table 2) was conducive to germination; alternating temperatures ranging between 20 and 32° C have been reported to induce maximum germination in leucaena (Pathak and Patil, 1982).
Post-emergence growth was quite vigorous in all treatments. The leaves were medium to dark green and the plants appeared to be healthy, which indicates leucaena's potential for growth on such soils. Treatment differences did not show up clearly, however, until the plants were harvested.
Soil and treatment effects
Statistical analysis of the data showed that neither inoculation with rhizobium nor addition of gypsum to the soil had a significant effect on the characters studied, except for shoot length of leucaena, which was significantly reduced (P<0.05) by adding gypsum. However, there were significant (P<0.05) differences among the soils for all characters studied, except for chloride content of the leucaena (Table 3).
Munns and Franco (1981) mentioned salinity, alkalinity, acidity and sodicity of soils, among other factors, as potential constraints on legume production in tropical areas.
Table 3. Effect of soil type on plant characters and nutrient contents of Leucaena leucocephala.
|
Treatments |
Soils |
||||
|
S1 |
S2 |
S3 |
S4 |
S5 |
|
|
Fresh wt. of tops (g/plant) |
1.43a |
0.78b |
0.69bc |
0.46cd |
0.46cd |
|
Dry wt. of tops (g/plant) |
0.46a |
0.28b |
0.24bc |
0.16bed |
0.16bcd |
|
Shoot length (cm) |
14.5a |
11.lb |
9.4cd |
7.3e |
9.2cd |
|
Root length (cm) |
13.0bc |
15.5a |
12.8bcde |
13.4b |
12.6bcde |
|
N % in shoots |
3.4cd |
3.5cd |
3.7bc |
4.4a |
3.9b |
|
N % in roots |
2.6bcd |
2.4bcd |
2.7b |
3.1a |
2.7bc |
|
Ca % |
1.1a |
0.9a |
0.5b |
0.4bc |
0.4bc |
|
Mg % |
0.35a |
0.30ab |
0.30abc |
0.20bcd |
0.20bcd |
|
Na % |
0.06b |
0.07b |
0.12a |
0.11a |
0.02c |
|
K % |
2.5a |
1.6b |
1.2bc |
1.0cd |
1.0cd |
|
Cl % |
0.8 |
0.6 |
0.8 |
0.6 |
0.7 |
Within rows, numbers followed by the same letter are not significantly different at P<0.05.
The soils used in this study comprise soil orders similar to those mentioned by Munns and Franco (1981), viz Aridisols, Vertisols and Entisols. Moreover, the same authors found that the most likely nutrient problems in such soils are deficiencies of P. Zn, K, Mo and S. Some of these deficiencies are unlikely in aridic soils because they are alkaline. However, P fixation via adsorption has been documented by El Mahi and Mustafa (1980).
Leucaena performs poorly in acidic soils that are highly saturated with Al and low in P and Ca (Chee and Devendra, 1982), constraints which are not experienced in the soils under study. on the other hand, Jones (1979), working on Leucaena leucocephala as a feed for ruminants in the tropics, indicated that the sodium content of the plant is consistently low. However, the mineral composition of well grown leucaena is usually adequate for productive livestock (Table 3).
Nodulation potential
Nodulation was very sparse (< 10 nodules per plant) in the inoculated treatments and plants did not nodulate in the uninoculated treatments. However, some nodules may have been lost due to the fragility of their attachment to the roots. All the nodules found were active (pink to brown). The authors believe that the poor nodulation of leucaena in this study may be attributable to:
a. The soil salinity or sodicity, or both, which might have inhibited the functioning of the symbiosis (Singleton and Bohlool, 1983). Unsuccessful inoculation with specific endophytes due to soil chemical factors was reported by Dommergues (1981). Adding gypsum to the soil at the rate recommended in Sudan (12.5 t/hectare) did not, apparently, increase nodulation.b. The fragile and narrow attachment of the nodule to the root (Halliday and Somasegaran, 1982).
c. Leucaena not being adapted to the area from which the soils were obtained. Halliday and Somasegaran (1982) reported that leucaena introduced to new areas is unlikely to encounter its specific rhizobial partner. This is substantiated by the convolution concepts of rhizobial strains and their hosts and by documented cases of leucaena failing to nodulate in habitats that are remote from its natural distribution. For example, Leucaena retusa, a native of Texas, has not nodulated in Hawaian soils. However, there are reports (Bohlool and Schmidt 1973; Vidor et al, 1981) of introduced strains both easily colonising areas and failing to do so. Nevertheless, as judged from the vigorous growth and healthy appearance of the plants in this study, leucaena is able to colonise these soils without inoculation. Effective nodulation is, however, important to reduce the period during which the seedling behaves as a non-legume, especially in soils that are deficient in N. e.g. those used in this investigation. Nodulation alone is not sufficient evidence of the productivity of leucaena (Halliday and Somasegaran, 1981). Work is currently underway in our laboratory to isolate salt-tolerant strains of Leucaena and other legumes.
d. Suppression of efficient nodulation might also be attributed to the N added as a starter dose. However, the authors believe that this is unlikely, since the soils used are inherently poor in nitrogen. An initial application of N is necessary for establishing plants in these marginal soils, because the plants may take up to So days to nodulate (Bushby, 1982). According to the author, even with inoculant levels at 200 times the normal rate the time to nodulate was slightly reduced. However, the poor nodulation on soils S1 and S5 (Table 1), which were non-saline, non-sodic and thus without any obvious deterents to plant infection, may support the theory that nodulation was suppressed by the N fertilizer.
Environmental factors
Temperature, solar radiation and rainfall affect the rate of growth of Leucaena and hence forage yield (Hegde, 1982). Different ranges of temperature have been reported to be conducive to the growth and development of Leucaena at different altitudes and latitudes, e.g. 0° C in winter to 41 C in summer (Hegde, 1982); 20° C to 32° C (Pathak and Patil, 1982) and 25° C to 30° C (Houming, 1982). These temperature ranges are similar to the temperature range reported in this work (Table 2). Moreover, Dijkman (1950) stated that optimum rate of growth occurs under direct sunlight. Pathak and Patil (1982) found that the above-ground growth of leucaena and nodulation initiation were best at 45% light intensity and 32.7° C, with fewer, larger nodules than at 100% light intensity and 35.4° C. In this work we did not measure the light intensity but temperature conditions were similar. Furthermore, few reports in the literature separate the effect of solar radiation from that of temperature.
In conclusion we think that the marginal soils studied can support leucaena growth. However, quick economic returns cannot be achieved without sensible management and use of cultivars that are better adapted to the chemical and the physical constraints inherent in these soils.
The authors are grateful to the NifTAL Project of the University of Hawaii, USA, for providing equipment, rhizobium strains, and leucaena seeds, and to the Arab Centre for Studies of Arid zones and Drylands (ACSAD), represented by the National Council For Research (NCR), for funding this work. We are also grateful to the Department of Biochemistry and Soil Science, Faculty of Agriculture, University of Khartoum, for providing research facilities.
Bohlool B B and Schmidt E L. 1973. Persistence and competition aspects of Rhizobium japonicum observed in soils by immunofluorescence microscopy. Soil Sci. Soc. Am., Proc. 37: 561-564. Brewbaker K L. 1978. Leucaena biomass productivity studies. Paper presented at the 20th. Annual Hawaii Forestry Conference, 22 September 1978, Kona, Hawaii, USA.
Brewbaker J L, Plucknett D L and Gonzalez V. 1972. Varietal variation and yield trials of Leucaena leucocephala (Koa Haole) in Hawaii. Hawaii Agric. Exp. qt., Res. Bull. No. 166.
Brewbaker J L, van den Beldt R and McDicken K. 1981. Nitrogen-fixing tree resources: Potential and limitations: In: Biological nitrogen fixation technology for tropical agriculture. CIAT, Cali, Colombia. pp. 413-425.
Bushby H V A. 1982. Rhizosphere population of Rhizobium strains and nodulation of Leucaena leucocephala. Aust. J. Exp. Agric. Anim. Husb. 22: 293-298.
Chapman H D and Pratt P F. 1961. Methods of analysis for soils, plants and waters. University of California, Berkeley, Division of Agricultural Sciences. 309 pp.
Chee W C and Devendra C. 1982. Research on Leucaena forage production in Malaysia. In: Leucaena research in the Asian-Pacific region. Singapore. pp. 55-60.
Dijkman M J. 1950. Leucaena - a promising soil erosion control plant. Econ. Bot. 4: 337-349.
Dommergues Y. 1981. Ensuring effective symbiosis in nitrogen-fixing trees. In: Biological nitrogen fixation technology for tropical agriculture. CIAT, Cali, Colombia. pp. 395-411.
El Mahi Y E and Mustafa M A. 1980. The effects of electrolyte concentration and sodium adsorption ratio on phosphate retention by soils Soil Sci. 130: 321-325.
Halliday J and Somasegaran P. 1982. nodulation nitrogen fixation and rhizobium strain affinities in the genus Leucaena. In: Leucaena research in the Asian-Pacific region. Singapore. pp. 27-32.
Hegde N. 1982. Leucaena forage management in India. In: Leucaena research in the Asian-Pacific region. Singapore. pp. 73-78.
Houming J. 1982. Introduction and trial planting of Leucaena in China. In: Leucaena research in the Asian-Pacific region. Singapore. pp. 123-126.
Jones R J. 1979. Value of Leucaena leucocephala as a feed for ruminants in the tropics. World Anim. Rev. 31:13-23.
Lavkulich L M. 1978. Pedology laboratory manual. Department of Soil Science, University of British Colombia, Vancouver, B.C. V6T IW5. 324 pp.
Munns D N and Franco A A. 1981. Soil constraints to legume production. In: Biological nitrogen fixation technology for tropical agriculture. CIAT, Cali, Colombia. pp. 133-152.
NAS (National Academy of Sciences). 1977. Leucaena: Promising forage and tree crop for the tropics. Washington D.C., USA.
NAS (National Academy of Sciences). 1979. Tropical legumes: Resources of the future. Washington D.C., USA. 331 pp.
Pathak P S and Patil B D. 1982. Leucaena research at the Indian Grassland and Fodder Research Institute (IGFRI). In: Leucaena research in the Asian-Pacific region. Singapore. pp. 83-88.
Singleton P W and Bohlool B B. 1983. Effect of salinity on the functional components of the soybean Rhizobium japonicum symbiosis. Crop Sci. 23: 815-818.
Thoma P E. 1983. Leucaena-Rhizobium compatability and nitrogen fixation. Desert Plants 5: 105-111.
USDA (United States Department of Agriculture). 1954. Diagnosis and improvement of saline and alkali soils. USDA Handbook 60.
Vidor C, Lavato P E and Miller R H. 1981. Survival of streptomycin-resistant mutants of Rhizobium phaseoli in non-sterile soil. In: Biological nitrogen fixation technology for tropical agriculture. CIAT, Cali, Colombia. pp. 201-205.
Walkley A and Black I A. 1934. An examination of the Degtjareff method for determining soil organic matter and proposed modification of the chromic acid titration method. Soil Sci. 37: 29-38.
![]()
![]()
![]()
{kind=link}