![]()
![]()
![]()
Kassu Yilala
Semi-Arid Food Grain Research and Development
Co-ordination Office, OAU/STRC
B.P. 1783, Ouagadougou
Burkina Faso
Abstract
Introduction
Mismatching of supply of nutrients and physiological states of animals in the Soudanian zone of Burkina Faso
Basic principles for the choice of source of nitrogen to utilise cereal by-products
Forage or dual-purpose legumes: Reliable source of N
Acknowledgements
References
The improvement in the efficiency with which ruminant animals transform crop residues into meat, milk and draught power will have important implications in the integration of animals in the strongly cereal crop-oriented production system of the Soudanian zone of Burkina Faso. There is a mismatching of supply of nutrients and the physiological states of animals in the production system. The abundant cellulosic energy could not be utilised effectively mainly due to the constraint of nitrogen (N) availability. The choice of source of N. non-protein or protein-N is crucial to the development of a feeding system that is sound biologically and economically.
The biological aspects of utilisation of crop residues with the aid of N supplements at ruminal and tissue levels are discussed to justify the choice of source of N that could fit into the objective conditions of the production system. The production system could be responsive to changes if the intervention is compatible with the long term productivity of cereal crop production through improved soil fertility and structure.
Amongst the sources of N emphasis was put on the use of forage or dual-purpose legumes for they could serve as the key link between animal and crop production systems, the latter through increased soil N and organic matter (OM) status. The patterns of in vitro degradation of OM and N of the forage legumes and sorghum and millet stovers were assessed and briefly discussed. It was shown that the forage legumes in addition to correcting the deficiency of rumen degradable nitrogen (RDN), if supplemented to cereal stovers, can also contribute a substantial amount of rumen-degradable organic matter (RDOM). However, due to the rapid degradation of N the available RDOM may not be able to match to the former. Suggestions are given to increase the supply of protein to the small intestine.
Mixed crop-animal production occurs virtually throughout the farming sectors of Burkina Faso. In the Soudanian zone cereal crop sorghum and millet, production is the foundation of agriculture and animal production the vital economic unit of the system.
Declining crop yield/unit area accompanied by progressively increasing requirement for food is forcing farmers to cultivate more land at the expense of grazing pasture and browses. The other aspect of this process is the increase in the production of cereal crop residues. There is already a heavy reliance on the use of the byproducts as feed during the long dry season, manifested by the large number of ruminants concentrated in the zone, while moving from the Sahel to the Guinean zones, immediately after harvest of cereals.
It is well documented that the utilisation of the energy components of such materials by ruminant animals is highly dependent on the efficiency of the fermentative activity of the microbes in the rumen. For optimum or maximum fermentation on a given diet a certain level of ammonia (NH3) concentration in the rumen is required. Otherwise feed intake may be reduced if NH3 concentration is limiting the rate of fermentation (Mehrez et al, 1977). Animals fed on such materials as their sole diet show low dry-matter intakes and decline in liveweight. There is ample evidence that with the inclusion of a source of nitrogen, NPN or protein-N, positive responses in intake and liveweight gain can be realised. In the Soudanian zone of Burkina Faso, as in other semi-arid regions, nitrogen is the most limiting nutrient for ruminant animal production. Therefore, the incorporation of this nutrient in a feeding system based on low-nitrogen fibrous diet is of paramount importance. The attempt in supplementing with nitrogen should be to maximise the utilisation of the residues through the fermentative activities in the rumen and to realise as large responses as possible at the tissue level of the host animal at a given physiological state.
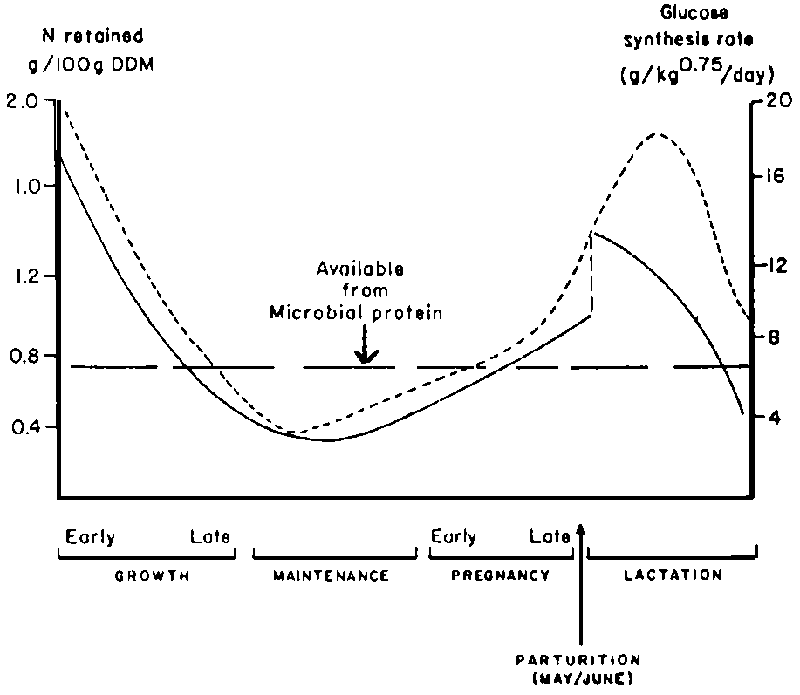
Improvement in animal productivity requires improved nutrition throughout the year with a guarantee for adequate supply, quantitatively and qualitatively, in the latter part of pregnancy, lactation and early growth periods. Orskov (1970) and Kempton et al (1977) have demonstrated animal needs of amino acids and glucose according to physiological states (Figure 1). Such concepts could serve as guidelines to the budgetting of available nutrients according to nutritional needs of the animal. The demand for amino acids and energy (in the form of glucose) are high during growth, late pregnancy and lactation. In Burkina Faso the main calving, lambing or kidding seasons are just before or during the rains (Hay/June), but some births also take place in October/November just after the rains, when cereal residues become abundant. The high demand for nutrients during the last stage of pregnancy in the former case, or during lactation in the latter, cannot be met adequately from the natural pasture and cereal residues; the nutritional status of the animals, lactation performance and growth of the young are reduced, and are reflected by high rates of pre-and post-weaning mortality of calves and lambs in the semi-arid regions, depending on the month of the year (ILCA, 1982).
|
* Physiological state |
Major sources of nutrients |
|
|
Growth |
||
|
|
- Preweaning |
- Might benefit from improved pasture between July and September through dam's milk during early pre-weaning period. |
|
|
- Postweaning |
- Exposed to mature natural pasture and crop residues. |
|
Maintenance |
- If between October and June all animals are exposed to standing mature dry pasture and cereal residues. |
|
|
Pregnancy |
- Last 10 to 8 weeks before parturition exposed to extremely dry mature natural pasture and cereal residues. Period of severe deficit of nutrients. |
|
|
Lactation |
- First 90 - 120 days lactating animals exposed to improved grazing conditions to be followed by cereal residues. |
|
Sources: Orskov (1970); Kempton et al, (1977).
The nutrient yield from the natural pasture increases between July and October, but the short rainy season accompanied by high evapotranspiration and high temperature results in rapid decline in the contents of nitrogen (Figure 2) and degradation of organic matter (OM) in the rumen (Table 1).
When available, farmers use groundnut and cowpea haulms, the latter grown in association with millet, to supplement cereal residues to selected animals. However, the quantities produced on the farm are not adequate to satisfy the requirements during periods of high nutrient demand.
Table 1. In vitro dry-matter digestibility (DMD) and in-sacco disappearance of DM of fallow natural pasture cut at different stages of growth, Soudanian zone, Burkina Faso.
|
Date of cutting |
In vitro |
Disappearance of DM in the rumen |
|
|
DMD |
Incubation |
(h) |
|
|
(%) |
0 |
48 |
|
|
21-08-85 |
63.4 |
26.7 |
71.4 |
|
11-09-85 |
61.4 |
22.7 |
65.5 |
|
20-02-85 |
59.1 |
19.6 |
55.6 |
|
23-10-85 |
57.1 |
15.7 |
58.7 |
|
14-11-85 |
54.1 |
13.7 |
57.9 |
|
LSD (P=0.05) |
2.3 |
3.3 |
4.1 |
Source: Yilala (1986b).
The feeding system that needs to be established under a particular condition should be based on the universal biological laws of nutrient utilisation by ruminants and also fit into the economic realities of the production system. Some aspects of the basic biological principles involved in the digestion in the rumen and utilisation by the host animal of cereal residues supplemented with NPN or protein-N, and the implications of these principles on the choice of sources of nitrogen to fit into the objective conditions of the production system in Burkina Faso will be discussed briefly.
1. Biological
1.1 Effects of source of nitrogen on microbial digestion and voluntary intake
Several studies have shown improvements in the intake of low-nitrogen fibrous diets due to urea supplementation (Egan, 1965; Egan and Moir, 1965; Kempton and Leng, 1979; Sriskandrajah et al, 1982), attributed to increased rate of digestion of cellulose in the rumen (Egan, 1965; Egan and Moir, 1965), probably resulting in increased rate of passage of feed, thereby possibly alleviating a physical limitation to intake (Blaxter et al, 1961).
The degradation of urea is know NH3 to be too rapid (Johnson, 1976), indicating that the efficiency of utilisation could be low when used with cellulosic diets, which yield energy too slowly for the efficient capture of NH3 by rumen micro-organisms (Meggison et al, 1979). To realise a more effective utilisation of urea its N may need to be released slowly (Orskov, 1982). The synchronisation of rate of degradation of N and carbohydrate components in the rumen is important for the synthesis of microbial protein (Meggison et al, 1979; Satter and Roffler, 1981). Microbial protein synthesised in the rumen is the major source of N to the host animal accounting for 60-85% of the total amino acids entering the small intestine (Orskov, 1982). The pattern of degradation, therefore, influences the choice of N source for efficient utilisation of cereal residues. A rapid degradation of N not matched to the release of OM from the carbohydrate could lead to a high absorption of NH3 from the rumen (Meggison et al, 1979). The availability of suitable carbon skeletons and ATP from plant material is a requirement for the NH3 released from dietary urea to be used for microbial protein synthesis (Czerkawski, 1986).
Figure 2. Pattern of change in the proportion of N and NDF in the DM of natural pasture on fallow land cut at different intervals from August 21 to November 14, 1985 (Soudanian zone, Burkina Faso).

Source: Yilala (1986b).
The NH3 not captured in the rumen is absorbed and converted into urea, partly to be transferred to the rumen and influence the digestion rate of cellulose or be lost in the urine depending on the concentration of NH3 in the rumen (Kennedy and Milligan, 1978). The synthesis of urea in the liver requires expenditure of energy, each mole of urea requiring 4 moles of ATP (Martin and Blaxter, 1965).
Compared with urea, protein-N sources could provide better conditions for the digestion of cereal residues by the microbes in the rumen, for the following reasons:
a. Although NH3 is the major source of N for microbial growth, some species are able to utilise or require the presence of peptides and/or amino acids for growth (Cotta and Russet, 1982; Czerkawski, 1986; Harrison and McAllan, 1980; Maeng and Baldwin, 1976; Pisulewski et al, 1981).b. Due to the variation in degradability, many protein-N sources release NH3 at slower rate than urea-N, more closely coinciding with release of energy from the cellulosic component and thus enhancing microbial production (Miller, 1982). This in turn will also stimulate increased rate of cellulose digestion and voluntary intake.
c. Protein-N can also serve as a good source of sulphur which usually is lacking in Lou-protein roughage diets (Miller, 1982). The supplementation of roughage diets with sulphur shoved increases in the digestion of cellulose (Gill et al, 1973; Gulati et al, 1985; Spears et al, 1976) for it is an essential element for ruminal microbes (ARC, 1980; Spears et al, 1976).
1.2 Effects of source of nitrogen on the response at the tissue level
The animal is mainly dependent on volatile fatty acids (VFA) as its source of energy. However, glucose is the major energy component required for various metabolic processes in the tissue. Because ruminants are unable to absorb sufficient glucose from the digestive tract to meet their needs (Lindsay, 1980), the availability of glycogenic materials such as propionic acid and amino acids is essential (Kempton et al, 1977). The fermentation of poor quality roughages such as cereal residues normally leads to the production of high proportion of acetic acid (Thomas and Rook, 1981) which is non glycogenic (Hovell and Greenhalgh, 1978). Thus cereal residues cannot supply adequate energy for the retention of N at the tissue level.
Under such conditions amino acids of microbial origin or catabolised from the tissues might through gluconeogenesis supply glucose for protein synthesis (MaCrae and Reeds, 1980), to utilise acetate for lipogenesis (Hovell and Greenhalgh, 1978), and for urea synthesis in the liver (Martin and Blaxter, 1965), indicating that the metabolism of absorbed amino acids contributes also to energy metabolism. The transformation of amino acids into glucose obviously reduces the efficiency with which the former are utilised for tissue synthesis (MaCrae and Reeds, 1980).
With no contribution from the urea to the energy pool the low N balance values for the urea-N, as compared to protein-N supplemented diets (Egan, 1965; Krzeminski, 1985) is not surprising. The large increase in liveweight gain (Saadullah et al, 1983; Kempton and Leng, 1979) or reduction in plasma urea and increase in N retention (Krzeminski, 1985) due to the inclusion of poorly degraded protein-N sources in straw diets containing urea supplements might, at least partially, be associated with the supply of glucose contributed by amino acids absorbed from the small intestine. Besides this, the improvements in N status of the animals were associated with increases in the intake of the fibrous diets (Egan, 1965; Kempton and Leng, 1979).
2. Objective conditions of production system
From the above it is clear that the form of N supplementation of cereal by-products should be considered if effective intervention is desired at small-scale farm level. Quite often it is urea-N, assumed to be easily available and cheap per unit of N. that is considered to improve the utilisation of cereal residues, either for direct supplementation or treatment of the byproduct. Treating residues with urea has been observed to enrich the N content and result in positive effects on digestibility, intake (Dolberg et al, 1981) and liveweight changes better than the direct application prior to feeding (Orskov, 1981).
As a result of these and simplicity in the method of treatment it is believed to be applicable under small-scale farm conditions,. However, under the prevailing conditions in Burkina Faso this does not appear to hold true, at Least for the time being, for two main reasons:
a. Treatment with urea requires large quantity of water (1 litre of water/kg of straw; Dolberg et al, 1981); the principal constraint of the production system in the Soudanian zone.b. NH3 loss could be up to 60% when straw is treated with urea (Sundstol, 1981) and such losses cannot be justified under conditions of limited N supply.
Protein-N sources such as oilseed cakes and those of animal origin are produced in limited quantity and seem to be beyond the economic reach of the farmers. The self-reliance of the farmer in the source of N. therefore, is a prerequisite to improve the efficiency of utilisation of the cereal residues and productivity of animals.
Forage legumes have enormous potential under such conditions. Since N is also the most limiting nutrient in the soils of Burkina Faso, the production of forage, or dual-purpose legumes could be compatible with the long-term productivity of the production system comprising soil fertility through N2 fixation. The increase in cereal grain yield could be accompanied with an increase in residue yield. Forage legumes could, therefore, serve as the key link for effective integration of animals into the strongly crop-oriented production system.
Intercropping of cereals with legumes such as cowpea is a common practice in the Soudanian zone of Burkina Faso. Groundnut is grown as a sole crop on small plots of land. The residues of these crops serve as sources of N to supplement the diets of selected animals. However, the quantity produced on the farm is low, compared to the DM of cereal residues, and cannot offset the deficiency of N (Table 2).
Studies with forage and dual-purpose legumes on fallow land showed the existence of great potential for the replacement of poor fallow pasture with legumes which could be conserved as hay for feeding as required. The legumes used were: Dolichos lablab, Stylosanthes hamata, Macroptilium atropurpureum, Vigna unguiculata (local and improved varieties) and Phaseolus aureus.
In general legumes are known to be less dependent on soil N for they obtain their N through biological fixation. As a result, the N content of all the legumes during the period of observation has always been above the critical level, for animal use, at all stages of growth compared to the natural pasture (Figure 3b). The critical level of N below which voluntary intake of DM is depressed is 1.12% (Whiteman, 1980). As expected there was a decline in the contents of N and increase in the contents of the cell wall components (Figure 3a). Differences were also noted in the solubility of DM due to species differences and stage of growth (Table 3).
Table 2. Estimated yields of dry matter (DM), metabolisable energy (ME) and rumen-degradable nitrogen (RDN) of cereal residues, legume haulms and legume hays.
|
|
Cereal residue |
Legume haulms |
Legume hay |
||||
|
Sorghum |
Millet |
Cowpea |
Groundnut |
Cowpea
|
D. lablab |
||
|
Sole |
Intercrop |
|
|||||
|
DM yield (kg/ha) 1 |
4209 |
3125 |
373 |
101 |
1497 |
3010 |
3010 |
|
ME yield (kg/ha) 2 |
31568 |
16875 |
2611 |
707 |
11527 |
26187 |
34595 |
|
RDN yield (kg/ha) 3 |
20 |
10 |
4.4 |
1.0 |
16.9 |
115 |
148 |
|
RDN required to match ME yield (kg/ha) |
40 |
21 |
3.3 |
0.9 |
14.4 |
33 |
43 |
|
RDN balance (kg/ha) 4 |
-20 |
-11 |
1.1 |
0.1 |
2.5 |
82 |
105 |
1. Values are averages for different villages in the Soudanian zone, Burkina Faso.2. Calculated using the in vitro digestible organic matter values.
3. Refers to the 24 h in vitro disappearance.
4. Assuming complete synchronization of release of energy and nitrogen in the rumen.
Table 3. Disappearance of dry matter (%) of forage legumes, grown on fallow land, from nylon bags in the rumen of cattle for incubation period of 0 and 48 h.
|
Species |
Age of cutting (days) |
|||||||
|
35 |
49 |
77 |
84 |
|||||
|
Incubation (h) |
Incubation (h) |
Incubation (h) |
Incubation (h) |
|||||
|
0 |
48 |
0 |
48 |
0 |
48 |
0 |
48 |
|
|
D. lablab (cv Highworth) |
52.9 |
84.9 |
55.9 |
89.6 |
40.3 |
80.1 |
40.0 |
83.0 |
|
V. unguiculata (cv KN-1) |
46.5 |
88.8 |
41.6 |
79.9 |
43.1 |
80.8 |
41.1 |
85.7 |
|
P. aureus |
45.1 |
82.9 |
46.2 |
86.3 |
40.1 |
86.0 |
42.5 |
85.9 |
|
S. hamata |
- |
- |
36.4 |
81.1 |
34.8 |
75.9 |
32.5 |
75.4 |
|
M. atropurpureum |
37.3 |
72.3 |
36.4 |
64.5 |
34.2 |
62.2 |
32.3 |
58.8 |
Source: Yilala (1986a).
Figure 3. Patterns of change in nitrogen (N) and neutral detergent fibre (NDF) contents of forage legumes compared to fallow land natural pasture as growth advances:
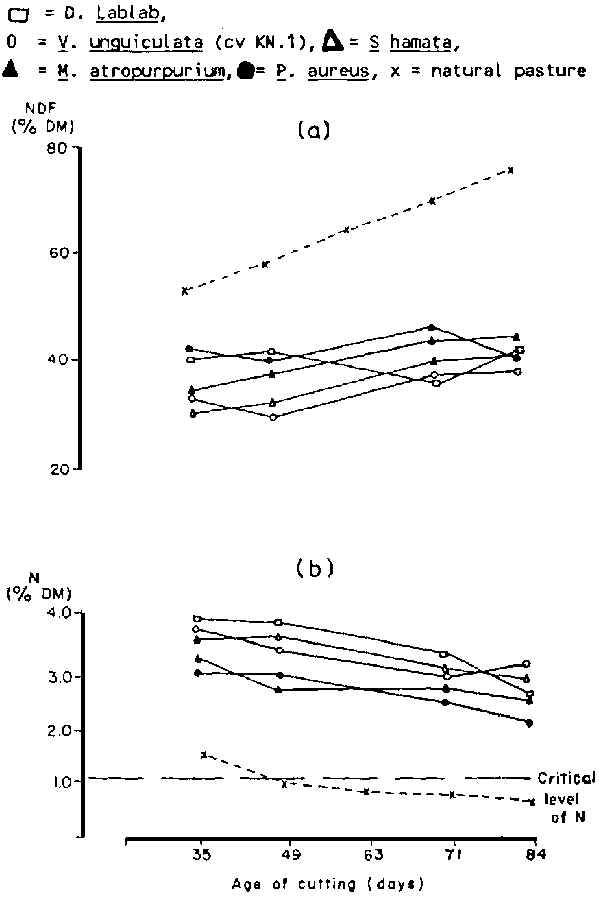
Considering the pattern of change with time in DM yield, N. NDF and lignin contents, digestibility and solubility of DM, the age of cutting that will fit into the conditions of the production system for conservation as hay was identified. Under the conditions of the Soudanian zone harvesting between 75 and 80 days of growth is suggested for all the species tested. This coincides with the period at which the labour requirement for cereal crop production is lowest, thus allowing the conservation of forage legumes at the preferred stage of growth.
1. In vitro degradability characteristics of forage resumes cut at 75 days of growth
The presence of N in the diet does not always guarantee the availability to the target microbes in the rumen. As noted earlier it is influenced by the pattern of degradability of the protein. The amount of protein that reaches the small intestine, besides degradability, is also influenced by the outflow rate from the rumen (Orskov and Robinson, 1981).
The pattern of N and OM in vitro disappearance in this study might indicate the characteristics of the legumes in their degradation in the rumen. Such information might allow effective screening of legumes to identify those whose N might be released in synchronisation with the release of OM from the cereal residues in the rumen (Table 4).
The results, however, need to be treated as indicative of the pattern of degradation and not to be equated with in vivo conditions. In addition to correcting the deficiency of rumen-degraded nitrogen (RDN) to utilise the rumen-degraded organic matter (RDOM) of the cereal crop residues, the forage legumes could also contribute a substantial amount of OM digested in the rumen. However, the N components of these legumes were rapidly degraded with a disappearance of not less than 80% within 6 hours of incubation.
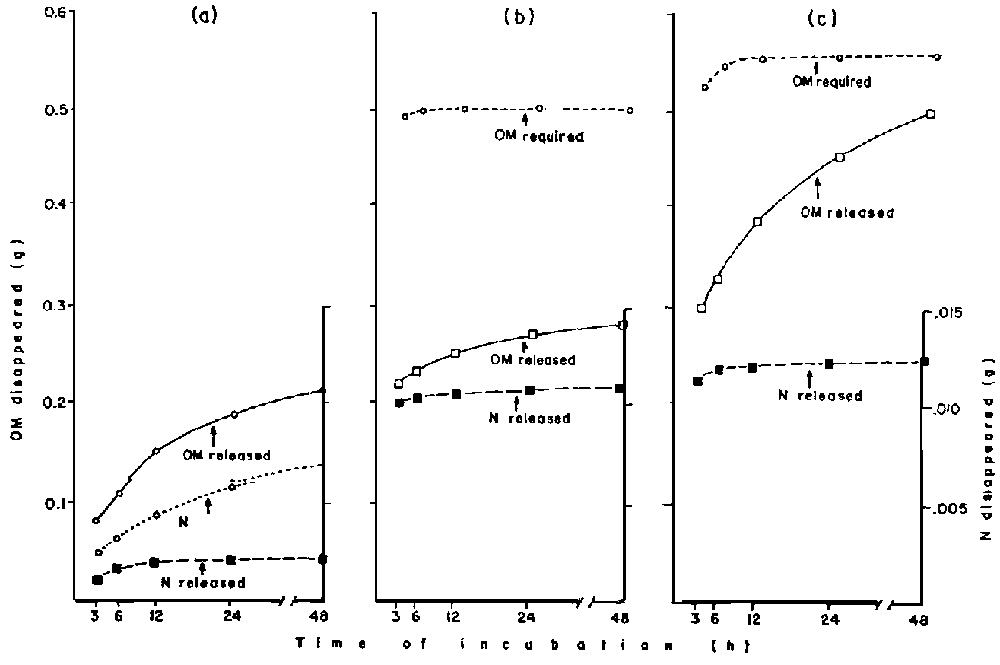
ARC (1980) estimated the ratio of RDN: ROOM required in the rumen to be 1:33.3. Based on this the OM and N releases of cowpea and millet stover were assessed for illustration purposes (Figure 4). In Figure 4 (a) it is noted that the OM released from the millet stover may not be utilised completely because of the inadequacy of N contained in it. Figure 4 (b) might illustrate a situation where cowpea N. is degraded rapidly and may not get adequate amount of OM released to match it, and the excess N could be lost in the urine. It is possible that the combination of the OM from cowpea and millet stover too may not provide adequate OM to match the N degraded (Figure 4(c)).
In places where conditions permit, forage proteins of high degradability could be treated with chemicals such as formaldehyde to increase their passage to the small intestine to be absorbed as amino acids. Whereas, in places where such possibilities do not exist, for example in Burkina Faso, other alternatives that could fit into the system need to be sought.
2. Suggestions to improve the utilisation of forage protein-N
There are reports (Kempton and Leng, 1979; Macrae and Ulyatt, 1980; Orskov and Robinson, 1981) which indicate protein deposition in animals to be positively related to the amount of amino acids absorbed from the small intestine. Such responses are higher in those animals fed rations with lower rather than higher concentration of degradable N (Cummins et al, 1980; Yilala and Bryant, 1985).
In order to improve the supply of protein to the small intestine when forage legumes are to be included in crop residue basal diets the following suggestions may help:
a. Increase the level of readily fermentable carbohydrate to capture the degraded N for microbial synthesis to be absorbed as amino acids of microbial origin. Probably growing the legumes in association with grass species with known water soluble carbohydrates and adopted to the climatic conditions of the semi-arid region might help. Chopping the straw might increase the overall digestible organic matter in the rumen.b. Combine the particular forage legume with other protein-N sources of less degradability so that the absorption of amino acids from the dietary source could be increased at the small intestine. Since protein-N of animal origin and oilseed cakes are scarce, legume forages and browses containing tannins could be introduced. Legumes containing a certain level of condensed tannins are known to increase the quantity of amino acids absorbed in the small intestine (Barry and Manley, 1983; Waghorn et al, 1987), although condensed tannins can also depress digestion in the rumen of the carbohydrate components (Barry and Manley, 1983).
In conclusion, the objective realities in the Soudanian zone of Burkina Faso favour the creation of conditions that will allow the effective integration of ruminant animals into the crop production system through recycling of resources. Forage legumes possess enormous potential as sources of N and could serve as the key link in the integration of crop and animal production systems (Table 5). So far farmers have not given due importance to the incorporation of forage crops in the system. Possibilities that could increase the production of forage proteins do exist:
a. Replacement of poor fallow pasture with forage legume/cereal rotation.b. Intercropping cereals with forage or dual-purpose legumes.
c. Production of browses along contour bunds or alley cropped with cereals.
The author is grateful to Mr. S. Zoundi and technical assistants for providing all the necessary help for the trials. The financial assistance of SAFGRAD (Semi-Arid Food Grain Research and Development) and the provision of facilities by INERA (Institut national pour l'etude et la recherche agronomique) are gratefully acknowledged.
ARC (Agricultural Research Council). 1980. The nutrient requirements of ruminant livestock. Commonwealth Agricultural Bureaux, Farnham Royal, UK.
Barry, T.N. and Manley, T.R. 1983. Quantitative digestion studies in sheep fed vegetative Lotus pedunculatus with different contents of condensed tannin. Proc. Nutr. Soc. New Zealand 8: 140-141.
Blaxter, K.L.; Wainman, F.D. and Wilson, R.S. 1961. The regulation of food intake by sheep. Anim. Prod. 3: 51-61.
Cotta, M.A. and Russell, J.B. 1982. Effect of peptides and amino acids on efficiency of rumen bacterial protein synthesis in continuous culture. J. Dairy Sci. 65: 226-234.
Cummins, K.A., Nocek, J.E. and Polan, C.E. 1980. Growth and nitrogen balance of calves fed rations of varying degradability and physical form. J. Dairy Sci. 65: 773-783.
Czerkawski, J.W. 1986. An introduction to rumen studies. Pergamon Press, Oxford.
Dolberg, F.; Saadullah, M. and Haque, R. 1981. Straw treatment in a village in Noakhali district, Bangladesh. In: M.G. Jackson, F. Dolberg, C.H. Davis, M. Haque and M. Saadullah (eds), Maximum livestock production from minimum land. Proc. Seminar held in Bangladesh, Feb. 1981. Bangladesh Agricultural University, Mymensingh, Bangladesh. pp. 205-224.
Egan, A.R. 1965. Nutritional status and intake regulation in sheep. III. The relationship between improvement in nitrogen status and increase in voluntary intake of low-protein roughages by sheep. Aust. J. Agr. Res. 16: 463-472.
Egan, A.R. and Moir, R.J. 1965. Nutritional status and intake regulation in sheep. I. Effects of duodenally infused single doses of casein, urea and propionate upon voluntary intake of a low-protein roughage by sheep. Aust. J. Agr. Res. 16: 437-449.
Fall, A.; Diop, M.; Sanford, J.; Wissocq, Y.J.; Durkin, J. and Trail, J.C.M. 1982. Evaluation of the productivities of Djallonke sheep and N'Dama cattle at the Centre de Recherches Zootechniques, Kolda, Senegal. ILCA Research Report, No. 3, ILCA, Addis Ababa, Ethiopia.
Gill, L.A.; Shirley, R.L. and Moore, J.E. 1973. Effect of methionine hydroxy analog on bacterial protein synthesis from urea and glucose, starch or cellulose by rumen microbes in vitro. J. Anim. Sci. 37: 159-163.
Gulati, S.K.; Ashes, J.R.; Gordon, G.L.R. and Phillips, M.W. 1985. Possible contribution of rumen fungi to fibre digestion in sheep. Proc. Nutr. Soc. Austr. 10: 96-106.
Harrison, D.G. and McAllan, A.B. 1980. Factors affecting microbial growth yields in the reticulo-rumen. In: Y. Ruckebusch and P. Thivend (eds), Digestive physiology and metabolism in ruminants. MTP Press Ltd., Lancaster. pp. 205-226.
Hovell, F.D. and Greenhalgh, J.F.D. 1978. The utilization of diets containing acetate, propionate orbutyrate salts by growing lambs. Br. J. Nutr. 40: 171-183.
Johnson, R.R. 1976. Influence of carbohydrate solubility on non protein nitrogen utilization in the ruminant. J. Anim. Sci. 43:184-191.
Kempton, T.J. and Leng, R.A. 1979. Protein nutrition of growing lambs. 2. Effect on nitrogen digestion of supplementing a low-protein cellulosic diet with either urea, casein or formaldehyde-treated casein. Br. J. Nutr. 42: 303-315.
Kempton, T.J.; Nolan, J.V. and Leng, R.A. 1977. Principles of the use of non-protein nitrogen and by-pass proteins in diets of ruminants. World Animal Review (FAO) 22: 2-10.
Kennedy, P.M. and Milligan, L.P. 1980. The degradation and utilization of endogenous urea in the gastrointestinal tract of ruminants: A review. Can. J. Anim. Sci. 60: 205-221.
Krzeminski, J.; Kulasek, G.; Motyl, T.; Jaworska, K.; Leontouricz, H. and Kozlowski, B. 1985. Protein supplement resistant against ruminal degradation as a factor improving utilization of urea in ruminant feeding. Archiv fur Tierernahrung. 35: 401-409. (Abstract (En): SDI Service-ILCA).
Lindsay, D.B. 1980. Amino acids as energy sources. Proc. Nutr. Soc. 39: 53-59.
MacRae, J.C. and Reeds, P.J. 1980. Prediction of protein deposition in ruminants. In: P.J. Buttery and D.B. Lindsay (eds), Protein deposition in animals. Butterworths, London. pp. 225-250.
Maeng, W.J. and Baldwin, R.L. 1976. Factors influencing rumen microbial growth rates and yields. J. Dairy Sci. 59: 643-647.
Martin, A.K. and Blaxter, K.L. 1965. The energy cost of urea synthesis in sheep. European Association for Animal Production Publication No. 11, Academic Press Inc., London, UK. pp. 83-97.
Meggison, P.A.; McMeniman, N.P. and Armstrong, D.G. 1979. Efficiency of utilization of non-protein nitrogen in cattle. Proc. Nutr. Soc. 38: 147A.
Mehrez, A.Z.; Orskov, E.R. and McDonald, I. 1977. Rates of rumen fermentation in relation to ammonia concentration. Br. J. Nutr. 38: 437-443.
Miller, E.L. 1982. The nitrogen needs of ruminants. In: D.J. Thomson, D.E. Beever and R.G. Gunn (eds), Forage protein in ruminant animal production. BSAP Occasional Publication (UK), No. 6, British Society of Animal Production, Surrey, UK. pp. 79-87.
Orskov, E.R. 1970. Nitrogen utilization by the young ruminant. In: H. Swan and D. Lewis (eds), Proceedings of the 4th Nutrition Conference for Feeds Manufacturers, University of Nottingham, London.
Orskov, E.R. 1981. Nutritional evaluation of poor quality roughages. In: M.G. Jackson, F. Dolberg, C.H. Davis, M. Haque and M. Saadullah (eds), Maximum livestock production from minimum land. Proceedings of a Seminar held in Bangladesh, Feb. 1981. Bangladesh Agricultural University, Mymensingh, Bangladesh. pp. 70-87.
Orskov, E.R. 1982. Protein nutrition in ruminants. Academic Press, London.
Orskov, E.R. and Robinson, J.J. 1981. The application of modern concepts of ruminant protein nutrition to sheep production systems. Livestock Prod. Sci. 8: 339-350.
Pisulewaki, P.M.; Okorie, Q.L.; Buttery, P.J.; Haresign, W. and Lewis, D. 1981. Ammonia concentration and protein synthesis in the rumen. J. Sci. Food. Agric. 32: 759-766.
Saadullah, M.; Haque, M.; Thomson, K.V.; Moller, P.D. and Sorensen, A.N. 1983. Response of calves fed ammonia treated straw to increasing levels of fishmeal. In: Maximum livestock production from minimum land. Proc. 4th seminar held in Bangladesh, May 1983. Bangladesh Agricultural University, Mymensingh, Bangladesh.
Satter, L.D. and Roffler, R.E. 1981. Influence of nitrogen end carbohydrate inputs on rumen fermentation. In: W. Haresign and D.J.A. Cola (eds), Recent developments in ruminant nutrition. Butterworths, London. pp. 115-139.
Spears, J.W.; Ely, D.G.; Bush, L.P. and Buckner, R.C. 1976. Sulphur supplementation and in vitro digestion of forage cellulose by rumen microorganisms. J. Anim. Sci. 43: 513-517.
Sriskandrajah, N.; Kellaway, R.C. and Leinbolz, J. 1982. Utilization of low quality roughages: Effects of supplementing with casein treated or untreated with formaldehyde on digests flows, intake and growth rate of cattle eating wheat straw. Br. J. Nutr. 47: 553-563.
Sundstol, F. 1981. Results of some experiments on ammonia-treated straw. In: M.G. Jackson, F. Dolberg, C.H. Davis, M. Haque and M. Saadullah (eds), Maximum livestock production from minimum land. Proceedings of a Seminar held in Bangladesh, Feb. 1981. Bangladesh Agricultural University, Mymensingh, Bangladesh. pp. 97-102.
Thomas, P.C. and Rook, J.A.F. 1981. Manipulation of rumen fermentation. In: W. Haresign and D.J.A. Cole (eds), Recent developments in ruminant nutrition. Butterworths, London.
Waghorn, G.C.; Ulyatt, M.J.; John, A. and Fischer, M.T. 1987. The effect of condensed tannins on the site of digestion of amino acids and other nutrients in sheep fed on Lotus corniculatus: Br. J. Nutr. 57: 115-126.
Whiteman, P.C. 1980. Tropical pasture science. Oxford University Press, Oxford.
Yilala, K. 1986a. Preliminary investigation on the establishment of forage legumes on fallow land and assessment of their nutritive value at different stages of growth. In: Farming Systems Research Report (SAFGRAD/INERA). Ed. Co-ordination Office OAU/STRC/SAFGRAD, Ouagadougou, Burkina Faso.
Yilala, K. 1986b. Assessment of the nutritive value of natural pasture on fallow tend at different stages of growth. In: Farming Systems Research Report (SAFGRAD/INERA). Ed. Co-ordination Office OAU/STRC/SAFGRAD, Ouagadougou, Burkina Faso.
Yilala, K. and Bryant, M.J. 1985. The effects upon the intake and performance of store lambs of supplementing grass silage with barley, fishmeal and rapeseed meat. Anim. Prod. 40: 111-121.
![]()
![]()
![]()
{kind=link}
{kind=link}