![]()
![]()
![]()
J.J. Hakiza, J.R. Lazier and A.R. Sayers
ILCA, P.O. Box 5689, Addis Ababa, Ethiopia
Introduction
Methodology
Results
Discussion
Acknowledgements
References
Appendix 1. Collections of stylosanthes fruticosa being evaluated in Ethiopia.
Abstract
A study was carried out on accessions undergoing agronomic evaluation and seed multiplication. A total of 93 accessions grown as spaced plants were involved and 18 simple morphological and agronomic data recorded. Principal component analysis, cluster analysis and K-means clustering were used on the data.
It was shown that although there existed a high level of agronomic heterogeneity and genetic variability among the accessions, more collections were inevitable to widen the genetic base of S. fruticosa and cover areas particularly in Africa where this species is prevalent. It was also realised that a more systematic study of these accessions for fairly long periods might give a better understanding of these materials. Characteristics of various groupings are discussed.
The importance of a germplasm collection to those utilising it is very dependent on the availability of accurate descriptions of the accessions and on the taxonomic identification of the germplasm (Engels, 1986). Where there is a substantial number of accessions of one species or genus within the collection, efforts should be made to classify these into like groups. The objective of this classification is to allow the reduction of the number of accessions that have to be grown in the next phase.
The genus Stylosanthes comprises 30 to 40 species (Burt et al., 1983; Stace and Edye, 1984), many of which are either agronomically unattractive or are of rare occurrence. By the beginning of 1984 the world collection of Stylosanthes spp. and S. fruticosa were as shown in Table 1. Stylosanthes species are adapted to quite different ecological situations and range from annuals to perennials. They also have different agronomic forms. This exemplifies the high range of variability within the genus.
S. fruticosa is regarded as an excellent forage legume in some parts of Africa. Its deep and strong tap root system makes it resistant to grazing. The prostrate forms have some value in protecting the soil against erosion. S. fruticosa is a species which has received less attention than its South American allies S. guianensis and S. humilis, particularly in Australia. It must be presumed to have considerable potential as a forage legume in regions with a single dry season. It is distributed in many countries in Africa as seen in Figure 1.
The latitude and longitude ranges within the current S. fruticosa collections were respectively between 12°00'N and 13°24'N and 02°19'E and 09°03'E for Niger, and for Ethiopia between 04°54'N and 07°11'N and 37°22'E and 39°30'E. Williams et al (1984), Lazier (1984), Skerman (1977), Thulin (1983) and IBPGR (1984) show that S. fruticosa is distributed in at least 23 African countries.
Table 1. World collection of Stylosanthes fruticosa and Stylosanthes spp.
|
Institution1/ |
S. fruticosa |
All Stylosanthes spp. |
|
CIAT |
10 |
2241 |
|
CSIRO |
88 |
1451 |
|
EMBRAPA |
0 |
1296 |
|
EPAMIG |
1 |
626 |
|
University of Florida |
23 |
838 |
|
ILCA* |
164 |
858 |
1/ CIAT = Centro International de Agricultura Tropical, Colombia.
CSIRO = Commonwealth Scientific and Industrial Research Organization, Australia.
EMBRAPA = Empresa Brasileira de Pesquisa Agropecuaria, Brazil.
EPAMIG = Empresa de Pesquisa Agropecuaria de Minas Gerais, Brazil.
ILCA = International Livestock Centre for Africa, Ethiopia.*The figures for ILCA are as of 31/12/86, and these were not included in the original table.
Source: Stace and Edye (1984).
Despite this wide distribution of S. fruticosa, especially in Africa, the current collection at the International Livestock Centre for Africa (ILCA) is mainly from Ethiopia and Niger. Two accessions were acquired from the Commonwealth Scientific and Industrial Research Organization (CSIRO), Australia. In order to search for materials of agronomic potential, it is essential to collect sufficiently variable germplasm from many diverse environments and evaluate their forage potential.
Some morphological and agronomic characters were studied for both identification purposes and assessment of agronomic value.
Of the 164 accessions of S. fruticosa in the genebank at ILCA, 102 accessions were planted for agronomic evaluation and seed multiplication at Soddo in Sidamo, Ethiopia during April, 1985. The geographical and climatic description of Soddo and the planting procedure were as given in the Zornia paper in these proceedings.
These S. fruticosa accessions had been collected mainly from Ethiopia and Niger (Appendix 1). Those from Ethiopia were predominantly from Wolayta in Sidamo. In order to assess the existence of variations between the accessions that were undergoing agronomic evaluation, a study was superimposed on this trial. Observations were made when the plants were about 18 months old at the end of the rainy season. Eighteen characters (morphological and agronomic) on 93 accessions were selected based on the practical and field considerations at that particular time. These are given in Table 2 with their character state codes.
Figure 1. Distribution of Stylosanthes fruticosa in Africa.
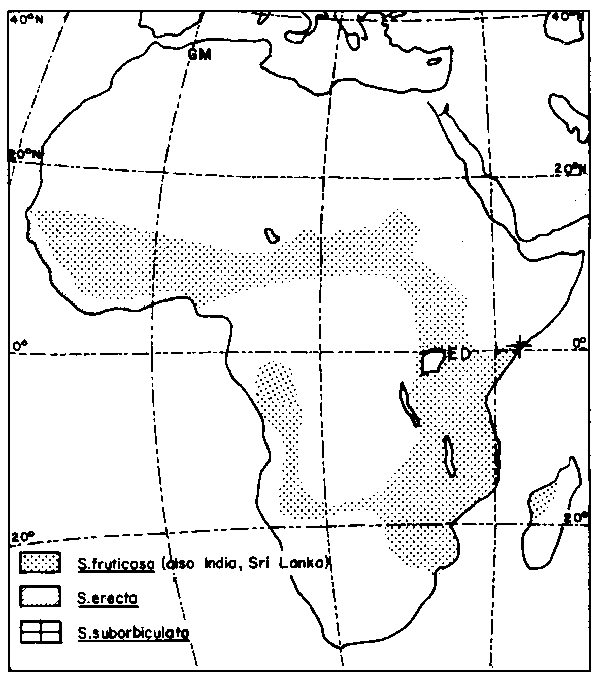
Source: Williams et al. (1984).
Principal component analysis and cluster analyses were done on the HP 3000 computer as described in the Zornia paper in these proceedings.
Table 2. Morphological and agronomic characters taken at Soddo, Ethiopia, on Stylosanthes fruticosa and their character states.
|
1. |
Plant growth form: 1 = erect; 2 = semi-erect; 3 = ascendant; 4 = ascendant/decumbent; 5 = decumbent, 6 = decumbent/prostrate; 7 = prostrate |
|
2. |
Plant height at centre of the plant excluding the central shoot (cm) |
|
3. |
Plant height to highest point of central shoot (cm) |
|
4. |
Canopy spread at the widest point (cm) |
|
5. |
Canopy spread at the narrowest point (cm) |
|
6. |
Mean number of branches originating from the main stem of all plants at or below 5 cm |
|
7. |
Length of the longest primary branch (cm) |
|
8. |
Average length of the spike (mm) |
|
9. |
Adaptability: 1 = negligible; 2 = very poor; 3 = poor; 4 = fair; 5 = good, 6 - very good |
|
10. |
Length of central leaflet of the leaf nearest to the spike (mm) |
|
11. |
Width of the central leaflet of the leaf nearest to the spike (mm) |
|
12. |
Central leaflet length/width ratio |
|
13. |
Distance between base of centre leaflet to junction point of the two outer leaflets: 1 = <1 mm; 2 = <2 mm; 3 = >2-<mm; 4 = >3-<4 mm; 5 = >4-<5 mm |
|
14. |
Stipule sheath length (mm) |
|
15. |
Plot cover: 1 = negligible; 2 = very poor; 3 = poor; 4 = fair; 5 = good; 6 = very good |
|
16. |
Height or distance of shoot tips from the ground level (mm) |
|
17. |
Expressivity of the central (main) shoot: 1 = negligible; 2 = very poor; 3 = poor; 4 = fair; 5 = good; 6 = very good |
|
18. |
Petiole length of the leaf nearest to the spike (mm) |
Principal Component Analysis
The correlation matrix for the characters under study (Table 3) shows a number of high correlations. Plant growth form was most highly correlated with the height or distance of shoot tips from the ground but had otherwise low correlations with other characters. Other high correlations included canopy spread at the widest point with the length of the longest primary branch; canopy spread at the narrowest point and adaptability; and adaptability with plot cover. These correlations indicate that principal component analysis may provide a useful summary of the variability in the data.
When principal component analysis was applied to the data, four components or factors were found to have eigenvalues greater than unity. These four factors explained 71.6% of the total variance with 4.176% eigenvalues of 4.176, 2.606, 1.920 and 1.318 respectively. Based on the sorted rotated factor loading (Table 4) the following interpretations were attached to the factors.
The positive end of factor one is associated with the attributes responsible for characters of agronomic interest such as plants which are highly branched and quite bushy. Factor two represents characters associated with large leaflets while factor three expresses the high plant growth structure. The fourth factor is concerned with high leaflet length to width ratio.
The factor scores of the individual accessions on the first three factors were plotted against each other. Some clustering could be identified, although most of the accessions appeared to be scattered randomly about the origin. On the plot of factor 1 versus factor 2, for example (Figures 2 and 3), about six clusters could be identified.
Cluster 1 was composed of six accessions. These were characterised by long and wide central leaflet of the leaf nearest to the spike petioles of the leaflet nearest to the spike being long and with distance between base of centre leaflet to junction point of the outer leaflets being between 1 and 2 mm.
Cluster 2 comprises ten accessions. These accessions have very good agronomic characters tending to those of S. scabra in growth form. They are quite bushy with a high branching pattern and very high adaptability and plot cover.
Cluster 3 is a uni-membered cluster composed of the accession 10539 whose central leaflet is very narrow and rather short. The accession, however, has similar agronomic features to those of the accessions in cluster 2, especially its wide canopy spread. This accession happens to be the one collected from the furthest east in the collection. It was collected from near Negele Borana in Sidamo, Ethiopia along the longitude of 39°30'E.
Cluster 4 is composed of ten accessions. Their main characteristic features are poor to fair adaptability and extremely poor to very poor plot cover. Exceptional cases are the accessions 13932, 13929 and 13973. However, 13973 has extremely poor plot cover. The rest of the accessions generally have few branches originating from the main stems at or below 5 cm.
Table 3. Correlation matrix for Stylosanthes fruticosa.
|
|
PLGRFM |
PLHTHT |
CANSPW |
CANSPN |
NOBR |
LPRBR |
ADAPT |
LFL |
LFW |
LFRAT |
DIST |
FLCOV |
JTSJT |
|
|
3 |
5 |
6 |
7 |
8 |
9 |
11 |
12 |
13 |
14 |
15 |
17 |
18 |
||
|
PLGRFM |
3 |
1.000 |
|
|
|
|
|
|
|
|
|
|
|
|
|
PLHTHT |
5 |
-.227* |
1.000 |
|
|
|
|
|
|
|
|
|
|
|
|
CANSPW |
6 |
.153 |
.193 |
1.000 |
|
|
|
|
|
|
|
|
|
|
|
CANSPN |
7 |
-.085 |
.314** |
.795** |
1.000 |
|
|
|
|
|
|
|
|
|
|
NOBR |
8 |
.154 |
.042 |
.482** |
.500** |
1.000 |
|
|
|
|
|
|
|
|
|
LPRBR |
9 |
.215* |
.160 |
.933** |
.799** |
.421** |
1.000 |
|
|
|
|
|
|
|
|
ADAPT |
11 |
.087 |
.384** |
.612** |
.663** |
.421** |
.585** |
1.000 |
|
|
|
|
|
|
|
LFL |
12 |
-.159 |
.108 |
.022 |
.113 |
-.040 |
.116 |
.127 |
1.000 |
|
|
|
|
|
|
LFW |
13 |
-.201 |
.076 |
-.057 |
.031 |
- 146 |
.036 |
-.031 |
.678** |
1.000 |
|
|
|
|
|
LFRAT |
14 |
.063 |
-.007 |
.080 |
.057 |
.141 |
.075 |
.147 |
.265* |
-.508** |
1.000 |
|
|
|
|
DIST |
15 |
-.233* |
.149 |
.160 |
.148 |
.059 |
.156 |
.060 |
.364** |
.529** |
-.244* |
1.000 |
|
|
|
PLCOV |
17 |
.108 |
.314** |
.539** |
.555** |
.319** |
.534** |
.798** |
.169 |
.063. |
077 |
.119 |
1.000 |
|
|
HTSHT |
18 |
-.630** |
.379** |
.111 |
.170 |
-.071 |
.061 |
.176 |
.052 |
.174 |
-.193 |
.210* |
.089 |
1.000 |
|
LPETR |
20 |
-.152 |
.121 |
.067 |
.227* |
-.028 |
.074 |
.053 |
.401** |
.454** |
-.141 |
.549** |
.111 |
.276** |
LPETR = 20
LPETR 20 1.000
Level of significance: 5% : > 0.209(*), 1%: > -/271(**)
Although this cluster exhibits a negative association of factors 1 and 2, there is no consistent pattern of expression of these characters for all the accessions in the cluster. These accessions, however, have a less than 1 mm distance between base of central leaflet to function point of the two outer leaflets. The central leaflets of the leaves nearest to the spikes are generally narrow and short, with a few exceptions being of medium length and fairly wide. These accessions other than those originating from Ethiopia have the longest spikes in the S. fruticosa collection at ILCA. The above mentioned features tend to separate accessions in cluster 4 from those of cluster 5.
Cluster 5 comprises nine accessions. These accessions have extremely poor to very poor plot cover and adaptability. They are of small stature and tend to form small bushes. Their leaflet and petiole characters are, however, almost similar to those of some accessions in clusters 1 and 2. It is interesting to note that although most of these accessions show poor adaptability, they were collected from Soddo, Sidamo in the neighbourhood of where they are currently being evaluated.
Cluster 6 is another uni-membered cluster comprising the accession 13924 from Niger. Like the other accessions appearing on the negative side of factor 1, it has very poor agronomic characters. It is also the only accession in the collection which was collected as far south in Niger as latitude 12°00'N.
Cluster Analysis
The 93 accessions of S. fruticosa in the final dendrogram were divided into ten clusters which distinctly are grouped into two main blocks comprising 15 and 77 accessions which will be referred to as groups A and B respectively. Group B comprising 77 accessions is solely composed of accessions collected from Ethiopia while group A comprises accessions from Ethiopia, Niger and CSIRO, Australia.
In group A the accessions which form the main cluster are 13942, 13850, 13860, 13893, 13890, 10829, 10523, 13919, 10431 and 10453. These are some of the accessions found with negative scores on factor 1 in the principal component analysis. These accessions are part of clusters 4 and 5 in the plot of PCA factor 1 versus factor 2 and are characterised by long spikes and short and narrow central leaflets of the leaf nearest to the spike. These accessions also have small bushes, very poor plot cover and adaptability.
Accessions in group B are characterised by large canopy spread, a fairly high number of branches originating from the main stem at or below 5 cm and good to very good adaptability. The above factor-loading matrix has been rearranged so that the columns appear in decreasing order or variance explained by factors. The rows have been rearranged so that for each successive factor, loadings greater than 0.5000 appear first. Loadings less than 0.2500 have been replaced by zero.
Table 4. Sorted rotated factor loadings for Stylosanthes fruticosa.
|
|
FACTOR 1 |
FACTOR 2 |
FACTOR 3 |
FACTOR 4 |
|
|
CANSPW |
6 |
.907 |
.000 |
.000 |
.000 |
|
LPRBR |
9 |
.890 |
.000 |
.000 |
.000 |
|
CANSPN |
7 |
.882 |
.000 |
.000 |
.000 |
|
ADAPT |
11 |
.813 |
.000 |
.000 |
.000 |
|
PLCOV |
17 |
.740 |
.000 |
.000 |
.000 |
|
NOBR |
8 |
.619 |
.000 |
.000 |
.000 |
|
LFW |
13 |
.000 |
.890 |
.000 |
.000 |
|
LFL |
12 |
.000 |
.767 |
.000 |
.565 |
|
DIST |
15 |
.000 |
.739 |
.000 |
.000 |
|
LPETR |
20 |
.000 |
.718 |
.000 |
.000 |
|
HTSHT |
18 |
.000 |
.000 |
.858 |
.000 |
|
PLGRFM |
3 |
.000 |
.000 |
-.790 |
.000 |
|
PLHTHT |
5 |
.296 |
.000 |
.640 |
.000 |
|
LFRAT |
14 |
.000 |
-.266 |
.000 |
.906 |
|
|
VP |
4.176 |
2.606 |
1.920 |
1.318 |
K-means Clustering
In the final it-means clustering analysis, divisions into 10 and 12 clusters were specified (Table 5). The relative importance of characters or variables in determining clusters can be judged by the "F-ratio" of the between to the within cluster mean squares. The most important variables were the average length of the spike, canopy spread at the narrowest point, the distance between base of centre leaflet to junction point of the two outer leaflets, canopy spread at the widest point, length of the longest primary branch, petiole length of the leaf nearest to the spike, plot cover and adaptability. The F-ratios for these variables were all above 10.0 with the highest being 28.0 for the average length of the spike. This can be illustrated by comparing clusters 2, 4 and 8. Cluster 2 is composed solely of accession 6745 which has the longest spike (of about 50 mm) among the collection; cluster 4 is also solely composed of 13342 which has the shortest spike (about 5 mm) and cluster 8 comprises accessions 13942, 13929, 13893, 13890, 13850 and 13860 whose spike lengths vary from 25 to 35 mm. The rest of the accessions had spike lengths ranging from 10 to 20 mm.
Figure 2. PCA for all characters Stylosanthes fruticosa.
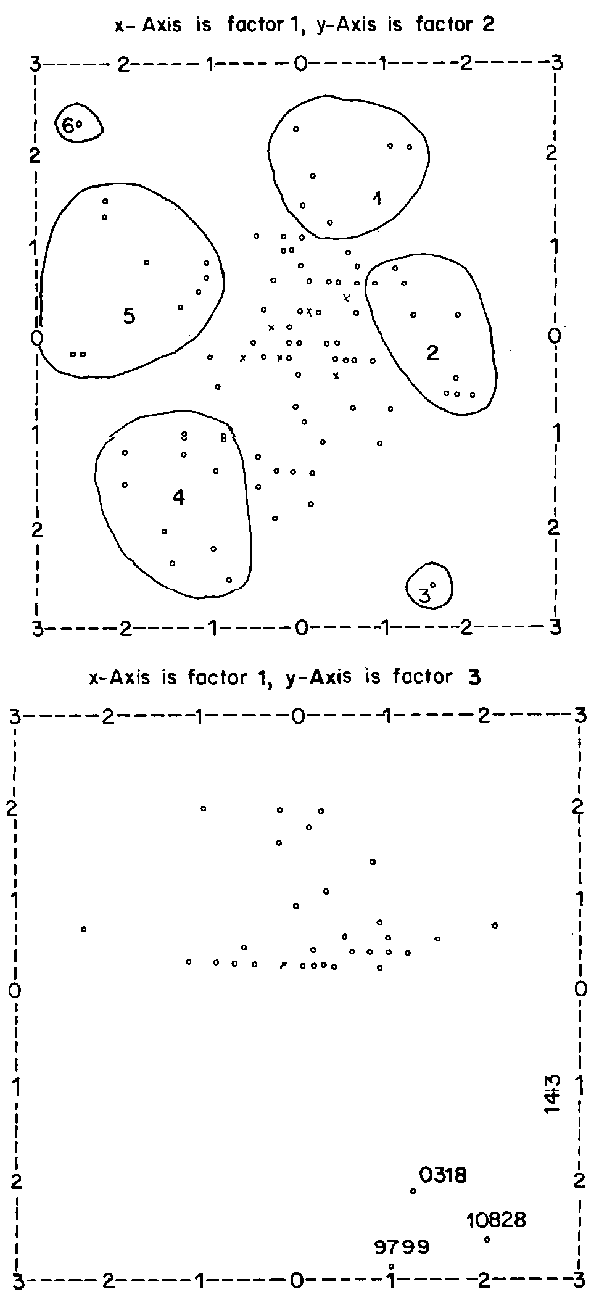
Scale is from -3 to +3. Factor scores greater than 3 are plotted as 3. Factor scores less than -3 are plotted as -3.
Figure 3. PCA for all characters Stylosanthes fruticosa.
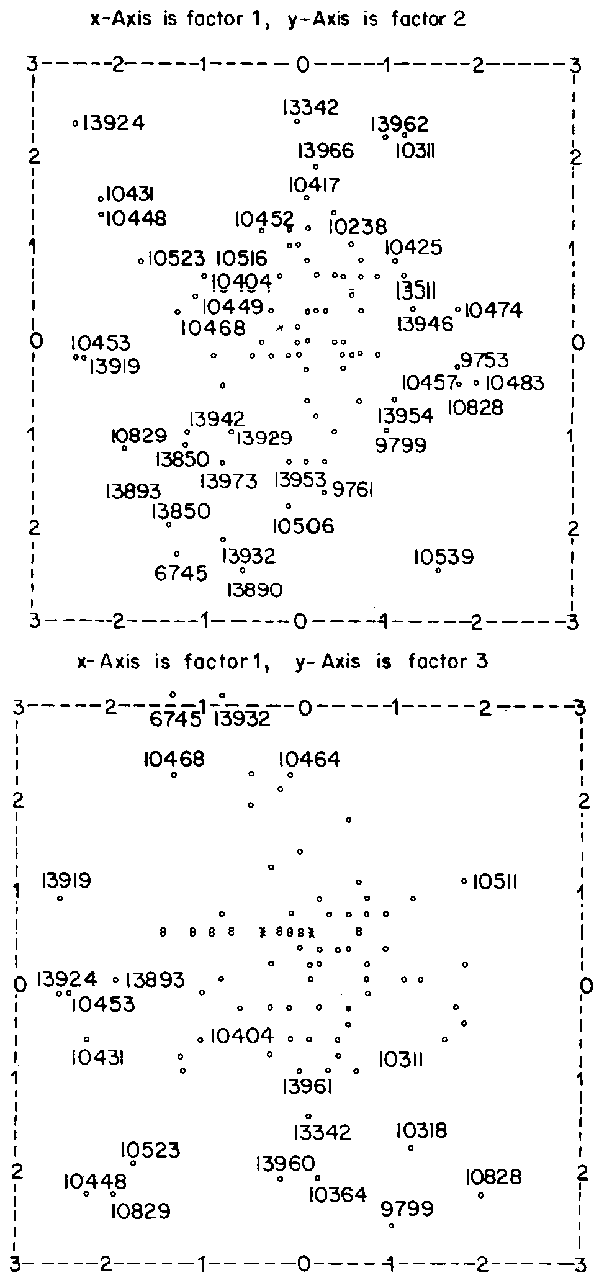
Scale is from -3 to +3. Factor scores greater than 3 are plotted as 3. Factor scores less than -3 are plotted as -3.
Table 5. Accessions constituting various clusters in the 10 and 12 group of K-means clustering.
Cluster analysis, principal component analysis and K-means clustering were used to group similar accessions. The 93 accessions available for study were agronomically and morphologically diverse. With cluster analysis, some strong clusters were formed. However, about one-third of the accessions were non-conformist. Accessions collected from Niger tended to form a group of their own while the Ethiopian materials also tended to form theirs. There were exceptions to this, however. Group A in the cluster analysis which had all the Niger accessions, except 13929, had materials of Ethiopian origin too. However, a sub-cluster existed that comprised wholly of materials from Niger. This cluster was also consistent in both principal component analysis and K-means clustering. It was not possible to record some of the most important characters in Stylosanthes species such as flowering, seed and fruit characters in this study.
For proper characterisation and evaluation of plant genetic resources it is necessary to start from seeding to harvesting in order to obtain more meaningful inferences. Where time is limiting, however, a study such as is described here can give guidelines on what characters to use during evaluation and possible variations existing among the materials. All the analyses gave similar grouping of accessions. These groups were related to the geographical location or types of environments from which these plants were collected rather than variations between accessions from similar geographical location. For instance, the Niger material tended to be grouped together, while the Ethiopian material tended to form "agrotypes".
Since ILCA's collections of S. fruticosa is from such a limited part of its range, further collections from other African countries are highly recommended on the basis of the following observations:
1. There is need to collect and preserve genetic materials that are in danger of disappearing as a consequence of the destruction of natural vegetation.2. There is need to increase the variability of collection from particular areas in relation to geographic, climatic and/or edaphic relevance.
3. There is need to collect genetic material in particular areas which in the past have been neglected or overlooked with regard to collecting activities.
Whereas it has been possible to obtain groups of accessions with particular characteristics based on the descriptions used in this study, the practical application of the findings has some limitations because of the omission of some important attributes. The variability existing within the species could therefore have been better described if more descriptors were used over a longer period of time. Despite the fact that little study has been done on S. fruticosa compared to the other Stylosanthes species, this characterisation and preliminary evaluation gives an indication of the variability or heterogeneity existing in S. fruticosa accessions currently available at ILCA.
We wish to acknowledge the technical assistance of Mr. James Ochang in growing and looking after the plants at Soddo, Ethiopia and other members of the Forage Legume Agronomy Group (FLAG) who provided transport and other necessary assistance.
One of the authors (J.J. Hakiza) wishes to acknowledge the financial support and authorisation to carry out the study by the International Board for Plant Genetic Resources (IBPGR) and the International Livestock Centre for Africa (ILCA).
Burt, R.L., Rotar, P.P., Walker, J.L. and Silvey, M.W. (eds.) 1983. The role of Centrosema, Desmodium and Stylosanthes in improving tropical pastures. Westview Tropical Agriculture, Series No. 6.
Engels, J.M.M. 1986. The systematic description of cacao clones and its significance for taxonomy and plant breeding.
IBPGR, Royal Botanic Gardens, Kew. 1984. Forage and browse plants for arid and semi-arid Africa. IBPGR Secretariat, Rome.
Lazier, J.R. 1984. Global ventures in Stylosanthes. IV. West Africa. In: H.M Stace and L.A. Edye (eds), The biology and agronomy of Stylosanthes. Academic Press, Melbourne, Australia.
Skerman, P.J. 1977. Tropical forage legumes. FAO Plant Production and Protection Series No.2. FAO, Rome.
Stace, H.M. and Edye, L.A. (eds). 1984. The biology and agronomy of Stylosanthes. Academic Press, Melbourne, Australia.
Thulin, M. 1983. Leguminosae of Ethiopia. Opera Botanica 68.
Williams, R.J., Reid, R., Schultze-Kraft, N.M., Coasta, S. and Thomas, B.D. 1984. Natural distribution of Stylosanthes. In: H. M. Stace and L. A. Edye (eds), The biology and agronomy of Stylosanthes. Academic Press, Melbourne, Australia. p.94.
|
|
ILCA No. |
Genus species |
Country of origin |
Original source or donor |
Collecting institute |
Other number |
Remarks about origin or source |
|
1 |
10388 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 47 N, 37 47 E |
|
2 |
10391 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 51.5N, 37 43.5E |
|
3 |
10404 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
4 |
10415 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
5 |
10417 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
6 |
10420 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
7 |
10423 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
8 |
10424 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
9 |
10425 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
10 |
10428 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
11 |
10482 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 50 N, 37 45 E |
|
12 |
10474 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 43.5E |
|
13 |
10471 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 43.5E |
|
14 |
10470 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 43.5E |
|
15 |
10468 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 46.5E |
|
16 |
10464 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 42 E |
|
17 |
10453 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
18 |
10452 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
19 |
10451 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 43.5E |
|
20 |
10450 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 43.5N |
|
21 |
10449 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 45.5N, 37 43.5E |
|
22 |
10448 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
23 |
10445 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
24 |
10440 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 46 E |
|
25 |
10437 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
26 |
10433 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
27 |
10431 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
28 |
10514 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 46.5E |
|
29 |
10488 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 50 N, 37 45 E |
|
30 |
10493 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 45.5N, 37 43.5E |
|
31 |
10502 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 43 E |
|
32 |
10506 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 42 E |
|
33 |
10507 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N |
|
34 |
10511 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44.5N, 37 42 E |
|
35 |
10513 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 46.5E |
|
36 |
10515 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 46.5E |
|
37 |
10516 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 46.5E |
|
38 |
10517 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 46.5E |
|
39 |
10523 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 43 E |
|
40 |
13945 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 58.5E |
|
41 |
13946 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53 N, 38 005E |
|
42 |
13947 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 31 E |
|
43 |
13949 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 31 E |
|
44 |
13950 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 31 E |
|
45 |
13951 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 31 E |
|
46 |
13953 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 31 E |
|
47 |
6745 |
Stylosanthes fruticosa |
Sudan |
CSIRO |
? |
CPI 412194 |
From Cunningham |
|
48 |
13978 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 35.5E |
|
49 |
13976 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 35.5E |
|
50 |
13975 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 35.5E |
|
51 |
13973 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 35.5E |
|
52 |
13972 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 35.5E |
|
53 |
13971 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 32 E |
|
54 |
13970 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 31 E |
|
55 |
13968 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 51 N, 37 34.5E |
|
56 |
13966 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 51 N, 37 31 E |
|
57 |
13965 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 35.5E |
|
58 |
13964 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 35.5E |
|
59 |
13962 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 51 N, 37 31 E |
|
60 |
13961 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 31 E |
|
61 |
13960 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 51 N, 37 31 E |
|
62 |
13959 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 51 N, 37 31 E |
|
63 |
13958 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 53.5N, 37 36.5E |
|
64 |
13954 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 52 N, 37 44 E |
|
65 |
9678 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
05 28 N, 38 16 E |
|
66 |
9752 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
05 28 N, 38 16 E |
|
67 |
9753 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
05 07 N, 38 17 E |
|
68 |
9761 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
04 38 N, 38 14 E |
|
69 |
9796 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
05 47 N, 39 17 E |
|
70 |
9799 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
05 36 N, 39 21 E |
|
71 |
10280 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 37 N, 37 34 E |
|
72 |
10288 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 25 N, 37 22 E |
|
73 |
10304 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 11 N, 37 36 E |
|
74 |
10311 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA- |
None |
06 44 N, 37 46 E |
|
75 |
10318 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 48 E |
|
76 |
10322 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 48 N, 37 45 E |
|
77 |
10364 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 07 N, 37 34 E |
|
78 |
10393 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 44 N, 37 39 E |
|
79 |
10396 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 42 N, 37 36 E |
|
80 |
10539 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
05 26 N, 39 30 E |
|
81 |
10828 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
05 43 N, 38 06 E |
|
82 |
10829 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
04 54 N, 38 06 E |
|
83 |
13942 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
12 21.2N, 02 22.2E |
|
84 |
13932 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
13 24 N, 09 03 E |
|
85 |
13924 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
12 00 N, 02 19 E |
|
86 |
13919 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
12 09.6N, 02 23.4E |
|
87 |
13893 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
12 17.8N, 20 33.1E |
|
88 |
13890 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
12 16 N, 02 27.6E |
|
89 |
13860 |
Stylosanthes fruticosa |
Niger |
ILCA/IBPGR |
ILCA IBPGR |
None |
12 29.9N, 02 24.4E |
|
90 |
13850 |
Stylosanthes fruticosa |
Niger |
ILCA/IBPGR |
ILCA IBPGR |
None |
12 27.8N, 02 23 E |
|
91 |
13342 |
Stylosanthes fruticosa |
7 |
CSIRO |
? |
CPI 41116 A |
From Cunningham Laboratory |
|
92 |
10432 |
Stylosanthes fruticosa |
Ethiopia |
ILCA |
ILCA |
None |
06 49 N, 37 47 E |
|
93 |
13929 |
Stylosanthes fruticosa |
Niger |
ILCA/IBPGR |
ILCA/IBPGR |
None |
12 24.4N, 02 22.6E |
![]()
![]()
![]()
{kind=link}