![]()
![]()
![]()
Abstract
Résumé
Introduction
Materials and methods
Results and discussion
Conclusion
Acknowledgement
References
B.A.O. Inyangala1, J.E.O. Rege1 and S. Itulya2
¹Animal Production Department, University of Nairobi
P O Box 29053, Nairobi, Kenya
2Animal Science Department, Egerton University
P O Box 536, Njoro, Kenya
Data on 969 lambs collected over a 10-year period (1978 to 1987) on Dorper sheep at Ol'Magogo were used in this study. Lamb traits studied were weights from birth to yearling and absolute growth rates between adjacent stages of growth. Heritability estimates from paternal half-sib analysis were: 0.15±0.07, 0.18±0.08, 0.39±0.11, 0.55±0.13, 0.53±0.13, 0.14±0.07, 0.28±0.09, 0.59±0.14, 0.49±0.12 and 0.49±0.12; for birthweight (PIRTHW), adjusted weaning weight (ADJWWT), adjusted six-months weight (ADJSMW), adjusted nine-months weight (ADJNMW), adjusted 12-months weight (ADJTMW), pre-weaning growth rate (GR1), weaning to six months (GR2), six to nine months (GR3), nine to 12 months (GR4), and birth to yearling (OVRGRT), respectively. Post-weaning heritability estimates were generally higher than those for pre-weaning suggesting a lower direct genetic influence early in life. Genetic and phenotypic correlations estimated between weights were mainly positive (0.15 to 0.99 and 0.02 to 0.98, respectively). There were generally high genetic and phenotypic correlations between adjacent weights, between weights and rates of growth, and among growth rates themselves.
Cette étude repose sur les données recueillies pendant 10 ans (1978 à 1987) sur 969 agneaux de race Dorper à OL'Magopo Naivasha. Les paramètres étudiés sont le poids des agneaux de la naissance à l'âge de 1 an, et leurs taux de croissance absolus à intervalles de temps consécutifs. Les coefficients d'héritabilité obtenus à partir de demi-frères paternels étaient de 0,15±0,07 pour le poids à la naissance, de 0,18±0,08 pour le poids au sevrage corrigé, de 0,39±0,11 pour le poids à 6 mois corrigé, de 0,55±0,13 pour le poids à 9 mois corrigé, de 0,53±0.13, 13 pour le poids à 12 mois corrigé, et en ce qui concerne la croissance, de 0,14±0,07 avant le sevrage, de 0,28±0,09 du sevrage à l'âge de 6 mois, de 0,59±0,14 de 6 à 9 mois, de 0,49±0,12 de 9 à 12 mois, et de 0,49 ±0,12 de la naissance à l'âge de 1 an. Les coefficients d'héritabilité des caractères étudiés après le sevrage étaient presque toujours supérieurs à ceux des caractères étudiés de la naissance au sevrage, ce qui signifie que l'influence directe du patrimoine génétique de l'animal est moins importante au cours des premiers mois de la vie qu'au cours des mois suivants. Les coefficients de corrélation génétique et phénotypique estimés entre les poids étaient essentiellement positifs, allant respectivement de 0,15 à 0,99 et de 0,02 à 0,98. Par ailleurs, il existait de fortes corrélations génotypiques et phénotypiques entre les poids moyens aux intervalles consécutifs, entre les poids et les taux de croissance et entre les taux de croissance eux-mêmes.
The potential for genetic improvement is largely dependent on the heritability of the trait and its genetic relationship with other traits of economic importance upon which some selection pressure may be applied. Information on heritabilities is essential for planning efficient breeding programmes and for predicting response to selection. Genetic correlations, on the other hand, are essential in predicting indirect responses to selection and are needed in order to determine the optimum weighting and expected response to multiple trait selection.
Statistical techniques presently utilised for the estimation of breeding values in selection programmes rely on estimates of genetic variation within and between traits of economic importance. The overall impact of any selection programme will depend on the direct and correlated responses that result from selection on the selection criterion. These responses can be predicted a priori by using estimates of genetic and phenotypic relationships between all traits of economic importance.
In Kenya, Dorper sheep are kept mainly for meat production. Thus, traits affecting economic viability include those associated with growth. Body weight and rate of gain are among the most economically important and easily measured traits of meat animals. Although weight is an important objective in selection, knowledge of the particular phase of the animal's growth upon which to base selection is of utmost importance. Genetic and phenotypic parameter estimates are scarce in sheep reared under Kenyan conditions and where such information is available, analytical methods used tend to be inadequate.
The purpose of this study was to estimate genetic and phenotypic parameters of growth traits in a flock of Dorper sheep at various stages of growth from birth to yearling.
The data used in this study consisted of growth records of lambs in a flock of Dorper sheep. These data were collected from records of the Sheep and Goat Development Project (SGDP) based at Ol'Magogo, a substation of the National Animal Husbandry Research Centre (NAHRC), Naivasha, between 1978 through 1987. A total of 969 records were available and were thus analysed for genetic and phenotypic parameters.
Bodyweights analysed included weight at birth, at three months (weaning), at six months, at nine months and at 12 months (yearling). Lambs were nursed by their dams up to weaning. Each lamb record included sire, dam and lamb identifications, type of birth, sex and dam breed. Three seasons of birth were defined on the basis of the monthly rainfall distribution. Two rainy seasons were identified with April and May (season 1) forming the peak of the long rains while October and November were classified as season of short rains (season 2). The remaining months were classified as dry season (season 3).
Parities were defined based on the number of times the ewes had lambed giving rise to parities 1, 2, 3 and 4, the latter comprising ewes with four or more lambings. Due to disproportionate distribution of data across years which resulted into discontinuity in the data, it was not possible to include actual years in the analysis. Therefore, to adjust for differences in weather conditions across years, periods of birth were defined by grouping adjacent years on the basis of annual rainfall pattern. This otherwise uncommon grouping method was implemented after examination of meteorological data, and was felt to be the best method under the circumstances. Period 1 consisted of the years 1978-1980 which received the highest rainfall, period 2 (1981-1985) received intermediate amounts while period 3 (1986-1987) received the lowest.
The absolute growth rates were derived by taking the difference in weight within the period and dividing it by the time interval in days. The absolute rate of gain for each lamb was calculated over five growth periods namely: Birth to weaning (GR1), weaning to six months (GR2), six to nine months (GR3), nine to 12 months (GR4) and birth to 12 months (OVRGRT). These, together with the bodyweights, constiute the 10 traits analysed in this study.
In view of the differences in actual age at which weights were taken, the latter were pre-adjusted as follows:
ADJWWT (adjusted 90-day weaning weight) = GR1 X 90 days + birthweight (BIRTHW)
ADJSMW (adjusted six-months weight) = GR2 X 90 days + ADJWWT
ADJNMW (adjusted nine-months weight) = GR3 X 90 days + ADJSMW
ADJTMW (Adjusted 12-months weight) = GR4 X 90 days + ADJNMW
Adjustment for fixed effects (sex, season of birth, period of birth and parity) was achieved by including them in the model.
Genetic and phenotypic parameters estimated were heritability of each trait, and genetic and phenotypic correlations among these traits as traits of the lamb.
Statistical analyses
The statistical model used to relate observations with independent variables was as follows:
Yijklmn = u + ai + bj + ck + dl + fm + eijklmn
where Yijklmn = the ijklmnpth observation.
u = an underlying constant for the trait
ai = effect of the i th sire (i=1,......,63) assumed random, N(0, s 2)
bj = effect of the j th season of birth (j = 1, 2, 3)
ck = effect of the k th parity (k = 1, 2, 3, 4)
dl = effect of the l h period of birth (l=1, 2, 3)
fm = effect of the m th sex (1 =male, 2=female)
e ijklmn = random error associated with the ijklmnpth observation; N(0, s 2) e
Parameters were estimated from covariances of relatives using paternal half-sib (PHS) analysis based on the Least Squares Method (Harvey, 1982) in which sires were crossclassified with fixed effects. The analytical complications introduced in half-sib analysis by use of litters was avoided by including only lambs born as singles. Heritability was estimated from variance components as
|
h2 = 4 s 2 /(s 2 + s 2) | ||
|
s |
s |
w |
where
h2 = heritability estimate
s 2 = sire variance component
s
s 2 = variance of records within sires
w
The approximate method of Swiger et al (1964) was used to estimate the standard error of the heritability estimate. Genetic correlations between two traits 1 and 2, r12 were estimated as
r12 = s s1s2 /(s s1s s2)
where s s1 s2 is the sire covariance component for the two traits and os1 and s s2 are square roots of respective sire variance components.
Genetic and phenotypic parameters
Heritability
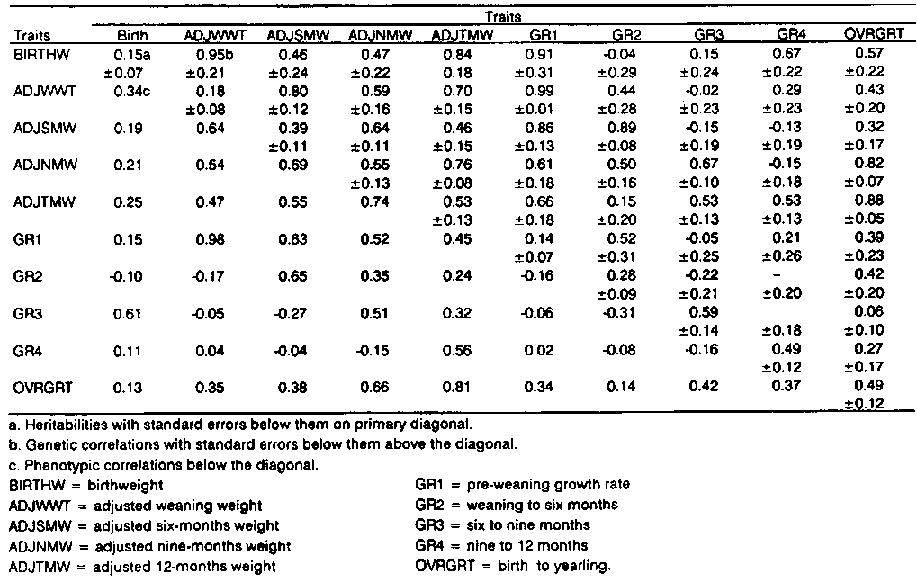
Heritability estimates are summarised in Table 1. Heritability of birthweight was found to be 0.15±0.07. This estimate compares favourably with estimates of 0.19 (Osman and Bradford, 1965) and 0.21 (Dzakuma et al, 1978). The estimate for weaning weight was 0.18±0.08 which was lower than 0.28±0.11 (Stobart et al, 1986) and 0.36±0.12 (Mavrogenis et al, 1980). Thus, in general, both estimates for birth and weaning weight were lower than those in the literature. Heritability for six-months weight was estimated at 0.39±0.11. This was slightly above the estimates of 0.28±0.10 (Dzakuma et al, 1978) and 0.21 (Ercanbrack and Price, 1972). The estimate for weight at nine months (0.55±0.13) compares favourably with the range of 0.30 to 0.50 (Rae, 1982). However, few studies have reported heritability estimates of nine-months weight. That for 12-months weight was estimated at 0.53±0.13 and was higher than values reported in other studies: 38±0.23 (sass and Acharya, 1970); 0.26±0.11 (Stobart et al, 1986); and 0.11±0.11 (Dzakuma et al, 1978). Heritability estimates of growth rates progressively increased from 0.14±.07 for pre-weaning growth rate to 0.59±0.14 for growth rate between six to nine months, then declined to 0.49±0.12 for rate of gain between nine to 12 months. The range of heritability for preweaning growth rate in the literature (Hundley and Carter, 1956; Givens et al, 1960; Vogt et al, 1967) was 0.18-0.37. Overall growth rate had a heritability estimate of 0.49±0.12. However, this was higher than 0.29±0.11 (Stobart et al, 1986). Rae (1982) reported a range of 0.20-0.40 for post-weaning growth rate while a high figure of 0.58±0.14 has been reported in Columbia sheep (Ercanbrack and Price, 1972).
It is clear from these and other results that post-weaning growth generally has higher heritability estimates than pre-weaning growth. This would indicate that environmental factors, in relation to additive genetic factors, had more influence on early lamb gain than on gains later in the lamb's life. This may be attributed to the high maternal influence associated with lamb growth performance early in life. High maternal influence has a tendency to increase the component of variance environmental to the lamb thereby lowering heritability estimates (Harrington et al, 1962; Thrift et al, 1973).
The best time to evaluate an animal's additive genetic value for a desired trait is under circumstances which assure maximum expression of the genes, that is under conditions when heritability is highest, provided that the genetic expression at this time is highly correlated with the genetic expression during the time period in life when the trait is most valuable or important (Ercanbrack and Price, 1972). However, one should not ignore the effect of time of evaluation on generation interval. Moreover, the desired genotype might be more accurately evaluated through another highly heritable trait, or an index, which is highly correlated genetically with it.
These results, therefore, indicate that to select lambs for their own genetic merit for weights and gains, it would be best to use bodyweight at nine months as the selection criterion rather than weaning weight as is often practiced. This trait should be superior to weaning weight or pre-weaning growth rate since it is much less influenced by maternal effects which tend to obscure the direct additive genetic effect for growth. Although the generation interval may not be considerably shortened, overhead costs should certainly be curtailed when the rest of the lamb crop is disposed of at this stage. On the other hand, selection of ewes as dams must be based on lamb performance pre-weaning and at weaning. In any case, the objective should be to choose a practical selection criterion which will maximise the annual rate of progress for the trait to be improved without seriously impairing merit in important correlated traits (Ercanbrack and Price, 1972).
Genetic correlations
Genetic correlations between growth traits at various ages was generally high and positive ranging from 0.15±0.24 to .99±.01 (Table 1). Genetic correlations between adjacent traits were generally higher than the ranges reported in the literature: 0.21-0.77 for correlation between birth weight and weaning weight (Osman and Bradford, 1965; Mavrogenis et al,: 1980), 0.13-0.22 for birthweight and 12-months weight, and 0.21 -0.72 for weaning weight and 12-months weight (Dzakuma et al,- 1978; Stobart et al, 1986). The high genetic correlations among these traits suggests that selection-for any one of these traits would result in considerable positive change in other traits. The complications brought about by maternal environment early in the lamb's life (hence the lowered estimate of direct genetic effects) may be overcome by utilising correlated response. One could concentrate on traits with high heritability as long as there exists a high positive correlation with other traits of economic value. Moreover, selection directed towards weights at later ages would minimise response in birthweight and possible increased frequency of dystocia (Shelton, 1964; Thrift et al, 1967; Olson et al, 1976; Martin et al, 1980; Mavrogenis et al, 1980). However, selection for weights at later ages would be expected to lead to increased yearling weights which is desirable for meat animals, but may be associated with increased maintenance costs for breeding animals and those kept for wool production.
Genetic correlations between growth rates and various weights and between growth rates themselves ranged from medium and negative to high and positive (-0.42 to 0.99). Statistically, most of the estimates were not significantly different from zero. Such (negative) estimates could be explained by the effect of compensatory growth obscuring underlying genetic relationships. Thus, in general, genetic correlations were low to high and positive. There is, therefore, potential for exploiting correlated response for most of these traits. Traits such as birthweight, weaning weight and pre-weaning growth rate could be incorporated in a selection index aimed at selecting for yearling weight since these traits are highly correlated with 12-months weight. However, care must be taken to avoid problems related to dystocia which may result due to selection for increased birthweights.
Phenotypic correlations
Phenotypic correlations between weights were all positive and generally high (0.19-0.74) (Table 1). As was the case for genetic correlations among weights, phenotypic correlations between adjacent weights were higher. The correlations tended to decrease as the time interval separating the observed weights increased.
In the case of genetic correlations between growth rates and weights, and among growth rates themselves, the phenotypic correlations ranged from low and negative to high and positive values (-0.17 to 0.98). The fact that there exists a negative (although low) correlation between pre-weaning growth rate and growth rate in the intervals weaning to six months and six to nine months (-0.16 and -0.06, respectively), may indicate that a faster pre-weaning growth rate because of substantial milk available tended to be followed by slower post-weaning growth rate. It seems logical to suggest, in general, that the low negative correlations which appear in some growth periods such as between weaning to six months and nine to twelve months (-0.08), and six to nine months and nine to 12 months (-0.16), are consequences of compensatory growth and not antagonistic as such. There was, as would be expected, a consistent positive phenotypic correlation between overall growth rate and all other growth rates of the lamb before yearling indicating common environmental effects.
From the results of this study' it is concluded that selection for weight and weight gain would best be based on post-weaning traits though the generation interval is likely to be slightly longer. Positive correlated response should be expected in other correlated traits due to the generally large and positive genetic correlations. Preliminary selection could also be conducted during the pre-weaning period since Inyangala (1989) reported favourable repeatability estimates for birth weight, pre-weaning growth rate and weaning weight. However, the high maternal influence on pre-weaning must not be ignored as it tends to mask the true genetic merit in the lambs.
This work received financial support from the Norwegian Agency for International Development (NORAD). Permission to use project data was granted by the Ministry of Livestock Development, Government of Kenya. The authors are indebted to the staff of the Sheep and Goat Development Project, Naivasha, for their support and cooperation.
Dass G S and Acharya R M. 1970. Growth of Bikaneri sheep. Journal of Animal Science 31 (1): 1 4.
Dzakuma J M, Nielsen M K and Doane T H. 1978. Genetic and phenotypic parameter estimates for growth and wool traits in Hampshire sheep. Journal of Animal Science 47: (5): 10141021.
Ercanbrack S K and Price D A. 1972. Selecting for weight and rate of gain in noninbred lambs. Journal of Animal Science 34:713.
Givens C S Jr, Carter R C and Gaines J A. 1960. Selection indexes for weaning traits in spring lambs. Journal of Animal Science 19:134.
Harrington R B. Brothers D G and Whiteman J V. 1962. Heritability of gain of lambs measured at different times and by different methods. Journal of Animal Science 21:78.
Harvey W R. 1982. User's guide for LSML76. Mixed model least-squares maximum likelihood computer program. Ohio State University, Columbus, Ohio, USA. (Mimeograph)
Hundley W S Jr and Carter R C. 1956. Heritability of growth rate and market grade in mutton lambs. Journal of Animal Science 15:1226 (Abstract).
Inyangala BAO. 1989. Genetic and phenotypic parameters for growth traits of the Dorper and Dorper x Red Maasai sheep. MSc thesis, University of Nairobi, Nairobi, Kenya.
Martin T G. Sales D I, Smith C and Nicholson D. 1980. Phenotypic and genetic parameters for lamb weights in a synthetic line of sheep. Animal Production 30:261.
Mavrogenis A P. Louca A and Robison O W. 1980. Estimates of genetic parameters for preweaning and postweaning growth traits of Chios lambs. Animal Production 30:271.
Olson L W. Dickerson G E and Glimp H A. 1976. Selection criteria for intensive market lamb production: Growth traits. Journal of Animal Science 43:78.
Osman A H and Bradford G E. 1965. Effects of environment on phenotypic and genetic variation in sheep. Journal of Animal Science 24:766.
Rae A L. 1982. Breeding. In: Coop I E (ed), World animal science. C. Production-system approach. 1. Sheep and goat production. Elsevier Scientific Publishing Company, Amsterdam, The Netherlands. pp. 15-55.
Shelton M. 1964. Relation of birth weight to death losses and to certain productive characters of fall-born lambs. Journal of Animal Science 23:355.
Stobart R H. Bassett J W. Cartwright T C and Blackwell R L. 1986. An analysis of body weights and maturing patterns in Western Range ewes. Journal of Animal Science 63:729.
Swiger L A, Harvey W R. Everson D O and Gregory K E. 1964. The variance of intraclass correlation involving groups with one observation. Biometrics 20:818.
Thrift F A, Whiteman J V and Ovejera A A. 1967. Evaluation of udder scoring in a ewe flock. Journal of Animal Science 26:206.
Thrift F A, Whiteman J V and Kratzer D D. 1973. Genetic analysis of preweaning and postweaning lamb growth traits. Journal of Animal Science 36:640.
Vogt D W. Carter R C and McClure W H. 1967. Genetic and phenotypic parameter estimates involving economically important traits in sheep. Journal of Animal Science 26:1232.
![]()
![]()
![]()
{kind=link}