![]()
![]()
![]()
3.1 Forage on offer
3.2 Forage intake
3.3 Energy requirements
3.4 Growth and milk production
3.5 Mortality
3.6 Reproduction
3.7 Herd management regime
3.8 The simulation process and initial herd composition
A general cattle herd simulation model in which a herd is simultaneously represented as both a biological and an economic system is used in the study. As the structure and detailed mathematical description of this model are presented elsewhere (Konandreas and Anderson, 1982), only its essential components and features are outlined here.
The model is time-dynamic, stochastic and non-optimizing, and treats simulated animals as individual entities. The parameters of mathematical representations of the various biological processes drawn from the literature are adjusted to particular systems under study, based on observations from these systems. Thus, the model is data-based where possible, and adequately modularized so that alterations and refinements can be made relatively easily. Another feature of the model is that it provides the user with an array of policy options so that herd performance can be studied under a variety of management regimes. These policy options allow for the simulation of certain parameters influenced by husbandry practices, such as breeding season and age of calf weaning. Additionally, input and output policy options can be specified with an adequate degree of detail relating to the management of the herd as an economic unit. On the input side, animals can be purchased and a range of supplements can be provided for increasing meat and milk production and/or for strategic reasons such as ensuring the survival of the breeding herd during drought periods. On the output side, milk and meat off take can be regulated as can the sale of surplus females over and above a planned herd size within the constraints of the available resource base.
The simulation of biological systems such as cattle herds requires, necessarily, one basic simplification. Although the links between the different interrelated processes of a system are time continuous phenomena, they must be specified in discrete time steps within a computer-simulated environment. The length of the time step used in this model is 1 calendar month. One month corresponds, in general, to the usual and practicable frequency of field data collection at the herd level. Such a correspondence is a prerequisite for model validation. Additionally, a monthly time step is within the accuracy required for specification of the management regime of the system in terms of breeding season, weaning age, supplementation strategies etc. Thus, the model can adequately simulate the impact of alternative management policies.
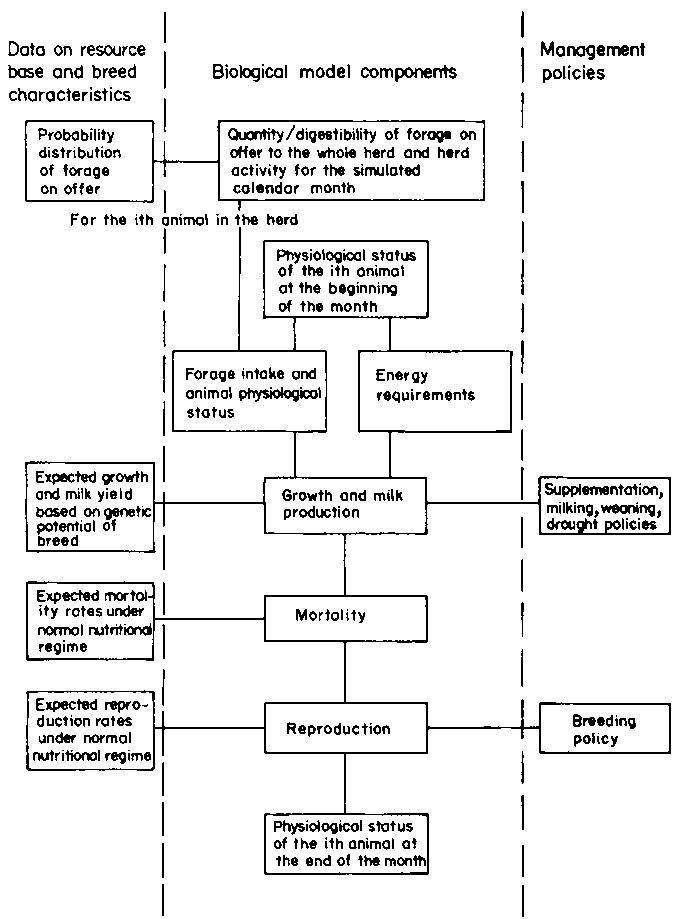
Within a time-dynamic environment, the various animal processes which determine the transition in the status of an animal from one month to the next must be mathematically described. These processes are illustrated diagrammatically in Figure 2. At the beginning of each month of simulation the model determines the quality and quantity of forage on offer to the herd and the level of animal activity for that calendar month. Forage on offer is simulated independently, based on historical time-series data. After the forage on offer has been obtained, the model determines the changes taking place in the status of each animal during the month, based on its genetic potential and using the endogenous biological processes regulated by exogenous management policies. Each animal in the herd is processed separately except for cows with suckling calves. As suckling calves secure at least some of their energy from their dams, their joint energy requirements must be determined simultaneously.
Six general components in the model account for changes in the biological status of animals during each month of simulation. These are forage on offer, forage intake, energy requirements, growth and milk production, mortality and reproduction.
Figure 2. Basic components of the simulation model.
The quantity and quality of forage on offer in the study area varies considerably from season to season within a year, and from one year to the next.
There are two ways in which this variability can be incorporated into a herd simulation model.
The first way consists of a quantitative, i.e. mathematical, description of the biological processes determining forage productivity, given the driving variables of the primary production system (rainfall, soil fertility, solar radiation, grazing pressure etc). Mathematical representation of the primary production system along these lines is difficult, as these biological processes are not well understood and data requirements are overwhelming (see for example Sullivan et al, 1981; van Keulen and de Wit, 1975).
The second way, which is used in this model, bypasses the underlying processes of the primary production subsystem and involves a statistical description of the quantity and quality of forage on offer based on field observations over several years, covering a wide spectrum of environmental variability. The seasonal level of activity of grazing animals is also associated with the degree of environmental variability. Animals respond to a situation of low forage density and inferior quality of forage on offer by exercising selective grazing, implying an increased level of activity.
Five year types have been identified to provide an adequate representation of environmental variability in the production system under study. At present with the stocking rates on the ranches studied, the quantity of forage on offer is not a constraint to animal intake. However, its quality varies considerably. Table I presents the digestibility, CP and animal activity vectors for the five year types identified. Also shown are the probabilities of occurrence of each year type and the corresponding months of the beginning of the growing season.
In 80% of all situations, growth starts in October and the forage consumed is of adequate quality (digestibility above 45% and CP above 5 %) until July. A below average year type implies a late start to the growing season in December or even January (5% probability), which results in an extended period (4 to 5 months) of very low quality of forage consumed (digestibility at or below 40% and CP as low as 3%). During these late-start-of-growth years, the onset of rains results in very high quality forage (digestibility 70% and CP as high as 13%), but due to the relatively short growing season the total quantity of forage produced is below normal. This high quality forage lasts for only 4 to 6 months, until April to May. In the simulation model, year types are drawn at random, based on the probability distribution of Table 1, with the further assumption of independence between successive draws.
The forage intake of extensively grazing cattle is influenced by the environment, the age and physiological status of individual animals and the quality of forage on offer. Considerable experimental work has been done, particularly in the last two decades, to quantify the separate influences of the individual determinants of intake 8. This work, although lacking standardization of measurement and, in some cases, adequate definition of the experimental animals involved, shows clearly that for a given quality of forage on offer, ad libitum intake is a function of body liveweight and the physiological status of individual animals (Conrad et al, 1964; Montgomery and Baumgardt, 1965; Elliot and Fokkema, 1961; Elliot et al, 1961; Hodgson, 1968). However, within a given functional form, estimated parameters can vary considerably between breeds and climatic conditions.
8 See overviews of experimental work by Balch and Campling (1962), Baile and Forbes(1974), end Cordova et al (1978).
The general voluntary intake relationship used in this model and referred to as physical limit is a modified version of the form suggested by Conrad et al (1964) and has the form
I = a W0.73/(1 - d) (3.1)
where
I = DM intake in kg/d,
W = body liveweight in kg,
d = digestible fraction of the forage on offer, and
a =a breed- and system-specific parameter whose value is a function of age and physiological status of individual animals.
At high digestibility levels, intake for mature animals is reduced due to chemostatic or thermostatic mechanisms (Conrad et al, 1964; Montgomery and Baumgardt, 1965; Baile and Forbes, 1974), implying a constant energy intake for these higher digestibility levels. It is therefore assumed in the model that for digestibility greater than 65%, feed intake is of a level such that the resulting metabolizable energy is equal to that obtained from the above relationship at the 65% digestibility level. This assumption implies a relationship (referred to as physiological limit) of the form
I = b W0.73/d (3.2)
where
b = 1.86a
Intake figures for different breeds and under various environmental conditions can vary considerably (see for example Cordova et al, 1978). Additionally, within a given breed and environmental regime intake is influenced by the physiological status of animals. For example, estimation of equation 3.1 using the data reported by Elliot et al (1961) and Elliot and Fokkema (1961) shows that cows in the last 3 months of pregnancy and lactating cows have a 7% and 15% higher intake respectively than dry, non-pregnant cows. The estimated coefficients "a" were 0.042, 0.045 and 0.049 for dry, pregnant and lactating cows respectively.
a Data compiled by APRU.
b Columns 1, 2 and 3 under each year type show: 1) average digestibility of forage consumed (%), 2) average CP in forage consumed (%), and 3) average distance walked (km/d).
Little is known about the voluntary intake of young calves. Hodgson (1968) experimented with calves from 3 to 6 months of age grazing on forage of digestibility ranging between 65 and 80% and observed that physiological limits are not constraining for young, fast-growing animals. Estimation of equation 3.1 using his figures yields a coefficient of 0.022, i.e. about 53% of the coefficient for dry cows obtained from the data of Elliot et al (1961).
The relative values of coefficient "a" for the different physiological conditions of animals are assumed as above for the purposes of modelling forage intake. However, the absolute levels of these coefficients are obtained from a calibration of equation 3.1 to the breeds and system under study. For this purpose a mature, dry and non pregnant reference cow is considered. Field data indicate that such a reference animal is in a state of liveweight equilibrium (i.e. neither gaining nor losing weight) for the month of July during a year of average forage quality. Stated alternatively, this implies that the daily DM intake during July in an average forage year is just sufficient to maintain body weight at the level of activity for that month. Solving equation 3.1 for intake requirements of the known quality for July, which are sufficient to cover maintenance and the animal's level of activity, yields an estimate of the intake coefficient "a" for the reference animal. The estimated value for the reference animal, along with those for the other animal classes as discussed above, are shown in Appendix Table A. 1. The intake coefficient for very young calves (3 to 6 months of age) is taken as 53% of the estimated coefficient for the reference animal, and is increased linearly until the level of the reference animal is reached at 18 months of age. Similarly, the intake coefficients of pregnant and lactating cows are taken as 107% and 115% of the estimated coefficient of the reference animal.
In addition to the above, there are other adjustments to the level of voluntary intake. When CP content of the forage on offer drops below 5%, which approximately corresponds to 40% digestibility, intake is reduced by the factor (d/0.4)0.6. Further, older animals (over 8 years) are assumed to have intake reduced by the factor [1 - 0.03 (age - 8)]. These correction factors have been argued to be appropriate adjustments to voluntary intake by Sanders and Cartwright (1979). Finally, the general model allows for adjustments to intake due to limitations in the quantity of forage on offer and grazing time limitations as a result of long walking distances to watering points or seasonal migrations. However, for the system under study these two factors are not constraining and thus not in effect.
Organic nutrients obtained from the feed consumed by an animal are used for a variety of functions, namely maintenance, construction of body tissues, the synthesis of milk and conversion to mechanical energy for work done by the animal. The net energy requirements for these activities are in general a function of the size of the animal and the level of each activity (MAFF, 1975; Blaxter, 1969; Webster, 1978). The conversion efficiency of nutrients to net energy for different body functions is higher the higher the quality of the feed consumed, and is also a function of its end use (Pigden et al, 1979; MAFF, 1975). Construction of body tissues, for example, has a lower conversion efficiency than net energy required for mechanical work.
In the model, the intake of feed energy is exactly balanced by its utilization for maintenance (plus pregnancy and lactation as appropriate) and weight gain or loss. The production and growth algorithms are different for calves and cows and weaned males. They determine liveweight change for calves or non-lactating cows and males, and both milk production and liveweight change for lactating cows. However, for the model to be able to predict the production levels of individual animals, parameters specifying the potential of the breeds under study must be provided.
Liveweight evolution from birth to maturity is a function of the genetic potential of the breed, the sex of the animal and its nutritional status at the different stages of its development. The model requires an age- and breed-specific average growth curve to be specified for both males and females. These average growth curves are used as a reference for determining the simulated condition and production potential of individual animals. Such curves can be estimated from recorded liveweight data (by age and sex) incorporating, to the extent possible, data acquired for the complete range of year types which can occur in the system under study. If this can be done then there will be minimum bias in the simulation towards higher or lower liveweight levels because the data to specify the model came from a particular sequence of favourable or unfavourable production years. When sufficient data are available it is possible to estimate not only averages but also measures of the distribution of liveweights around the means at any age. These statistics can then be used to estimate upper and lower limits of liveweight fluctuations (defining the boundaries of feasible live-weights), such that observed liveweights will lie within these limits with a specified degree of confidence.
Estimation of average growth curves in the model requires a minimum of three point estimates of liveweight evolution: weights at birth, at the age of growth slowdown (inflection point), and at the age of maturity. The model then fits a continuous and monotonically increasing curve consistent with these point estimates. Data on these three points of the growth curves for both T and ST genotypes are presented in Table 2.
For T cattle under ranching conditions in Botswana, average liveweights at birth are 31 kg and 28 kg for males and females respectively. The trend of the growth curve is almost linear up to about 18 months of age, at which time average weights are 310 kg and 280 kg for males and females respectively. Males reach a mature weight of 720 kg at about 72 months of age, while females reach a mature weight of 480 kg at about 54 months of age. The genetic growth potential of ST types is marginally higher. Average birth weights are 8% higher than for T types (APRU, 1980b), i.e. about 34 kg and 31 kg for males and females respectively. At 18 months of age and at maturity ST liveweights are about 11 % above T liveweights. Thus, average 18-month liveweights for ST cattle are taken as 340 kg and 310 kg for males and females respectively. Maturity is assumed to be reached at the same age as in T cattle, with mature weights of 800 kg and 516 kg for males and females respectively.
Liveweights of individual animals are assumed to be distributed normally around their age-specific means, with a coefficient of variation of 0.30 for calves up to weaning age, dropping linearly to 0.25 by 18 months of age and remaining constant thereafter. This coefficient implies that in two thirds of all situations the liveweights of mature T females would be between 352 kg and 607 kg and those of mature ST females between 379 kg and 653 kg. Similarly, with a 95% confidence level (which is taken as determining the limits of permissible liveweights in the model), the minima of female liveweights are 244 kg and 257 kg and the maxima are 714 kg and 768 kg for T and ST cows respectively. Such upper and lower boundaries exist for all ages and both sexes, calculated on the basis of corresponding mean liveweights and coefficients of variation, and define the feasible sets for liveweight evolution.
These liveweight boundaries are assumed to operate for all animals of each genotype, and are used as references for defining the liveweight condition of individual animals in the herd. This condition index is then used to modify their potentials for reproduction and milk production. Additionally, as explained in Section 3.5, the lower liveweight boundary represents the limit below which animals are assumed unable to survive unless they recover their lost weight 9.
9 It is recognized that this is a simplification of the complex factors which cause fluctuations in liveweights, as substantial differences are observed between individuals of the same breed. However, since data are not adequate to ascribe particular genetic potentials to each animal in the simulated herd, it is assumed here that all animals of the breed under study are genetically equivalent as regards their potential for liveweight growth and their tolerance to liveweight fluctuations. It is also assumed that animals of the same class and physiological status have identical constraints on intake and identical conversion efficiencies of feed to energy for production.
Specification of milk yields involves three components: data on the maximum potential yield per lactation of a cow in ideal condition and at the age when its reproductive capacity is maximal; age-specific milk yields per lactation (expressed as a fraction of the maximum potential yield); and finally, information for specifying the lactation curve within a given lactation.
The maximum potential yield per lactation for T cows in the system studied is assumed to be 1900 kg and that of ST cows 3500 kg 10. It may be achieved by animals between 5 and 8 years of age. For animals older than 8 years maximum potential milk yield per lactation decreases gradually to 70% of this level by 12 years of age. For animals younger than 5 years maximum potential milk yield per lactation decreases gradually to 70% of the above level for animals in their first lactation. Appendix Table A.2 provides complete details of the assumed effect of age on potential lactation yields.
10 The maximum potential milk yield of a pure Scow is about 5100 kg per lactation. The maximum potential milk yield of an ST F1 cross is estimated as midway between the yields of the two pure breeds, that is about 3500 kg. Additionally, the butter fat content of T milk is assumed to be 52 g/kg, which corresponds to an energy content of 3.5 MJ/kg. The butter fat content of pure S milk is assumed to be 40 g/kg and, assuming again that the butter fat content of ST milk is midway between that of the two breeds, it follows that the energy content of ST milk is 3.3 MJ/kg.
Table 2. Summary of production parameters used in the model for the T and ST genotypes in the environment of the study area a.
|
Attribute |
Genotype |
Males |
Females |
|
Growth parameters: |
|||
|
Liveweight at birth (kg) |
T |
31 |
28 |
|
ST |
34 |
31 |
|
|
Expected liveweight at 18 months (kg) |
T |
310 |
280 |
|
ST |
340 |
310 |
|
|
Expected liveweight at maturity b (kg)
|
T |
720 |
480 |
|
ST |
800 |
516 |
|
|
Coefficient of permissible liveweight variability |
|
0.25 |
0.25 |
|
Mortality parameters c: |
|||
|
Survival rate to 1 year (%) |
|
97 |
97 |
|
Survival rate to 2 years (%) |
|
96 |
96 |
|
Mortality rate for 3- to 8-year-olds (%/year) |
|
0 |
0 |
|
Mortality rate for 12+ year-olds (%/year) |
|
2 |
2 |
|
Fertility parameters: |
|||
|
Reproductive maturity |
|
|
|
|
- expected age in months (and liveweight in kg) |
T |
|
20 (302) |
|
for animals in normal condition |
ST |
|
20 (328) |
|
- earliest age in months (and liveweight in kg) |
T |
|
12 (318) |
|
for animals in very good condition |
ST |
|
12 (332) |
|
- latest age in months (and liveweight in kg) |
T |
|
42 (233) |
|
for animals in very poor condition |
ST |
|
42 (251) |
|
Expected annual calving rates c |
|
|
|
|
-for 2-year-old cows(%)
|
T |
|
80 |
|
ST |
|
88 |
|
|
- for 5- to 8-year-old cows (%)
|
T |
|
92 |
|
ST |
|
97 |
|
|
- for 12+ year-old cows (%)
|
T |
|
80 |
|
ST |
|
88 |
|
|
Lactation parameters: |
|||
|
Maximum potential milk yield per lactation |
T |
|
1900 |
|
for 5- to 8-year-old cows (kg) |
ST |
|
3500 |
|
Fraction of maximum potential milk yield for cows in their first lactation (%) |
|
|
70 |
|
Fraction of maximum potential milk yield for 12+ year-old cows (%) |
|
|
70 |
|
Maximum lactation period (months)
|
T |
9 |
|
|
ST |
|
10 |
|
a All data relating to the genetic potential of the two genotypes in the environment of the study area have been taken or constructed from APRU (1980b, 1981), Buck et al (1976), Pratchett et al (1977), Rennie et al (1977), Trail et al (1977) and personal communication with scientists at APRU. See text for additional qualifications on individual parameters reported here.
b Maturity is reached at 72 months for males and 54 months for females.
c Under adequate nutritional regime.
The maximum duration of any given lactation in this system is assumed to be 9 months and 10 months for T and ST cows respectively. Additionally, it is assumed that 33% and 28% of the milk yield of a given lactation is produced during the first 2 months post-partum for T and ST cows respectively. On the basis of this information the model approximates the shapes of the lactation curves for the two breeds as presented in Appendix Table A.3. Thus, for example, the simulated absolute maximum daily milk yield of a T cow between 5 and 8 years of age, during the first 2 months post-partum, is calculated as 10.45 kg/d, and that of an ST cow as 16.33 kg/d. These maxima may or may not be achieved, depending on the feeding regimes of individual animals during their lactation period.
The mortality component of the model determines whether an animal dies during a given month of simulation. Mortality is modelled as a probabilistic process qualified by the age of the animal and its nutritional status.
Mortality due to epidemics is minimal in ranching systems in Botswana as a result of effective disease control measures. Mortality is usually accounted for either by nutritional stress due to occasionally inadequate feed supplies, or by a complex set of factors not directly related to their nutritional status.
The lower boundary of liveweights, as obtained from the assumptions made in Section 3.4, corresponds to the poorest condition in which an animal can remain in the simulated herd. If an animal loses sufficient weight or does not gain weight as it should in accordance with its increasing age, it eventually develops an age - liveweight combination below this lower boundary. When an animal's liveweight drops below this boundary and the lost weight is not recovered within the subsequent month of simulation, then it is assumed in the model that death due to nutritional stress occurs. This source of death is deterministic in the sense that simulated death inevitably occurs when these conditions apply.
Animal losses not relating to nutritional status (referred to as "normal" losses) are dependent on age. For animals over 2 and up to 8 years of age such losses are insignificant. However, losses of younger animals which are not related to nutritional stress can be substantial. Overall mortality to 1 and 2 years amounts to 5.5% and 6.5% respectively in the system under study (including both nutritional stress and all other causes). APRU (1980b) identified nine causes of mortality to 2 years of age (stillbirths, accidents, predators etc.), which accounted for 50% of all reported deaths. The causes of the residual 50% of deaths are unknown. It is assumed here that in the total mortality to 2 years causes not associated with nutritional stress account for 75% of all reported mortalities, with the residual 25 % being the result of nutritional stress. This implies a "normal" mortality rate of 0.75% (75% of 1%) for animals between 1 and 2 years of age.
For animals over 2 and up to 8 years, "normal" annual mortality is assumed to be zero, with all probable mortalities for animals within this age interval being the result of nutritional stress. Finally, for animals over 8 years of age "normal" annual mortality rates are assumed to increase gradually to 2% by 12 years of age, reflecting problems associated with older age. This assumption implies an annual mortality rate of 0.1%, 0.5 %, 1.1% and 2%, for animals in the age groups 8-9, 9-10, 10-11 and 11+ years respectively.
The mortality information provided to the model to account for "normal" losses is used to generate monthly probabilities of mortality which are the test values in a binomial distribution trial. Death occurs if a randomly selected number between 0 and 1 is below the appropriate test value for the animal under consideration.
The reproduction component of the model determines whether a non-pregnant cow conceives and a pregnant cow gives birth during a given month. Conception is modelled as a probabilistic process and is further qualified depending on whether the animal is a heifer or a mature cow. In addition to the nutritional status of individual animals, conception may be influenced by exogenous management practices, indicated as breeding policy in Figure 2, regulating the length of the breeding season and specifying a minimum weight and age of individual animals before they are first mated.
The specification of female fertility requires firstly information on reproductive maturity, and secondly age-specific expected calving rates for mature cows obtained from the system under consideration. The fertility and availability of males are assumed not to be limiting factors.
Onset of puberty is a function of weight and age, and on average occurs at about 20 months of age for both T and ST animals. For animals with a very favourable liveweight growth, puberty can be reached by 12 months of age, and at the other extreme animals with a very poor liveweight growth reach reproductive maturity at 42 months of age. Table 2 presents the ages for reproductive maturity associated with different liveweight growth conditions, and the corresponding liveweights at maturity. These liveweights have been obtained from the expected liveweight evolution curves and associated liveweight boundaries for the two breeds as shown in Figure 3. Two exemplary cases are also demonstrated in this figure. In Case I, an ST female with a normal growth pattern reaches reproductive maturity at 18 months weighing 330 kg (point R). In Case II, a T female with below average growth reaches delayed reproductive maturity at 30 months weighing about 270 kg (point R) 11.
11 A female reaching reproductive maturity does not necessarily conceive. Her probability of conception is a function of the expected calving rate (successful conceptions) for her age-class, and additionally of the breeding policy in effect (see Section 3.7).
Figure 3. Liveweight - age combinations for reproductive maturity.
The probability of successful conception for all animals of reproductive age is assumed to be a function of age, and within an age-class a function of the liveweight conditions of individual females. Maximum reproductive capability is assumed to apply for animals between 5 and 8 years of age. Before 5 and after 8 years reproductive performance is reduced. Table 2 provides data on expected calving rates by age for T and ST genotypes. The detailed age-specific calving rates and corresponding expected monthly probabilities of conception are given in Appendix Table A.4. Actual conception rates for cows within a given age group can be lower or higher than these expected values, depending on the condition of individual cows (Buck et al, 1976)12. Thus, if the model selects a succession of 2 years with favourable production conditions simulated then conception rates will be above the average, and vice-versa for a sequence of unfavourable production conditions.
12 Expected conception probabilities apply to cows of normal liveweight for their age. The probability conception drops linearly by 10% for animals 20% below normal weights, and by a further 40% for animals near the lower liveweight boundary. This is closely in line with results reported by APRU (1981), whereby animals weighing less than 340 kg had a reconception success about 30% below animals weighing over 450 kg. For animals with liveweights above normal, the probability of conception is assumed to increase linearly by 10% for animals 15% above normal weights, and to decrease thereafter to 90% of the expected probability level for very heavy animals (near the upper liveweight boundary).
A well defined management regime is required by the simulation model. Management options in the model should, to the extent possible, replicate practices in the real system. In the ranching system as it is presently managed, specific weaning, breeding and sale policies are followed. The specification of these policies in the model are described in this section and summarized in Table 3.
Breeding is completely controlled in this system and occurs only during 3 months of the year, from January to March inclusive. This implies carvings from October through December. In addition to the season of breeding, management has control over the age and weight of animals to be bred. Individual animals are bred only if they are older than 24 months and maintain a liveweight of at least 270 kg for T and 290 kg for ST animals during the breeding season.
Calf weaning takes place at 7 months of age, regardless of the condition of the calf and the milk yield of its dam at that time. As carvings take place from October to December, weaning occurs from May to July. However, weaning necessarily occurs at an earlier age in the case of death of the dam. No milk is removed for human consumption. All the milk produced is consumed by calves, and dams cease lactating immediately their calves are weaned.
The sales policy specifies the conditions under which males and surplus females are removed from the herd. Males are sold at the end of June after they have reached at least 2.5 years of age. Sales of females take place so that a breeding herd size of 40 animals is maintained 13. Selective disposal is followed with the objective of progressively eliminating the most unproductive animals until the target herd size is achieved. Productivity is measured by the simulated reproductive performance of individual animals.
13 The size of the breeding herd, at 40 animals, was arbitrarily selected here. It is large enough to reflect the dynamics of the herd and small enough not to present serious computational problems. It does not necessarily reflect an economically optimum herd size. For example, APRU (1980) reports that the optimum for a beef cattle production herd is somewhere near 600 LUs or about 200 breeding cows.
Animals reaching 13 years of age are compulsorily sold regardless of their past reproductive performance, as their present potential for reproduction is considerably reduced. These old cows are sold during August so that they can complete their current lactation period, if they happen to be lactating. Second in the order of sale are cows with relatively long calving intervals. These sales take place during June, when animals are in relatively good condition, and only if the time since their last calving is over 16 months and they have not yet conceived. As carvings occur at the earliest during October and conception takes place at the latest during March, this policy implies that cows must miss two complete breeding seasons to be sold because of poor reproductive performance. Third in the order for sale are heifers which have not yet conceived. Again, the sale of females in this category takes place in June. Finally, if the aggregate of sales from the above categories does not reduce the size of the breeding herd below 40 animals, older animals are sold, starting with the oldest and progressively lowering the sale age from 13 down to 10 years of age. June is again the month of sale for females in this category.
Table 3. Summary of management practices in the system under study.
|
Practice | ||
|
Weaning: |
|
|
|
Age at weaning (months) |
|
7 (both sexes) |
|
Breeding: | ||
|
Minimum breeding age (months) |
|
24 |
|
Minimum weight for breeding (kg) |
T |
270 |
|
|
ST |
290 |
|
Breeding season (inclusive months) |
|
Jan.-March |
|
Sales: | ||
|
Minimum sale age (months) for males and calendar month of sale |
|
30 (June) |
|
Minimum sale age (months) for old cows and calendar month of sale |
|
156 (August) |
|
Target size of breeding herd a |
|
40 |
|
Calendar month for sale of surplus females |
|
June |
|
Minimum infertile period for sale of less productive females (months) |
|
16 |
a See footnote 13 for additional qualifications.
Experimentation with this herd simulation model consists of a quantitative description of the productivity of the livestock system under study, evolving over a predetermined number of years. Figure 4 is a schematic representation of this production system showing its inputs and outputs. The production process starts at the beginning of the simulation period, with an initial herd which evolves into the final herd at the end of it. Intermediate inputs and outputs during this period include the feed consumed and the milk and meat produced. The evolution of the herd is a stochastic process. Starting with the same initial conditions and management regime, there exists an infinite number of realizations of the evolution of the herd through the entire simulation period. Thus, in the context of a stochastic simulation model, the outcome consists not of a single realization but of a probability distribution of realizations. Realizations differ from each other not only because of the variability in the forage regime, but also because of the inherently random outcome of the various biological processes of conception and mortality, the sex of offspring etc., embedded in the model.
A simulation experiment for a livestock production system under a given management regime involves a series of independent realizations of the system's outcome over a specified time horizon. The time horizon considered here is 15 years, and each experiment is replicated 10 times over the 15-year period to establish the statistical significance of the simulated results.
An initial total herd size of 60 animals is considered, consisting of females only with an age composition as shown in Appendix Table A.5. Out of the total herd 40 animals are of reproductive age (i.e. over 2 years of age). The first month of the simulation is October, at which time 34 out of the 40 cows are pregnant and expected to calve during October, November and December. Initial liveweights are assumed slightly below the average liveweights corresponding to the animals' age, to reflect the generally below average liveweights prevailing in the real system at the beginning of October.
Figure 4. Summary of inputs and outputs of the system under study.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
{kind=link}