![]()
![]()
![]()
* Based on papers presented by A Blair Rains, P N de Leeuw, J C Bille and W Doppler and discussion led by R Rose Innes.
Herbaceous vegetation
Woody vegetation
Pasture management
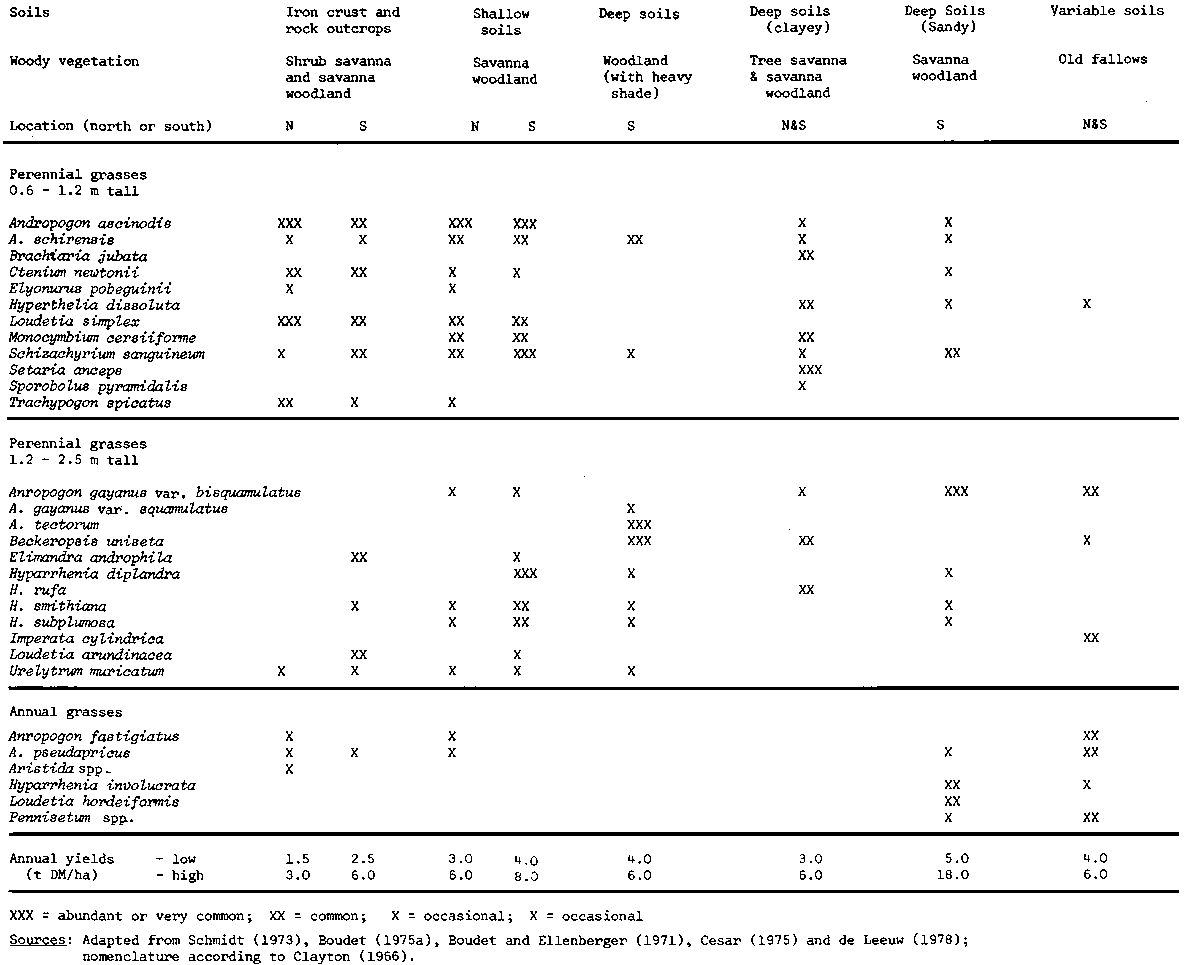
According to Rattray (1969), the grass cover of the subhumid zone can be classified into two broad belts: the northern part of the zone where Andropogon gayanus predominates and the southern area where the Hyparrhenia species are more important. More recent rangeland surveys, however, have shown that although climatic variation brings about a modest segregation of grass species, soil conditions, in particular soil depth and texture, and site history, especially related to cultivation and burning, have a much greater impact on the species composition of savanna vegetation than climate (Boudet, 1975a; de Leeuw, 1978a). The principal grass species of the subhumid zone are shown in Table 4 in relation to soil types and woody vegetation.
Biomass Production
The upper and lower limits of end-of-season herbage yields at a number of sites in the subhumid zone are shown in Table 4. Yields range widely from 1 500 to 18 000 kg DM/ha. In contrast to the modest effect of latitude on the distribution of grass species, the quantity of herbage growth increases markedly from north to south on rangelands with similar species composition. For instance, annual yields at Olokomeji, near Ibadan in southern Nigeria, were found to be twice as high as at Shika further north (see Table 5). It has been suggested that the main factor determining yield is the longer growing season in the south, rather than the higher mean annual rainfall (de Leeuw, 1978a).
In addition to the effects of latitude, grass yields are closely related to the catenary sequence of soils. The lowest yields are recorded on stony, shallow and gravelly soils which have a low, sparse grass cover, partly consisting of annual species. Highest yields are associated with deep loamy soils, usually ferrisols, in the derived savannas where tall grasses thrive (Lamotte, 1975; Boudet, 1975a, Egunjobi, 1973). On shallow soils over ferruginous crust at Shika, yields rarely reach 2 500 kg DM/ha, compared to a production of 6 000 kg on deep, sandy alluvial clays in the same area (see Table 5). Similar differences in yields according to soil types have been recorded by Afolayan (1975) in the Borgu Game Reserve in Nigeria and by Lamotte (1975) in Ivory Coast.
Shade from the woody canopy not only affects species composition of the grasses (see Table 4), but also affects herbage yields. At Afaka, near Kaduna in Nigeria (10° 35° N), the annual herbage yield in places without tree cover was found to average 3 200 kg DM/ha, compared with 2 300 kg DM/ha in more shady places at the same location. However, the shade tolerance of the tall woodland grasses, called 'fringe grasses' by Rose Innes (1977), is high; they are only suppressed in areas where the tree canopy is almost entirely closed. This explains why in Isoberlinia tomentosa woodland in northern Nigeria with a tree cover of 66%, a dense tall perennial grass cover (53%) survived, yielding 3 500 kg DM/ha annually, when subjected to irate annual burning (Afolayan, 1975).
In shrub savanna, woody cover is usually inversely related to grass yield, since low shrubs compete directly with grasses for light, and probably for moisture and soil nutrients as well. Rose Innes and Mansfield (1976), working on the Kaduna plains in Nigeria, estimated the standing biomass on four- to eight-year-old wooded fallows at 1 000 to 1 200 kg DM/ha, or less than half the average measured on older tree savannas or undisturbed savanna woodland.
Despite these large differences in end-of-season yields, most perennial grasses in a monsoonal climate follow a similar pattern of growth. After a short initial period of slow growth in the early rainy season, vegetation growth is linear for three to four months until stem elongation occurs preparatory to flowering, when growth again accelerates. This period of rapid growth begins in late August for short grasses in the northern part of the zone, but occurs several weeks later in the south. Tall grasses generally mature later than shorter species.
As the end of the growing season approaches, growth rates decline, as shown in Table 5. At Shika in Nigeria, on shallow gravelly soils under shrub savanna, the growth rate of the grasses begins to drop in mid-October, whereas at sites further south or with deeper soils, the growth rate begins to drop more than one month later. The sites under comparison included a savanna woodland area at Olokomeji, a tall grass savanna on deep colluvial soils along the Shika River and a tall Andropogon gayanus grass savanna on deep sandy soil at Fashola, all in Nigeria. These differences in rangeland production are reflected in daily vegetative growth rates which can vary from 15 to 57 kg DM/ha, increasing to a range of 29 to 145 kg DM/ha towards the end of the rains. Egunjobi (1973) recorded a peak daily production of 270 kg DM/ha on Andropogon gayanus fallow at Fashola, which is in contrast to only 32 kg DM/ha for the same species at Shika (Haggar, 1970).
In the drier areas of the subhumid zone, growth rates diminish rapidly as soon as the residual moisture supply in the soil is exhausted. In southern Mali, which is relatively dry, low daily growth rates of 0.8 to 2.5 kg DM/ha were measured early in the dry season (Boudet and Ellenberger, 1971), while at Shika, which is also a relatively dry area, very little growth was observed after cutting in October (Haggar, 1970). In more humid regions, the growing season is more prolonged. Piot and Rippstein (1975) recorded a daily growth rate of 2.7 to 4.5 kg DM/ha on the Adamawa Plateau in Cameroon at the end of a 240-day growing season, while in the even more humid forest savanna mosaic, Boudet (1975a) recorded daily growth rates early in the dry season of 5 kg/ha for Loudetia arundinacea on shallow soils and 7 to 8 kg/ha for Hyparrhenia diplandra on deeper soils.
Table 5. Herbage accumulation in ungrazed rangeland at four sites in Nigeria (tonnes DM/ha)
|
|
May 1 |
June 1 |
July 1 |
Aug 1 |
Sept 1 |
Oct 1 |
Nov 1 |
Dee 1 |
|
Shika (11° N) |
0.10 |
0.35 |
0.75 |
1 45 |
2.40 |
3.20 |
3,10 |
3.00 |
|
Shika River (11° N) |
0.35 |
0.65 |
1.20 |
2.70 |
4.10 |
4.10 |
6.15 |
- |
|
Olokomeji (7° 25' N) |
1.30 |
2.00 |
3.00 |
3.70 |
4.80 |
5.65 |
6.00 |
6.10 |
|
Fashola (7° 55' N) |
1.70 |
3.20 |
5.20 |
5.50 |
11.60 |
16.20 |
15.30 |
- |
Sources: For Shika and Shika River, de Leeuw (1971a) (data collected in 1968); for Olokomeji, Hopkins, private communication (data collected in 1957-58); for Fashola, Egunjobi (1973).
As the dry season continues, growth is further reduced until the soil moisture deficit is replenished by the early rains and growth recommences. Growth rates at the beginning of the rains are also greatly affected by the climate and the depth of the soil as shown in Table 6.
Table 6. Daily growth rates of burned vegetation during the second half of the dry season and the early rainy season at five sites in the subhumid zone
|
|
Daily Regrowth (kg DM/ha) |
|
|
Late Dry Season |
Early Rainy Season |
|
|
Savanna on shallow soils at Yanfolila, Mali (11° N) |
1.5 |
- |
|
Shrub savanna on shallow soils at Shika, Nigeria (11° N) |
1.9 |
8.8 |
|
Grass savanna on deep, low-lying soils at Shika, Nigeria (11° N) |
6.1 |
13.6 |
|
Savanna woodland at Olokomeji, Nigeria (7° 25' N) |
- |
22.9 |
|
Andropogon gayanus grass savanna at Fashola, Nigeria (7° 55' N) |
- |
41.3 |
Sources: For Yanfolila, Boudet and Ellenberger (1971), for Shika, de Leeuw (private communication), burning between late December and mid-April, figures derived from regression equation, y = 1.93 x + 14.4 (n = 28, r = 0. 81), where x = days after burning and y = DM yield in kg/ha; for Olokomeji, Hopkins (private communication); for Fashola, Egunjobi (1973), burning in late February.
Nutritive Value
Several elements may be considered in evaluating the nutritive value of savanna forage, such as the nitrogen, calcium, phosphorus and potassium content. However, crude protein (CP) content, measured as a percentage of dry matter, is closely related to the other parameters of nutritive value, and may be used as a sensitive indicator of overall herbage quality. The crude protein content of forage has also been shown to have an overriding influence on animal performance (Zemmelink, 1974).
The feeding value of forage, as measured in terms of crude protein content, varies with the season and the physiological status of the plant, to a degree that differs among the different grass species. Most perennials, for example, are capable of some regrowth almost throughout the year, but the decline in crude protein content with time and plant maturity is very rapid: in fact, crude protein content is rarely over 6% of total herbage for more than three months out of the year.
The fall in crude protein content with the age of the plant seems most pronounced for tall grasses, where the decline is about 1% for every 10 days of plant growth (Boudet, 1975b). For some species, crude protein content also depends on the season during which the plant grows: a plant is physiologically younger, and thus of higher nutritive value, if it grows during the dry season than a plant of the same age growing during the rains. During the rainy season, as long as residual soil moisture is available, crude protein content declines at the same rate as the decline in growth.
For many species, defoliation by grazing, cutting or burning stimulates regrowth of higher nutritive quality than that of continuous growth during the same period. For other species with strictly fixed life cycles, such as Hyparrhenia diplandra, aging continues regardless of defoliation and the feeding value is inadequate from August until the following rains.
For various rangelands around Shika, forage growth of 60 to 90 days was found to contain between 4 to 6% CP (Haggar, 1970; de Leeuw, private communication). During the late dry season, fairly high crude protein content has been recorded after burning or grazing. At Yanfolila in Mali, Boudet and Ellenberger (1971) found 7.2 ±0.4% CP after 20 to 30 days' regrowth on upland savanna and 9.5 ±1.6% CP on lowland savanna. After 55 to 65 days' regrowth on upland savanna at Shika, de Leeuw (private communication) found 5.3 ±0.2% CP, while Boudet (1975a) found 15.7 ±1.4% CP for Echinochloa/Oryza spp. on flood plains. Table 7 shows the percentages of crude protein found at different times of the year for upland shrub savanna under three different treatments.
Data on the crude protein content of total herbage may be unreliable in that they do not take into account the variation in crude protein between different parts of the plants. The figures in Table 8 show that the crude protein content of Andropogon gayanus is much higher in the top growth than in the rest of the plant. If this pattern also applies to other tall grass species, selectively grazing animals will obtain a diet of higher nutritive value during most of the rains and into the early part of the dry season than would be assumed from an assessment of the crude protein content of total herbage alone, As the dry season progresses, however, the crude protein content diminishes throughout the plant. Out of 273 grass samples analysed at Shika, 85% contained less than 4% CP, and 25% contained less than 2. 5% CP (Brinckman and de Leeuw 1975).
Table 7. Crude protein content under three different rangeland treatments at Shika, Nigeria (% DM)
|
|
1 May |
1 June |
1 July |
1 Aug |
1 Sept |
1 Oct |
1 Nov | |
|
Ungrazed, burned | ||||||||
|
|
Upland |
8.1 |
8.1 |
6.7 |
4.7 |
3.9 |
3.5 |
2.5 |
|
|
Old fallow a |
9.3 |
9.4 |
6.5 |
4.2 |
4.2 |
4.2 |
3.0 |
|
|
Fadama grassland |
9.9 |
10.4 |
6.5 |
4.1 |
3.7 |
3.0 |
2.6 |
|
Grazed, burned b | ||||||||
|
|
Green herbage |
- |
9.7 |
8.5 |
7.2 |
6.0 |
4.9 |
3.7 |
|
|
Total herbage |
- |
8.8 |
7.5 |
6.1 |
4.8 |
3.5 |
2.1 |
|
Grazed, not burned b | ||||||||
|
|
Green herbage |
- |
7.7 |
7.1 |
6.4 |
5.7 |
5.0 |
4.3 |
|
|
Total herbage |
- |
3. 7 |
3. 6 |
3. 5 |
3.4 |
3.2 |
3.1 |
a. Old fallow on deep colluvial soils with Setaria anceps, Hyparthelia dissoluta, Andropogon gayanus and Sporobolus pyramidalis as dominant species.b. Lenient wet-season grazing; green herbage refers to the upper 30 cm of the sward.
Source: de Leeuw (1971a).
Table 8. Crude protein content (CP) and in vitro digestibility (dig.) for different plant components taken from Andropogon gayanus pasture towards the end of the growing season
|
|
12 September |
3 October |
26 October |
|||
|
CP (%DM) |
dig. (%DM) |
CP (%DM) |
dig. (%DM) |
CP (%DM) |
dig. (%DM) |
|
|
Young leaves |
6.8 |
66.0 |
8.7 |
56.6 |
6.6 |
54.4 |
|
Old leaves |
2.1 |
49.5 |
3.1 |
29.2 |
3.4 |
39.4 |
|
Young stems |
3.1 |
64.1 |
5.4 |
56.8 |
4.6 |
42.6 |
|
Old stems |
1.2 |
53.6 |
1.5 |
44.9 |
1.1 |
32.5 |
|
Whole plant |
4.1 |
- |
3.7 |
- |
3.0 |
- |
Sources: For leaf and stem data, Haggar (1970); for whole plants, Haggar and Ahmed (1971).
The importance of woody vegetation for livestock production in the subhumid zone lies in its competition with grasses and its contribution of browse to animal nutrition, which can be particularly important at times when grazing is in short supply. In the more humid areas, most management systems follow a strategy of reducing the growth of trees by burning. Mechanical measures, such as felling or ringing, and chemical treatment have also been practiced occasionally. In general, it seems most economic to clear young plants before reforestation becomes advanced.
Browse is not considered as important a component of animal diets in the subhumid zone as in the Sudanian and Sahelian zones farther north because the number of woody species acceptable to stock is limited and grass is generally available. At Shika in Nigeria, only 11 of 42 shrub species identified were readily consumed, while 17 species, which accounted for over 50% of the shrub canopy, were avoided altogether (de Leeuw, 1979) Boudet (1975a) and Boudet and Ellenberger (1971) reported similar findings from other sites in the subhumid zone. They recorded only 10 shrub species which are acceptable to livestock.
The usefulness of browse lies mainly in the high feeding value it provides at seasons when other roughage sources are of low quality. From 41 samples of subhumid species, for example, two-thirds were found to have a crude protein content of over 10%. In general, browse contains more calcium, phosphorus, magnesium, and potassium than grasses (Brinckman and de Leeuw 1975). Piot (1970) discusses the value of a number of common subhumid species, including Daniella oliveri, Gardenia ternifolia, Hymenocardia acida, Lophira lanceolata, Piliostigma thonningi and Vitex madiensis. Leaves of these species were found to contain 8 to 17% CP and 0.2% P. Other species utilized as browse are Khaya senegalensis, Afzelia africana, Pterocarpus erinaceous and the fruits of Parkia clappertonia and Prosopis africana.
Actual data on the browsing behaviour of stock are scarce. Van Raay and de Leeuw (1974) found that late in the dry season (March to early July) 15 to 20% of the feeding time of semi-nomadic herds in the Sudanian zone was spent browsing, while sedentary herds in the subhumid zone spent only 3 to 6% of their feeding time on browse. Paddocked cattle at Shika browsed for 12 to 15% of their total feeding time in March and June, compared with only 4% in February when many shrubs are leafless or have only mature leaves. The pronounced seasonality of browsing behaviour is associated with the new flush of foliage which appears on the browse plants with the onset of the hot and humid weather preceeding the rains, a period when grass tends to be in short supply. In many parts of the northern subhumid zone, herdsmen climb large trees and lop off the branches for their animals to supplement the low-growing shrubs, coppicing shoots and fallen leaves and fruit that are readily accessible.
High acceptability and intake rates have been recorded for woody species in the Miombo woodlands of Zambia (Rees, 1974) and for thicket species on the Accra plains in Ghana (Rose Innes and Mabey, 1964). Considering the scarcity of data, further assessment is needed of the browse value of the woody species of the subhumid zone.
Grazing
The effects of grazing on pasture growth and species composition depend on the frequency and timing of defoliation, the selectivity of the grazing animals, and soil and climatic conditions. In general, certain species, such as Andropogon tectorum, tend to disappear under even relatively light grazing, to be replaced by species such as Brachiaria brizantha or Setaria sphacelata, without decreasing overall pasture productivity. As grazing increases, most of the palatable grasses are replaced by short-cycle annuals, such as Sporobolus and Eragrostis spp., or by species which the animals reject, such as those of the Malvacae and Solanaceae families. Extreme overgrazing is followed by denudation and soil erosion in the drier northern parts of the zone, or by bush encroachment in the south, forming dry Arthrosamanea forests, secondary forests or composite forests with Harungana.
Under traditional management, pastures in the subhumid zone are grazed primarily during the dry season. There is little information on changes in species composition or herbage yield following prolonged wet-season grazing. However, work at Shika suggests that cattle grazing rangeland with a variety of grass species tend to be rather unselective, avoiding few species completely (de Leeuw, 1979). Grazing behaviour appears to be governed by the age of the herbage, rather than by preference for particular species, leading eventually to a mosaic pattern of closely grazed and ungrazed patches (Cesar, 1975).
It is likely that changes in the species composition of pastures under heavy grazing are due to differences in the ability of particular species to withstand repeated defoliation, rather than to selective grazing behaviour per se. Grazing tends to favour the shorter grasses, such as Setaria, Bracharia and Paspalum, and those which mature early, such as Sporobolus pyramidalis and Hyperthelia dissoluta, rather than the tall and late-maturing Hyparrhenia spp. However, the resistance to grazing of the tall perennials varies considerably, depending on the timing and frequency of defoliation and the soil type. Piot and Rippstein (1975) observed that populations of H. rufa and H. filipendula decreased on shallow soils under grazing, but increased on deep colluvial soils. A similar pattern was observed by Cesar (1975) in Ivory Coast, while in Kafanchan and on the Jos plateau in Nigeria, Hyparrhenia species have been found to withstand prolonged heavy grazing and on protection to revert to tall stands similar in composition to ungrazed savanna.
Intensive grazing may lower overall pasture yields due to succession from tall to shorter grasses and also result in lower net annual herbage production among those grass species which remain. At Shika, it was found that growth 60 to 90 days after defoliation averaged 27% less than uninterrupted growth. On the Adamawa Plateau of Cameroon, frequent defoliation (every 20 to 30 days) resulted in cumulative yields which were 25% lower than yields with less frequent cutting every 60 to 80 days.
However, the rapid seasonal decline in the nutritive value of the herbage can be minimized by defoliation. Several trials have shown that yields of digestible nutrients, including protein, from grazed areas are higher than from cut areas; similarly the yield of crude protein from an area which had been grazed during the first part of the season was shown to be nearly as high as that of an ungrazed adjacent area. These results must be attributed to the smaller proportion of mature herbage on grazed areas and to the return of dung and urine. On a lightly stocked paddock the plant nutrients obtained from urine and dung are unlikely to increase herbage production, but on a heavily stocked paddock the return of nutrients (80% of the ingested nitrogen) is likely to be significant. The distribution of dung and urine on a paddock will always be uneven, however, and much will be lost by oxidation.
When animals are confined in paddocks, they tend to graze a limited area closely to which they frequently return. In this way, they provide themselves with a supply of comparatively nutritious herbage throughout the growing season. The grazed lawn can be composed of the original natural species which adopt a prostrate habit or of sub-climax species. In the ungrazed areas, a certain amount of the more palatable leafy material from the tussocks will be consumed. In appearance there can be very distinct grazed and ungrazed areas, or there can be frequent tussocks growing in a matrix of grazed grass. The proportion of grazed to ungrazed grasses is probably determined by the level of stocking.
The major constraint on most wet-season grazing systems is the lack of herbage of suitable quality during the dry season. Although repeated defoliation maintains the level of nutrients, there is only a limited amount of palatable material on offer at the beginning of the dry season. During this season, animals therefore need access to a larger area where they can graze selectively. This area could be tightly stocked with mature animals during the growing season, who would also have access to the tightly grazed lawns. For increased dry-season fodder production, sod seeding with legumes has also given promising results in a number of situations.
The detailed system of grazing (whether continuous, rotational or deferred) has to be adapted to the type of grassland and to its condition. However, a number of trials have indicated little difference in animal performance or in the botanical composition of pastures between different systems of grazing management, except for degraded areas or those under extremely high stocking levels. The stocking level affects liveweight gain per animal, liveweight gain per unit area and the pattern of selective grazing.
With many types of grassland, an increase in the number of livestock per unit area produces changes in gain per animal and in gain per unit area which are well known. However, comparison of the results obtained from wet-season grazing trials undertaken on medium to tall savanna grasses, particularly Andropogon, Hyparrhenia and similar genera, suggests that the regression between stocking level and gain per animal is not linear and that the highest gains are not obtained at the lightest stocking level. The regression for stocking level and gain per unit area is also different in the case of these grasslands. In Nigeria (Shika) unimproved shrub savanna stocked at a rate of one tropical livestock unit (250 kg liveweight) to 1. 6 to 2. 0 ha during the growing season gave better liveweight gains than either lighter or heavier stocking (de Leeuw, 1971a).
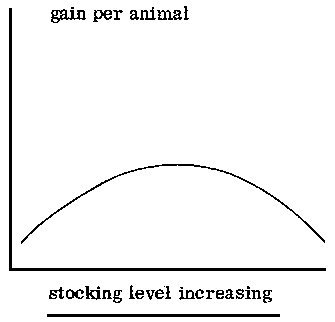
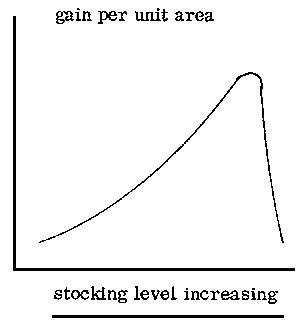
Because of the importance of protein for animal production, the supply of this nutrient could be used as a basis for estimating carrying capacity. A 250 kg animal requires 200 g digestible crude protein per day (160 g for maintenance, plus 18 g per 100 g liveweight gain) or 75 kg digestible crude protein per year. In fodders containing 7% crude protein, this level of digestible protein is approximately equivalent to 140 kg crude protein per year. The sustained yield of protein varies according to site and soil type from 60 kg to 200 kg per ha annually throughout the subhumid zone: if by frequent grazing the protein content is maintained at a satisfactory level and the total yield of protein is not affected by the frequent defoliation, then an area which provides 140 kg of crude protein will support an animal of 250 kg liveweight. If, in order to control shrub regrowth, a burn every third or fourth year is considered necessary, there will be a reduction in the carrying capacity by a third or a quarter, i. e., from 1:2 ha to 1:2. 6 ha. Carrying capacity will also be lower on Isoberlinia or Daniella woodlands than on open savanna. Where the tree canopy is 60% or greater, the carrying capacity will he nearly 1:6 ha.
A rotational grazing system was successfully implemented on relatively moist savanna at Ubangi, Zaire over a period of ten years with very little decrease in productivity or disturbance to the species composition of the grassland (IEMVT, 1974). The dominant grass species in this area is Hyparrhenia diplandra, which can produce 12 000 to 16 000 kg DM/ha annually when undisturbed. A five-paddock rotational grazing system was initiated with a very low stocking rate, designed to result in annual consumption of only 3 000 kg/ha, or 20 to 25% of total potential herbage production. Each paddock was burned once a year and grazed for a period of three months, beginning 100 days after burning. Analysis indicated a favourable crude protein content of about 10% of dry matter for the pastures under grazing.
The dry-season use of subhumid pastures by livestock that move north during the rains can be regarded as a rotational grazing system on a large scale. The shrewd use of a variety of fodder resources by transhumant producers is evidenced by the generally more prosperous appearance of their animals compared with sedentary herds in the same areas, though their freedom of movement is becoming increasingly limited by the spread of townships, roads and cultivated fields.
Evidence points to great resilience of pastures in the subhumid zone under transhumant dry-season and light wet-season grazing, but the rangeland does deteriorate seriously under prolonged heavy grazing and it is important to avoid stocking levels which are too close to the 'peril point' where the grasslands deteriorate rapidly. At Shika, for example, overgrazing has created large areas of stable 'terminal' grassland, consisting of low-yielding prostrate species such as Brachiaria stigmatisata, Tephrosia pedicellatum, Sporobolus pyramidalis and annual weeds. In parts of Ivory Coast, Cesar (1975) found that annual sedges and grasses replaced virtually all perennials under similar heavy grazing conditions.
Fire
During the dry season, savanna rangelands have traditionally been burned to clear tall grass and stimulate the replacement of old and unpalatable material with fresh herbage. This use of fire has long been a controversial issue. Van Raay (1974) condemns it out of hand, while others have considered it a useful management tool available to traditional farmers and pastoralists. In Nigeria, sedentary pastoralists often blame the use of fire on the transhumants, or bororo, but in fact they themselves make extensive use of the regrowth after fires to supplement the low protein levels available from the desiccated grazing late in the dry season. ILCA herd behaviour studies at Kachia near Kaduna indicate that animals spend over four hours a day grazing fire regrowth during the latter part of the dry season after crop residues have declined in importance.
The response of the savanna ecosystem to frequency and time of burning has received considerable attention, but most published studies have focussed on changes in the tree cover rather than on effects on grasses. This emphasis is due to the general opinion that frequent fires are essential to prevent bush encroachment and have little or no effect on the fire-tolerant grasses which dominate most savannas (Ramsey and Rose Innes, 1963; West, 1965). Afolayan (1977) has reported that under northern subhumid savanna conditions, late burning for two consecutive years can reduce the regeneration of tree seedlings by up to 80%, while in areas which are burnt early in the season or not burnt at all the number of seedlings increases.
It has generally been found that changes in grass species composition under burning are slow, and short-term effects are difficult to detect (de Leeuw, 1978a; Ramsay and Rose Innes, 1963). However, early burnt areas may tend to become dominated by annual grasses and fortes while late burnt areas tend to become dominated by perennials (Afolayan, 1977). West (1965) found that burning increased grass production in moist savanna areas, but reduced yields in more arid regions. This conclusion is supported by data from five locations in Nigeria presented in Table 9.
The effects of burning warrant further study in different parts of the subhumid zone. With increased cropping and sedentarization of livestock production, grazing pressure is likely to become greater and the role of fire and the results of burning will acquire an increased significance. Changes in crude protein content and dry matter yield should be investigated, as well as the impact on subsequent yields of early grazing of the regrowth.
Table 9. The effect of early and late burning on herbage yield at five locations in Nigeria (tonnes DM/ha/year)
|
|
Early Burning |
Late Burning |
Protected Not Burned |
|
Isoberlinia shrub savanna at Shika a |
2.20 |
2.23 |
2.18 |
|
Detarium savanna woodland at Borgu b |
2.09 |
3.62 |
1.56 |
|
Isoberlinia woodland at Borgu c |
1.93 |
3.51 |
1.83 |
|
Terminalia macroptera tree savanna at Borgu d |
4.11 |
6.89 |
3.78 |
|
Andropogon gayanus grass savanna e at Fashola |
17.83 |
16.37 |
15.27 |
a. Yield after six years of burning (1968-74) on upland site; early burning in late November or December, late burning in March or early April.b. Sites located in Borgu Game Reserve (10° N); mean yields over two years (1974-75) with years of burning unknown; early burning in late November or December, late burning in March or early April. This site on shallow soil over ironstone; dominant grass species are Loudetia flavida, Hyparrhenia involucrata and Andropogon pseudapricus.
c. As above, but with 66% canopy cover on deep colluvial soil; shade-tolerant grass species dominate.
d. As above, but on deep valley soils, seasonally flooded; dominant grasses tall Hyparrhenias, including H. rufa and H. smithiana.
e. Deep sandy soil with tall grasses dominant; burned one year (1969/70) on 22 December and 27 February; latitude 7 ° 55' N.
Sources: For Shika, de Leeuw (personal communication); for Borgu, Afolayan (1975 and private communication); for Fashola, Egunjobi (1973).
Cutting
The effects on pasture lands of grazing and successive cutting are similar but not identical. Cutting retards the development of inflorescent stems and may extend the period of growth by 30 to 45 days, but productivity declines, stabilizing at a lower level during the third year of any particular cutting strategy. In some cases, the species composition may also be slightly modified. The reduction in yields over a period of three years at Bouar in Central African Empire under a system of cutting three times a year is shown in Table 10.
Table 10. Green matter yields at five sites at Bouar (CAE) when cut three times a year (tonnes/ha/year)
|
|
Year 1 |
Year 2 |
Year 3 |
|
Forest savanna |
11.0 |
8.5 |
7.7 |
|
Savanna on concretions |
6.3 |
5.0 |
5.0 |
|
Savanna on colluvial soil |
16.1 |
14.9 |
13.0 |
|
Savanna on ferralitic soil |
19.5 |
14.5 |
11.8 |
|
Overgrazed savanna |
14.6 |
12.0 |
11.7 |
Source: Bille and Hédin (1968).
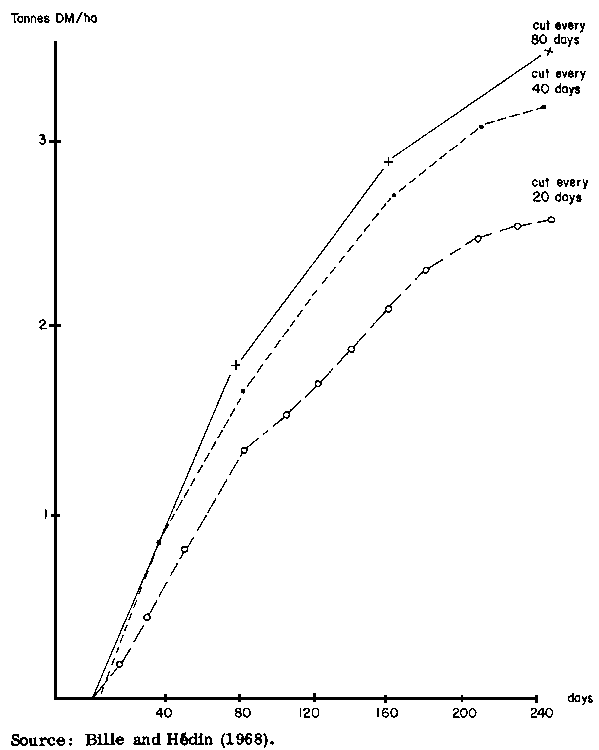
Productivity may decline at a faster rate if the utilization rate is increased. In another experiment carried out at Bouar (Bille and Hédin, 1968), dry matter yields were 3 400 kg/ha on pasture cut three times during the growing season, diminishing to 3 200 kg/ha when cut six times, 2 800 kg/ha when cut eight times and 2 500 kg/ha when cut 12 times, as shown in Figure 4. The time of defoliation also affects growth rates: Haggar (1970), working at Shika, found that Andropogon gayanus regrew at a daily rate of 11 kg/ha when cut at the vegetative stage before August 12, but regrew at a rate of only 5 kg/ha when cut later in the season.
Figure 4. Pasture productivity at Bouar (CAE) when cut 3, 6 and 12 times (tonnes DM/ha)
Sown Pasture
The advantages of pastures sown with high-yielding forage grasses and legumes lie mainly in the higher levels of forage production achieved, both in terms of quantity and quality. Leguminous species are particularly attractive because of their ability to convert and utilize atmospheric nitrogen. However, the high costs of pasture sowing and the level of management required in order to sustain productivity limit the adoption of this management strategy to certain specific situations. Comprehensive reviews of pasture sowing, covering species selection and adaptation, as well as the effects of fertilizer application and management on quality and yield, are found in Blair Rains (1963), Ruthenberg (1974), Dumas and Coulomb (1978) and de Leeuw and Agishi (1978).
Research in several countries has served to identify promising pasture grass and legume species and appropriate methods of sowing and maintaining improved pastures. Stylosanthes humilis has been identified as a very attractive legume for this purpose and has been sown successfully in Australia from aeroplanes without cultivation on burned pastures at the beginning of the growing season. Broadcasting without cultivation failed when it was tried at Shika in Nigeria because the soil formed a crust which the seeds or seedlings could not penetrate. Techniques of sowing without cultivation might be more successful on sandy soils, but competition from the perennial grasses must also be reduced and a fairly large population of legume seedlings must be established at the outset. Experience from elsewhere in West and Central Africa has shown that Stylosanthes seed can be established successfully by broadcasting from the ground when competition from existing grasses is reduced by prior burning or heavy grazing. Once a pasture is established with 20 to 30% Stylosanthes, the grazing animals can maintain it, at least for a few years on fairly open land, by ingesting and excreting the seed if they are allowed to graze when the seed is mature but still in the heads. No cultivation is required.
Though it has been shown possible to establish Stylosanthes, all large-scale trials in West Africa have failed eventually because the Stylosanthes has disappeared after a few years. On very fertile land, this has been due to competition from the grasses, while on overgrazed or eroded land the soil becomes compacted and cannot be penetrated by the Stylosanthes seeds. The plants are also destroyed by fire. In addition, Stylosanthes tends to become stemmy after two or three years and is subject to termite attack. Faster-maturing varieties are needed, which will produce seed before the pastures are burned at the end of the growing season, though usually such varieties also produce less green foliage during the dry season. The varieties which have shown promise so far include Stylosanthes humilis, an annual and S. guyanensis, a perennial. Cultivars of other species which require further evaluation in the area include S. hamata, Macroptilium atropurpureum and Centrosema pubescens.
The costs and returns of livestock production systems based primarily on sown pastures have been estimated by Ruthenberg (1974) and de Leeuw and Agishi (1978), Unfortunately these estimates are difficult to interpret or compare because of rapid inflationary trends in both input costs and beef prices. Ruthenberg concludes that breeding ranches grazing N'Dama cattle on pastures sown with Stylosanthes are on the whole not viable economically, but fattening and growing-out operations may break even if the livestock weight produced annually reaches 200 kg/ha. De Leeuw and Agishi compared beef production based on pastures sown to pure grasses and to grass-legume mixtures, as well as production based on unimproved rangelands. Supplementation with high-protein concentrates during periods of pasture deficiency was also considered, resulting in a comparison between six production systems, as shown in Table 11. The difference between gross income and costs for the six production systems ranges from US $ 22 to 198 per ha. The most attractive strategy in economic terms appears to be grazing on natural savanna with supplementation. The second most attractive strategy is year-round grazing on improved legume-based pasture.
An economic analysis was carried out recently of beef production and pasture improvement programmes initiated in the subhumid zone at Avetonou, Togo, comparing extensive and intensive ranching conditions (Doppler, 1979). Pasture improvement strategies considered include pastures planted with Panicum maximum - Centrosema pubescens, Stylosanthes guyanensis - Cynodon dactylon and natural pasture improved with C. dactylon. The primary objective of this analysis was to consider the economic incentives for pasture improvement from the viewpoint of a small-scale farmer and a livestock producer on a somewhat larger scale.
These combinations of grass and legume species were chosen to maximize fodder availability during the dry season. The natural pasture improved with Cynodon dactylon gave a relatively constant yield during the dry season, while the Stylosanthes guyanensis - C. dactylon combination provided good standing hay. The application of fertilizer in September at the end of the rains was also analysed. This approach appeared to he more expensive than the provision of alternative feeds during the dry season, such as grass silage, maize silage or concentrates. However, the use of fertilizer is nevertheless desirable because it tends to curie the growth of unpalatable weeds and shrubs.
All the systems of improved pasture management devised were found to be uneconomic in terms of beef production. An increase of cattle prices of approximately 50% would be necessary to obtain a positive rate of return. Alternatively, methods of establishing and maintaining improved pastures would have to be developed at substantially reduced costs.
In addition to poor profitability, the pasture improvement systems devised at Avetonou would require substantial financing to meet their high investment costs. Under the extensive ranching system tested, an area of 28 000 ha would he stocked with 6 012 cows and 301 bulls at a value of about US $ 1.1 million. Such a capital-intensive operation with low profitability would involve an unacceptably high level of risk, even if credit for the initial investment were available. It would also he very difficult to procure the required number of animals. Based on this analysis, it was concluded that, given present production techniques and cost-price relationships, no viable large-scale beef ranches could he expected to emerge from the private sector and any large-scale government ranches would have to rely on considerable injections of outside funds.
Livestock production based on sown pastures might be economically justified on well-managed grazing reserves if used to replace savanna grazing for selected classes of stock, such as pregnant and lactating cows or young calves, to provide night grazing in order to extend the daily grazing period, or to provide a reserve of fodder for emergencies such as uncontrolled fires or drought. Used in this way, improved pastures might help relieve one of the major constraints on livestock production under traditional pastoralist systems, which is the short grazing day combined with the long distances covered during the daily orbit (van Raay and de Leeuw, 1974). It is important, however, that sown pastures on a grazing reserve be kept under the strict control of family or cooperative units who are also responsible for maintenance costs.
Table 11. Input-output analysis of six livestock production systems in Nigeria
|
a. Wet Season - six months |
||||
|
|
Grass Pastures |
Legume Pastures |
Natural Savanna |
|
|
(no supplement) |
(no supplement) |
(supplement) |
(no supplement) |
|
|
Stocking rate (kg lwt/ha) |
800 |
300 |
150 |
150 |
|
Daily gain (kg) |
0.50 |
0.50 |
0.60 |
0.25 |
|
Yield for season (kg lwt/ha) |
288 |
108 |
65 |
27 |
|
b. Dry season - six months |
||||||
|
|
Legume |
Pastures |
Legume |
Pastures |
Natural |
Savanna |
|
supplement |
no supplement |
supplement |
no supplement |
supplement |
no supplement |
|
|
Stocking rate (kg lwt/ha) |
600 |
600 |
300 |
300 |
50 |
150 |
|
Daily gain (kg) |
0.50 |
0.25 |
0.50 |
0.25 |
0.25 |
- 0.04 |
|
Yield for season (kg lwt/ha) |
216 |
108 |
108 |
54 |
35 |
- 7.00 |
|
Annual yield (kg lwt/ha) |
252 a |
198 a |
216 b |
162 b |
100 b |
20 b |
|
Total Income (US$) |
408.20 |
316.80 |
345.60 |
259.20 |
160.00 |
32.00 |
|
Management costs (US$/year) |
152.00 |
152.00 |
,112.00 |
112.00 |
9.60 |
9.60 |
|
Cost of supplements (US$/year) |
68.80 |
- |
35.20 |
- |
28.80 |
|
|
Total costs |
220.80 |
152.00 |
147.20 |
112.00 |
38.40 |
9.60 |
|
Margin (US $/ha) |
182.40 |
164.80 |
198.40 |
147.20 |
121.60 |
22.40 |
|
Input/output ratio |
0.56 |
0.53 |
0.43 |
0.43 |
0.24 |
0.33 |
a. Stock grazed on grass pastures for six months and on legume pastures for six months, with both pastures left ungrazed for half the year; therefore the annual production is the mean of the two grazing periods.b. Legume pastures and savanna assumed to he grazed year round; thus annual production is the sum of the two grazing periods.
c. Monetary output calculated on the assumption that 1 kg liveweight = 1 Naira, and 1 Naira = US $ 1.60.
Source: de Leeuw and Agishi (1978)
Pasture sowing might also be an appropriate strategy for arable farms in the subhumid zone. In areas where pressure on the land is still low, fallow periods are relatively long - they may last from five to seven years. Natural fallows could he oversown with forage species, in particular with Stylosanthes or the last crop in the rotation could be undersown (Thomas, 1975; de Leeuw, personal communication). These fallow pastures would need to be properly managed under the control of individual farmers; they could be used in the same ways as sown pastures in grazing reserves.
Improved pastures could also be an important component of livestock production on breeding and fattening ranches. Experience with ranch development in the subhumid zone, on Mokwa Ranch in Nigeria (Hübl et al., 1974), suggests the advantages of a balanced combination of arable farming, including such crops as sorghum, maize and soya beans, followed by several years of grass-legume pastures (Panicum, Centrosema, Stylosanthes guyanensis) on the better soils and the less productive land left to natural savanna. This would provide an income from the sale of crops to offset clearing and cultivation costs as well as a diversity of roughages, including crop residues, to make possible a wide range of management and feeding strategies.
Fodder Crops
Several experimental fodder production projects have been carried out in the subhumid zone. High yields have often been reported, but it has not always been clear whether these yields were sustained over several years or how much fertilizer was required. Fodder trials carried out at Bobo Dioulasso in Upper Volta (Sikora et al., 1976), including annual and perennial grass species and legumes, gave daily productivity levels ranging from 160 to 190 kg DM/ha for the annuals, with a growing period of about 100 days, and from 180 to 210 kg DM/ha for the perennials, with a growing period of about 240 days. The legumes produced about 100 kg DM/ha daily. Results by plant species are presented in Table 12.
Table 12. Fodder productivity at Bobo Dioulasso, Upper Volta
|
Species |
Growing Period |
Annual DM Yield |
Daily DM Yield |
|
(days) |
(kg/ha) |
(kg/ha) |
|
|
Brachiaria ruziziensis |
243 |
49 811.00 |
204.98 |
|
Panicum maximum |
212 |
38 359.00 |
180.94 |
|
Stylosanthes graciis |
245 |
23 010.00 |
134.73 |
|
Centrosema pubescens |
244 |
16 982.00 |
69.60 |
|
Glycine wightii (javanica) |
245 |
15 844.00 |
64.83 |
|
Sorhum rio |
125 |
19 704.00 |
157.63 |
|
Canavalla eusiformis |
122 |
22 948.00 |
188.09 |
|
Stylosanthes humilis |
117 |
8 089.00 |
69.13 |
|
Cajanus cajan |
71 |
7 031.00 |
99.03 |
Source: Sikora et al. (1976).
A fodder trial carried out over a three-year period at Bouaké in Ivory Coast (Roberge et al., 1976) suggests that yields generally drop over the years: without fertilizer, productivity during the third year was only 47% of the level reached in the first year. Even with substantial fertilizer applications (up to 712 N. 675 K2O and 216 P2O5 annually), productivity had decreased by 25% by the third year. However annual production of Panicum maximum was maintained at 40 000 kg DM/ha under irrigation (though the level of water intake was not specified) and with fertilizer applications at the rate of at least 550 N. 200 P2O5, 810 K2O, 220 CaO and 290 MgO.
Use of Crop Residues
Crop residues are an important component of the forage supply of traditionally managed herds, particularly along the northern fringes of the subhumid zone and in the drier areas to the north. Residues comprise only a minor proportion of the total forage supply throughout the year because their availability is highly seasonal, but in the early part of the dry season after the harvests, from December to February, they have been shown to account for 75% of the total roughage supply of transhumant animals and 50% of the total supply available to sedentary herds. The contribution of crop residues from various sources has been estimated by van Raay and de Leeuw (1974) under transhumant and sedentary pastoral systems, as shown in Table 13.
Table 13. Sources of annual roughage intake as % of total daily grazing time under two pastoral management systems in northern Nigeria
|
|
Transhumant |
Sedentary |
|
|
Pastoralists |
Pastoralists |
||
|
Savanna Grazing |
|||
|
|
Upland savanna and fallows |
59 |
60 |
|
|
Lowland grass savanna (valleys) |
13 |
22 |
|
|
Browse |
9 |
4 |
|
Total |
81 |
86 |
|
|
Crop Residues |
|||
|
|
Sorghum |
12 |
10 |
|
|
Millet |
1 |
|
|
|
Groundnuts |
3 |
2 |
|
|
Cowpeas |
- |
2 |
|
|
Cotton |
3 |
|
|
Total |
19 |
14 |
|
Source: Derived from van Raay and de Leeuw (1974).
In the Kano Close Settled Zone, an intensive agricultural area in northern Nigeria, crop residues account for over 80% of the total year-round forage supply. The period during which these residues are available is extended in this area mainly through fodder conservation and storage (Hendy, 1977). The annual production of crop residues has been estimated by several authors (Rose Innes and Mansfield, 1976; van Raay and de Leeuw, 1970), but general estimates of yield levels based on particular situations are of limited value due to the wide variety of crop mixtures planted, levels of fertilizers and other inputs used. However, the estimates provided by Hendy (1977) for the intensive farming system in the Kano Close Settled Zone suggest yield levels which are likely to obtain in other similar areas of the subhumid zone. These estimates are presented in Table 14.
Table 14. Average roughage resources on farms in the Kano Close Settled Zone of northern Nigeria
|
|
% of Cultivated |
DM Yield |
CP Content |
(% DM) |
% of Total |
|
Land under each crop |
(t/ha) |
During Growing Season |
After Harvest |
Digestible CP Produced |
|
|
Sorghum leaves |
40 |
1.6 |
6.0 |
3.0 |
14 |
|
Millet leaves |
27 |
1.0 |
6.0 |
3.0 |
3 |
|
Groundnut haulms |
16 |
1.5 |
12.7 |
7.7 |
46 |
|
Cowpea haulms |
13 |
1.6 |
17.5 |
12.5 |
36 |
|
Other crops |
4 |
1.5 |
|
4.0 |
1 |
a. The high CP content of sorghum and millet refers to leaves stripped during the growing season; for legumes, the CP content is based on early harvested and well cured hay.Source: Derived from Hendy (1977).
Hendy estimates annual production of all residues at 1 400 kg DM/ha, of which 40% is legume hay, mostly of good quality. Because 75% of the land in this area is under crops, producing substantial quantities of roughage, a high stocking level of two tropical livestock units (equivalent to 250 kg each) can be maintained per ha, composed mainly of sheep, goats and donkeys.
Throughout most of the subhumid zone, the traditional utilization of crop residues has been limited to the grazing of sorghum stover by transhumant herds and local sheep and goats. Moreover, there is a gradual shift from north to south in the zone, from cereal to root crop farming with a resulting decrease in available residues. Sorghum retains an important role in areas where a yam-sorghum cropping pattern predominates, but is replaced by maize in areas where yam-cassava production is more important. Forage - producing legumes are also much less important towards the south.
The utilization of crop residues implies a mixed production system combining arable farming and livestock. Yet development planning in some countries of the region, notably in Nigeria, is focusing, at least in part, on large-scale mechanized cereal production with the role of crop residues for livestock feeding likely to be minimal. In this context, research is needed on the relative profitability of mono-cropping in various areas compared with the possibilities for mixed production systems including a livestock component.
![]()
![]()
![]()
{kind=link}