![]()
![]()
![]()
* Based on papers presented by K J R MacLennan, S N H Putt, B K Na'isa and S M Touré and discussions led by H E Jahnke.
The problem
Livestock production strategies in the tsetse-infested zone
Techniques for the control or prevention of animal trypanosomiasis
Economic assessment of trypanosomiasis eradication and control programmes
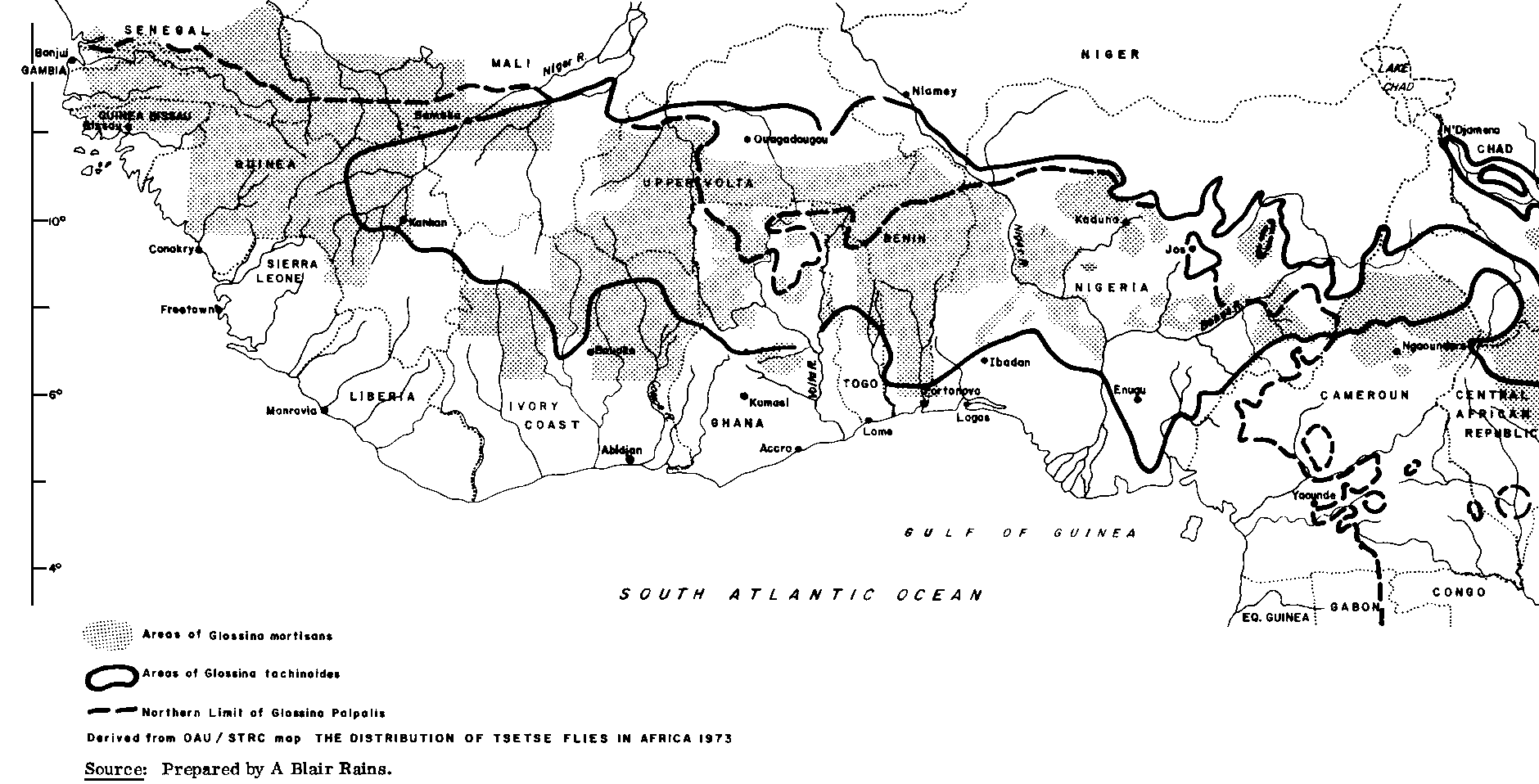
The entire subhumid zone as defined for the purpose of this report, is infested, or potentially infested, with tsetse flies, except for a few small high-altitude areas. These flies are the biological vectors which transmit several species of trypanosomes, causing human sleeping sickness and trypanosomiasis in domestic livestock. Although there are other constraints and other disease problems in the subhumid zone, tsetse-transmitted trypanosomiasis, accompanied in some areas by onchocerciasis, must be viewed as one of the major impediments to intensified livestock production. Tsetse flies infest approximately one-third of the African continent, or roughly 10 million km2, out of which 7 million km2 could profitably be used for livestock production (see Figure 6). Based on projected stocking rates, the potential production of about 120 million head of cattle is foregone (FAO, 1977c). Some of the best watered and most fertile land in West Africa is affected; a large proportion could be used at once by pastoralist livestock producers with practically no other development inputs if it were cleared of tsetse (MacLennan, 1979).
Figure 6. Tsetse distribution in West and Central Africa
The Tsetse Population
Tsetse flies of the subhumid zone are usually classified taxonomically into three types: the forest species, primarily Glossina fusca, G. nigrofusca and G. medicorum; the riverine species, mainly G. palpalis and G. tachinoides; and the savanna species, primarily G. morsitans and G. longipalpis. A more complete list of tsetse species is given in Table 15. Although these groups are described in terms of their most usual habitats, riverine tsetse may also be found in woodlands and other non-riparian habitats in the higher-rainfall areas and the forest species are occasionally found in patches of relict forest which occur frequently in parts of the derived savannas of the subhumid zone (Keay, 1953, 1959; OAU/STRC, 1973). Sleeping sickness is transmitted by riverine tsetse, while all three types transmit animal trypanosomiasis to a greater or lesser extent.
There are important local seasonal fluctuations in tsetse populations in areas which have pronounced wet and dry seasons. During the dry season, particularly if it is hot, there is a general regression of tsetse flies. This is also encouraged by traditional grass burning among hunters, farmers and pastoralists of the subhumid zone, which reduces the tree and shrub cover that forms the fly's natural habitat.
Long-term fluctuations in tsetse populations, particularly of the savanna type, also occur over wider areas (Ford, 1971). There was a widespread retreat of savanna tsetse towards the end of the last century when the host populations were sharply reduced by rinderpest panzootics, though much of the area freed of tsetse at that time has since been reinfested. Small-scale local reductions have also occurred as a result of increased cultivation and hush clearing or government tsetse eradication projects (Aiyedun and Amodu, 1976; Lester, 1945). However, in many areas the introduction of domestic livestock, notably cattle and pigs, has resulted in expansion of the tsetse population due to the new source of food. The habitat can also become more suitable for tsetse flies whenever overgrazing leads to increased bush infestation. The introduction of certain types of arable agriculture can also lead to increased tsetse infestation and outbreaks of trypanosomiasis. Habitats for the riverine species are found in mango groves and areas where guava, cashew, cocoa, banana and sugarcane are cultivated, while trees and hedges planted around family compounds and coco-yam plots may also harbour tsetse colonies (Baldry, 1969a; Challier, 1973; Touré, 1974).
Table 15. Species of tsetse (Glossina)
|
1.
|
Savanna tsetse: morsitans group, sub-genus Glossina |
|
Savanna: G. morsitans, a G. swynnertoni, G. longipalpis a |
|
|
Savanna and thicket: G. pallidipes |
|
|
Evergreen thicket: G. austeni |
|
|
2. |
Riverine tsetse: palpalis group, sub-genus Nemorhina, G. palpalis, ab G. fuscipes, a G. martinii, G. quanzensis, G. caliginea, a G. pallicera, ab G. tachinoides, a |
|
3
|
Forest tsetse: fusca group, sub-genus Austenina |
|
Rain forest: G. tabaniformis, a G. nigrofusca, G. haningtoni, G. nashi |
|
|
Forest edge: G. fusca, a G. medicorum, ac G. fuscipleuris, a G. schwetzi, G. severini, G. vanhoofi |
|
|
4. |
Others G. brevipalpis, G. longipennis |
a. Present in the subhumid zone.
b. Challier suggests that G. palpalis and G. pallicera should also be included as forest species.
c. Challier suggests that G. medicorum is found in riverine areas in the savanna zone.
Source: MacLennan (1979).
Since 1952, G. morsitans advances in the central and eastern parts of northern Nigeria have covered at least 25 900 km2. Similarly in central Cameroon, an advance of G. morsitans has occupied about 20 860 km2 of valuable agricultural land since 1950 and, unless halted, is likely to extend considerably further. Consequences for the rural economy have been serious: transhumant livestock producers are forced to move away, contributing to increased grazing pressure in the remaining tsetse-free zone, and the settled agricultural population is impoverished.
These instances of reinvasion and orientation to a man-made environment point to the fact that tsetse eradication in the subhumid zone is not likely to be accomplished by increased settlement and agricultural activity alone without specific eradication programmes. The problem is likely to assume increasing prominence in the future.
Sleeping Sickness
In humans, the prevalent form of trypanosomiasis in West Africa is Gambian sleeping sickness, caused by Trypanosoma gambiense, which is transmitted by infected riverine tsetse. Savanna tsetse have never been directly implicated in an outbreak of Gambian sleeping sickness, though their possible role has been suggested by several authors (Duggan, 1962; Ford, 1971).
Not all riverine tsetse populations harbour trypanosomes pathogenic to humans. The disease must be introduced to a fly community through an external source which, it is generally thought, can only be an infected person. There is circumstantial evidence indicating an alternative reservoir host (e. g. Molyneux, 1973; Scott in Mulligan, 1970), but none has so far been positively identified in the field. Once the disease is introduced into a community, it is transmitted from person to person through the infected flies. The chief factor governing the incidence of the disease is the intimacy of contact between tsetse flies and the human population: epidemics have occurred with very low overall densities of tsetse infestation, but where the localized communities of flies have been living and feeding in close contact with the human hosts (Nash, 1944, 1958 and in Mulligan, 1970).
Gambian sleeping sickness can occur in many forms, ranging from a chronic debilitating condition to a more acute illness. Both frequently proceed to neurological involvement, causing the syndrome which gives the disease its name and usually results in death. With the chronic form of the disease, the principal cause of death eventually is often another disease contracted because of the patient's lowered resistance (Lester, 1945).
Sleeping sickness is found principally in agrarian communities or among groups of people who work or live near rivers and come in close contact with tsetse flies. The pattern of the disease varies considerably in different areas, existing at low levels or in epidemic proportions in its chronic or acute form (Duggan, 1962). The substantial spread of this disease in the 1920s and 1930s gave rise to the first tsetse eradication programmes in West and Central Africa. Due to vector eradication, combined with continuous surveillance, chemoprophylaxis and treatment, the disease at the moment is successfully suppressed in the subhumid zone, though scattered episodes continue to occur. At present, 50 to 100 cases are recorded in Nigeria every year.
In spite of the present recession, however, continous vigilance is required to keep sleeping sickness at a low level (Thomson, 1968). As already mentioned, disease problems will not automatically be eliminated with increased human settlement: in many parts of the subhumid zone, sleeping sickness has even become an urban problem (Baldry, 1969a, 1970; Challier, 1973; Touré, 1974). For example, riverine tsetse have found a suitable habitat in Bamako, Mali in the extensive mango plantations near the town, and sleeping sickness can only be kept under control by continuous efforts. In Ivory Coast and Cameroon, cocoa and coffee plantations established in the moister parts of the subhumid zone have led to major sleeping sickness outbreaks which have only been brought under control with considerable difficulty and at substantial cost. Another disturbing factor is the recent suspicion that the T. gambiense strain, which causes sleeping sickness in West Africa, may be closely related to T. rhodiense, the strain responsible for the virulent East African form of the disease, and that the rhodiense strain may arise as a mutant from the general T. brucei population present in game animals.
Animal Trypanosomiasis
In natural populations, tsetse flies transmit trypanosomiasis between wildlife hosts in what McLennan refers to as the sylvatic cycle. Transmission to domestic animals occurs when livestock are introduced into an area where the disease is present. The disease can also be transmitted between domestic animals, or strains can be transferred back to wild animals and later reintroduced into domestic herds. E these strains have become drug resistant, they retain their resistance throughout the cycle of transmission (Gray and Roberts, 1971a, 1971b).
Unlike the human disease, animal trypanosomiasis is carried by nearly all tsetse species. However, the actual infection rate and severity of the disease is often related to the type of tsetse and the density of the population, as tsetse species vary in terms of feeding preference, infection rate and transmission capability (MacLennan, 1979). Low levels of infestation by the riverine species cause low levels of infection, even among highly susceptible exotic livestock breeds, while larger numbers of the same riverine species give rise to more severe trypanosomiasis problems (Jordan, 1961; Page, 1959). Generally, flies of the savanna group cause very serious disease problems, even when they are present in small numbers (Kirkby, 1963; Leeflang, 1975). Some members of the forest group also undoubtedly feed on cattle when they have the opportunity and can be effective carriers of the disease.
All mammalian species of domestic livestock are susceptible to trypanosomiasis, but their susceptibility to different strains of trypanosome varies, as shown in Table 16. Cattle, for instance, are highly susceptible to T. congolense and T. vivax, while pigs are subject to fatal outbreaks caused by T. simiae. There is also a wide variance in pathogenicity among different strains of trypanosome species. Within livestock species, susceptibility also varies significantly between breeds: for example, the N'Dama and West African Shorthorn cattle breeds and the West African Dwarf breeds of pigs, horses, sheep and goats show a substantial degree of tolerance to the disease, compared with Zebu or European stock. Stress factors are also important determinants of susceptibility. Losses can take the form of rapid mortality or chronic debilitation which depresses production potential in terms of growth, meat, milk, fertility and draught power.
Table 16. Species of pathogenic trypanosomes and susceptible hosts a
|
|
Humans |
Cattle |
Sheep |
Goats |
Pigs |
|
T. gambiense |
+++ |
- |
- |
- |
- |
|
T. rhodiense |
+++ |
- |
- |
- |
- |
|
T. brucei |
- |
+ |
+ |
++ |
± |
|
T. congolense |
- |
+++ |
++ |
++ |
± |
|
T. equiperdum |
- |
+ |
+ |
+ |
± |
|
T. evansi |
- |
++ |
+ |
+ |
± |
|
T. simiae |
- |
- |
+ |
++ |
+++ |
|
T. suis |
- |
- |
- |
- |
++ |
|
T. uniforme |
- |
++ |
++ |
++ |
- |
|
T. vivax |
- |
+++ |
++ |
++ |
- |
a. Pluses and minuses indicate degree of susceptibility.
Source: Stephen, in Mulligan (1970).
Transhumant and Semi-Settled Production
The traditional patterns of transhumance enable most livestock to avoid tsetse-infested areas at times of greatest risk. During the dry season, when grazing and water resources in the north are depleted, the tsetse population in the infested areas retreats into the moister localities. Transhumant herdsmen are able to bring their animals slowly down into the subhumid zone, grazing on crop residues and manuring the farmers' fields along the way, to utilize the well-watered fodder resources of the subhumid grasslands. These herdsmen use their knowledge of the areas of greatest tsetse infestation to avoid trypanosomiasis risk, though this is not always possible and disease outbreaks occur which can be serious. The situation is particularly difficult in times of drought when movement options are restricted. At the beginning of the rains, the transhumants avoid further risk by returning to the northern areas as soon as water and pasture are sufficient. Thus, during the rainy season in the six northern states of Nigeria, 86% of the cattle are found on only 46% of the land, corresponding with the area which is free of tsetse or only lightly infested.
Farmers and pastoralists can also be found settled in areas of light riverine infestation. These producers accept limited levels of trypanosomiasis among their herds, and they may also move their animals short distances during the season of peak infestation. In Nigeria, levels of infection in these areas have also been reduced because transhumant herds which come down seasonally from the north are now less likely to he infected due to tsetse eradication programmes in their home areas. If serious disease incidence does occur, pastoralists are usually able to move away to safer locations. The White Fulani Zebu, which form the basis for most of these herds, also appear to have a small degree of tolerance to light trypanosome risk arising from low-level riverine tsetse infestation (MacLennan, 1979).
When cattle are trekked down to southern markets along traditional trade routes, areas of infestation often cannot be avoided and tsetse populations have built up in some places based on the constant food source available from the cattle passing through. This problem may be alleviated with the present general trend away from trekking in favour of transportation by lorry. In Nigeria, for example, in 1974, the control posts at Wawa, Kontagora and Yelwa reported a total of 65 185 cattle passing through on the hoof and 15 051 in lorries: by 1978, 80 911 were reported in lorries and only 5 939 on the hoof (Nigeria, Tsetse and Trypanosomiasis Division, 1978).
Production Based on Trypanotolerant Breeds
Small-scale livestock production is also carried out in the southern part of the subhumid zone based on the trypanotolerant N'Dama and West African Shorthorn cattle and West African Dwarf sheep and goats (Pagot in IEMVT, 1974). Large-scale ranches have also been operated successfully in Congo and Zaire based on introduced trypanotolerant cattle.
Trypanotolerance has been defined by an FAO expert consultation (1976) as:
a hereditary biological property which enables certain breeds, or certain individuals, to live normally in a naturally infected environment while carrying pathogenic trypanosomes without themselves showing any clinical signs of the disease.
Tolerance is not absolute: it can be broken down by stress factors such as exposure to other diseases, heavy parasite burdens, parturition, lactation, inadequate nutrition or hard work (Touré, 1977). There is evidence that trypanotolerant cattle may be more susceptible than other breeds to certain diseases, notably rinderpest, but they appear to have greater resistance to others, such as dermatophilosis. When infected with trypanosomiasis, trypanotolerant animals develop anaemia, and several authors have claimed that their growth and reproductive performance are lowered. However, at some N'Dama multiplication centres where high abortion and calf mortality rates were initially attributed to trypanosomiasis infection, closer investigation revealed that high levels of helminthiasis were also a factor.
The hereditary nature of trypanotolerance was demonstrated by Esuruoso (1977), based on observations of a trypanotolerant Muturu herd kept in Nigeria. A group of Muturu were kept in a tsetse-free area and later housed continuously in fly-proof accommodation in the rain forest, to prevent any trypanosomiasis infection. The second generation of this group was experimentally infected with T. vivax, along with a group of Zebu cattle. The clinical course of the disease and the haematological changes in both types of cattle were similar, but the Muturu survived the infection for a period twice as long as the Zebu, indicating an innate, though partial, ability to tolerate the disease.
One immediate constraint on the increased utilization of trypanotolerant livestock is the general shortage of breeding stock, however. The multiplication of trypanotolerant breeding stock needs to be carried out on a priority basis. Whenever trypanotolerant stock are introduced into a new environment, prophylactic drugs should be administered for a short period while the animals adjust to new trypanosome strains and levels of infestation. This strategy has been followed by extension programmes in several countries with good results.
Intensive Production with Susceptible Breeds
Livestock susceptible to trypanosomiasis are sometimes kept in the tsetse-infested zone under chemoprophylactic regimes. Such schemes have generally been undertaken in areas of dense settlement or on ranches where careful control of drug use is possible and where there has been a sufficient modification of the vegetation and fauna to make the area less inviting for resident tsetse populations. However, it has not proven possible to modify the environment sufficiently to make it totally uninfestable, since it is simply technically, financially and administratively impossible to exert comprehensive control of land use on the scale that would be required.
The problems are diverse. For one thing, all game reserves, national parks and forest reserves in the subhumid zone are almost certainly tsetse infested, and in several cases tsetse populations have advanced through these areas into pastoral grazing zones, agricultural areas or government stock farms (Wilson, 1958). For this reason, reforestation projects, even in the Sahel, should anticipate the need for tsetse-control measures. In Nigeria north of Bauchi, for example, an integrated agricultural project was initiated for settled Fulani who practiced stall feeding and used oxen for ploughing. A G. morsitans population which occupied a nearby forest reserve spread to the pastures, causing a major trypanosomiasis outbreak which could not be contained by trypanocidal drug treatment alone. Several examples of this type of problem have occurred elsewhere in the subhumid zone.
A second type of problem has occurred in a number of areas apparently free of savanna tsetse infestation, but where the introduction of cattle has resulted in an incidence of trypanosomiasis. In these cases, the disease problem brought to light tsetse populations, originally too small to be detected, which were able to escalate rapidly with the new food supply. It appears that a penumbra of flies exists around a tsetse-infested area which is very difficult to detect. Given this penumbra, no susceptible livestock are safe within several miles of a primary tsetse focus. Either a nearby tsetse population may advance into the livestock production area or a small number of local flies may increase rapidly in response to the new source of food (Leeflang, 1975; MacLennan in IEMVT, 1974). In either case, satellite tsetse populations can become established and an intense transmission cycle rapidly escalates, causing serious losses in spite of treatment with trypanocidal drugs.
An example of this situation arose at the government livestock farm at Shika, in Nigeria, at the northern limit of the subhumid zone. Initial surveys before the farm was established did not reveal any G. morsitans infestation. The farm functioned successfully for a number of years, and then T. vivax was discovered as the cause of high abortion rates. This signalled the outbreak of a serious trypanosomiasis problem. Though few flies were ever detected, they were later found breeding on the farm in an atypical habitat (Kirkby, 1963).
Another example arose with the beef fattening operation which was set up at Mokwa Ranch, also in Nigeria, in an area of heavy tsetse infestation. The herd was maintained under prophylactic treatment and animals were not grazed within 0.8 km of the ranch boundary. However, because of poor pasture management, trees and shrubs encroached on portions of the ranch, and a reinfestation of tsetse led to a serious incidence of trypanosomiasis when the ranch veterinarian went on leave and treatment was relaxed. Similar advances of the tsetse -population have also led to serious disease problems along traditional cattle trek routes (Baldry, 1969b; Ferguson, 1964b; Riordan, 1971; Yesufu and Mshelbwala, 1973).
Domestic pigs are sometimes kept in tsetse-infested areas. They appear to tolerate relatively mild infections of T. congolense and T. brucei, but if T. simai is introduced through infected flies, a fatal outbreak is likely, even though the overall tsetse population is extremely small (Baldry, 1964).
If draught oxen are introduced onto farms in the tsetse-infested zone, heavy losses can result. Where small numbers of oxen are widely dispersed over a number of holdings, prompt treatment is difficult to organize, leading in some cases to drug resistance, anaemia and myocarditis. However, in other parts of the subhumid zone, for instance in northern Benin, the use of draught oxen has increased substantially without tsetse control measures and without serious losses. Much depends on the degree of risk from tsetse, the breed of animal, and husbandry and stress factors.
This variety of experience points to the need for more precise information on tsetse populations and a clearer demarcation of the areas where control measures are necessary.
When considering the various approaches to a trypanosomiasis problem in a particular area, a basic distinction must be made between eradication and control objectives. In a control situation, measures are taken against the disease and/or the vectors in a defined area within a zone of infestation. While the initial costs may be limited control involves vector and disease surveillance on a continuous basis, with a substantial technical and supervisory commitment. An eradication programme, by contrast, involves the elimination over a larger area of the tsetse population which carries the disease. Once the vectors are eliminated, a tsetse-free status is maintained. Either an entire tsetse belt is eliminated or a large area is treated which can subsequently be protected with a minimum of financial and technical resources. A special issue is raised in many cases where the area of infestation crosses national boundaries. In these instances, the most effective approach to eradication would can for international cooperation on a regional basis.
After a substantial eradication operation has been carried out, the surveillance and supervision requirements become less over time, limited to the vulnerable perimeter of the eradication zone if the entire local tsetse belt has not been eliminated. However, this type of operation can only be undertaken in situations where it is technically feasible and within the limits of available funds. Such a programme must be based on detailed knowledge of the local tsetse distribution and habitat relationships and requires a substantial field capability.
The nature of tsetse distribution and reproduction makes eradication possible over large areas of the drier savanna zone. Present levels of knowledge and technology are now being developed to make possible the extension of eradication programmes into the moister areas. Where eradication is still not technically or financially feasible, control measures must be accepted, though production based on susceptible livestock with prophylactic treatment is usually not feasible except in areas of relatively low risk and where treatment can be rigidly controlled, as the experiences already described demonstrate. Production based on trypanotolerant livestock is another possibility.
Of the range of techniques available or being developed, some are more appropriate for eradication operations and some more appropriate for control. Many techniques are used for both types of operation, applied on different scales and in different infestation circumstances.
Habitat Modification
Bush Clearing. Bush clearing was the main technique used to control riverine tsetse populations in the campaigns to suppress sleeping sickness before the advent of modern insecticides. Methods vary from complete removal of the woody and shrub vegetation, known as sheer clearing, to selective clearing of only that vegetation known to be important to the survival of a particular tsetse species. Selective clearing becomes more difficult in humid environments where the tree cover is dense (Keay, 1953) and the fly population less dependent on particular types of trees or shrubs (Davies, 1964; MacLennan, 1967; MacLennan and Na'isa 1971).
Early clearing campaigns were carried out by hand, but more recently mechanized methods have been introduced involving the use of bulldozers and drag chains. Clearing operations carried out with motorized saws and arboricides have been disappointing. In all cases, the major problems have been the physical control of rapid regenerative growth and the costs of both the initial clearing and the control of regrowth, which requires continuous supervision and countermeasures.
The technical problems and, more importantly, the high costs of these operations are only justified in particular circumstances, such as when limited clearing is required along rivers to control sleeping sickness or when a barrier strip is cleared along the perimeter of a tsetse eradication area or an intensive livestock development project. Because of the cost, clearing is usually limited to a one-mile-wide (1. 6 km) strip, usually accompanied by periodic applications of insecticide along troth sides or only along the infested side of the cleared area.
Large-scale clearing might be justified when coupled with a programme of intensive land use, involving the development of arable or mixed farming or intensive ranching with improved fodder production and appropriate stocking rates to prevent the reinvasion of trees and shrubs. However, in Africa this approach has seldom proved economic in terms of beef production, although, in Latin America for example, it can be economically attractive provided the land to be cleared has sufficiently high agricultural potential. Even in such cases, however, the area covered is likely to be small in terms of overall livestock production needs and the size of the tsetse-infested zone.
Wildlife Elimination. Control of tsetse populations by eliminating their wildlife hosts has been attempted mainly in eastern Africa, and especially in Uganda (Wood, 1968). In Nigeria, along the Jama'ara Katagum River, warthog shooting and trapping were encouraged in the early 1960s as a measure to control the G. morsitans population (Davies, 1964). The operation was apparently successful, though mainly through the use of insecticides. Game eviction is likely to be used in future only in exceptional circumstances, due to organizational difficulties and the pressure of world environmental opinion.
Housing. Under zero-grazing systems, susceptible livestock can be protected from trypanosomiasis risk by keeping them in fly-proof shelters. This might be practicable and economically feasible in the case of intensive dairying or pig production in the tsetse-infested zone (MacLennan, 1979).
Chemotherapy and Prophylaxis
A small number of curative and prophylactic drugs have been developed and are widely used in Africa for the treatment of animal trypanosomiasis. The same drugs are often used for both curative and prophylactic treatment: the distinction lies in the quantity of the dose and sometimes in the form of administration. A drug may be administered curatively at three to five times the level of the prophylactic dosage.
Drugs most often used for prophylaxis and treatment are listed in Tables 17 and 18. Novidium was the first to be developed, but by about 1964 strains of trypanosomes were appearing which were resistant to this drug. Berenil was introduced at that time, but again resistance developed quickly. A few other products have been introduced since then, but drug resistance and cross-resistance are becoming an increasingly serious problem. In spite of this, the commercial pharmaceutical firms have not developed any new drugs for some time. This type of research and testing work involves substantial investments, and the potential African market is perhaps not sufficiently attractive to justify, from a strictly commercial point of view, the level of expenditure required. The Tsetse and Trypanosomiasis Division of the Nigerian Federal Livestock Department has been actively involved in testing trypanocidal drugs and is carrying out research on the mechanisms of drug resistance.
Table 17. Trypanocidal drugs used for prophylactic treatment of domestic livestock
|
Name of Drug |
Trade Name |
Trypanosomes Affected |
Length of Protection |
Treatment of Relapses |
|
Isometamidium chloride |
Samorin |
T. vivax |
2-4 months |
Diminazene |
|
Trypamidium |
T. congolense |
|
|
|
|
|
T. brucei |
|
|
|
|
Pyrithidium bromide |
Prothidium |
T. vivax |
2- 4 months |
Diminazene |
|
|
T. congolense |
|
Isometamidium |
|
|
Quinapyramine chloride and sulfate |
Antrycide |
T. brucei |
2 months |
Suramin |
|
Prosalt |
T. evansi |
|
|
|
|
Suramin-quinapyramine complex |
|
T. simiae |
young pigs: 3 months |
Isometamidium |
|
T. evansi |
adult pigs: 6 months |
Isometamidium |
||
|
|
|
Diminazene |
Source: FAO (1979a).
Several livestock production projects have been carried out in tsetse-infested areas under chemoprophylactic drug regimes. This type of operation has been successful in areas where the level of tsetse infestation is sufficiently low, but it is not recommended under high infestation conditions. With medium infestation, it has been found that a high level of management and veterinary and entomological supervision is essential to avoid major disease outbreaks or a general increase in drug-resistance. The timing and dosage of the drug regime must be strictly adhered to and blood samples must be examined on a regular basis, requiring a degree of simple laboratory support. It has been the usual experience that block treatment of an entire herd at appropriate intervals gives better results than waiting for an individual positive diagnosis, because in cases where prophylaxis is waining infection may occur before it becomes detectable by blood slide examination (MacLennan, 1979). This situation greatly favours the development of drug-resistant strains. A different drug should be introduced when curative treatment becomes necessary, but even this may not always be effective because by now cross-resistance to several trypanocides has become a widespread phenomenon. In some eases, reinfection has been detected only 10 days after treatment with a prophylactic.
Table 18. Trypanocidal drugs used for curative treatment of domestic livestock
|
Name of Drug |
Trade Name |
Trypanosomes Affected |
Treatment of Relapses |
|
Homidium bromide
|
Ethidium
|
T. vivax |
Diminazene |
|
T. congolense |
Isometamidium |
||
|
Homidium chloride
|
Novidium
|
T. vivax |
Diminazene |
|
T. congolense |
Isometamidium |
||
|
Diminazene aceturate
|
Berenil
|
T. congolense |
Isometamidium |
|
T. vivax |
|
||
|
(T. brucei) |
|
||
|
(T. evansi) |
|
||
|
Quinapyramine sulfate
|
Antrycide sulfate
|
T. congolense |
Isometamidium |
|
T. vivax |
|
||
|
T. brucei |
|
||
|
T. evansi |
|
||
|
Isometamidium chloride
|
Samorin Trypamidium
|
T. vivax |
Diminazene |
|
T. congolense |
|
||
|
(T. brucei) |
|
||
|
Suramin
|
|
T. evansi |
Quinapyramine |
|
T. brucei |
|
Source: FAO (1979a).
The costs of carrying out such programmes and the risks involved often mean that livestock projects based on chemoprophylactic treatment become uneconomic It has been estimated that the total costs of the management, veterinary supervision and services required may amount to 10 times the cost of the drugs alone, and, perhaps more importantly, the type of personnel and the supervision required is not widely available in developing countries.
The problems of treating transhumant herds or draught animals widely dispersed on small farms are even greater. In many countries, the demand for trypanocidal drugs far exceeds the capacity of the official distribution system, resulting in the widespread illicit use of drugs. In such a situation, it is impossible to ensure that the drugs are properly administered, and the use of drugs at sub-curative levels or the use of the wrong drug or dosage for the particular situation has contributed to the sharp increase in drug-resistance. Breakdown resulting in serious disease episodes is not unusual, even on projects with high levels of management.
The use of trypanocidal drugs has been more successful in situations where treatment has been of limited duration, for example when cattle must be trekked through tsetse hefts on the way to slaughter. These herds usually come from areas which are free of infection and may therefore have -little or no resistance, particularly under the stress of travel; consequently they often suffer heavy losses. Since normally only a short period of protection is required, a properly supervised chemoprophylactic programme can be of great benefit in reducing losses. A similar argument in favour of prophylactic treatment can be made in the ease of feedlot operations.
Insecticide Spraying
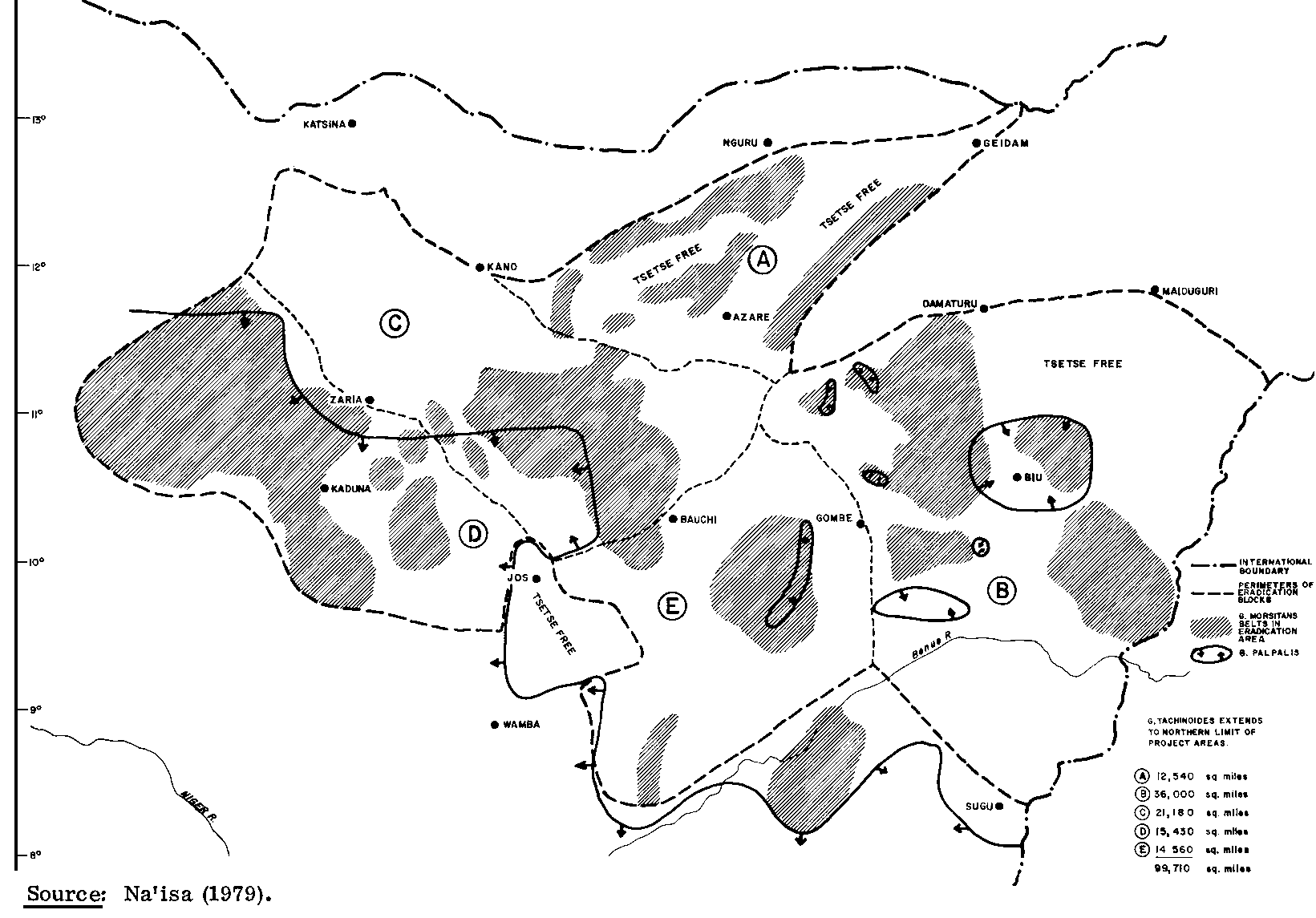
Spraying programmes involve placing lethal doses of insecticide on trees or shrubs where tsetse flies rest or in the atmosphere where they fly. A number of different methods have been developed, and extensive eradication programmes based on spraying have been carried out in Nigeria, as shown in Figure 7. As of 1977-78, a total of 204 802 km2 had been cleared in that country, including 190 232 km2 by ground spraying units, 12 366 km2 by helicopter and 2 204 km by fixed-wing aircraft.
Figure 7. Tsetse eradication blocks in Nigeria
In the francophone countries, programmes have usually been more modest so far and in most cases are still at the experimental stage, for example in Senegal, Mali, Chad and the Central African Empire (Finelle, 1964; Grovel et al., 1970; Tibayrenc, 1977; Touré, 1973). Some of this work is particularly interesting because it has focused on the precise distribution of resting riverine species, allowing very selective insecticidal treatment. For example, a radio-marking technique has been described for detecting flies resting during the day time Laveissière, 1976; Laveissière et al., 1978; Challier 1973; Bois et al., 1977; Challier et al., 1978b).
Spraying programmes may be classified according to the method of application, for instance from a team walking along the ground with knapsack sprayers, from helicopters or from fixed-wing aircraft. The duration of the effects of the spray also vary, from transient or non-persistent to persistent or residual formulations. In addition, the areas treated vary: with selective spraying, only certain patches or groups of vegetation are sprayed and only those parts of the plants which the flies are known to frequent, but with non-selective spraying wide swaths of vegetation are covered. A variety of equipment and insecticides is available, suited to different circumstances. These are described in detail in FAO's Insecticides and application equipment for tsetse control (1977e).
Ground Spraying with Residual Insecticides. This method involves a single placement of persistent insecticide on the preferred tsetse resting places (Davies, 1964; MacLennan, 1967). The insecticide remains lethal for about eight weeks until au the pupae in the ground have hatched. To achieve this degree of persistence, relatively heavy rates of application are required, but the application is very selective, based on detailed knowledge of the ecology of the target species in each particular area, as preferred resting places vary considerably among tsetse species and in different climatic zones (MacLennan, 1967; MacLennan and Cook, 1972; Scholtz et al., 1976).
Spraying is done- at the end of the dry season when the flies' habitat is somewhat reduced. Swaths may be sprayed on either side of streams or at intervals of about 150 m in dense woodland. In savanna areas, small habitat features may be sprayed, such as individual trees or thickets. Large teams of labourers are required, with supervisory personnel, and a network of camps, tracks and access paths must be constructed. Although the necessary level of organizational and logistical support is considerable, this type of operation brings secondary benefits to the local community in terms of employment and increased access to remote areas indeed, a high proportion of total expenditure remains in local hands.
This approach has proven highly successful in eradicating both savanna and riverine tsetse species in the drier areas to the north of the subhumid zone. It has also been successful, though more difficult and more expensive, in the less densely wooded parts of the subhumid zone itself (see Keay, 1953). In moister environments with denser vegetation, the tsetse habitats become more diffuse and difficult to recognize and a larger proportion of the vegetation must be sprayed to achieve eradication (MacLennan and Na'isa, 1971). The method is technically and financially efficient if only 10% of the vegetation has to be sprayed, but if more than about 18% of an area must be covered, costs escalate and there is greater concern about the environmental side effects. Moister climatic conditions also reduce the period of effective persistence of the insecticide. In areas of dense settlement in the moister areas, however, the tsetse's resting places may be reduced by bush clearing, farming and intensive grazing. In such circumstances, selective ground spraying may be an effective means of eradicating tsetse, though it will probably be more useful as a control technique.
Helicopter Spraying with Residual Insecticides. Persistent insecticide is sprayed selectively on preferred tsetse habitats from small, low-flying helicopters. As with ground spraying, detailed knowledge of the local tsetse habitats is important, and considerable logistical support is needed to clear a number of small temporary landing places, to service and maintain the helicopters in the field, to carry out tsetse surveys both before and after spraying and to provide protection against reinvasion.
This approach has been made feasible by the development of ultra low volume insecticide formulations and spray equipment specially designed for use on helicopters (Lee, 1977; Spielberger et al., 1977). Droplet size and the rotation speeds of the atomizers used for distributing the spray are of crucial importance because deposits must be large enough to be lethal, numerous enough to give adequate coverage and heavy enough to penetrate the tree canopy from the air. This technique is also very sensitive to meteorological conditions. As with ground spraying, it is more effective during the dry season; in addition, windy conditions must be avoided. A condition of temperature inversion must exist, where ground temperatures are lower than those at, say, 7 m, because, when the lower air is warmed by the sun, turbulence develops which can carry small droplets upwards and away from the target areas. Spraying is thus restricted to short periods around dawn and dusk.
The helicopter is flown one or two metres above the tree canopy at speeds of 32 to 40 kph over savanna country. The speed is reduced over forests to give a down draught strong enough to disperse the insecticide below the trees. Insecticide is sprayed along linear swaths oriented according to preferred tsetse habitats, as with ground spraying. If the operation proceeds smoothly in areas where only 10% of the vegetation must be covered, a unit of two helicopters can spray about 3 600 km2 in one season. Though much less logistical support is needed than for ground spraying, the cost of the helicopter can more than offset the savings on labour and transport.
Compared with ground spraying, larger amounts of insecticides are required since spraying within the swaths is much less selective. As a result, the impact on non-target organisms is substantially greater (Koeman et al., 1978). The costs and environmental effects also rise considerably if more than a certain percentage of the area must be sprayed: as with ground spraying, the upper limit is probably about 18%. This means that, although the technique has proven effective in eradicating riverine and savanna tsetse populations in the northern subhumid and Sudanian zones, it becomes less effective in more humid regions.
Non-Residual Insecticide Spraying from Fixed-Wing Aircraft. This technique shows promise for application in moister areas, though as yet only limited trials have been conducted in the subhumid zone (Lee, 1977). Exceedingly small doses (6-12 g/ha) of non-residual insecticide are sprayed into the air over an entire area, killing all adult tsetse as they fly. Applications are repeated, usually about five or six times at 10- to 19-day intervals over a period of about two months. This should be sufficient to kill all the flies emerging from pupae before they have a chance to deposit pupae themselves. Knowledge of the timing of the tsetse reproductive cycle is of crucial importance, as this varies in each local area according to prevailing temperatures (Glascow in Mulligan, 1970; Phelps and Burrows, 1969).
Usually, a rectangular area is sprayed along flight lines about 300 metres apart, oriented transversely to the prevailing wind. The aircraft flies about 7 m above the tree canopy, closely controlled by personnel on the ground. A very high standard of navigation is required, calling for an experienced pilot and sophisticated navigation and emission control equipment. Meteorological conditions are even more critical than with helicopter spraying, but over flat terrain spraying can be carried out at night. However, if only a single aircraft is available, the operation becomes highly vulnerable to delays resulting from equipment failure, and if a delay between application goes beyond the period of the tsetse reproductive cycle, the whole series of operations must be repeated.
As with helicopter spraying, the formulation of the insecticide and the rate of emission are crucially important. An attractive feature of this technique is the low dosage of insecticide required, which keeps costs and environmental side-effects at a minimum.
Insecticide Formulations and Environmental Effects. Over the years, only DDT and dieldrin have proven effective in eradicating tsetse by ground spraying operations (Lycklama and Nijeholt, 1965; MacLennan, 1967). Of the two, DDT has extremely long-lasting effects on tsetse populations in drier areas and is less toxic to humans, but dieldrin is more effective in the denser habitats of the more humid regions (Koeman et al., 1971). For helicopter spraying, special ultra low volume formulations of dieldrin and endosulphan are used, and endosulphan concentrates are used for fixed-wing spraying operations.
In Nigeria, a number of studies have been carried out on the environmental effects of tsetse eradication operations (Koeman and Pennings, 1970; Davies, 1964; Koeman et al., 1971). Ground and helicopter spraying with dieldrin and endosulphan causes heavy mortality rates among fish, frogs, reptiles, insectivorous birds, fruit bats and one type of monkey at the time of spraying. However, tsetse elimination is largely achieved by a single application, and the readiness with which these species return to an area which has been sprayed depends on the distance of the nearest neighbouring population and whether the habitat is substantially changed, for example by land clearing and the introduction of cultivation, after the eradication campaign (FAO, 1977f; Koeman et al., 1971). Apart from some elevated residual levels among frogs, studies conducted one year after spraying found very little insecticide remaining, indicating that any serious long-term accumulation problem is unlikely. In areas visited nine years after ground spraying with dieldrin and DDT, all bird and fish species previously affected had returned in substantial numbers (MacLennan, 1973).
These studies indicate that ground spraying, which is more discriminative, has fewer side-effects than helicopter spraying (Koeman et al., 1978), and that dieldrin is more damaging to birds and mammals, while fish and reptiles are more susceptible to endosulphan. It is possible to reduce the environmental effects of spraying in many situations by using a combination of methods and insecticides: this approach is already being followed in Nigeria. Environmental effects can also be minimized by spraying particularly vulnerable habitats, such as fringe forests around streams, from the ground, rather than from helicopters.
Studies have been carried out elsewhere in Africa on the environmental effects of non-residual endosulphan spraying from fixed-wing aircraft (Wood and Turner, 1975). It was reported that no serious side-effects occurred, but there is little information on the effects of this technique in the subhumid zone. The very low levels of insecticide used suggest that environmental side-effects may not be too severe
Over the years, a number of other insecticide formulations have been tested, including organophosphorous and carbamate compounds, but they have proved less effective in eradicating tsetse flies than the insecticides already in use and have shown similar adverse environmental effects. Recently, small-scale tests have been conducted involving ground and helicopter spraying with synthetic pyrethroids, and the results have been encouraging (Challier et al., 1978a). The different pyrethroid compounds tested proved highly toxic to tsetse with relatively few side-effects, though crustaceans and some other insects were affected. Extremely low dosage levels are required. Further testing is now being carried out by the Nigerian Tsetse and Trypanosomiasis Division to identify improved formulations with reduced repellent effects and increased persistence.
Other Techniques
Sterile Male Release This technique involves releasing large numbers of male tsetse flies which have been sterilized by radiation or exposure to chemicals. Because the female tsetse apparently mate only once, if the sterile males reach them before the males in the natural population they will not reproduce.
This method may prove effective in eliminating small residual tsetse focii which are often difficult to detect or to eradicate by conventional methods (Jordan, 1976). If large populations are involved, the majority must be eliminated first by spraying with non-residual insecticides. To be effective, the sterilized males have to outnumber the natural male population by at least 3 to 1.
At present, breeding colonies to produce a sufficient number of sterilized males are expensive and difficult to maintain. If more than one species of tsetse is to be controlled, separate breeding colonies have to be established for each species. Though there are also some doubts as to whether artificially reared males can compete successfully with natural males in terms of survival and mating ability, cultured males from irradiated pupae have been shown in some tests to be fully competitive. Trials have been carried out at Bobo Dioulasso in Upper Volta and at Tanga in Tanzania. At Bobo Dioulasso, production of 20 000 sterilized males a month is envisaged (Itard, 1974), whilst in Tanzania 6 500 sterile male G. morsitans were released weekly over an area of 100 km2. Consequent on the development of an effective chemical attractant, it is possible to envisage large-scale field trapping and automatic sterilization and release which would circumvent the difficulties encountered with colonization.
Attractants. The effectiveness of all tsetse eradication methods would be greatly enhanced if local fly populations could be brought together in one place through the use of long-range olfactory attractants. More accurate survey and detection would be possible, spraying operations could he concentrated in smaller areas, and other methods, such as the release of sterile males, would become economically feasible. Work is being carried out in this field and, though no readily available practical attractant has been developed as yet, some promising advances have been made.
Techniques Still at the Experimental Stage. Growth regulating hormones have been identified which affect, for example, the maturation of the- tsetse puparium (Mouchet, 1974). Research is also being conducted on tsetse control through the use of larvacides (Jordan and Trewern, 1978), sterile hybrids, tsetse parasites and viral and bacterial diseases which affect the flies (Laird, 1977). Cattle dipping or spraying against ticks has been shown to have a limited effect on tsetse.
The search for a trypanosomiasis vaccine is being carried out at the International Laboratory for Research on Animal Diseases (ILRAD) in Nairobi. Although this would he a most attractive solution; it is made difficult by the number and variability of trypanosome antigens. However, recent work at ILRAD and elsewhere has opened up several promising avenues of research in this area, giving an indication that an immunological solution is not as impossible as was once thought (Murray et al., 1979).
In the francophone countries, a biconical tsetse trap has been designed which should not he too difficult or expensive to construct. The most successful design developed so far is a device with a blue lower cone (Challier and Laveissière 1973). In Ivory Coast, blue cloth screens impregnated with an insecticide, decamethrine, have been used experimentally on cocoa and coffee plantations, and preliminary results are promising. In areas with a small local tsetse population, the regular use of traps could have a significant effect because the flies are not very prolific.
It is difficult to quantify the economic losses attributable to animal trypanosomiasis, though these are no doubt substantial. It is likely that most of the direct economic losses are incurred by pastoralists who are forced to bring their herds through infested areas, mainly on transhumance or on the way to market, or who suffer a low but constant level of losses in areas of light riverine infestation. Devastating losses can also occur when tsetse of the savanna type advance into an area where livestock are kept by local producers or on government stock farms.
Trypanosomiasis also has a significant effect on the viability of livestock development projects. Many of these projects are economically marginal, and the considerable expenses of surveillance, management and treatment of infected animals and/or vector control can be enough to make a livestock project show a loss.
More important than actual mortality rates or production losses, however, is the loss of production potential of the abundant fodder resources in areas which are not fully utilized because livestock cannot be maintained on a permanent basis. The presence of savanna tsetse not only prevents the utilization of valuable fodder resources directly, but when the flies infest strategic locations, such as around water sources, grazing in extensive adjacent areas becomes hazardous.
These underutilized land resources in the subhumid zone are becoming increasingly valuable as the human population expands, resulting in a sharply increasing demand for land and meat and milk products. The relatively sparsely populated areas of the subhumid zone have assumed particular importance as a destination for immigrant settlers from the overpopulated humid zone to the south and Sudanian zone to the north.
At the same time, population pressure and the extension of cultivation in the Sahelian and Sudanian regions have pushed traditional livestock producers more deeply into the tsetse-infested areas for longer periods of time, resulting in greater animal losses.
In Nigeria, the problem of competition for land became acute in the 1950s and led to the initiation of the present anti-tsetse programme. Tsetse clearance made possible the year-round utilization of grazing areas in the subhumid zone, relieving the pressure on ecologically fragile areas farther north which were being increasingly utilized for arable farming. It allowed many of the pastoralists to settle in subhumid areas and made it possible for the size of the national herd to remain stable in spite of increased pressure on grazing resources in large parts of the country and the effects of the Sahelian droughts.
Although crop production can take place in tsetse-infested areas without the introduction of livestock, the land resources in such a situation are often not exploited to their full potential. For one thing, the production increases obtainable from the use of animal draught power are foregone, and, to increase crop production substantially, mechanized farming must be introduced from the outset. Livestock and crop production tend to be mutually dependent. Farmers need livestock to supply them with meat and milk products and to renew the fertility of their fields with manure, while pastoralists need farmers as a market outlet and to provide them with both staple foods and crop residues which are an important source of forage for their livestock. Livestock also put to productive use the otherwise neglected grazing resources of the farming areas. Such an interrelationship between arable farming and livestock production, whether it occurs on one mixed holding or involves a relationship between two groups of people, makes possible a more balanced and more productive use of the land. This is only feasible when an area is at least largely free of trypanosomiasis.
Costs of Eradication and Control Programmes
Information on costs is only available for the two tsetse eradication methods which have been used in West Africa on a fairly wide scale, the selective application of long-lasting insecticides by ground spraying and by helicopter. From the records of the Tsetse and Trypanosomiasis Division of the Nigerian Federal Livestock Department, it is possible to estimate the costs of ground spraying at N 150 to 200 (US $ 240 to 320) per km2 while costs of helicopter spraying are about 250 (US $ 400) per km2. These costs can be broken down as shown in Table 19, though, as explained previously, they will vary according to ecological conditions and the scale of the operation.
Table 19. Breakdown of costs of tsetse eradication programmes
|
|
Ground Spraying |
Helicopter Spraying |
|
Labour |
38% |
3% |
|
Insecticides |
18% |
36% |
|
Flying time |
- |
51% |
|
Junior Staff |
23% |
3% |
|
Senior Staff |
16% |
2% |
|
Vehiclesa |
2% |
2% |
|
Depreciation |
3% |
3% |
|
Total |
100% |
100% |
a. Imported items; the total foreign exchange component for ground spraying is 23%, for helicopter spraying 92%.Source: Putt (1979).
To the costs of the original operation must be added those of a certain amount of respraying, say 20%, to ensure complete eradication. This brings the total costs of ground spraying up to N 180 to 240 (US $ 283 to 384) per km2 and the total costs of helicopter spraying up to N 300 (US$ 480) per km2. Then to these basic costs must be added a share of the other costs of running the Tsetse and Trypanosomiasis Division. These cover headquarters costs, off-season activities such as surveys, salaries of permanent staff outside the spraying periods and replacement costs for buildings, vehicles and equipment. These costs are relatively low for the helicopter unit because it is run with a small supervisory staff only, amounting to about 50% of the basic expenditure, bringing the total costs up to N 375 (US $ 600) per km2. By contrast, the ground spray units employ considerable numbers of people and consequently require more substantial support services. The costs of this support probably doubles and may even treble the basic cost, bringing the total to somewhere on the order of N 360 to N 500 (US$ 576 to 800) per km2. These figures are only provisional: actual costs vary according to ecological conditions, as explained previously, and the scale of the operation.
From these figures it would appear that helicopter spraying is an attractive approach. However, 90% of the variable cost of the helicopter unit must be paid in foreign exchange. Furthermore, the figures presented here are based on the official exchange rate: if the real cost in naira of the helicopter operation were calculated, this would be almost double the figures quoted.
On the other hand, it has been estimated that when a livestock development project is carried out in a tsetse-infested area, the full costs of eradication contribute only 2 to 4% of the total investment costs, if these are limited to the provision of water supplies and animal health services. Where further investments are envisaged, the costs of tsetse eradication will only comprise 1 to 2% of the total investment (Jordan et al., 1977).
No precise information is available on the costs of tsetse control, as compared with eradication. It can be assumed that the initial costs are significantly lower, but the costs of control recur continuously over an indefinite period. Similarly, the costs of prophylaxis and treatment with trypanocidal drugs have often been estimated based on the cost of the drugs alone, whereas information from Botswana suggests that the full costs of a treatment programme, including personnel and equipment, can range up to ten times the cost of the drugs administered (Negrin and MacLennan, 1977).
Benefits of Tsetse Eradication
The principal effects of tsetse eradication have been the spontaneous changes in land use which have resulted. Yet because these changes are also due to other factors, it is difficult to quantify to what extent they are attributable to tsetse eradication alone Some of the changes which can be expressed in numerical terms might be:
- increased population in the area due to immigration,
- increased agricultural production due to the introduction of animal draught power,
- increased carrying capacity, in terms of the additional fodder made available,
- increased livestock numbers and livestock production, and
- halting or reversing large-scale tsetse advances.
Since the benefits accrue in the years after the costs for tsetse eradication are incurred, they have to be discounted. Both the discount rate used and the assumptions about the time span over which incremental production is attributable to tsetse eradication are as important in determining the economics of an eradication programme as the costs and the benefits accruing in any one year. For this reason, it is difficult and misleading to generalize. Each situation requires careful analysis and elaboration of all the implicit and explicit assumptions involved. For example, if the cost of eradication is N 500 per km and the discount rate is 12% (at present value of 1 per annum: 5-year factor of 3. 6950), then the minimum annual level of benefits which must accrue in five years to cover the costs of eradication are about N 140/km2 annually to break even.
![]()
![]()
![]()
{kind=link}
{kind=link}