![]()
![]()
![]()
J. E. Halver
University of Washington
Seattle, Washington
1.1 Thiamine
1.2 Riboflavin
1.3 Pyridoxine
1.4 Pantothenic Acid
1.5 Niacin
1.6 Biotin
1.7 Folic Acid (Folacin)
1.8 Vitamin B12
1.9 Ascorbic Acid
1.10 Inositol
1.11 Choline
1.12 p-Aminobenzoic Acid
1.13 Lipoic Acid
Water-soluble vitamins include eight well-recognized members of the vitamin B complex: thiamine, riboflavin, pyridoxine, pantothenic acid, niacin, biotin, folic acid and vitamin B12; the water-soluble essential nutritional factors: choline, inositol, ascorbic acid; and vitamins with less-defined activity for fish: p-aminobenzoic acid, lipoic acid and citrin. The first eight, though required only in small amounts in the diet, play major roles in growth, physiology and metabolism. Choline, inositol and ascorbic acid are required in appreciable quantities in the diet and sometimes are not referred to as vitamins but as major dietary nutrients. A typical vitamin test diet for fish control over the water-soluble vitamins is listed in Table 1.
Table 1 - Water-Soluble Vitamin Test Diet H-440 1/
|
Complete test diet (g) |
Vitamin mix Mineral mix (mg) (mg) |
|||||
|
Vitamin-free casein |
38 |
Thiamine HCl |
5 |
USP XII No. 2 |
plus |
|
|
Gelatin |
12 |
Riboflavin |
20 |
AlCl3 |
15 |
|
|
Corn oil |
6 |
Pyridoxine HCl |
5 |
ZnSO4 |
300 |
|
|
Cod liver oil |
3 |
Choline chloride |
500 |
CuCl |
10 |
|
|
White dextrin |
28 |
Nicotinic acid |
75 |
MnSO4 |
80 |
|
|
a -Cellulose mixture 2/ |
9 |
Calcium |
50 |
KI |
15 |
|
|
|
|
|
pantothenate |
|
|
|
|
a -Cellulose |
8 |
Inositol |
200 |
CaCl2 |
100 |
|
|
Vitamins |
|
Biotin |
0.5 |
per 100 g of salt mixture |
|
|
|
|
|
Folic acid |
1.5 |
|
|
|
|
Mineral mix |
4 |
L-Ascorbic acid |
100 |
|
|
|
|
Water |
200 |
Vitamin B12 4/ |
0.01 |
|
|
|
|
Total diet as fed |
300 |
Menadione (K) |
4 |
|
|
|
|
|
|
a -Tocopherol acetate (E) 3/ |
40 |
|
|
|
1/ Diet preparation: Dissolve gelatin in cold water. Heat with stirring on water bath to 80°C. Remove from heat. Add with stirring - dextrin, casein, minerals, oils, and vitamins as temperature decreases. Mix well to 40°C. Pour into containers; move to refrigerator to harden. Remove from trays and store in sealed containers in refrigerator until used. Consistency of diet adjusted by amount of water in final mix and length and strength of beating.2/ Delete two parts a -cellulose and add two parts CMC for preliminary feeding
3/ Dissolve a -tocopherol in oil mix
4/ Add vitamin B12 in water during final mixing
Experimental avian polyneuritis, a condition resembling beriberi, was produced by Eijkman in Java in 1886 and the anti-beriberi factor was crystallized and named "vitamine". The term "thiamine" was introduced when the chemical nature of the factor was established. Thiamine was isolated from rice polishings in 1926 and synthesized in 1936.
1.1.1 Chemistry
Thiamine hydrochloride

Thiamine hydrochloride is a water-soluble, colourless, monoclinic, crystalline compound. It is comparatively stable to dry heat but is rapidly broken down in neutral or alkaline solutions and is split by sulphites into constituent pyrimidine and thiazole moieties. It has a characteristic yeast-like odour. The pyrimide ring is relatively stable, but the thiazole ring is easily opened by hydrolysis. Several derivatives are stable to heat and appear to be more completely soluble in weak alkaline solutions than thiamine itself and still show biological activity in animals. These derivatives include thiamine propyl disulphide, benzoylthiamine disulphide, dibenzoylthiamine, and benzoylthiamine monophosphate. Both thiamine hydrochloride and thiamine mononitrate have been successfully used as the active vitamin in test diets for fish nutrition studies.
1.1.2 Positive functions
Thiamine functions in all cells as the coenzyme cocarboxylase, thiamine pyrophosphate, which participates in the oxidative decarboxylation of pyruvic acid to acetate for entry into the tricarboxylic acid (TCA) cycle.
Thiamine pyrophosphate

Thiamine pyrophosphate is also a coenzyme of the transketolase system by which direct oxidation of glucose occurs in the cytoplasm of cells via the pentose phosphate pathway. Erythrocyte (RBC) levels of metabolites of this system have been used to indicate thiamine status in experimental animals, including salmon and trout.
Thiamine is essential for good appetite, normal digestion, growth and fertility. It is needed for normal functioning of the nervous tissue and the requirement is determined by the caloric density of the diet.
1.1.3 Deficiency syndrome
Deficiency signs in salmonids include impaired carbohydrate metabolism, nervous disorders, poor appetite, poor growth, and increased sensitivity to shock.
A trunk-winding symptom in eels has been reported, together with haemorrhage at the base of the fins. Skin congestion and subcutaneous haemorrhage occurs in carp fed thiamine-deficient diets. Typical symptoms observed in salmonids, carp and catfish are listed in Table 2. Thiamine deficiency has also been reported in marine flatfish started on clam neck diets stored long enough for thiaminase present to hydrolyze the thiamine in the ration. Typical nervous paralysis occurred with rapid mortality from physical shock.
Table 2 Vitamin Deficiency Syndromes
|
Vitamin |
Symptoms in salmon, trout, carp, catfish |
|
Thiamine |
Poor appetite, muscle atrophy, convulsions, instability and loss of equilibrium, oedema, poor growth |
|
Riboflavin |
Corneal vascularization, cloudy lens, haemorrhagic eyes - photophobia, dim vision, incoordination, abnormal pigmentation of iris, striated constrictions of abdominal wall, dark colouration, poor appetite, anaemia, poor growth |
|
Pyridoxine |
Nervous disorders, epileptiform fits, hyperirritability, ataxia, anaemia, loss of appetite, oedema of peritoneal cavity, colourless serous fluid, rapid postmortem rigor mortis, rapid and gasping breathing, flexing of opercles |
|
Pantothenic acid |
Clubbed gills, prostration, loss of appetite, necrosis and scarring, cellular atrophy, gill exudate, sluggishness, poor growth |
|
Inositol |
Poor growth, distended stomach, increased gastric emptying time, skin lesions |
|
Biotin |
Loss of appetite, lesions in colon, colouration, muscle atrophy, spastic convulsions, fragmentation of erythrocytes, skin lesions, poor growth |
|
Folic acid |
Poor growth, lethargy, fragility of caudal fin, dark colouration, macrocytic anaemia |
|
Choline |
Poor growth, poor food conversion, haemorrhagic kidney and intestine |
|
Nicotinic acid |
Loss of appetite, lesions in colon, jerky or difficult motion, weakness, oedema of stomach and colon, muscle spasms while resting, poor growth |
|
Vitamin B12 |
Poor appetite, low haemoglobin, fragmention of erythrocytes, macrocytic anaemia |
|
Ascorbic acid |
Scoliosis, lordosis, impaired collagen formation, altered cartilage, eye lesions, haemorrhagic skin, liver, kidney, intestine, and muscle |
|
p-Aminobenzoic acid |
No abnormal indication in growth appetite, mortality |
1.1.4 Requirements
In determining dietary thiamine requirements of fish, some considerations must be placed upon the composition of dietary ingredients in the ration. Fat content of the diet may affect not only caloric intake but also the thiamine requirement because cocarboxylase participates in the oxidation of that through a-ketoglutarate. Therefore, fish on a high fat diet and low thiamine intake might take longer to develop deficiencies and will give an erroneous requirement. Table 3 lists the thiamine requirements of selected fish species.
1.1.5 Sources and protection
Common sources for thiamine are dried peas, beans, cereal bran, and dried yeast. Fresh glandular tissue is also a good source for thiamine and other members of the vitamin B water-soluble complex, but is seldom used in modern commercial fish diets.
Thiamine can be easily lost by holding diet ingredients too long in storage or by preparing the diet under slightly alkaline conditions or in the presence of sulphide. Wet or frozen diets pose a problem because moisture content increases the chance of enzymatic (thiaminase) hydrolysis and subsequent destruction of thiamine. Obviously, wet or moist diet, preparations containing any fresh fish or shellfish tissue must be used immediately.
1.1.6 Antimetabolites and inactivation
Acetylcholine is an antagonist to thiamine and pyrithiamine. Oxythiamine and normal butylthiamine are specific antimetabolites.
Several thiaminases occur which destroy thiamine. These rupture the thiazole ring at the sulphur bond making the residue inactive. Freshwater fish tissues have high thiaminase activity as do tissues from clams, shrimp and mussels. Thiaminases have also been found in beans and mustard seed and in several micro-organisms. Thiaminase activity is low in most saltwater fish tissues, however, and the enzyme is inactivated by heating or prolonged pasteurization. Thiamine present in fresh Torula yeast is relatively unavailable to fish, but the yeast becomes an excellent thiamine source after rupturing the cells by steam treatment or by dehydration.
1.1.7 Clinical assessment
Clinical assessment of thiamine status have been made by measuring erythrocyte transketolase activity. Typical saturated levels for thiamine activity in sea salmon range from 15 to 20 g of thiamine/g of wet liver tissue. Fingerling chinook or coho salmon reared in 10 or 15 C water on test diets containing the listed amount or more of thiamine hydrochloride: assayed at 8-10 g of thiamine/g of wet liver tissue. These liver storage levels and normal erythrocyte transketolase activity in the absence of any deficiency sign will indicate an adequate thiamine intake for that fish population.
Following the recognition that the original water-soluble B consisted of more than one factor, a new growth-promoting factor consisting of yellow-green pigments was isolated in 1879. This new factor was designated G by some and 83 by others. The second water-soluble vitamin discovered was given its specific chemical name riboflavin. Lactoflavin, hepatoflavin, and ovoflavin were also shown to be identical with the pure riboflavin. Riboflavin occurs in the free form only in the eye, whey and urine.
Table 3 Vitamin Requirements for Growth 1/
|
Vitamin (mg/kg dry diet) |
Rainbow trout |
Brook trout |
Brown trout |
Atlantic salmon |
Chinook salmon |
Coho salmon |
|
Thiamine |
10-12 |
10-12 |
10-12 |
10-15 |
10-15 |
10-15 |
|
Riboflavin |
20-30 |
20-30 |
20-30 |
5-10 |
20-25 |
20-25 |
|
Pyridoxine |
10-15 |
10-15 |
10-15 |
10-15 |
15-20 |
15-20 |
|
Pantothenate |
40-50 |
40-50 |
40-50 |
* |
40-50 |
40-50 |
|
Niacin |
120-150 |
120-150 |
120-150 |
* |
150-200 |
150-200 |
|
Folacin |
6-10 |
6-10 |
6-10 |
5-10 |
6-10 |
6-10 |
|
Cyanocobalamin |
* |
* |
* |
* |
0.015-0.02 |
0.015-0.02 |
|
myo-Inositol |
200-300 |
* |
* |
* |
300-400 |
300-400 |
|
Choline |
* |
* |
* |
* |
600-800 |
600-800 |
|
Biotin |
1-1.5 |
1-1.5 |
1.5-2 |
- |
1-1.5 |
1-1.5 |
|
Ascorbate |
100-150 |
* |
* |
* |
100-150 |
50-80 |
|
Vitamin A |
2 000-2 500 I.U. |
* |
* |
- |
* |
* |
|
Vitamin E 2/ |
* |
* |
* |
- |
40-50 |
* |
|
Vitamin K |
* |
* |
* |
- |
* |
* |
1/ Fish fed at reference temperature with diets at about protein requirement
2/ Requirement directly affected by amount and type of unsaturated fat fed
* Denotes a requirement, the level of which has not been established
Table 3 (concluded) Vitamin Requirements for Growth 1/
|
Vitamin |
Carp |
Channel catfish |
Eel |
Sea bream |
Turbot |
Yellowtail |
|
Thiamine |
2-3 |
1-3 |
2-5 |
* |
2-4 |
* |
|
Riboflavin |
7-10 |
* |
* |
* |
* |
- |
|
Pyridoxine |
5-10 |
* |
* |
2-5 |
* |
* |
|
Pantothenate |
30-40 |
25-30 |
* |
* |
* |
* |
|
Niacin |
30-50 |
* |
- |
* |
- |
- |
|
Folacin |
- |
* |
* |
- |
* |
- |
|
Cyanocobalamin |
- |
* |
- |
* |
- |
- |
|
myo-Inositol |
200-300 |
* |
- |
300-500 |
- |
- |
|
Choline |
500-600 |
* |
- |
* |
* |
- |
|
Biotin |
1-15 |
* |
* |
- |
* |
* |
|
Ascorbate |
30-50 |
30-50 |
- |
* |
- |
* |
|
Vitamin A |
1 000-2 000 I.U. |
* |
- |
- |
- |
* |
|
Vitamin E 2/ |
80-100 |
* |
- |
- |
- |
* |
|
Vitamin K |
* |
* |
- |
- |
- |
- |
1/ Fish fed at reference temperature with diets at about protein requirement
2/ Requirement directly affected by amount and type of unsaturated fat fed
* Denotes a requirement, the level of which has not been established(Adapted from: J.E. Halver. In Fish Nutrition, 1972, 39 p; NAS/NRC, 1973;1977)
1.2.1 Chemistry
Riboflavin

Riboflavin is a yellow-brown crystalline pigment. The vitamin is very slightly soluble in water but soluble in alkali. It is insoluble in most organic solvents. Riboflavin is stable to oxidizing agents in strong mineral acids and in neutral aqueous solution. It is also stable to dry heat but is irreversibly decomposed on irradiation with ultraviolet rays or visible light, breaking down to lumiflavin. Riboflavin phosphate is the chemically active group of Warburg's 'yellow' enzyme.
1.2.2 Positive functions
Riboflavin functions in the tissues in the form of flavin adenine dinucleotide (FAD) or as flavin mononucleotide (FMN). The flavo-proteins function as enzymes of tissue respiration and are involved in hydrogen transport to catalyze the oxidation of reduced pyridine nucleo-tides (NADH and NADPH). Thus, they function as coenzymes for many oxidases and redunctases such as cytochrome e reductase, D- and L-amino acid oxidases, xanthine and aldehyde oxidase, succinic dehydrogenase, glucose oxidase and fumaric dehydrogenase. Riboflavin is also involved with pyridoxine in the conversion of tryptophan to nicotinic acid and is most important in the respiration of poorly vascularized tissues such as the cornea of the eye. Riboflavin is involved in the retinal pigment during light adaptation and lack of it causes impaired vision and photophobia in experimental animals, including fish.
1.2.3 Deficiency syndrome
A summary of deficiency signs in fish is included in Table 2. Tissue riboflavin is exhausted in young salmonids after 10 to 12 weeks on riboflavin-deficient diets in 10-15 °C water systems. Poor appetite and poor diet efficiency are the first signs, followed by photophobia, mono or bilateral cataract's, corneal vascularization, eye haemorrhage, incoordination, and general anaemia. Dark pigmentation, coupled with striated constrictions of abdominal wall in salmon have been noted. Skin atrophy has been reported for some fish species and abnormal pigmentation of both skin and iris has been noted. Replacement of riboflavin in the diet reduced the symptoms except when cataracts have developed. This irreversible condition will continue in monolateral cataracts throughout the life of the fish, whereas bilateral cataracts largely result in eventual starvation and death of the afflicted animal. The first specific signs have consistently appeared in and about the eyes of salmonids, carps and catfish.
1.2.4 Requirements
The requirements of fish for riboflavin under experimental conditions in 10-15 °C water supplies is listed in Table 3. Values for the trout are slightly lower than those reported for salmon. The requirements may vary depending upon the balance of other dietary ingredients, caloric density and environmental conditions under which the fish is raised. The values established under these standard test conditions should serve as tentative-levels which will satisfy biological demands for normal growth, health, and physiological function. Most of these studies have been made on very young fish, often initially feeding fry, with the logical assumption that their vitamin needs would be more than that of larger fish having advanced metabolic enzyme systems capable of synthesizing at least some of these vitamins to partially meet the requirements.
1.2.5 Sources and protection
Riboflavin is widely distributed in plants and in animal glandular tissues. Milk, liver, kidney, heart, yeast, germinated grains, peanuts, soybeans and eggs are rich sources. Keeping feeds from sunlight or intense artificial light is necessary to minimize loss of the vitamin by conversion to lumiflavin.
1.2.6 Antimetabolites and inactivation
When the ribose group in the molecule is replaced by other groups, analogues are formed which either have reduced activity or become antimetabolites. Galactoflavin is an antagonist to riboflavin and inhibits growth of rats when the diet contains this compound. Flavin monosulphate inhibits D-amino acid oxidase and appears to act as a competitor and inhibitor for growth of Lactobacillus casei. The hydroxyethyl analogue is an antagonist for riboflavin function in both rats and bacteria and also shows antifungal activity.
1.2.7 Clinical assessment
Liver tissue of actively feeding sea salmon contains 6-8 g of riboflavin/g of wet tissue. In a freshwater environment, young fish fed test diets in 15 ºC water systems showed liver storage of 3.5-4.0 m g/g. Riboflavin content of blood plasma does not change significantly in riboflavin deficiency in experimental animals. However, the erythrocyte riboflavin content has been reported at around 10m g/100 ml blood for man on low riboflavin intake and approximately twice that level on high intake. Urinary excretion of riboflavin has been used clinically to indicate dietary status, with 200 m g/24 it suggesting adequate intake and less than 100 m g indicating low intake. Excretion of 50 m g or less daily is strong indication of extended dietary deficiency. Several studies show that excretion of less than 200 m g of riboflavin/g of creatinine would be indicative of deficiency.
A new factor which could cure dermatitis in rats was reported by Györgi in 1935 and was named vitamin B6. The name pyridoxine is also attributed to Györgi. The active material was isolated in 1939 and pyridoxine was subsequently synthesized. First quantitative requirements for fish were described in 1944. Pyridoxine deficiency has since been reported in trout and in salmon.
1.3.1 Chemistry
Pyrodoxine hydrochloride

Pyridoxal

Pyrodoxamine

Compounds which have vitamin B6 activity include: pyridoxine, pyridoxal and pyridoxamine. Pyridoxine hydrochloride is readily soluble in water and is, heat stable in either acid or alkaline solution. It acts as a coenzyme in a number of enzyme systems. Pyridoxine is sensitive to ultraviolet light in neutral or alkaline solutions. Pyridoxamine and pyridoxal in dilute solutions are labile compounds which are rapidly destroyed on exposure to air, heat, or light. Therefore, most vitamin supplementation is in the form of pyridoxine hydrochloride and analysis for pyridoxine activity by microbiological assay of diet ingredients probably measures pyridoxal phosphate and other intermediates as well.
1.3.2 Positive functions
Pyridoxal phosphate is the coenzyme, codecarboxylase, involved in the decarboxylation of amino acids. It is also the co-factor of the 22 different transaminases present in animal tissues. As codecarboxylase, pyridoxal phosphate has been shown to participate in the decarboxylation of 5-hydroxytryptophan to produce 5-hydroxytryptamine or serotonin. As a co-factor of the enzyme desulphydrase converts cysteine to pyruvic acid. Pyridoxal phosphate is the co-factor for the synthesis of d -amino-levulinate, the latter a precursor of haeme. Many neuro hormones require pyridoxal phosphate as a coenzyme in their synthesis. Pyridoxine is also involved in fat metabolism, especially of the essential fatty acids. It is involved in the synthesis of messenger RNA which determines amino acids sequence in polypeptide synthesis. Pyridoxine plays a most important role in protein metabolism and, as a result, carnivorous fish have stringent requirements for the vitamin in the diet and stores are rapidly exhausted.
1.3.3 Deficiency syndrome
Signs of pyridoxine deficiency in fish are listed in Table 2. Since salmonids, ictalurids, and very young cyprinids are carnivorous, with protein requirements for young animals between 40-50 percent of the ration, pyridoxine stores are rapidly exhausted when fish are held on pyridoxine-deficient rations. Acute deficiency signs occur in salmon after 14-21 days on a high protein diet of pyridoxine and the entire population dies within 28 days in 12-15 C water. Deficiency is followed by general nervous disorders, and alteration in control of melanophore contraction. Postmortem rigor mortis occurs very rapidly. Rapid and gasping breathing with flexing of the opercules is a common observation and oedema in the peritoneal cavity with colourless serous fluid occurs in fish on some experimental treatments.
Salmon, trout, carp and yellowtail exhibit premortem rigor a few hours before death. Recovery at late stage deficiency is very unlikely unless fish are injected with pyridoxal phosphate. Handling the animals generally induces more damage than vitamin administration will correct. Recovery among fish still feeding is equally rapid and dramatic upon administration of pyridoxine hydrochloride in the diet. Deficiency signs disappear within a day or two. Erythrocyte and plasma transaminase activities reflect the deficiency state.
1.3.4 Requirements
Pyridoxine requirements for trout, salmon, carp, and yellowtail are listed in Tables 3 and 4. Supplementing commercial fish diets with pyridoxine is desirable because of the important role the vitamin plays in protein metabolism.
1.3.5 Sources and protection
Good sources of pyridoxine are yeast, whole cereals, egg yolk, liver and glandular tissues. Pyridoxine compounds in phosphorylated form present in agricultural products are fairly stable but are sensitive to ultraviolet radiation. Some pyridoxal phosphate will be lost on exposure to air. Free forms or pyridoxal and pyridoxamine are rapidly destroyed by air, light and heat when moist.
1.3.6 Antimetabolites and inactivation
Antagonists may compete for reaction sites of the apoenzyme or may react with pyridoxal phosphate to form inactive compounds. Deoxypyridoxine is a potent pyridoxine antagonist because of competition for apoenzyme sites but is a useful agent to accelerate pyridoxine deficiency in experimental animals. This same compound inhibits tyrosine decarboxylase. Methoxypyridoxine is another antagonist. Oxopyrimidine (2-methyl-4-amino-5-hydroxymethyl-pyrimidine) produces liver damage and inhibits glutamate decarboxylase.
1.3.7 Clinical assessment
Plasma and erythrocyte transaminase activity in most instances reflects pyridoxine status of fish although high tryptophan load in the diet tends to increase the pyridoxine requirement. Liver storage measured by microbiological assay showed 5-6 V-g of pyridoxine activity/g in fresh sea salmon liver; whereas, fingerling salmon fed a 50 percent protein diet in freshwater had 2-3 mg/g of wet tissue. Assay of diet for pyridoxine by microbiological methods gives a truer representation of total pyridoxine activity including intermediates.
A chick dermatitis was cured by Elvehjem and Köehn in 1935 using a factor containing b -alanine. The term pantothenic acid was given earlier by Williams to a yeast growth factor which was later recognized as the anti-dermatitis factor. Pantothenic acid was synthesized by Stiller in 1940. Phillips observed clubbed gills in trout fed pantothenic acid-deficient diets in 1945. Rucker detected the same condition in salmon fed low pantothenic acid diets.
1.4.1 Chemistry
Pantothenic acid

Pantothenic acid may be considered as dihydroxydimethylbutyric acid bonded to b -alanine. The free acid is a yellow, viscous oil and therefore the compound generally used in fish diet preparation is the calcium salt. This salt is a white crystalline powder readily soluble in water, mild acid, and is almost insoluble in organic solvents. It is stable to oxidizing and reducing agents and to autoclaving, but is labile to dry heat, hot alkali, or hot acid. Pantothenol has almost as much activity as pantothenic acid for growth of chicks. Pantothenic acid acetate, benzoate, and diphosphate esters are biologically active for animals but not for lactic acid bacteria. The optical isomer L-pantothenic acid, appears physiologically inert. Some organisms may utilize a portion of the molecule. Bacteria appeared to require only the dihydroxydimethylbutyric acid, and some yeasts utilize only g-alanine. Animals, however, need the entire pantothenic acid molecule or its reduced alcohol form.
1.4.2 Positive functions
Pantothenic acid is part of acetyl coenzyme A which occurs in many enzymatic processes involving 2-carbon compounds. It has been shown to be required by all animal species studied and by many micro-organisms.
The acetyl coenzyme A system is involved in the acetylation of aromatic amines and choline; condensation reactions for synthesis of acetate, fatty acids, and citrate; the oxidation of pyruvate and acetaldehyde; and is essential for the development of the central nervous system. The 2-carbon fragment called 'active acetate', or acetyl coenzyme A, is an essential intermediate in metabolism. It is involved in acylation of acetate, succinate, benzoate, propionate and butyrate. Pantothenic acid is involved in adrenal function and for the production of cholesterol. Coenzyme A is also involved in many other steps of intermediate metabolism of carbohydrates, fats and proteins.
1.4.3 Deficiency syndromes
Deficiency signs for pantothenic acid are summarized in Table 2. Under standard test conditions with deficient diet fed in 10-15°C water systems, salmon and trout exhaust pantothenic acid stores within 8-12 weeks. Fish stop feeding and close examination of gill filament show proliferation of epithelial surface plus swelling and clubbing together of the filaments and lamellae. The opercules become distended and the surface of the gills is often covered with an exudate. Fish become prostrate or sluggish. Necrosis, scarring and cellular atrophy of the tender gill elements occur and anaemia develops after prolonged deficiency. Dietary gill disease has been adequately described and correlated with pantothenic acid deficiency. The same type of symptom has been observed in salmon, trout, eel, carp and catfish. After replacement of pantothenic acid in the diet, recovery is rapid for those fish still feeding and gross deficiency symptoms disappear after about four weeks on the recovery diet although evidence of necrosis and scarring remains.
1.4.4 Requirements
Dietary requirements for salmon, trout and other fish are listed in Table 3.
1.4.5 Sources and protection
Good sources for pantothenic acid are cereal bran, yeast, liver, kidney, heart, spleen and lung. Fish flesh is a relatively rich source, although the content is only about 20 percent of pantothenic acid found in animal glandular tissue. Royal jelly probably contains the greatest amount as it contains over 500 m g of pantothenol/g dry weight. The calcium or sodium salt of pantothenic acid is relatively stable and can be incorporated into either moist or dry fish diets. Some loss is incurred during autoclaving and excessive heat should therefore be minimized during diet preparations. Since the free acid is labile to heat and also to acid and alkali, some loss can be expected during moist diet preparation or during storage. Certain cereal brans may have pantothenic acid bound in a form unavailable to fish because of the low digestibility coefficient and should not be relied upon as the sole pantothenic acid source in the diet.
1.4.6 Antimetabolites and inactivation
Since pantothenic acid affects the respiration of many types of cells, compounds like 6-mercaptopurine, 2,6-diaminopurine, and 8-azaguanine, which inhibit growth of tumors are antagonistic toward pantothenic acid. Pantoyltaurine is an antimetabolite of pantothenic acid and has been used to accelerate deficiency syndromes in experimental animals.
Methyl-w -pantothenic acid was reported to interfere with the formation of acetyl coenzyme A and accelerates deficiency syndromes in animals. This compound inhibits sulfanilamide acetylation in pigeon liver homogenates but did not prevent citric acid formation. Pantothenic acid can be used to overcome these inhibitory effects, including the reversal of mitosis blockage by 7-mecaptopurine in animals. High levels of calcium pantothenate in the diet also has a transient effect on the migratory urge of salmon.
1.4.7 Clinical assessment
Assay of pantothenic acid content in the diet may be misleading unless care is exercised in proper hydrolysis of the raw materials being assayed since pantothenic acid is only slowly liberated by normal hydrolytic procedures. Complete hydrolysis with enzyme preparations will liberate all of pantothenic acid from biologically active material of glandular tissues, fish flesh, yeast, and bran.
Salmon feeding actively in the oceans showed liver pantothenic acid content of 18-20 m /g of fresh tissue. Young chinook and coho salmon fingerlings reared in fresh water at 12-15°C showed maximum liver storage at about 14-16 m g of pantothenic acid/g of fresh liver tissue. Load tests and acetylation reactions of sulfanilamide have been used in man and other experimental animals but have not yet been extended for assessment of pantothenic acid status in fish.
Nicotinic acid was synthesized in 1873 but was left on the shelf as an organic compound unrelated to the severe pellagra afflictions occurring throughout the world at that time. Sixty years later the compound was shown to be present in coenzymes I and II and two years thereafter Elvehjem cured 'black tongue' in dogs with the vitamin. Niacin was postulated to be part of factor H for fish in 1937, but deficiency symptoms were not adequately described until reported in trout in 1947.
1.5.1 Chemistry
Nicotinic acid

Niacinamide

Niacin is the preferred nomenclature. Nicotinic acid amide or niacinamide is the common form in which the vitamin is physiologically active. Niacin is a white, crystalline solid, soluble in water and alcohol. It is stable in the dry state and may be autoclaved for short periods without destruction. It is also stable to heat in mineral acids and alkali. Niacin is both a carboxylic acid and an amine and forms quaternary ammonium compounds because of its basic nature. Acidic characteristics include salt formation with alkali. Niacin can be esterified easily, then converted to amides. Niacinamide is a crystalline powder soluble in water and ethanol and the dry material is stable up to about 60°C. In aqueous solutions it is stable for a short period when autoclaved. It is the form in which the vitamin is normally found in niacinamide adenine dinucleotide (NAD), and in niacinamide adenine dinucleotide phosphate (NADP).
1.5.2 Positive functions
The major function of niacin in NAD and NADP is hydrogen transport in intermediary metabolism. Most of these enzyme systems function by alternating between the oxidized and reduced state of the coenzymes NAD-NADH and NADP-NADPH. Oxidation-reduction reactions may be anaerobic as when pyruvate acts as the hydrogen acceptor and lactate is formed, or the reactions may be coupled to electron transport systems with oxygen as the ultimate hydrogen acceptor; such aerobic reactions occur in respiration. Both NAD and NADP are involved in the synthesis of high energy phosphate bonds which furnish energy for certain steps in glycolysis, in pyruvate metabolism, amino acid and protein metabolism, and in photosynthesis. An interrelationship between thiamine and niacin exists since both vitamins are coenzymes in intermediary metabolism whereby food is oxidized to furnish energy for physiological functions, to maintain homeostasis, and for body temperature maintenance in homeotherms.
1.5.3 Deficiency syndrome
The deficiency signs in fish are listed in Table 2. Stores of niacin are more slowly exhausted under experimental conditions than are some of the other vitamins resulting in less defined and more slowly developing symptoms.
Niacin deficiencies in fish were experimentally induced in the late forties and early fifties by using basal diets which had a low niacin content. Loss of appetite and poor food conversion were the first signs noted. Then the fish turned dark and went off feed, followed by the appearance of lesions in the colon, erratic motion, oedema of the stomach and colon and muscle spasms while fish were apparently resting. A predisposition to sunburn in fish confined in the open in shallow ponds or raceways was described. Common carp showed a congestion of the skin with subcutaneous hemorrhages. Common symptoms of niacin deficiency in most fish studied were muscular weakness and spasms, coupled with poor growth and poor food conversion.
1.5.4 Requirements
Niacin requirements in young fish experimentally tested are listed in Table 3.
In homeotherms on a balanced test ration the niacin requirement is generally estimated to be about ten times that of the thiamine requirement. These rations generally contain considerable carbohydrate material to furnish energy to maintain body temperature. In fish the requirement appears to be twenty to thirty times that of the thiamine needs determined for the same test conditions and test rations. This difference may be due to low carbohydrate content of young fish diets and the higher protein content of these rations. Conversion of tryptophan to niacin occurs in the mammalian liver and possibly also in the liver of fish. This conversion may account for the slow development of the niacin-deficiency syndrome in fish. However, after 10-14 weeks on diets devoid of niacin, deficiency symptoms did occur in several species of fish. The symptoms were reduced by replacement of niacin in the ration even when high protein diets containing an excess of tryptophan were fed. Too much niacin inhibits growth.
1.5.5 Sources and protection
Niacin is found in most animal and plant tissues. Rich sources are yeast, liver, kidney, heart, legumes, and green vegetables. Wheat contains more niacin than corn and the vitamin is also found in milk and egg products. The vitamin is very stable since it is generally found in coenzyme form in raw materials. Niacin added to the diet as a supplement remains relatively unaltered during diet manufacture, processing, and storage.
1.5.6 Antimetabolites and inactivation
Pyridine-3-sulphonic acid and 3-acetylpyridine are compounds structurally related to niacin and are antimetabolites for this vitamin in animals and in micro-organisms. Additional niacin can overcome the anti-metabolic effect. Deficiency symptoms in rats may be induced by 6-amino niacinamide. The symptoms are reversed by addition of ten times more niacinamide than the antimetabolite. Thioacetamide has been reported to be a niacin antagonist in fish.
1.5.7 Clinical assessment
Salmon feeding actively in the oceans showed liver niacin content of 70-80 m g of niacin/g of fresh tissue. About half of this amount is present in fingerling salmon raised in fresh water at 12-15°C and fed test rations containing 40-50 percent protein and niacin supplements 500-750 mg/kg dry diet. Urinary metabolities of niacin have been measured in other animals on standard niacin load in test rations containing a standard tryptophan load. The technique is well developed to measure the N1-methyl derivative in mammalian urine. These data have not been reported for fish but metabolism chambers are available for collecting branchial and urinary wastes from large fish intubated with different diet material.
Certain foods, notably liver and kidney, were found to be protective against a form of dermatitis resulting from the consumption of egg white by rats, in the early thirties. It was soon learned that a specific component of egg albumin, avidin, rendered dietary biotin unavailable, hence producing the symptoms. Biotin was variously called coenzyme R and 'vitamin H'. It was isolated by Du Vigneaud in 1941 and synthesized by workers of Merck and Company in 1943. Biotin was once thought to be part of factor H for fish. Blue slime patch disease due to biotin deficiency was reported in trout.
1.6.1 Chemistry
Biotin

Biotin is a monocarboxylic acid slightly soluble in water and alcohol and insoluble in organic solvents. Salts of the acid are soluble in water. Aqueous solutions or the dry material are stable at 100 C and to light. The vitamin is destroyed by acids and alkalis and by oxidizing agents such as peroxides or permanganate. Biocytin is a bound form of biotin isolated from yeast, plant, and animal tissues. Other bound forms of the vitamin can generally be liberated by peptic digestion. Oxybiotin has partial vitamin activity but oxybiotin sulphonic acid and other analogues are antimetabolites inhibiting the growth of bacteria.
Avidin, a protein found in raw egg white, binds biotin and makes it unavailable to fish and other animals. Heating to denature the protein makes the bound biotin available again to the fish. Biocytin or e -biotinyl lysine (the epsilon amino group of lysine and the carboxyl of biotin being combined in a peptide bond) is hydrolyzed by the enzyme biotinase making the protein-bound biotin available.
1.6.2 Positive functions
Biotin is required in several specific carboxylation and decarboxylation reactions, including the carboxylation of pyruvic acid to form oxaloacetic acid. It is part of the coenzyme of several carboxylating enzymes fixing CO2 such as propionyl coenzyme A involved in the conversion of propionic acid to succinic acid in methylmalonyl coenzyme A. Biotin is also involved in the conversion of acetyl CO2 to malonyl coenzyme A in the formation of long chain fatty acids. It has possible involvement in citrulline synthesis and may have effects on purine and pyrimidine synthesis. It is involved in the conversion of unsaturated fatty acids to the stable cis form in the synthesis of biologically active fatty acids.
1.6.3 Deficiency syndrome
Some signs of biotin deficiency in salmonids are skin disorders, muscle atrophy, lesions in the colon, loss of appetite, and spastic convulsions. Haematology discloses fragmentation of erythrocytes. Poor growth is a common symptom and has been reported for salmonids, common carp, goldfish (Carassius auratus) and eel. Blue slime patch disease in brook trout deficient in biotin appears typical for this species. Fish reared in 10 -15 C water exhaust biotin stores in 8-12 weeks and the first signs are anorexia, poor food conversion, and general listlessness before the more acute deficiency symptoms become detectable. General deficiency signs are listed in Table 2.
1.6.4 Requirements
The biotin requirements for young salmon, trout, carp, goldfish and eel raised under experimental conditions appear to be about the same. Brown trout appear to require nearly twice as much biotin in the diet as do brook or rainbow trout. Requirements are listed in Table 3.
1.6.5 Sources and protection
Rich sources of biotin are liver, kidney, yeast, milk products, and egg yolks. Nut meats contain good supplies of biotin. The diet should be protected from strong oxidizing agents or conditions which promote oxidation of ingredients. Raw egg white should not be incorporated into moist fish diets. Cooking will inactivate the avidin.
1.6.6 Antimetabolites and inactivation
Raw egg white has already been discussed which irreversibly binds biotin and makes it unavailable to young fish. Many biotin homologues with different side chain lengths inhibit the growth of bacteria. Oxybiotin, in which sulphur is replaced by oxygen, has about the same biological activity as the natural biotin. On the other hand, oxybiotin sulphonic acid inhibits biotin activity.
1.6.7 Clinical assessment
Measurement or urinary excretion of biotin in animals does not prove a good clinical method since biotin is synthesized by several organisms in the gut. In fish, this technique needs to be explored and may well be valuable as a clinical tool since the gastrointestinal tract of many freshwater fish contains only small quantities of bacteria. Biotin is one of the most expensive vitamins to add to fish rations. Salmon feeding actively in the oceans had liver biotin concentrations of 10-12 mg of wet liver tissue. The concentrations in the liver of young salmon fingerlings in fresh water fed test diets containing an excess of biotin were between 6-8 mg of the vitamin/g of tissue. Fish with these levels of biotin in the liver should probably be in sound biotin nutritional status.
A factor effective in curing metaloblastic anaemia in monkeys was found in yeast and liver concentrates and was designated vitamin M in 1935. In 1939 an anti-anaemic factor was found in liver and called vitamin BC. These were later shown to be the same substance with folic acid as the active ingredient. Folic acid was synthesized in 1946 and was soon used in fish diets for preventing purified diet anaemia.
1.7.1 Chemistry
Folic acid

Folic acid crystallizes into yellow spear-shaped leaflets which are soluble in water and dilute alcohol. It can be precipitated with heavy metal salts. It is stable to heat in neutral or alkaline solution, but unstable in acid solution. It deteriorates when exposed to sunlight, or during prolonged storage. Several analogues have biological activity including pteroic acid, rhizopterin, folinic acid, xanthopterin and several formyltetrahydropteroyl-glutamic acid derivatives. These have closely allied ring structures and many have been isolated as derivatives in various animals or microbiological preparations. One simple form, xanthopterin, present in the pigments of insects, is shown below and is of special interest because of early work with this compound as the anti-anaemic factor H for fish.
Xanthopterin

1.7.2 Positive functions
Folic acid is required for normal blood cell formation and is involved as a coenzyme in one-carbon transfer mechanisms.
In the presence of ascorbic acid, folic acid is transformed into the active (5-formyl-5, 6,7,8) tetrahydrofolic acid. Folic acid is involved in many one-carbon metabolism systems such as serine and glycine interconversion, methionine-homocysteine synthesis, histidine synthesis, and pyrimidine synthesis.
Several coenzyme forms of the active vitamin have been isolated. Folic acid is involved in the conversion of megaloblastic bone marrow to normoblastic type. It has a role in blood glucose regulation and improves cell membrane function and hatchability of eggs.
1.7.3 Deficiency syndrome
Macrocytic normochromic anaemia occurs in several experimental animals, including fish fed diets devoid of the folic acid. Increasing numbers of senile cells are observed as the deficiency progresses until only a few old and degenerating cells are found in the blood of deficient fish. Anterior kidney imprints disclose only adult cells and no preforms present. Other signs observed have been poor growth, anorexia, general anaemia, lethargy, fragile fins, dark skin pigmentation, and infarction of spleen.
1.7.4 Requirements
Requirements of fish for folic acid are listed in Table 3. The requirement seems to be about the same for trout and salmon. Marginal macrocytic anaemias occur in fish fed diets containing marginal amounts of folacin. Individual fish ingesting adequate amounts of the vitamin show little variation in total erythrocyte counts.
1.7.5 Sources and protection
Yeast, green vegetables, liver, kidney, glandular tissue, fish tissue, and fish viscera are good sources of folic acid. Insects contain xanthopterin which has folic acid activity. At one time the yellow pigment of xanthopterin was identified as the fish anti-anaemic factor H, but subsequent experiments showed only partial activity and that folic acid itself was a much more potent antimacrocytic anaemia factor. Insects may contribute significantly to the folic acid requirements of wild fish, but in scientific fish husbandry artificial diets are more reliable sources. Activity is lost during extended storage and when material is exposed to sunlight. Therefore, dry feeds should be carefully protected during manufacture and moist diet rations should be carefully preserved. Both types of fish diets should be fed soon after manufacture to assure minimal loss of folic acid activity.
1.7.6 Antimetabolites and inactivation
One antagonist of folacin is 4-aminopteroylglutamic acid or aminopterin. This material, when incorporated in the diet of guinea pigs and rats, induces anaemia and leucopenia and has been used to treat leukemia in man. Amethopterin (4-amino-N10-methylpteroylglutamic acid) can also be used to induce deficiency by inhibiting purine, pyrimidine, and nucleic acid protection; viz., inhibition of nucleic acid synthesis results in macroytic anaemia.
1.7.7 Clinical assessment
Haematology is used as a simple clinical tool to assess haemopoiesis in fish. Anterior kidney imprints easily disclose normal distribution of immature cells and preforms undergoing reticulosis. Salmon actively feeding in the ocean and young salmon fingerlings raised in fresh water and fed diets rich in folacin show liver storage of 3-4 m g of folic acid/g of wet tissue. Microbiological assay is preferred for assessment of total folic acid in dietary raw materials because the total biological activity it measures includes all the various coenzyme forms and folic acid analogues. Assessment of the dietary intake of folic acid is important for intensive cold water fish husbandry. In pond culture, aquatic and terrestrial insects, algae, etc., may also be available and folic acid in the supplementary feed may not be as critical. Since folic acid is labile in storage, excess amounts are generally added to manufactured feed in anticipation of storage losses. However, prudent fish husbandry dictates rapid use of manufactured rations with minimum storage. Routine periodic haematology of fish assures proper nutritional status for maximum production and sound health. The author has noted in several series of experiments that when fish diseases occur through inadvertent contamination of the water supply, those groups of fish partially or completely deficient in folic acid were among the first to show acute disease symptoms. Therefore, folic acid must also play an important role in resistance to disease.
The antipernicious anaemia factor found to be contained in liver was isolated and crystallized in the mid forties. This substance, named vitamin B12 by its discoverers, was later to be recognized as essential for growth of chicken fed diets entirely of plant origin and was designated animal protein factor (APF). When anaemic salmon were injected with crystalline B12 in combination with folic acid and xanthopterin positive haemopoiesis occurred within a few days, and the salmon showed rapid recovery from the anaemia.
1.8.1 Chemistry
Vitamin B12 or cynacobalamin has the following approximate chemical structure.
Cyanocobalamin
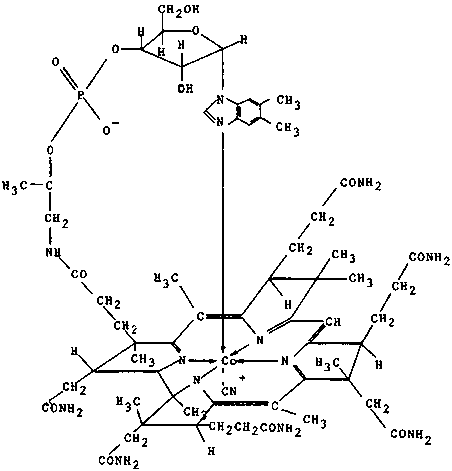
The molecule has a planar group and a nucleotide group lying nearly at right angles to one another. This cobalt-containing vitamin has a net charge of one at the central cobalt atom to which is attached a replaceable cyano group. Vitamin B12 is stable to mild heat in neutral solution, but is rapidly destroyed by heating in dilute acid or alkali. Crude concentrates are more unstable and rapidly lose activity. The compound is similar to the porphyrins in its spatial configuration with a central cobalt atom linked to four reduced pyrrole rings in the haeme series. Replacing the cyanide ion with a variety of anions produce derivatives which have comparable biological activities; viz., hydroxocobalamin, nitritocobalamin, chlorocobalamin and sulphatocobalamin.
1.8.2 Positive functions
Cyanocobalamin is involved with folic acid in haemopoiesis. It is required by many micro-organisms and is a growth factor for many animals. The animal protein factor present in fish and animal by-products was not recognized until crystalline vitamin B12 was injected into anaemic chinook salmon fingerlings in 1949 and positive haemopoiesis was observed. A coenzyme incorporating vitamin B12 is involved in the reversible isomerization of methyl-malonyl coenzyme A to succinyl coenzyme A and in the isomerization of methylaspartate to glutomate. Cyanocobalamin is involved in the coenzyme for the methylation of homocystine to form methionine. It is also involved in several other one-carbon reactions and in the synthesis of labile methyl compounds. One vitamin B12 containing coenzyme acts in methylation of the purine ring during thymine synthesis. Vitamin B12 is also involved in cholesterol metabolism, in purine and pyrimidine biosynthesis, and in the metabolism of glycols.
1.8.3 Deficiency syndrome
Deficiency signs in young pigs, chicks, and rats show abnormal blood elements, poor growth, and pernicious anaemia. An intrinsic factor is necessary for good absorption of the vitamin from the gut. This factor is a low molecular weight mucoprotein which normally occurs in gastric juice, and especially in hog gut mucosa. Pernicious anaemia in chinook and coho salmon is characterized by fragmented, erythrocytes with many aberrant forms present. Haemoglobin levels are inconsistent and erythrocyte counts have a range extending from frank anaemia to a near normal blood pattern. Cyanocobalamin stores in fish tissues are slowly exhausted and only after 12-16 weeks on test do the symptoms appear in deficient salmon populations. Poor appetite, poor growth, poor food conversion, and some dark pigmentation can be observed before frank anaemia is detected.
1.8.4 Requirements
The figures listed in Table 3 represent estimates of requirement based on fish that showed normal haematology at the end of the 16-week feeding period, and maximum liver storage of the vitamin determined by Lactobaccilis leichmanii microbiological assay.
1.8.5 Sources and protection
Rich sources of vitamin B12 are found in fish meal, fish viscera, liver, kidney, glandular tissues, and slaughter house wastes. Since vitamin B12 is labile on storage, and in mild acid solution is easily destroyed by heating, care must be exercised in diet preparation containing flesh or meat scraps.
1.8.6 Antimetabolites and inactivation.
The vitamin B12 coenzymes are very unstable under light, which rapidly decomposes the coenzymes. Photo-sensitivity is increased in dilute acid solutions.
1.8.7 Clinical assessment
Generalized anaemia with fragmentation of erythrocytes and extremely variable haemoglobin levels and erythrocyte counts indicate possible B12 deficiency. Prompt response in individual fish is obtained by injecting B12 alone or in combination with folic acid in the ratio of 1 part vitamin B12 to 100 parts folic acid. Careful interpretation of haemotological data will enable one to distinguish one form of anaemia from the other.
Experimental work to cure scurvy with fruit juice was described by Lind in 1753, but nearly 200 years elapsed before the exact chemical compound responsible for reducing the symptoms was defined. The isolation of ascorbic acid was due to Szent-Györgyi of Hungary and C.G. King of the U.S.A. Vitamin C synthesis was accomplished in 1933 after the chemical structure of ascorbic acid was established by British and Swiss workers.
McCay and Tunison reported scoliosis in brook trout fed formalin-preserved meat in 1934 and McLaren observe haemorrhages in trout fed rations low in ascorbic acid. It was not until the sixties that a critical need for L-ascorbic acid by trout and salmon was demonstrated.
1.9.1 Chemistry

L-ascorbic acid is a white, odourless, crystalline compound, soluble in water but insoluble in fat solvents. It is readily oxidized to dehydroascorbic acid, the less biologically potent form. Ascorbic acid is very stable in acid solution because of the preservation of the lactone ring, but in alkaline solution hydrolysis occurs rapidly and vitamin activity is lost. It is very heat labile and prone to atmospheric oxidation, especially in the presence of copper, iron, or several other metallic catalysts. The reduced form is the most biologically active but several derivatives or salts are obtainable which have varying degrees of ascorbate activity.
1.9.2 Positive functions
L-ascorbic acid acts as a biological reducing agent in hydrogen transport. It is involved in many enzyme systems for hydroxylation; i.e., hydroxylation of tryptophan, tyrosine, or proline. It is involved in the detoxification of aromatic drugs and also acts in the production of adrenal steroids.
Ascorbic acid is necessary for the formation of hydroxy proline which is a constituent of collagen, a component of intercellular material in bones and soft tissues. Ascorbic acid plays a synergistic role with vitamin E as intracellular antioxidants and free radical traps. The conversion of folic acid to folinic acid requires vitamin C. Ascorbic acid is involved in the formation of chondroitin sulphate and intercellular ground substance. Labelled ascorbic acid fed to fish previously deficient in the vitamin was shown to be rapidly mobilized and fixed in areas of rapid collagen synthesis and became concentrated in the thick collagen of the skin and in cartilagenous bones, as well as in the glands of the anterior kidney. Ascorbic acid is also involved in erythrocyte maturation.
1.9.3 Deficiency syndrome
Scurvy with impaired collagen formation, perifollicular haemorrhages, loose teeth and poor bone formation, anaemia, and oedema have been reported in other animals. Deficiency signs in fish are generally related to impaired collagen formation. Fish soon show hyperplasia of jaw and snout. The same symptoms have been observed in trout, salmon, yellowtail, carp, guppies and char. Histologically, hypertrophy of the adrenal tissues and haemorrhage at the bases of fins have been observed in coho salmon. Deficiency signs cease to develop and new growth becomes normal upon replacement of ascorbic acid in the ration. Anaemia eventually develops in extremely deficient fish and scoliosis and lordosis do not repair but are walled off by new growth around the afflicted areas of the spine when ascorbic acid is once again added to the ration.
1.9.4 Requirements
In the rainbow trout, reasonable blood and anterior kidney storage levels were obtained with an intake of about 100 mg of vitamin C/kg of dry ration in 10°, 12°, or 15° C water systems. When wound repair experiments were initiated, however, or when fish were exposed to other stress then the requirements doubled or tripled. When severe abdominal or intramuscular wounds were inflicted, young fish needed at least 500 mg of active ascorbate for tissue repair comparable with control fish receiving 1 g or more of ascorbate in the diet/kg of dry diet. Coho salmon appear to need about half of these requirements for adequate tissue levels and for maximum severe wound repair rates. This phenomenon is illustrated in Table 4 showing growth response and tissue repair for rainbow trout and coho salmon. The requirement for ascorbic acid is related to stress, growth rate and size of the animal, as well as to the other nutrients present in the diet. A compromise value of about 200 mg of ascorbic acid/kg diet for trout and salmon raised in freshwater systems between 10-15°C would ensure reasonable tissue storage levels and furnish some excess for mild stress conditions and for ascorbic acid loss from the diet through oxidation. Large common carp can synthesize some ascorbate and the requirement for this species may be dependent on fish size and the environment in which they are reared.
Table 4 Growth and Tissue Ascorbate
|
Vitamin C diet treatment |
Trout |
Salmon |
||||
|
Average weight at 24 weeks |
Ascorbate concentrate 1/ |
Average weight at 24 weeks |
Ascorbate concentrate 1/ |
|||
|
blood |
kidney |
blood |
kidney |
|||
|
mg/100 g |
g |
m g/g |
m g/g |
g |
m g/g |
m s/s |
|
0 |
2.4 |
2/ |
2/ |
5.0 |
22.3±2.2 |
89 |
|
5 |
9.6 |
34.4±2.9 |
125 |
6.0 |
30.5±1.2 |
132 |
|
10 |
10.6 |
34.6±1.3 |
137 |
5.7 |
35.8±1.6 |
265 |
|
20 |
10.1 |
38.8±3.3 |
132 |
6.1 |
34.2±2.3 |
183 |
|
40 |
10.2 |
46.8±6.2 |
162 |
6.3 |
33.7±2.0 |
225 |
|
100 |
10.8 |
51.0±4.6 |
247 |
6.0 |
37.8±2.3 |
321 |
1/ Average of five samples for blood (± S.D.) and two for head kidney tissue
2/ No fish available for assay
1.9.5 Sources and protection
Ascorbic acid is widely distributed in nature with citrus fruits, cabbage, liver, and kidney tissue good sources for the vitamin. Fresh insects and fish tissues contain reasonable amounts of the vitamin. Synthetic ascorbic acid is also readily available. Fish food should be protected from oxidizing agents and kept sealed or frozen until used to prevent loss of the vitamin.
1.9.6 Antimetabolites and inactivation
D-ascorbic acid, the optical isomer of the active form, has no vitamin activity and competes for sites of several enzyme reactions mediated by L-ascorbic acid. 6-deoxy-L-ascorbic acid, dehydroascorbic acid and L-glucoascorbic acid have very little activity.
1.9.7 Clinical assessment
Ascorbic acid status for experimental animals is normally attempted by tissue ascorbate analysis. Most of the assays previously used measure total ascorbate and not biologically active L-ascorbic acid. Consequently these are fraught with errors and misconception of true vitamin C status. In fish tissues, blood and liver do not adequately reflect the ascorbic acid intake and status, whereas assay of the anterior kidney which contains adrenal tissue provides a fairly representative picture of tissue storage of the vitamin. Stress rapidly reduces the ascorbic acid content of this tissue with concurrent production of adrenal steroids. Conversely, dietary repletion is reflected by up to four or five-fold storage levels from the deficient state. Examination of fragile support cartilage in the gill filaments under low magnification will detect early hypovitaminosis before clinically acute symptoms become noticeable. However, the best tissue for routine clinical analysis to assess vitamin C status in trout and salmon appears to be the anterior kidney with samples selected from the junction of the two forward wings. Tissues are blotted free of blood with filter paper, and then assayed for total ascorbate by one of the improved quick methods to determine total ascorbate.
Muscle 'sugar' was discovered by Scherer in 1850 and was characterized as inositol in 1887. Inositol was shown to be an alopecia (a form of hairlessness) preventing factor for mice in 1940. Poor growth and poor food passage in inositol-deficient fish was observed in salmon and carp.
1.10.1 Chemistry
Inositol

Seven optically inactive and two optically active isomers of hexahydroxycyclohexane can exist. One of the optically active forms, myo-inositol, is a white crystalline powder soluble in water and insoluble in alcohol and ether. The material can be synthesized, but is easily isolated from biological material in free or combined forms. The mixed calcium-magnesium salt of the hexophosphate is phytin. Isomers have little biological activity but will compete in chemical reactions. Inositol is a highly stable compound.
1.10.2 Positive functions
Myo-inositol is a structural component in living tissues. It has lipotropic action by preventing accumulation of cholesterol in one type of fatty liver disease and is involved with choline in maintaining normal lipid/metabolism. It is a growth-promoting substance for micro-organisms and prevents an alopecia in mice. It is a reserve carbohydrate in muscle as well as a major component of phosphoglycerides in animal tissues.
1.10.3 Deficiency syndrome
Poor growth, increased gastric emptying time, oedema, dark colour, and distended stomachs are symptoms observed in salmon, trout, carp and catfish held for long periods on inositol-deficient test rations.
A 'spectacle-eye' condition described for rats has not been observed under the experimental conditions used in fish studies. The major deficiency sign is inefficiency in digestion and food utilization and concomitant poor growth leading to a population of fish with distended abdomens.
1.10.4 Requirements
Inositol needs have been measured in only two species of salmon and carp and the requirement is high for maximum growth and for maximum liver storage. Whether liver storage is accurate criteria to determine requirements is debatable since inositol intake was compared with maximum growth rate and diet conversion to develop a tentative requirement for young fish for this 'muscle' sugar. Requirements of some species are listed in Table 3.
1.10.5 Sources and protection
Myo-inositol occurs ubiquitously in large amounts wherever biological tissue is found. Wheat germ, dried peas, and beans are rich sources. Brain, heart, and glandular tissues are very good sources of biologically active inositol. Citrus fruit pulp and dried yeast also contain inositol. The compound is stable.
1.10.6 Antimetabolites and inactivation
Seven optically inactive and one optically active but biologically inactive stereo isomers occur. Since inositol is synthesized in the biologically active form by many microorganisms in the gut, only large quantities of chemically synthesized inactive isomers added to diets would interfere with the metabolism of inositol for growth and normal physiological function. Biologically inactive stereoisomers of myo-inositol do not compete for critical sites in metabolism. Methyl derivatives and mono-, di-, and triphosphoric acid esters occur naturally. Salts of the hexaphosphate or phytin make the bound inositol practically unavailable to the animal.
1.10.7 Clinical assessment
Assessment in fish has been based on lack of deficiency signs coupled with the most efficient food conversion. Salmon feeding actively in oceans show 1-1.5 mg of inositol/g of fresh liver tissue and young fingerlings raised in freshwater at 10-15°C had 600-700 m g/g of liver tissue. An alternate, better assessment may be based on a standard muscle section or whole carcass analysis for free or for bound inositol. Projection of inositol intake from normal fish diet ingredients should indicate an excess of this particular vitamin.
Methylation as a basic metabolic process was postulated by Hoffmeister. Methyl transfer was shown in vivo by Thompson and the interrelationships between choline, methionine, and homo-cystine were shown by du Vigneaud in 1939-42. Trout fed low choline rations developed haemorrhagic kidneys and salmon showed an aversion to food in choline-deficient diet.
1.11.1 Chemistry
Choline

Choline is a very strong organic base and forms many derivatives widely distributed in animal and vegetable tissue. One derivative, acetylcholine, is involved in the transmission of nerve impulses across synapses. Choline is very hygroscopic, very soluble in water, and is stable to heat in acid, but decomposes in alkaline solutions.
1.11.2 Positive functions
Choline acts as a methyl donor in trans-methylation reactions. It is a lipotropic and antihaemorrhagic factor preventing the development of fatty livers. It is involved in the synthesis of phospholipids and in fat transport. Acetylcholine transmits the excitory state across the ganglionic synapses and neuromuscular junctions. Choline is essential for growth and good food conversion in fish.
1.11.3 Deficiency syndrome
Deficiency signs include poor growth and poor food conversion accompanied by impaired fat metabolism. Haemorrhagic kidneys and intestines have been reported in trout and increased gastric emptying time has been observed in salmon.
1.11.4 Requirements
The requirements for salmon are listed in Table 3.
1.11.5 Sources and protection
Rich sources of choline are wheat germ, beans, brain, and heart tissue. Choline hydrochloride, the commercially available form, may inactivate a -tocopherol and vitamin K when in direct contact with these vitamins and care should be exercised in selecting properly protected (gelatin coated) fat soluble vitamins in fish diet preparation.
1.11.6 Clinical assessment
Choline status of fish can be estimated from the assay of choline content of the dietary ingredients with the absence of deficiency signs. Maximum liver storage may not be the best criteria to determine choline nutritional status but has been used to assess the tentative requirement listed for the two species of salmon.
p-Aminobenzoic acid

Para-aminobenzoic acid is a white crystalline powder which is water soluble and heat and light stable in aqueous and mild alkaline solution. It is a growth promoting vitamin for micro-organisms which require the vitamin for folic acid synthesis. Large intake has been shown to counteract the antimetabolite effect of sulphonamides in bacterial culture. No positive function or deficiency signs have been observed in fish.
Lipoic acid

Lipoic acid is both fat soluble and water soluble. Its active form is the amide derivative known as lipoamide. Lipoic acid functions as a coenzyme in a a -ketoacid decarboxylation. It was discovered independently in several laboratories during the period 1945-50 and shown to be an essential component of multienzyme, the pyruvate dehydrogenase complex. Pyruvate is converted to 'active acetaldehyde' which in turn is picked up by lipoamide. Subsequent oxidation of the aldehyde results in the reduction of lipoamide to a disulphydryl form. The multi-enzyme unit also includes thiamine pyrophosphate, coenzyme A, and flavin adenine dinucleotide. Glandular tissues are good sources of lipoic acid. No requirements have been determined for fish.
Fat-soluble vitamins A, D, E, and K differ from the water-soluble vitamins in their accumulative action. Little evidence has been recorded for hypervitaminosis with the water-soluble vitamins since these compounds are rapidly metabolized and excreted when intake exceeds liver or tissue storage capacity, but hypervitaminosis is a common occurrence in fish and other animals when large quantities of any one of the fat-soluble vitamins are ingested. Toxicity symptoms involving vitamins A and D are indistinguishable from deficiency symptoms for the same vitamins. On the other hand, symptoms of excess vitamins E and K intake are more discrete. Fish rations may often be enriched with fish oils to increase caloric density of the ration resulting in excessive intake of the fat-soluble vitamins.
A fat-soluble factor that promoted growth in rats was described by Hopkins and by Osborne and Mendel at the beginning of this century. McCollum and Simmonds cured xerophthalmia, a disease of the eye, with this material. The chemical structure of vitamin A and its relationship to b -carotene was shown by Von Euler in 1928. Active vitamin A was synthesized in the mid-thirties.
2.1.1 Chemistry
Vitamin A alcohol

The relationship of the vitamin A alcohols to naturally occurring b -carotene is as follows:
Beta-carotene

Retinene, the aldehyde form of vitamin A, has been isolated from the retina of dark adapted eyes and is involved in vision in dim light. Retinoic acid, which is the oxidized form of vitamin A alcohol, has been shown to have some vitamin A activity. Vitamin A1 is found in saltwater fish, whereas vitamin A is more abundant in freshwater fish. Interconversion of the two forms in living fish tissue has been demonstrated. Fish oils contain vitamin A as free alcohols or esters. Vitamin A alcohol occurs as a light coloured viscous oil which is heat labile and subject to air oxidation. Beta-carotene occurs as an orange, crystalline compound which is more stable to heat and oxidation. Vitamin A is water insoluble but is soluble in fat and organic solvent.
2.1.2 Positive functions
Vitamin A is essential in maintaining epithelial cells, preventing atrophy and keratinization of epithelial tissues. It is combined with a protein in visual purple and is important in night vision. Vitamin A also prevents xerophthalmia in rats and young children. Vitamin A promotes growth of new cells and aids in maintaining resistance to infection.
The interrelationships of cis- and trans-vitamin A in the rhodopsin cycle in mammalian vision are shown below:
2.1.3 The syndrome of deficiency or excess
Hypovitaminosis A is characterized by poor growth, poor vision, keratinization of epithelial tissue, xerophthalmia, night blindness, haemorrhage in the anterior chamber of the eye, haemorrhage of the base of the fins, and abnormal bone formation. Nerve degeneration has been reported in pigs, chickens, rats, rabbits, and ducks, but only occasionally observed in fish after long periods of deficiency. Hypervitaminosis A has been described in fish and in other animals and involved enlargement of liver and spleen, abnormal growth, skin lesions, epithelial keratinization, hyperplasia. of head cartilage, and abnormal bone formation resulting in ankylosis and fusion of vertebrae. Hypervitaminosis A is reflected in very high liver oil vitamin A content and elevated serum alkaline phosphatase. Removal of excess vitamin A from the diet promotes rapid recovery.
2.1.4 Requirements
Dietary requirements for vitamin A alcohol are listed in Table 3. Fish apparently require vitamin A for growth in light, but not in darkness. Therefore, the requirement for maximum growth and reproduction is related to exposure to light and reflects observations in other animals that near normal growth will occur at very low vitamin A intake in protected environments where fish are not exposed to stress, infection, and ultraviolet radiation.
2.1.5 Sources and protection
Cod liver oil is the best known source of vitamin A although black sea bass, swordfish, or ling cod oils contain much more of the vitamin. Whale liver oil contains kitol which has little or no biological activity until heated above 200° C when one molecule of biologically active vitamin A is generated per molecule of whale kitol. This biologically inactive kitol may be deposited in the whale to avoid hypervitaminosis A during excessive vitamin A intake. The possibility of hypervitaminosis A occurs when tuna, shark, or ling cod viscera are used in preparation of moist fish diets. Synthetic vitamin A preparations, such as vitamin A palmitate, are available and are often used to supplement rations low in fish meal, fish viscera, or carotenes. Some fish species seem able to utilize b-carotene as a vitamin A source, whereas others are unable to split the g-carotene molecule and vitamin A must be added to the diet.
2.1.6 Clinical assessment
Vitamin A status can best be assessed by the absence of deficiency signs and by assay of liver oil for vitamin A content. Assay for the vitamin in blood or plasma has not been found to be useful.
Rickets was induced with test diets by Hopkins, and Mellanby cured the disease in dogs by adding cod liver oil to the ration. Ultraviolet light exposure was shown to have an anti-rachitic effect, and provitamin D activity had been ascribed to ergosterol. Crystalline vitamin D was isolated by Angus and activated 7-dehydrocholesterol was isolated by Windaus.
2.2.1 Chemistry
Vitamin D2

Vitamin D2 or ergocalciferol, is one of several biologically active forms of vitamin D. Vitamin D3 or activated 7-dehydrocholesterol has the chemical formula C27H44O and contains a more simplified, unsaturated 8-carbon side chain. Vitamin D3, also known as cholecalciferol, is formed in most animal tissue by the rupture of one of the ring bonds of 7-dehydrocholesterol by ultraviolet radiation. Cholecalciferol is a white, crystalline compound soluble in fat and organic solvents and is stable to heat and oxidation in mild alkali or acid solution. The conversion of ergosterol to vitamin D2 and several side compounds is shown below.
Conversion of ergosterol to Vitamin D2
2.2.2 Positive functions
Vitamin D is essential for maintaining calcium and inorganic phosphate homeostasis. It is involved in alkaline phosphatase activity, promotes intestinal absorption of calcium, and influences the action of parathyroid hormone on bone. Fish may sequester calcium from water through the gill membrane; thus the major function of vitamin D for other animals may not be necessary to satisfy calcium requirements for fish.
2.2.3 Syndrome of deficiency or excess
Hypovitaminosis D for fish has not been described. Rickets and abnormal bone formation described in detail for animals has not been observed in fish fed low vitamin D diets. However little work has been done under carefully controlled conditions with young growing fish and only alteration in alkaline phosphatase activity has been reported on different vitamin D diet intake. Hypervitaminosis D, however, has been reported. Brook trout fed large doses of vitamin D showed impaired growth, lethargy, and dark colouration. High intake of vitamin D mobilizes phosphorus and calcium from the bone and tissues and may result in fragile bones, poor growth, and poor appetite related to the nausea described in man afflicted with hyper-vitaminosis D. Arterial and kidney lesions due to excessive intake of vitamin D reported for rats and dogs have not been described histologically for fish nor has hypercalcaemia been described. This area needs to be explored because of the potential for hypervitaminosis D in fish fed diets containing various fish viscera which might contain large amounts of the vitamin D. Tuna liver oil may contain, for example, 100 to 1 000 times as much active vitamin D as cod liver oil.
2.2.4 Requirements
No requirements of fish for vitamin D have been described to date, but little work has been done in this area using highly purified diets and controlled experiments with young growing fish. Likewise, no demonstration of synthesis of the vitamin in fish reared on diets devoid of vitamin D or vitamin D precursors has been reported. Therefore, actual requirement of vitamin D for maintenance of calcium-phosphorus homeostasis in the young growing animal has not been adequately investigated and the true requirements of vitamin D in young fish have not been defined.
2.2.5 Sources and protection
Vitamin D requirements of many animals can be met by ultraviolet irradiation of the skin where cholesterol derivatives are converted to calciferol. Since the vitamin is fat soluble and accumulates in lipid stores, fish liver oil is a rich source of the material. Content varies tremendously in liver oil, however, with values of about 25 I.U./g present in shark liver oil and over 200 000 I.U./g in albacore tuna liver oil. Cod liver oil contains from 100-500 I.U./g and animal liver contains some vitamin D. One international unit (I.U.) is equal to 0.025 mg of crystalline vitamin D.
2.2.6 Clinical assessment
Absorption maxima in the ultraviolet region can be used to detect provitamins D in the non-saponifiable fraction of oils. Concentrated preparations of vitamin D can be assayed by the Carr-Price antimony trichloride reaction when assay is necessary to determine biologically active materials in liver oil of fish on different treatments. The chick assay may not apply to fish liver storage levels because the biologically active form for fish has not been determined. Therefore, clinical assessment for hypervitaminosis A must rely on crude methods for determination of vitamin D in liver oil samples by the Carr-Price reaction or by measuring absorption in the UV spectrum.
The existence of an antisterility vitamin was postulated by Evans and Bishop. This factor was named 'vitamin E' by Sure. The active tocopherol was isolated, characterized, and synthesized by Karrer.
2.3.1 Chemistry
Alpha-tocopherol

Vitamin E is composed of a class of compounds known as tocopherols. One of the most important tocopherols is a -tocopherol. Eight naturally occurring tocopherol derivatives have been isolated and all belong to the D series. Synthetic a -tocopherol is a racemic DL-a -tocoperhol mixture. The derivatives of tocol or of tocotrienol are named, alpha, beta, gamma, delta, epsilon, eta, zeta1 and zeta2-tocopherol as shown in Table 5. The pure tocopherols are fat-soluble oils which are capable of esterification to form crystalline compounds. The tocopherols are stable to heat and acids in the absence of oxygen, but are rapidly oxidized in the presence of nascent oxygen, peroxides, or other oxidizing agents. The tocopherols are sensitive to ultraviolet light and are excellent antioxidants in the free form. The esters are more stable and are commonly used as dietary supplements-anticipating hydrolysis in the gut and absorption of the free alcohol to act as an active intra- and intercellular antioxidant. Ethyl derivatives on the aromatic ring are also active. Oxidation products of a-tocopherol can be reduced with hydrosulphite to a -tocopherylhydroquinone or, in the presence of ascorbic acid, to a -tocopherol.
Table 5 The Tocopherols
|
a -Tocopherol (alpha) |
5,7,8-trimethyltocol |
|
b -Tocopherol (beta) |
5,8-dimethyltocol |
|
g -Tocopherol (gamma) |
7,8-dimethyltocol |
|
z 2-Tocopherol (zeta2) |
5,7-dimethyltocol |
|
h -Tocopherol (eta) |
7-methyltocol |
|
d -Tocopherol (delta) |
8-methyltocol |
|
e -Tocopherol (epsilon) |
5,8-dimethyltocotrienol |
|
z 1-Tocopherol (zeta1) |
5,7,8-trimethyltocotrienol |
Oxidation - reduction of a -tocopherol
2.3.2 Positive functions
The tocopherols act as extra and intracellular antioxidants to maintain homeostasis of labile metabolites in the cell and tissue plasma. As physiological antioxidants, these usually protect oxidizable vitamins and labile unsaturated fatty acids. Vitamin E prevents encephalomalacia in chicks, erythrocyte haemolysis in several animals and steatitis in mink, pigs, and farm animals. The tocopherols also prevent exudative diathesis in chicks, white muscle disease in lambs and calves, and dietary liver necrosis in rats. They act with selenium and with vitamin C for normal reproductive activity and are involved in prevention of nutritional muscular dystrophy in the chick, the yellowtail and carp. The tocopherols act as free radical traps to stop the chain reaction during peroxide formation and stabilize unsaturated carbon bonds of polyunsaturated fatty acids and other long-chain labile compounds.
Vitamin E is involved in the maintenance of normal blood capillary permeability and the integrity of heart muscle. It was first shown to be involved in prevention of sterility and foetal resorption in rats. It may likewise affect embryo membrane permeability and hatch-ability of fish eggs.
2.3.3 Syndrome of deficiency or excess
Deficiency signs in fish are listed in Table 2. One of the first signs is erythrocyte fragility closely followed by anaemia, ascites, xerophthalmia, poor growth, poor food conversion, epicarditis, and ceroid deposits in spleen and liver. Muscle dystrophy and xerophthalmia have been described in yellowtail and carp. Non-specific forms of degenerative conditions have been described in several species of fish fed large quantities of polyunsaturated fatty acids with inadequate tocopherol in the ration. Hypervitaminosis E results in poor growth, toxic liver reaction, and death.
2.3.4 Requirements
Requirements of fish for vitamin E are shown in Table 3. The exact requirement of the fish for a -tocopherol may depend upon the amount and type of polyunsaturated fatty acids content of the ration. Polyunsaturated labile fish oils may invoke an increased requirement for intracellular antioxidants.
In addition, the amount of tocopherol needed as a supplement to the ration will depend upon the form of the vitamin used, the method of diet preparation, and the storage conditions before feeding.
2.3.5 Sources and protection
Vegetable oils are rich sources of tocopherols. Synthetic a -tocopherol in esterfied acetate or phosphate form is commonly used as a diet supplement. These esters are much more stable than the free form which is rapidly lost by air oxidation or in the presence of labile compounds like the polyunsaturated fish oils. Addition of antioxidants such as BHT (butylated hydroxyanisole) and BHT (butylated hydroxytoluene) protects fats and other labile compounds in the ration from oxidation, but these antioxidants have no vitamin E activity.
2.3.6 Clinical assessment
The erythrocyte fragility test indicates the physiological state of fish. Absence of histologically detectable ceroid in liver and spleen from representative samples in the population is a good clue on the presence of adequate amounts of physiological antioxidants in the fish. A barbituric acid test for oxidation of components has not been applied to fish tissues as a clinical tool for assessment of nutrition state except when liver oils become saturated with very labile polyunsaturated fatty acids such as when feeding squid or saury oils. These assays are good indicators of the state of oxidation of the finished ration, but the absence of deficiency signs and normal erythrocyte fragility are better clinical tests. Analysis for tocopherol is difficult and time-consuming and only applicable under critical research experiment situation.
The name vitamin K (for 'koagulation') was proposed by Dam. Dam isolated the vitamin from alfalfa and from fish meal in 1939. It was synthesized later that year.
2.4.1 Chemistry
Vitamin K2 series; n may equal 6, 7 or 9 isoprene units

The K vitamins K2 contain 6,7, or 9 isoprene units in the side chain which varies from 30-45 carbon atoms. Many isosteres of vitamin K have been identified in animal tissues, plant tissues, and micro-organisms. The structure of phthiocol and menadione (K3) are shown below:
Pthiocol

Menadione

Although fairly stable compounds, they are destroyed upon oxidation and exposure to ultraviolet radition. Menadione is very reactive and is subject in aqueous media to chemical interaction.
2.4.2 Positive functions
Vitamin K is involved in the hepatic synthesis of blood clotting proteins - prothrombin and proconvertin. A simplified scheme of blood coagulation is shown below:
Substituted forms of vitamin K are strongly bacteriostatic and may serve as an alternate defence mechanism for bacterial infections. Vitamin K is structurally related to flavo compounds which function as electron carriers in the electron transport system. The primary role of vitamin K is to maintain a fast normal blood clotting rate which is so important to fish living in a water environment.
2.4.3 Syndrome of deficiency or excess
A summary of deficiency signs is listed in Table 2. Prothrombin time in salmon fed diets devoid of vitamin K was increased three to five times and, during prolonged deficiency states, anaemia and haemorrhagic areas appeared in the gills, eyes, and vascular tissues. Increased blood clotting time has also been reported for other fish reared on diets with low vitamin K content. Interrelationships with other vitamins have not been documented in fish where the primary deficiency signs are slow blood clotting and haemorrhage, severe anaemia, or death in wounded fish. Haemorrhagic areas often appear in fragile tissues such as the gills.
2.4.4 Requirements
Nutritional deficiency signs in fish occur within 10-14 weeks. Quantitative requirement studies have not been completed are needed in order to define the vitamin K-requirement of rapidly growing young fish, especially those held in high population density under intensive fish husbandry conditions.
2.4.5 Sources and protection
Vitamin K sources are green, leafy vegetables. Alfalfa leaves are among the best sources of vitamin K. Low levels are found in soybeans and animal liver. Synthetic menadione is also available. Although vitamin K in ground alfalfa is fairly stable, synthetic material should be protected from exposure to ultraviolet light and to excessive oxidizing or reducing conditions. The use of rapidly cured dehydrated alfalfa is essential to minimize formation of the physiological antagonist, dicumarol. The diet should be kept dry, prepared with minimum exposure to air oxidation, and fed as soon as practicable after manufacture to minimize vitamin K loss.
2.4.6 Antimetabolites and inactivation
Two vitamin K antagonists, dicumarol and warfarin are shown below:
Dicumarol

Warfarin

Dicumarol was isolated from spoiled sweet clover hay and shown to prevent the normal function of vitamin K in maintenance of normal blood clotting times. Early work was with cattle and hogs but was soon extended to rats, other experimental animals, and man. Dicumarol is an anticoagulant and has been used to prevent thrombosis in animals and man. It is not an antimetabolite competing for vitamin K sites but plays another role in preventing normal blood clotting. Vitamin K counteracts the dicumarol effect quantitatively. Another anticoagulant, warfarin, is a common rat poison which has five to ten times the anticoagulant activity of dicumarol. The effects of warfarin can also be reversed by administration of vitamin K.
2.4.7 Clinical assessment
Vitamin K status of fish is determined by measuring blood clotting time. Chemical methods for determining menadione content of active metabolite after storage are available but have not been applied to measure vitamin K activity in the liver or tissues of young, growing fish.
![]()
![]()
![]()

{kind=link}
{kind=link}
{kind=link}
{kind=link}