![]()
![]()
![]()
1. IDENTITY
2. DISTRIBUTION, ECOLOGY AND METABOLISM
3. LIFE HISTORY
4. POPULATION STRUCTURE AND MORTALITY
5. PRODUCTIVITY OF THE RESOURCE
6. METHODS OF HARVESTING AND HARVESTING CYCLE
7. EQUIPMENT USED FOR HARVESTING AND CULTURE OF EUCHEUMA
8. PROTECTION AND MANAGEMENT OF THE RESOURCE
9. UTILIZATION
10. REFERENCES
by
Maxwell S. Doty
Department of Botany
University of Hawaii
Honolulu, Hawaii
|
ABSTRACT Three commercial kinds of Eucheuma are being used for their carrageenans. The three kinds, the annual production of each and their specific carrageenans are, respectively: cottonii, 30,000 tons producing kappa carrageenan; spinosum, over 6,000 tons producing iota carrageenan; and gelatinae, about 100 tons producing a mixture of gamma, beta and kappa carrageenans. The principal carrageenan uses are as suspensoids in dairy, luxury human and pet foods and drug products. Over 95% of the annual commercial Eucheuma crop is from farms in the tropical far western Pacific. In addition to foreign exchange earnings for those countries exporting the seaweed and the final use values of its carrageenans, the labor-intensive farming of Eucheuma is of great socio-economic value to the often nearly indigent shore dwelling families who grow it. |
1.1 Taxonomy
1.2 Nomenclature and terminology
1.3 Morphology and anatomy
Eucheuma production from wild crop sources has dwindled to insignificance, and over 95% of the crop is farmed E. alvarezii Doty (as cottonii), E. denticulatum [Burman] Collins & Hervey (as spinosum) and E. gelatinae [Esper] J. Agardh (as gelatinae). Only the latter is predominantly from wild crops. The others are almost all farmed.
The taxonomy of the Eucheuma section Cottoniformia, which produces the cottonii of commerce, is most difficult due to the wide range of environmentally induced forms. This was not realized until strain selection commenced in the early 1970's (Doty & Alvarez 1975) and one kind, Eucheuma alvarezii var. tambalang Doty, was found to be consistently different physiologically and to have its own gamut of variation in form. It proved to be so much more easily grown that it has completely replaced the other species, such as E. striatum (elkhorn variety), formerly dominating the farming.
Likewise, many of the former wild crop components of commercial lots of Eucheuma were either never farmed (such as E. cottonii), are no longer farmed (e.g., E. striatum), or are not now recognized (e.g., E. spinosum, E. edule, E. muricatum, E. okamurae and E. serra) (Doty & Norris 1985).
Table 1. provides the key characteristics of the three commercial species. They are illustrated in Figures 1, 2, and 3. Doty & Norris (1985) provide a key to the commercial species and those sometimes seen in wild crop lots of Eucheuma.
Table 1. Distinguishing Characteristics of the Three Kinds of Eucheuma Currently in Commerce. A common and a scientific binomial name are given for each. The presence and nature of special core tissues usually requires microscopic study.
|
|
Morphological attributes |
| |
|
Common names and the species of Eucheuma |
Branching |
Axis & habit |
Carrageenan and sulfate content |
|
SPINOSUM E. denticulatum (Figure 1) |
Whorled or at predictable intervals |
Terete, erect w/a rhizoidal core |
Iota. 3 0 % sulfate or over |
|
COTTONII E. alvarezii (Figure 2) |
Irregular |
Terete, erect & w/o rhizoidal core |
Kappa. 28% 1 sulfate or less |
|
GELATINAE L. gelatinae (Figure 3) |
bi-lateral or dorsi-ventral |
Flattened & erect to procumbent w/a rhizoidal core |
Gamma, beta & kappa 20% sulfate |
Fig. 1. Eucheuma denticulatum branch showing the cylindrical axis with rings of spinous primary initially-determinate branches, some of which have become indeterminate but are yet small.

Fig. 2. Eucheuma alvarezii branch showing its cylindrical axis with branches that are commonly enlarged maximally just beyond a basal curvature toward the light.
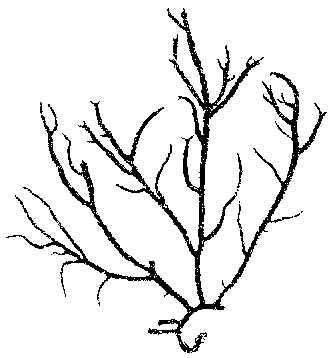
Fig. 3. Eucheuma gelatinae showing the flat branches arising from marginal cylindrical teeth. The scale bars represent 3 centimeters.
The first name applied to a Eucheuma was Fucus denticulatus N. L. Burman in 1768 and the use of this specific epithet is continued today as E. denticulatum (Burman) Collins & Hervey 1917. That Linnaeus (in 1771) sought to substitute Fucus spinosus for Burman's name, which has been used as E. spinosum (L.) J. Agardh, has no standing (Dixon 1962) in view of the priority of Burman's epithet, a priority acknowledged by Linnaeus himself.
Eucheuma was first distinguished as a genus and named by J. Agardh (1847), who included seven species one of which was E. "spinosa" based on Sphaerococcus spinosus C. Agardh, in turn based on earlier species including Fucus spinosus Linnaeus and F. muricatus Gmelin, in turn based (Dixon 1962) on Burman's yet earlier F. denticulatus. The Lectotype is the specimen in the Linnaean Herbarium as #1274-104 and illustrated by Turner (1808) as Figure "a" of his Plate 18.
There is no question as to the nomenclature of this species, Eucheuma alvarezii Doty (1985). The Holotype is in the B.P. Bishop Museum in Honolulu, Hawaii, bearing the author's voucher number 28727.
Among the species originally included in Eucheuma by J. Agardh (1847) was E. gelatinae, the specific epithet for which was first applied by Esper(1797) in the combination Fucus gelatinus. Thus today one uses E. gelatinae (Esper) J. Agardh for this third commercial entity. The Lectotype is Esper's material in the Hamburg (Germany) Herbarium.
The thalli of commercial Eucheumata are often up to a kilogram in mass but in one form of cottonii they have been weighed at 56 kilograms alive. Basically it is a multiaxial filamentous red algal genus which becomes strongly pseudoparenchymatous. The nature of their triphasic life history results in their relegation to the family Solieriaceae of the order Gigartinales as these taxa are defined by Kylin (1956) and more recent authors.
The species vary greatly in form as a result of the environments in which they grow. This is particularly true of those in the section of the genus to which the cottonii forms belong. Commercial spinosum and cottonii are composed of cylindrical branches that are rigid. Mature gelatinae branches are apically flat, somewhat flexible, have marginal teeth and arise from a cushion of strongly compressed branches.
Gabrielson (1983) and Gabrielson & Kraft (1984) provide much of the structural detail which Kylin (1956) does not include. None of these authors treats the sections of the genus, Anaxi-ferae, Cottoniformia or Gelatiformia, from which the commercial cottonii and gelatinae crops arise. Weber-van Bosse (1928) and
Doty (1985) provide some of the structural details of the section Cottoniformia but they are still largely undefined anatomically. (See Section 4.5).
2.1 Geographical distribution
2.2 Local horizontal and vertical distributional determinants
2.3 Effects of non-distributional ecological determinants
2.4 Environment and growth
2.5 Life form versus environment
The two major commercial forms, spinosum and cottonii, are native to the Old World Tropics (Weber-van Bosse 1928; Laite & Ricohermoso 1981) westward to East Africa (Anderson 1953; Mshigeni 1982, 1984). A very small amount of the third, gelati-nae, comes from the Philippines and China (Hainan Island and Taiwan). These three species of the genus are almost entirely restricted to the same brightly lit waters (Figure 4) in which coral reefs form. Eucheuma is now being commercially produced in the far western Pacific. Cottonii has been introduced eastward through Micronesia into Kiribati and Tonga to the Society and Hawaiian Islands.
In some seasonally substropical/temperate parts of the Southern and Northern Hemisphere, e.g., in Australia, Florida (U.S.A.), the Gulf of California (Mexico), and southern Japan, a variety of non-commercial species may flourish during the warmer brighter times of the year. The anticipated micro-forms persisting during the off-season are not known (See Section 3).
Tolerance to aerial exposure appears to govern the upward distributional limits of the Eucheuma species, although otherwise, the brighter the light the faster they grow. Possibly the tolerance of spinosum is greater than that of cottonii and gela-tinae is the least tolerant. Since Eucheuma grows almost entirely in open ocean water in areas of high water motion, light and temperature are not significant factors in local distribution. Water motion is most important. In some cases there may be some influence of run-off from land. Spinosum especially appears to be sensitive to water quality.
Both the commercial wild and farmed kinds of Eucheuma are mostly found in water less than a half meter deep at extreme low tide level. The greatest depth recorded is that of Weber-van Bosse (1913), 26 fathoms. For Eucheuma, deep water habitats are of no commercial importance.
Tides of greater range allow less working time at a site for either wild crop harvesting or farming than do tides of lower range. Also tides may exert an influence on productivity through light intensity and water motion control. Thus generally commercial production is in areas with low tide ranges.
Pigmentation is characteristically more or less dense at brighter light levels. Thalli may be black in intense light and relatively redder in the shade or in deeper water due to relative abundance of phycoerythrin. Pale yellower thalli are found in some bright light conditions.
Some cottonii forms may become negatively phototropic in intense light, and the spininess of some spinosum forms may be reduced to bilaterality, and the axial segments may then suffer some dorsiventral compression.
Cottonii thalli exposed to air tend to form "heads," or become prostrate and dorsiventrally flattened. Bushy thalli are formed in turbulent water, and trailing cottonii forms are found (Doty 1985) where current dominates. Cottonii may form erect cane-like forms seasonally in pools or deeper water.
At greater depths the species may change in morphology as though etiolated in a phycological sense. See Dawes (1981, Chapter 14) for the basic physiological nature of the responses expected of such a seaweed as Eucheuma as its extremes of vertical distribution are approached.
Cottonii and spinosum are composed of cylindrical branches (Figures 1 & 2) which, in cottonii, sometimes reach diameters of over two centimeters. The rigid and bush-like thallus form enhances thinning of the boundary layers wherever they are subjected to water motion. Thus, since materials move into and out of algae such as Eucheuma by diffusion, their growth increases as water motion increases, up to the point where drag or force produces excessive physical damage. Only gelatinae tolerates direct wave action. It is (Figure 3) the smallest of the three, and the fronds are apically flat and pliable.
Commercial production is from level sedimentary bottoms that are under water except during the lowest of tides. Level sedimentary bottoms are desirable for they are uniform and easily workable whereas sloping or irregular areas are not. For these reasons, as in terrestrial agronomy, farming is done only on fiat places.
Local horizontal and vertical distribution are correlated with the variations in the above factors. The chemical nature of the substratum is of little importance. The physical nature of the substratum is important as a stable anchorage for wild crop thalli. Its stability correlates with water movement, which, in turn, is essential (See Section 2.5) physiologically. In the Hawaiian experiment (See Section 2.4) water movement was found to be the most important environmental factor (Doty 1979) influencing growth. (See Section 5.2).
A few laboratory respirometric studies of Eucheuma have been done, but only some of them related to micro-nutrient water content; e.g. by Dawes et al. (1974a & b), Mathieson et al. (1974) and Glenn & Doty (1981). For the commercial forms of Eucheuma, beyond the results in Section 5.2, there have been no quantitative simultaneous measurments of seaweed growth and environmental quality; thus, to date no statistically valid cause-and-effect relationship has been experimentally demonstrated between, e.g., Eucheuma growth or hydrocolloid quality and any putatively causal micro-nutritional element in the farm environments. Clear water and stable salinities (of 29 to 34‰) are desirable.
Temperature and light are felt to be relatively independent of water motion and its quality. It is not economic for farmers to provide control of these factors directly, so in selecting sites it is essential all be within tolerable limits. Water motion especially is a factor that must be taken into account in selecting sites. It often affects sediment in the water, which, in turn, affects light.
Mineral nutrition is surely important in view of the high mineral content of the species. Table 2 provides various cation as well as nitrogen and phosphorus, values for the commercial species. They are similar among other Eucheuma species. Unfortunately, the variation in these elements in different commercial lots is not publicly known. Arsenic is below the U.S. Federal Drug Administration limits.
Table 2. Inorganic Cation Plus Phosphorus and Nitrogen Concentrations in Different Eucheuma - related Materials. The cation determinations were made by atomic adsorption methods in replicate. The means shown for K, Na, Ca and Mg are in terms of per cent dry weight; Zn, Cu, Fe and Mn are given as parts per million dry weight. Phosphorus and nitrogen are in terms of percent dry weight. The values for Fe were from thalli ground in porcelain.
|
Cations plus nitrogen and phosphorus | |||||||||||
|
Source |
K |
N |
Na |
Ca |
Mg |
P |
Zn |
Cu |
Fe |
Hn | |
|
A- THALLI rinsed and redried free of surface salt | |||||||||||
|
Spinosum |
11.91 |
0.46 |
4.79 |
0.589 |
0.758 |
0.043 |
13 |
7.5 |
66 |
14 | |
|
Cottonii |
9.33 |
1.19a/ |
3.97 |
0.234 |
0.619 |
0.030 |
16.5 |
2.5 |
52 |
4 | |
|
B- SALTS as crystals washed from thallus surfaces | |||||||||||
|
Cottonii |
52.82 |
--- |
0.290 |
0.120 |
0.027 |
0.012 |
1.1 |
6.5 |
18 |
5 | |
|
Spinosum |
36.52 |
--- |
10.45 |
0.240 |
0.15 |
nil |
18 |
1.1 |
24 |
1.1 | |
a/N value was that of a different Sample No. 26551.
In an unpublished study the fixed nitrogen and dissolved phosphorus contents of the water, as well as other factors, were determined weekly as the water passed through about 15 meters of (usually) dense Eucheuma planting. Among the results (Table 3) there appeared to be a reduction of 24% of the nitrate/nitrite ion as the water passed through the seaweed and 6% of the phosphate. This provided the material used by Glenn & Doty (1981) and is spoken of below as the 55-week Hawaiian experiment or study. In a different (unpublished) study the average N/P ratio in 9 species of the genus was near 15 though perhaps 12 or 13 was the mode. Thus, in the present case, the N/P ratio in the water of 8.5 may indicate non-detection of ammonium ion uptake due to its being produced (or recycled) by the animal population that had become resident in the planting.
Table 3. The Hawaiian Eucheuma Experimental Habitat. The values are mean microgram atoms per liter from all weekly measurements made over the 55 weeks of a 1976-77 study period.
|
Sample site |
Water motion |
Micro-nutrients |
||
|
NH3-N |
NO3+NO2-N |
PO4-P |
||
|
Upstream |
|
1.98 |
1.44 |
0.66 |
|
Downstream |
|
2.01 |
1.10 |
0.62 |
Fertilizing is not thought to be economically worthwhile though it does provide some increases in productivity.
Water movement of all kinds is of major significance. It affects growth in such ways as change in mass (growth rate), differentiation (form), reproduction and production of the hydro-colloid materials for which Eucheuma is prized. These topics are discussed above and in Section 5.2, below.
In the commercial Eucheuma species, the relationships of form to environment have not been described adequately, and they are only cursorily mentioned (Doty 1985; Doty & Norris 1985) with respect to their cause and effect relationships. Yet these relationships are important for farm production. For example, in regard to the space occupied and diffusion as a whole, the Eucheuma thallus is analogous in its relationships to the (sea) water in the same way the root system of a terrestrial flowering plant is related to the soil water. Unpublished results indicate that, in a desirable commercial Eucheuma habitat, the water in a one-liter thallus space may be laterally changed over a half million times a day.
The wild crops appear to follow the triphasic Polysiphonia life history scheme common for red algae. A diploid vegetative phase (Phase 1) produces haploid non-motile spores called tetra-spores. The tetraspores produce haploid gametophytes (Phase 2) that in turn produce diploid carposporophytes parasitic in the female thalli. The carposporophytes (Phase 3) release diploid carpospores which initiate the diploid tetrasporic stage again.
The details of the reproduction of Eucheuma from a classical point of view are readily available in Kylin (1956), and considerably updated by Gabrielson (1983) and by Gabrielson & Kraft (1984). Superfically spinosum, cottonii and gelatinae all appear to be triphasic though the strains being farmed may not be.
The contumacy of the species of red algae for the above described triphasic life history is well known to those who have sought it. There is no detailed study of the life history of any member of this genus. Male thalli are as yet unknown for the commercial forms. Santos & Doty (1978) found both cystocarpic and tetrasporic thalli in quantity in only 6 of the 15 species in which they were sought. They did not find them common in the commercial species and found no male thalli. Shallow water, line-farmed Eucheuma (and, incidentally, pond-grown Gracilaria thalli) are almost always sterile. While this may be a blessing to the marine agronomist and farmer in some ways, it hinders utilizing the advantages of sexual reproduction in improving the crop.
Eucheuma alvarezii carpospores produce morula-like tetra-sporophytic embryoes (Figure 5) that attach by rhizoids and differentiate an erect pyriform, then cylindrical form as the typical cluster of apical cells develops opposite the substratum. The younger embryos have a thick gel coat and a large number of very fine hairs, each ten or twenty times longer than the diameter of the embryo. Perhaps they may represent the micro-form in which some species of the genus persist in "off" seasons.
The Eucheuma farmed crop is produced without regard to the life history phase concerned. Prices paid to the commercial producers is the same for the two macroscopic stages.
4.1 Population structure of wild crops
4.2 Population structure of farmed crops
4.3 Morbidity, mortality and health
4.4 Grazing
4.5 Storm and other mechanical damage
4.6 Wound healing, regeneration, and secondary attachment
Eucheuma farm production is by vegetative propagulae grown to harvest size on farms from stocks that appear to live indefinitely. Therefore the conventional phenomena of population structure and mortality have little meaning.
Nothing has been published on the population structure of the communities in which Eucheuma appears. Overharvesting appears (Doty, 1973a,b; Doty & Alvarez 1973; Parker 1974) to cause wild crop population declines. Exports from the Philippines climbed in the 1960's to at least 800 tons in 1966 but fell in the late 1960's (Doty 1973a/b) and the early 1970's (Doty 1978) to less than one fourth of that amount. At that time most of the world's supply was coming from the Philippines. Earlier, when the crops were nearly all coming from Indonesia, the wild crop harvesting was not nearly as intense and apparently at most places well below the level of maximum sustained yield. Yet it was observed in two places along the southwest coast of Sulawesi (Indonesia) during this time that such harvests were bringing in spinosum of very small sizes that had been harvested from between finger coral branches as though all possible was being picked.
A nearly ten-fold decrease in wild crop density, seemingly related to over-harvesting, appears to have taken place (Table 3 in Doty 1972) at least at one time in the Sulu Sea area. A great deal of community composition information, including the masses of other seaweeds found in the random ring-toss samples, is available in the same data report and in Kraft (1969), a similar raw data report. There is almost nothing more, either published or in the grey literature on the commercial species comparable to that published on E. isiformis by Dawes et al (1967, 1974a,b) or in the brief study by Kraft (1972) on E. arnoldii and neither of these has data that imply cause-and-effect relationships. Kraft does describe E. arnoldii var. alcyonida as (loosely quoting) being restricted to association with the branched coelenterate corals in silt-free, rapidly moving water near the seaward margins of reefs.
Kraft (1969) provides a wealth of information on the communities in which commercial Eucheuma species occur, based on one or a few observation periods at many Philippine sites over a wide geographic range. Doty (1972) describes repeated observations of the same populations of commercial species relative to the non-Eucheuma seaweeds at some 31 sites just in the Sulu Archipelago. These two appear to be the largest sources of community information on the genus.
Farming quite naturally was developed as a result of over-harvesting and to obtain desirable size, stability and quality of crop. Farming routines soon result in very uniform populations. The very valuable work of Lim et al. (1982) at one site for a good many months is the major source of published information.
No attempt is being made here to describe farm siting, construction and operation beyond what is to be found in other sections of this case study. The early history of the farming (Doty 1973a,b; Doty & Alvarez 1973, 1975, 1981; Parker 1974; Deveau & Castle 1976; Lim & Porse 1981) provides the essential generalities. Each farming area displays characteristics which are to a large extent the result of field expediency.
Farms nowadays consist (Figure 6) of parallel rows of stakes bearing nylon monofilament line to which there are tied branches of Eucheuma broken from previous farmed, or from wild crop, thalli. For good growth the habitat is especially important in reference to bottom elevation, water movement, the nearness and nature of the bottom materials and the pests that may be in the area. The successful farms are over sedimentary bottoms with the thalli near or on the bottom and where problems such as those discussed in the following subsections are minimal.
In experiments, individual vegetative thalli of cottonii have lived for 14 months, and one farmed population is known to have thrived by vegetative cuttings for over 10 years. Farmed strains of spinosum and gelatinae have not been as closely observed but they seem to have prospered for years.
Thallus colors and growth rates are often considered indicative of the health of a planting. There is no finished study of the former but it is well known that yellowing indicates a lowering of vitality in the red algae. In Chondrus this is sometimes associated with the production of a higher yield of stronger gel and can be (Neish & Shacklock 1971) artificially induced by holding the live thalli in high light and low mineral nutrient water for a few days before the final harvesting. (See Section 2).
It is practical in test plots to use the coefficient of variation (mean growth rate/standard deviation) as a comparative measure of population health. This value, C, may be high due to genetic variability in the population or due to uneven grazing, responses to environment, different ages of seedstock branches orientations and exposures of the thalli. Values of 0.20 are good, less than 0.10 excellent.
The lack of healthy growth, very low rates of growth and lack of reproductive fertility may be due to environmental imbalances or agronomic practices. It must be recognized that farms are often located where Eucheuma does not occur naturally, and this may not always be just due to a lack of suitable substratum. The major cottonii form, tambalang, may not become fertile on farms, though the typical form is freely fertile in its native habitat. Whether this is due to its genetic constitution or to farm practice is unknown, but juvenile thalli are rarely if ever seen. Juvenile spinosum thalli are often seen on farms.
Other specific poor health situations seen on farms are discussed below. (Gross physical damage is included under Section 4.5).
The most common symptom of malaise, ice-ice, is often spoken of as a disease. Actually it is only a symptom. No microorganism-caused or contagious disease is known (Uyenco et al. 1981). Infections and infestations are known. In itself the word ice-ice is (Doty & Alvarez 1975) a malay treatment of the Middle English word, ice, applied to the white segments that appear between branches usually in the more basal parts of the thalli.
Ice-ice seems to be related most often to intolerable physio-chemical stresses, such as increased ceil permeability following low growth rates or relatively low counter-movement of photosynthate. The onset is a sharply limited greening of a segment which, the next day, is clearly green and in one or very few more days very pale, then white. The segment may remain there for a day or two but soon dissolves away, separating the two adjacent parts of the thallus, which seem to be otherwise unaffected. In this way ice-ice acts as an agent in vegetative reproduction: the ice-ice does not normally spread to the adjacent segments. The two such thalli resulting continue their normal growth though one is unattached. Of course, the one not attached may become lost to the farm.
A second malaise is "pitting," the formation of a cavity penetrating the cortex in one place and expanding in the medullary regions beneath. It can lead to thallus breakage. This phenomenon has been studied by various students and by Dr. Gordon Sill (unpublished personal communication) who carried out Koch's postulates with pure cultures of the bacteria isolated from them. In no case has the pitting been obtained without a mechanical wounding of the cortex. In time the pit surfaces become covered by regenerated cortex and are then the same color and nature as the adjacent natural cortex. At least if small, the pits do not adversely affect the thalli. It does not spread.
A third malaise is "tip darkening," relative to the adjacent segments below the affected branch apices. This may well be due to senescence or cold weather. In time the lower parts of the thallus so affected may lose color and disintegrate; the tips fall to the bottom. Isolated, or after a change in environmental conditions, the tips may resume growth and regenerate new thalli of normal size and appearance.
A fourth malaise is "tip discoloration" relative to the adjacent segments below the affected apices. This is usually a change toward pinkness and eventually the tips soften, discolor further, become white and dissolve away. It is as though respiratory metabolism were affected and usually has been thought to be (unpublished) related to intolerably warm water or aerial exposure
A fifth phenomenon is a slowing of growth, the appearance of epiphytes and usually the onset of pigment loss followed by a general decay through softening of all tissues. Possibly this is most often due to intolerable changes in water motion, salinity or mineral content such as appear at some sites seasonally or randomly. Moving the thalli to higher water-motion areas usually offsets this if done soon enough. It indicates a poor season, place or way of farming.
In some cases, general decay has been seen to result in the death and disappearance of ail but the very oldest basal, often horizontal but unattached, segments. These may remain alive though reduced to an unbranched cylinder. If the environment becomes favorable again such segments may generate a plethora of vigorous shoots and regenerate a new thallus of normal form.
In one case in Sitangkai (Philippines) a great deal of die-off occurred, with the thalli displaying the above general discoloration symptoms. Similar losses have occurred in the Malay-sian part of the Sulu Archipelago. This general die-off phenomenon has been thought due to pools of fresher water forming off the mouths of the large rivers that drain into the sea along the north shore of Borneo (i.e., in the Malaysian State of Sabah) and, breaking away, drift so as to temporarily cover reef-flat farm areas to the east and south. In some dialects of the region this phenomenon is apparently referred to as HAUS, pronounced as is the English word for a residence. (See below under micro-grazers.)
Grazing organizms can be recognized as individual species, with some being categorized as micro- forms and others as macro-forms. Yet, in reality, they form the continuum illustrated through the following two subsections. In different places and different times different ones of them are a problem. Some that may prevent farming in one place will be completely absent in another place.
4.4.1 Micro grazers
Micrograzing occurs when organisms, generally less than 2 cm long, both take up residence on a thallus and consume the thallus materials in some way. It has not been studied in Eucheuma. In the slowing of growth, described above, there have been cases of nematodes attaching in great numbers to the decadent thallus surface. Ordinarily micrograzers are not visible on healthy Eucheuma.
Micrograzing forms could be defined to include certain sizeable echinoderms that are sometimes bad pests in Eucheuma farms. There are two types in this category. The first is a Tripneustes that arrives in planktonic form and settles on the seaweed. As it grows it becomes visible, orange, and up to perhaps 5 or 6 cm in diameter. In serious infections there can be at least one of these sea urchins developing to adulthood in almost each Eucheuma thallus. Their grazing hollows the central area of the thalli affected. As they sever major branches, those branches not tied to a farm line fall from the farm planting (Figure 6) and may be lost.
The second type of similarly arriving micrograzer that becomes a macro-organism is exemplified by synaptid holothurians. In Hawaii, as Ophiodesma spectabilis, they arrive as net piankters and become up to 2 cm in diameter and half a meter long. These are first obvious as residents of Eucheuma thalli when they are soft pink objects just above a centimeter in length. They soon grow to be sizeable, adherent but otherwise innocuous, snake-shaped animals that may slip Eucheuma branch tips into their "mouths" where the seaweed disintegrates.
4.4.2 Macro-grazers
These grazing organisms arrive as adults, and they are usually 5 or more centimeters in their major dimension.
The first macrograzer is the common asteroid echinoderm, Protoreaster nodosus, which climbs into Eucheuma thalli when this alga is farmed on microalgal-dominated (as to color) sedimentary bottoms. Once there it everts its stomach over the branches, and the branches die where they are covered. This often causes major branches to separate from the part secured to the farm lines and become lost, as do branches separated by ice-ice. This can be a bad pest, but it appears to be restricted to near or within sea grass, Enhalus acoroides or Thalassia hemprichii, communities. The activities of Protoreaster have been described (Thomassin 1976) in regards its microalgal herbivory.
The heterospinous long-spined "poisonous" sea urchins, usually Echinothrix or Diadema, are notorious pests. They combine the ill effects of the above-mentioned Tripneustes and Protoreaster with the threat of injury to the farmer who may try to remove them. Fortunately their populations are usually downstream (on falling tides) from the best farm areas.
Fishes of different kinds browse on Eucheuma avidly. The rabbit fishes (Siganidae) and puffers (Tetraodontidae as well as Canthigasteridae) are common. The former are by far the most destructive. When about two centimeters long they appear as dark schools by the "millions" (often with the longest days of the year) and mow the diatoms off the edges of such eel grasses as Thalassia. As they grow larger they will turn to any Eucheuma available. The schools are gradually decimated as the individuals lengthen. Medium-sized Siganus species will attack a thallus here and there among the many on a farm and remove the normally reddish to greenish olivaceous brown cortex leaving a still-living white skeleton which, as though displaying a heroic ice-ice symptom, disintegrates in a few days. By the time the schools of thousands are reduced to scores; perhaps 4 to 5 months after their initial appearance as vast numbers of tiny individuals/the siganids nip off the branches beginning at the tips and continue this grazing until some inhibiting diameter is reached...perhaps related to the size of the fishes in the local school. After some months the remaining few rabbit fish are timid and, being fast agile swimmers, are usually seen from above the water surface as swiftly darting shadows. Though by now relatively few, they can still destroy a Eucheuma farm. Human presence and active overfishing may well control them.
The grazing of fishes can reduce dense Eucheuma beds to a few large-diameter segments that persevere as gnarled relics among the bottom debris. Russell (1983) has provided an often perceptive and quantitative study of this and related non-Siganid fish grazing phenomena. He reports the fish eating as much as 20 tons per month of live cottonii from a patch about 3 meters horizontally wide and 500 meters long.
Turtles are the largest of the macrograzers of concern. They pose a special problem for two reasons. First; it is usually the green turtles that are the problem, and they seem to take large bites and crawl over a farm, thus devastating their way through it. Secondly, the farmers pay no attention to the purported endangered species nature of this (or other) species. Human feelings about turtles are very strong in different places in very different ways. In some places the turtles are considered in a class with pork and therefore anything to do with them may be rejected. In some places they are traditional parts of particular celebrations. In yet other places they are avidly sought out and eaten.
For the most part, mechanical damage comes about as a result of a few discrete causes. Water motion tends to move the thalli about and, especially when they are large, this can cause branches to become abraded or break off entirely and be washed away. Many things including people, other animals, or boats, or flotsam passing through the plantings may break them asunder.
Damage, whatever the cause, is soon repaired by production of a new cortical layer covering the wound. Branches may regenerate from this new surface, if brightly lit. Near preexisting meristoderm or axial tissue, one or more cellular lumps may appear on the new cortical layer and differentiate a central tuft of apical cells and, then, a new branch.
Secondary attachments are normally formed when an apical cell tuft is held against a surface. Spinosum has many tips at right angles to the stems on which they are borne and is often seen with apical attachments to a wide range of objects including other Eucheuma thalli. Cottonii tips are mostly directed toward the light, parallel to each other and thus away from surrounding objects; thus its having secondary attachments is not common.
Wound healing from breakage or abrasion, e.g., by being rubbed against a coelenterate coral, is rarely seen to have led to attachment directly. Russell (1983) found that where corals and algae had rubbed together due to water motion, the corals more often grew around the seaweed than visa versa.
As a result of heavy grazing or nearly lethal conditions, large-diameter short segments of Eucheuma thalli may remain alive and fall onto the bottom, where their surfaces regenerate. During a successive favorable growth period these old gnarled segments may produce hosts of new branches quickly. Such regeneration is described by Russell (1983) when there is respite from grazing. (See Section 4.3).
5.1 Global tonnage and dollar productivity
5.2 Natural determinants of Eucheuma production
5.3 Genetics and high technology in Eucheuma production
5.4 Relative contributions of sexual and vegetative reproduction
5.5 Anticipated agronomic improvement of Eucheuma production
Productivity data in terms such as world crop, number of families concerned or dollar values are very scarce or nonexistent, and those discussed below are much of what is available. There are even fewer published values for productivity in terms of tons/ha/year for even one site, but Table 4 provides a few of the earliest.
Table 4. Eucheuma Growth Rates. E. gelatinae plus three other kinds, i.e., spinosum, elkhorn and tambalang, which are believed to be referable to E. denticulatum, E. striatum and E. alvarezii, respectively; with the latter two being cottonii. The many measurements of Lim et al. (1982) and Doty (unpublished) made in the field are not summarized here.
|
Author |
Kind |
Percent per day |
Tons/ha/year |
|
Tham (1969) |
sp.a/ |
1.1 to 4.7 |
|
|
Doty (1971) |
elkhorn |
2.64 |
|
|
Doty (1973a,b) |
elkhorn |
1.5 to 5.5; x 2.3 |
36 |
|
Parker (1974) |
elkhorn |
1.5 to 5.5c/ (x 2.3d/) |
36 |
|
|
2 to 4 |
13 |
|
|
Dawes, et al. (1974b) |
spinosum |
1 to 2b/ |
|
|
Doty & Alvarez (1975) |
elkhorn |
1.73 |
42 |
|
|
tambalang |
>5.20 |
|
|
Glenn & Doty (1981) |
(all three) |
2.3 to 5.34b/; |
|
|
Lim et al. (1982) |
|
2 to 6e/ |
|
|
Liu & Zuang (1984) |
gelatinae |
0.41g/ |
|
|
|
0.44 to 0.63g/ |
|
|
|
Doty & Glenn (unpub.f/)
|
spinosum |
3.51 |
|
|
elkhorn |
3.51 |
|
|
|
tambalang |
5.05 |
|
a/Species unknown. The author and Dr. Tham Ah Kow were working cooperatively at the time and only with cottonii.
b/Determined from respirometry.
c/Range in first week.
d/Mean of same thalli the second week.
e/Determined simultaneously from weight change.
f/Each value was reduced from 55 consecutive weeks' weights.
g/Calculated from data provided.
Laite & Ricohermoso (1981) provide the statistics on the early growth of Eucheuma exports leading to the undocumented estimate of 35,000 tons in 1985-86. If 30,000 tons of the 35,000 probably is farmed, then between 1000 and 2000 hectares is probably in Eucheuma farms. Lim & Porse (1981) provide yet other tonnages. These two sources of information concerned the Philippines at a time when perhaps 90 percent of the world's Eucheuma was coming from there. At present (Adnan & Porse In Mss) some few thousands of tons per year come from Indonesia too. Other places produce at best a total of a few hundred tons per year.
Most of the Eucheuma crop goes to the U.S.A., Denmark, France, Australia, Japan and China. These markets are largely for pre-export-treated material and thus their tonnages do not reveal the total crop harvested. Their declared values do not necessarily indicate the dollars returned to either the first seller or the exporter.
The import values of Eucheuma shown in Table 5 are for the countries importing cottonii predominantly in rough dried form. The discrepancies between general export prices of $350 (cottonii) and $400 (farmed spinosum) to $600 (wild clean spinosum) for those in this table are considered to have been produced largely by the import tonnages being mixtures of kinds and qualities as well as proportions of semi-processed and raw seaweed.
Table 5. Declared Dollar Value of Eucheuma Exported from the Philippines, 1980 and 1984. The terms are metric tons and U.S. dollars per ton. Only countries receiving very predominantly non-modified cottonii in 1980 are included.
|
Country of destination |
1980 |
1984 |
||
|
Tonnage |
Value |
Tonnage |
Value |
|
|
Argentina |
154 |
357.14 |
256 |
437.50a/ |
|
Denmark |
4859 |
342.25 |
5408 |
429.36a/ |
|
France |
2360 |
296.19 |
3129 |
285.39 |
|
Spain |
1802 |
330.74 |
879 |
470.99a/ |
|
South Korea |
191 |
314.14 |
842 |
451.31a/ |
|
HongKong |
55 |
363.64 |
2223 |
25.1.46 |
|
Taiwan |
60 |
333.33 |
453 |
238.41 |
a/These high values must be presumed to reflect part of export was AMC or spinosum.
5.2.1 Physical determinants of productivity
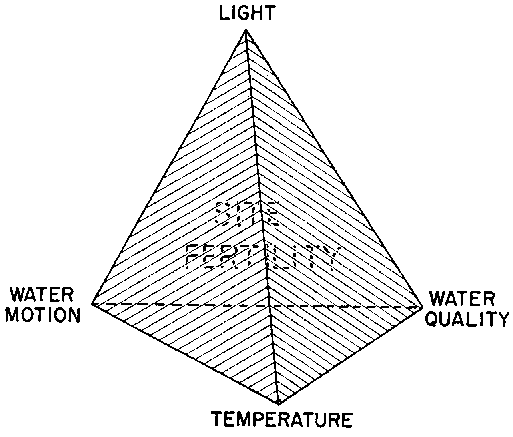
For wild crops the summation of the four complexes of physico-chemical factors (categorized as water motion, light, temperature, and water quality) was initially (Figure 7 and Doty 1971, 1979) considered to control "site fertility." Later, as farming came to dominate Eucheuma production, farm "productivity" in terms of mass/area/time, became the objective and water motion was recognized (Doty 1982a) as a prime factor in both successful site selection and productivity. Unless farm site selection has provided these four conditions suitably, and favorable tidal and bottom relationships as well, farm productivity is not satisfactory. (The similarly essential biological factors are considered in Section 5.2.3).
Figure 7. An hypothetical compensatorial relationship of site fertility to environment. Each of the four vertices of the tetrahedral figure represents respectively one of the four classes of physical environmental elements. The volume so established by the four plane surfaces of the tetrahedron represents both site fertility and productivity. (From Doty 1982b)
5.2.2 Seasonality and longer term variation in productivity
Field conditions have frustrated attempts to obtain reliable measurements of Eucheuma seasonality, except for the 55-week Hawaiian unpublished study of Doty & Glenn. Seasonal farm production data is undependable because of the lag between the time the seaweed is harvested and the time it reaches a level in the market chain where the quantity can be measured consistently. Added to this, one has no way of correcting the market chain for farmer, consolidator and extractor effort and these elements in themselves, have few seasonal constraints. The Hawaiian study reveals some of the environmental parameters (Table 3) within which Eucheuma thrives; it also indicates that spinosum and the two principle strains of cottonii grow (Table 6) at different rates and may have different seasonalities. Where these seaweeds are grown commercially nearer the equator, seasonality has appeared to be weaker except in monsoon areas.
Incident sunlight varies with latitude and longitude (List 1966) and varies (Figure 8) as much as 20 percent over periods of a few years. Sunlight intensity has a seasonal maximum, and commercial crop productivity seems to vary with it.
Sea temperatures are usually correlated with (but lag behind) sunlight intensity in an annual cycle. The global sea temperature (Figure 4) shows the area of the Indo-Pacific where the 23 to 33°C temperature range occurs. Geographically this temperature range indicates the latitudes favorable to Eucheuma commercial production.
Several year variations in water temperature and sea level changes surely influence Eucheuma production. These long-period changes are seemingly related to variations in the Southern Oscillation Index (see Ramage, 1986, for review) and this may in turn be related to such items as the pattern of variation in sunlight (Figure 8).
5.2.3 Human and other biological determinants of population density
The most important factors regulating the degree to which the highest economically sustained yields are obtainable are found among the biological factors controlling pests (Section 4.3 and 4.4), genetics (See Section 5.3) and the dedication with which wild and farmed crop agronomic practice are applied.
Table 6. Mean Growth Rates for Cottonii and Spinosum on the Hawaiian 55-week Experiment Plot. Cottonii was represented by "A" Eucheuma alvarezii and "B" by E. striatum with spinosum being represented by "C" E. denticulatum. "UP" and "DN" refer to the place of growth at, respectively, the upstream or downstream edge of the plot and growth is in terms of percent increase in weight per day. Only upstream values are given in the seasonal part of the table.
|
Season |
Growth rates as percent per day | ||
|
1976-7 |
A |
B |
C |
|
Spring |
5.25 |
3.95 |
3.51 |
|
Summer |
6.23 |
3.33 |
4.05 |
|
Fall |
4.41 |
2.98 |
3.52 |
|
Winter |
4.15 |
3.76 |
3.35 |
|
UP X |
5.05 |
3.51 |
3.61 |
|
DN X |
2.07 |
1.43 |
2.29 |
Though not done, it would apparently be relatively easy to produce new strains by traditional hybridization. There is no place equipped to do this now. There are two principle barriers to success: one is that the thalli are large and require both a large amount of desirable water and its movement; so in vitro work is limited. The other is the lack of recognizable, genetically-different strains and males in this genus. Eucheuma has not yet proven amenable to gene mechanics or protoplast fusion, and the costs and returns of this may not be favorable from an economic standpoint.
Sexual reproduction has little influence on crop production: all farmed crops are reproduced vegetatively.
Sexual reproduction and rearing crops from spores or very small vegetative propagulae of some kind do not seem practical in view of farm conditions. The size of Eucheuma thalli makes it economic in the Old World Tropics to break off and plant individual large pieces of the thalli, and this has become the routine for starting a farm or individual crop. Thus macro-vegetative reproduction is used. Inducing regeneration (Section 4.6) is not a part of the farming manipulations.
The farming routines advised (e.g., (Doty 1973a,b; Doty & Alvarez 1973), i.e., of harvesting and selling the small thalli after a crop growth period and replanting with vegetative fragments of the largest, would seem to be effective insofar as local genetic variability in the crop is concerned when farming is initiated from wild thalli. After a few crops, further use of this selection procedure produces neglible results insofar as improvement is concerned.
Early in the development of Eucheuma farming, a very significantly better variety was found (E. alvarezii var. tambalang Doty) by a selection program involving over 23 strains and species of wild cottonii. Such improvement has not been obtained in the case of spinosum despite many efforts. The most striking strains are strongly green thalli, principally, but other color and some morphological varieties are seen commonly. None of these appears to be superior from either the farm production or gel extraction points of view.
All farming involves environmental enhancement; in the case of Eucheuma it is largely provision of substratum. It may be concrete posts (Liu & Zhuang 1984) or nets (Doty 1973a,b, 1979; Doty & Alvarez 1973, 1975; Deveau & Castle 1976; Parker 1974) or more commonly lines nowadays (Figure 6) that hold the thalli in place in a desirable environment. It appears to be essential for such an environment to have water movement so strong near the sedimentary bottom that, at peak intensities, the seaweed would be washed away if they were not attached. The lower growth rates reported (e.g., Table 4) may be due to the plantings having been off the bottom or in calmer water.
Raking the bottom under the farm lines and fertilizing seem to enhance productivity, but perhaps will not produce economically significant increases in it. There is no shortage of suitable farm area, and the capital cost per hectare is very low; so unless there is a great labor saving, there is not much pressure for higher Eucheuma productivity.
Crop quality, not its productivity, is the greatest agronomic concern. The quality of the Eucheuma produced is affected by the agronomic practices of the farmers. The highest crop yields are produced by frequent harvesting, and this leads to harvesting what may be called immature material. Rational harvesting routines that replant young material and sell the oldest can be instituted; these will yield more carrageenan per ton of seaweed and higher quality-gel. Table 7 provides examples of value indices for a better standard cottonii and for a low grade cottonii, such as one can expect from poor harvesting practices. This table shows that if a crop indexed at 0.280 is worth $350 dollars per ton, a crop valued at but 119/280ths of that is worth only $145 per ton to the same processor if used for the same period.
Table 7. Linear Factoring as a Method of Evaluating Eucheuma for Extraction. The Value Index resulting is equal to the product of the individual evaluations in the columns to the left. While the individual percent values given may be characteristic they refer to no particular lot or source of the seaweed. The terms used are the Dry to Wet Weight Ratio (=Dry wt.), Clean Anhydrous Carrageenan Yield (=CAV) and Gel Strength (=Gal str.) and are standard terms to which a cost of processing (=Process) factor is added.
|
Seaweed quality |
Dry wt. |
CAY |
Gel str. |
Shipping |
Process |
Value index |
|
Standard |
0.70 |
0.40 |
1.00 |
1.00 |
1.00 |
0. 280 |
|
Low Grade |
0.60 |
0.30 |
0.60 |
1.00 |
1.10 |
0. 119 |
6.1 General harvesting cycle
6.2 Farm productivity
6.3 Eucheuma farming as an alternative means of employment
Wild crop production involves repeated periods of harvesting, regrowth and reharvesting with the addition of pre-sale treatment. Farming adds maintenance of the farm structures, planting, replanting and pest control. (See Section 4). Harvesting is breaking off part of each thallus by hand and removing it from the wild crop or farm area.
Immediately after harvesting, the most common treatment is sun drying. For this purpose the seaweed is usually scattered on rock or bamboo surfaces as this obviates sand, soil and trash being added. The more sophisticated drying is on slatted bamboo decks surrounding farm houses erected at the farm sites.
It takes about three days of hot sun to produce commercially dry Eucheuma, during which time the farmer should be alert to protecting his harvest from rain. Rain washes off salt and small organic solubles and causes bleaching. Such losses in weight and color increase the value of the crop, but this is at the farmer's expense, unless the greater effort and loss of weight bring a compensatory higher first sale price. Bone-dry Eucheuma rinsed in sea or fresh water for less than a minute will lose about 16% of its bone-dry weight.
To obtain greater value in the dried Eucheuma, there have been buying stations located among the farms. These consist of platforms surrounding the station's building(s) upon which, after the live crop is purchased, the cleaning and drying of the harvested lots is done uniformly by the consolidator. The buying stations, like the farm buildings, are constructed on posts driven into the reef flat surface so as to endure storm winds and waves. They are, thus, over water but elevated above the highest sea levels.
Initially the buying stations merely dried the seaweed and transported it the relatively long distance (for individual farmers) to the main consolidation areas. Recently there has arisen the commendable practice of purchasing live seaweed at the buying stations with the post-harvest treatment being done by the first purchaser. Under this regime variously sophisticated post-harvest treatments have been introduced at the buying stations.
The productivity of a farm depends on the inherent nature of the strain of seaweed and the agronomic practices being used, as well as the environmental conditions. (See Section 5).
Site fertility and the agronomy applied are so variable that there is little consistency in yield from place to place. Furthermore it is almost impossible to obtain such information without living at the farm being studied. Two of the best reports of field observations are Lim et al. (1982) and Adnan & Porse (In Mss) though incomplete in many respects. Yields as high as 74 (spinosum) to 104 (cottonii) dry tons per hectare per year have been verbally reported. However, a usual range (Table 4) of 15 to 30 tons per hectare per year is recognized.
Returns to the farmer vary from place to place. Lim & Porse (1981) provide the only published recent first sale values. Outside the Philippines first sale values are believed to range from perhaps 10% to 80% of the export value. Doty (1986) provides a method of relating the farmer's productivity to his cash returns from first sale, assuming 75% of the export price is returned as is common in the Philippines. The grey literature contains more information.
Farm growth rates of Eucheuma (Table 4) routinely range from 2 percent per day toward ten percent per day, depending upon strain and environment. Better quality Eucheuma is produced when the growth rates are such that the crop doubles in size in each lunar cycle of 28 days. This is a growth rate equal to 2.51 percent per day. More practically speaking, a growth rate of 3.5 percent per day is thought to offset the generally anticipated negative factors and provide a doubling of the crop and so yield a harvest of one half of it every month. In some places the thalli will double in weight in less than 14 days.
It is obvious that in starting a farm, the farmer should plant the largest thalli feasible. However, the larger the thalli become, the more susceptible they are to physical damage and loss. Thus the size at which harvesting would best take place, the mature size, is different in different places, though the strain and agronomic treatments may otherwise be the same.
The mature size is usually between 512 and 1028 grams. Especially seasonally, the mature size may be (Figure 6) much larger. The size of large mature Eucheuma thalli make it practical to handle, tie in place, tend and harvest individuals. The productivity can be calculated from the number of thalli involved, the mature size of the thalli, and time between harvests. Many farms have about 100,000 thalli per hectare.
A large, successful farm could require several people for its proper operation, and a small farm might be successful even though it does not require all the time of even one person. The farming of Eucheuma is not on an annual cycle; it is rather continuous in the same sense as poultry farming. Eucheuma farming is excellent as an alternative means of employment.
When the farm is small, the artisanal fisherman who has become a Eucheuma farmer can often continue his traditional ways of obtaining a livelihood, and it is often his wife who farms the seaweed. Thus, Eucheuma farming is a ready way of improving family income.
The farms are on reef flats or other horizontal bottoms near extreme low-water level. Since working at high tide is often much more exhausting than working at low tide, seaweed farming tends to become a part-time occupation. Furthermore, since the tides are not governed by calendar or solar time, but vary in lunar month (siderial time) cycles, the formal employment conventional to the developed world is hardly appropriate. It is sometimes difficult for those not working in the sea to cope with this idea, that favorable conditions for work are not in accord with the civil or religious calender days, weeks, weekends, holidays or seasons.
Incentive and free enterprise are major factors in farm productivity. Early experience (Doty 1973a,b) in developing the farming of Eucheuma led to the statement "having the farms small, and owned and operated by the workers who benefit in proportion to their investment of material and effort seems to be the key to successful production of this seaweed." From a different series of experiences, Lim & Porse (1981) have pointed out that Eucheuma from an absent-owner estate-managed farm costs $0.34 per kg to produce. Comparably from a resident-owner farm hiring workers, they found the cost was $0.37 per kilogram, but only $0.20 per kilogram from a family farm where there are no paid employees. Doty & Alvarez (1981) provided the further figures of 2 to 3 kg/man-hour on a non-seasonal family farm and 1 to 2 kg/man hour on an estate farm.
7.1 Wild resource management
7.2 General statements regarding capitalization
7.3 Sources of credit and insurance of the crop
Wild crop harvesters normally forage on the reef flats for much of their livelihood and so are already equipped to forage for Eucheuma. This equipage may be as little as just the essential knowledge of what seaweed is to be harvested and where it is found. The first level of sophistication might be a container in which to accumulate the Eucheuma as it is picked. Their locally made family boat is often used. While harvesters usually carry a large knife; this and even hats and shoes are not requisite to wild crop harvesting.
When wild crop harvesters or fishermen shift to farming Eucheuma, they may start with a single piece of monofilament line obtained at no cost and, as they sell their crop, use some of the proceeds to obtain more line. Creating circumstances which allow them to do this is one of the better ways to get this farming undertaken. Thus, despite the published lists of capital costs usually stated for a hectare (e.g./Doty 1973a/b), the costs may increase so gradually from negligible to such as the example tabulated (Table 8) as to be no barrier to undertaking farming. Most farms are probably near one tenth of a hectare or far less than a hectare in extent.
Table 8. Estimated Costs for Capital Investment in a Eucheuma Farm. Parameters are one acre of average quality bottom and Eastern Sabah (Malaysia) minimal costs in 1982 Malaysian dollars. 2.47 acres = 1 hectare.
|
I. Material expenses: | ||||
|
Item & Description |
Number needed |
Length needed |
Weight needed |
Cost in M$ in 1980 |
|
A. For the farm: |
|
|
|
|
|
1) Mangrove posts, 2 inches in diameter by 3' @ M$1.68 each |
565 |
3' or 1 m |
|
942.38 |
|
2) Mangrove posts, 12 feet long @ M$0.30 each |
4 |
12' or 3 m |
|
1.20 |
|
3) Nylon mono filament line 120 Ib test strength (this is not the #120 on most packages) |
|
14,700 |
5.8 lbs or 2.6 kg |
100.00 |
|
4) Tie-tie material cut into pieces 10" long (25.4cm) each split into 3 pcs. |
4,839 |
1,344' or 409 m |
|
20.00 |
|
B. Seedstock |
|
|
|
|
|
1) Seedstock, M$0.20 per kilo, (100g per plant initial, weight) at 1.5'x1.5' |
17,689 |
|
1,769 kg |
354.00 |
|
C. Total estimated cost of materials |
1418.58 | |||
|
II. Labor expenses: | ||||
|
A. Mangrove posts | ||||
|
1) Delivered to site |
100.00 | |||
|
2) Total cost of labor (for cutting and sharpening 565 stakes at $0.10 each) |
60.00 | |||
|
B. Staking (installation costs) | ||||
|
1) 565 stakes per acre, 10 minutes to set a stake or ca 100 man-hours per acre = 25 4-hour low tide periods with travel time each day @ M$8.00/such 4-hr day. | ||||
|
Labor cost |
200.00 | |||
|
C. Tying of seedstock | ||||
|
1) 125 to 240 thalli per hour - 1000 to 1920 thalli per man-day @ 50/man-days per acre @ $10.00/day at dense spacing | ||||
|
Cost of 5,000 thalli |
1,000.00 | |||
|
D. Setting and planting of monofilament lines (4 hours low tide working time per day.) | ||||
|
1) 32 man-hours per acre to make and install mono-lines and obtain thalli, M$8/day for 8 days. |
64.00 | |||
|
2) 16 man-days to set up monolines and plant 4,839 (a minimal number of) thalli on one acre |
128.00 | |||
|
E. Estimated total cost of labor w/o supervisory time. |
1552.00 | |||
|
III.. Equipment needed | ||||
|
1 crow bar |
18.00 | |||
|
2 heavy knives (e.g. bolo) |
12.00 | |||
|
2 stainless steel knives |
7.60 | |||
|
4 rattan baskets |
28.00 | |||
|
1 plastic container for water |
10.00 | |||
|
1 plastic container for gasoline |
10.00 | |||
|
Miscellaneous |
14.40 | |||
|
Total |
100.00 | |||
|
IV. Total estimated cost of materia1s and installation of a one acre monoline Eucheuma farm |
3070.00a/ | |||
a/In 1985 this was equivalent to about U.S.$1330.
When government or tradition intervenes, rather than cost of materials, etc., a stumbling block to farming from the would-be-farmer's point of view could be obtaining the right to farm.
An investor would find the weakest place in Eucheuma farming to be in the availability of competent personnel for selecting a site, installing a farm and managing it. The cost of such a manager could be equivalent to the local costs for a non-college-graduated school teacher. In addition the investor would have to obtain clearance to use the site and obtain the initial planting material. Table 8 displays the materials and their costs for one acre farms constructed in easternmost Sabah, Malaysia, for the State Fisheries Department. These Sabah costs are surely more nearly maximal than minimal if for no other reason than that they included (Table 9) two surrounding sets of nets and devices to control herbivorous fish (largely Siganus) and Chelonia midas, the green turtle.
The amount of time required is increased by the travel time to and from the farm construction site and by the tides. At the Sabah sites to which Tables 8 & 9 refer the time on the job was about four hours per man day. This would be different elsewhere.
Essentially there are no sources of credit or insurance for seaweed crops. In practice the most to be had is provided by the strong ties among the pre-sale seaweed producers and credit with the local merchants.
8.1 Management of seaweed resources
8.2 Regulation of seaweed
As in many such agricultural industries, there are three levels of management concern plus the government. These may be separated conveniently into a) pre-first sale level of wild crop harvesters and farmers; b) post-first sale entrepreneurs and exporters, and c) the ultimate processors. Hollenbeck (1983) and Vellin & Hollenbeck (1982) provide much more.
8.1.1 Commercial relationships
Management of wild crop harvesting has always been largely in local, usually informal or traditional, hands. A typical operation is for a post-first sale entrepreneur to make arrangements to buy Eucheuma from a shoreside village merchant. This merchant is often the de facto local government insofar as the harvesters are concerned. In Indonesia he is (Lee Hollenbeck, personal communication) often the traditionally recognized village leader who gets the artisanal fishermen to collect seaweed, often a given or traditional amount. The village leader accumulates the seaweed and sells or barters it to the entrepreneur or trader, who will probably be but a representative of a traditional exporter. In a money-economy situation the village leader, the trader and the exporter may be considered second level interests. The third level interests, the processors, establish commercial relationships which are often agreements to buy a certain tonnage within certain time/quality limits "free and on board" a ship bound for the port nearest the processor's plant.
Table 9. Fish and Turtle Exclosure Fences and Trap Costs. The list of materials is for a square one acre farm ca 210 feet on a side in an area where the tides may rise as high as two meters. Values are in 1982 Malayaian dollars.
|
Item & Description |
Number needed |
Length needed |
Weight needed |
Amount in M$ |
|
A. For fencing: |
|
|
|
|
|
1) Mangrove posts |
148 |
12 ft. long & 2 " diam. |
|
44.40 |
|
2) Stranded fish net Twine: PE. 380/6 Mesh: 2 1/4" Width: 300 cm Length: 150 ft./roll |
3 rolls |
880.6 ft. |
|
750.00 |
|
3) Border-line for stranded fish net, 150 Ib test. |
|
|
2 kilos |
24.00 |
|
4) Wire for securing fish net to posts, 16 inches long |
444 pcs. |
592 ft. |
|
20.00 |
|
5) For turtle net, 2.5 x 5 meters in standard net units |
60 pcs. |
946 ft. or 288.5m. |
5 kilos |
228.00 |
|
6) Mangrove posts for turtle net fencing |
60 pcs. |
12 ft. by 2" in diam. |
|
20.00 |
|
7) Wire for securing turtle nets to posts, wire pieces 16" long. |
174 pcs. |
232 ft. |
|
8.00 |
|
B. Two fish traps: |
|
|
|
|
|
1) Mangrove stakes, 3 meters long by 2 1/2 " in diam. |
8 pcs. |
10 ft. |
|
2.40 |
|
2) Mangrove stakes, 2.5 meters by 2 1/2" in diam. |
8 pcs. |
9 ft. |
|
2.40 |
|
3) Many rove stakes, 1 meter by 2 1/2" in diam. |
8 pcs. |
4 ft. |
|
1.20 |
|
4) Stranded fish net; Twine: PE. 380/6 Mesh: 1 1/4" Depth: 300 cm |
|
32 ft. |
|
7.00 |
|
C. Total fencing & trap material costs |
1,107.40a/ | |||
a/In 1985 this was equivalent to about U.S.$480.
8.1.2 Government roles
Two governmental groups are often the official bodies of concern: economic development authorities and the fisheries departments (often) of the agriculture department. They usually arise after-the-fact due to their lack of entrepreneurial and phycological capability as well as incentive, the need to heed the wishes of the politico/economic interests and their traditional fin/shellfish concerns.
The private sector has been the initiator (Doty 1973b; Adnan & Porse In Mss) and has been initially in control of the first two levels in almost all successful production areas: e.g., to the extent that there is production in Indonesia, Philippines, Taiwan and Tanzania. In Indonesia the Agency for the Assessment and Application of Technology and in Kiribati and Malaysia the fisheries departments have been very positively active in seeking to establish farming.
In the Philippines the national fisheries department research section played an unusually successful role by coordinating and contributing its facilities to the expatriate academic and processing interests of industry at the initial development level of both the wild crop harvesting and later the R & D leading to pre-eminence of the Philippines in both cottonii and spinosum farming.
There are no effective formal mechanisms for consultation between the levels of concern in the Eucheuma industry.
There are essentially no regulations effectively governing the harvesting/culture of Eucheuma. What arrangements exist for leasing or private ownership of nearshore or intertidal areas for this purpose have been ineffective or have been applied at the initiative of local fisheries officers and the private sector by what might be called field expediency. In southernmost Philippines, village councils have shouldered the responsibility for individual farmer concessions under the umbrella protection of master concessions from the government obtained by the processors. In some other places government participation in efforts to "facilitate" farmer leasing have been as much a barrier to success as an aid.
The concession plans in Indonesia and Malaysia; at least, were established so as to leave passageways for local boat traffic. On reef flats there is no conflict with fisheries useage or with ocean navigation. The farms result in enhanced fish crops, and fish traps and Eucheuma farms do not compete for the same space.
To date no specific legislation is known that affects Eucheuma farming. In Hawaii it has been effectively prevented in the past by the classification of all sub-tidal land as conservation, a designation that prevents its economic development. This situation in Hawaii is changing favorably through new legislation. In the Philippines legislative and executive bodies have considered legislation and various regulations, but these have not been effected. Regulations have been instituted to obtain advantages for individuals, but they are ineffective and largely ignored. Notably in one African country a monopoly has resulted in no development of farming and apparently no increase in export during the past 20 years. In two other countries government has entrusted development to innocent hands that despite goodwill have been unable to move to the economic export level. In some others nothing has happened. Only in those countries where there was no contrary legislation and the ultimate processing companies from abroad have sent in expatriate management has introduction led to farming and regular export.
9.1 Chemical and nutritional content
9.2 Human food
9.4 Animal Fodder
9.5 Manure
9.6 Industrial products and processes
Industry's only concern for Eucheuma is its carrageenans. Carrageenan is a cell wall component which represents 40 to 75 percent of the seaweed's salt-free dry weight. In addition to carrageenan the cell walls of a Eucheuma contain a small amount of cellulose and minor amounts of other unspecified materials.
Eucheuma reaches the ultimate consumer as whole or particulated, dried material in one of several forms. Commonly the carrageenan may be extracted carrageenan (with or without chemical modification) or in the non-extracted modified form.
Basically carrageenans are d-galactopyranoses alternately paired by alpha 1-3 and beta 1-4 linkages. Their molecular weights may be as high as 400,000. They are typically sulfated though sulfate is lacking (Greer & Yaphe 1984) in beta carrageenan. Seven types are recognized and referred to by the greek letters, beta, gamma, kappa, lambda, iota, mu and nu. Infra-red absorption peaks are present for total sulfate and, respectively, that at any of the 2, 4 or 6 positions. In addition, the different carrageenans are distinguished by their solubilities in water at different temperatures (Greer & Yaphe 1984) and reactions to different salt solutions as well as the physico-chemical properties of their gels. At present there is considerable question as to their structure, e.g., whether they are purely one or another of the seven types listed above or whether there are blocks of one type in combination with blocks of other types. Relatively low sulfate content and 3-6 anhydro linkages are found to be related to high gel strengths. Milk reactivity and gel strength may be related to potassium and sulfate, respectively, at the 4-carbon positions. Various other radicals may be attached especially at 6-carbon positions.
The nutritional value of Eucheuma and its contained carrageenan are unknown. Its effect on humans is believed to be nil and, in foods containing it, the carrageenans are minor components by weight, though often indispensable for contributing a specific quality to the product.
Culinarily speaking, Eucheuma, the fresh or non-dried seaweed itself, adds delightful variety, novelty, and bulk to peoples' diets in various countries where it grows, e.g., as a fresh salad component or as pickles.
The extracted carrageenans provide stabilization and quality to the products in which they are combined. Today, many instant foods that come in powdered form for mixing with cold milk to make them comestible owe their speed and permanence of suspension to the carrageenan modification they contain. A desirable mouth feel is sometimes obtained by adding a carrageenan. Some major toothpastes contain it for its way of holding water in the product and, thus, extending shelf life. The successful suspension of chocolate in milk is perhaps best done with carrageenan. Most if not all evaporated milk nowadays contains carrageenan as the component that makes possible its pleasant flavor and thick, creamy quality.
Aside from "extracted carrageenan", there is the alkali-treated Eucheuma (sometimes spoken of as AMF or AMC, i.e., alkali modified flour or alkali modified carrageenan or semi-processed carrageenan). Making alkali-treated Eucheuma seems to consume most of the cottonii Eucheuma produced, for it is far less expensive than extracted carrageenan. It can usually substitute for extracted carrageenan where a little cloudiness from the small percentage of cellulose present does not interfere.
Both the alkali-modified Eucheuma products and the extracted carrageenans must compete with other hydrocolloids on the basis of amount required to do the job and cost per pound. An outstanding exception is their special effect in some milk products wherein carrageenan is often essential or nearly so. Carrageenans form synergistically desirable substances with other hydrocolloids and are chemically modifiable in many ways, giving them a wide variety of useful special qualities. Guiseley et al. (1980), Whistler & BeMiller (1973) and Glicksman (1969, 1982) provide four of the most useful treatises on applications of the different carrageenans and their modifications.
The carrageenans and Eucheuma are not normally used in animal fodder. However, the largest volume of alkali-treated Eucheuma is in pet foods where it is used as a powdered, granulated or hydrated gel component in the various formulations.
Eucheuma has essentially no use as a manure. Though its addition as an organic component in conditioning sandy soils is in some cases attractive, the cost is high. Some effort has been put forth to obtain use of the filter aid waste from the carrageenan extraction plants as a soil conditioner. Such waste filter aid is the major nonsoluble product from Eucheuma processing
The carrageenans extracted from Eucheuma are modified by proprietary methods and even the relatively simple alkali-treatment process is hardly publicized. The assayed plant yields of extracted carrageenan may vary (Adnan & Porse In Mss) from 23 to 36 percent. A simple diagrammatic flow sheet is provided (Figure 9) though it is hardly applicable to any one actual alkali-modification plant.
The carrageenan extraction process itself is well known to be a hot water, high pH, process. The principal costs are in heat and separating the carrageenan from the extraction water. Three approaches are in use for water removal after centrifugation, filtering and sometimes vacuum removal of some water: a) spray drying on steam-heated drums as in the making of powdered milk; b) cold-water extraction of the seaweed (when Chondrus is the source) such that the lambda carrageenan is separated from the kappa, with the kappa being extracted and precipitated separately later; and c) precipitation with alcohol. Each process has its merits, and some large companies use more than one method to obtain different qualities in their final products. Moss & Doty (In Press) provide a flow sheet (Figure 10) for the latter plant type.
Aside from the raw seaweed material, the principal costs of carrageenan (Moss & Doty, In Press) are in inventory, warehousing, water, power, alcohol recovery (only in alcohol precipitation plants), filter aid (diatomite or expanded mica as a rule) and chemicals (largely calcium and potassium hydroxides).
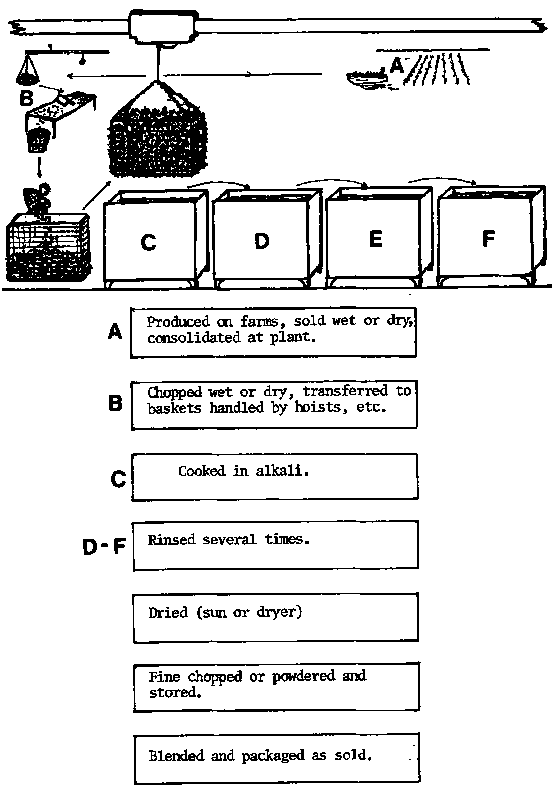
Figure 9. The alkali-treatment process for cottonii. Depending upon end product specifications, live or dried cottonii with or without preliminary treatment is often treated first in a hot alkali solution containing about 8.5 percent potassium hydroxide and then rinsed in successive tanks of water. The seaweed may be handled in baskets by an overhead traveling crane. Following a final draining, the treated material may be dried whole, or chopped and dried. The lower part of the figure shows a succession of processing steps that may follow depending on control laboratory analysis and desired nature of the final product
Agardh, J.G., 1847 Nya alger fran Mexico. Ofvers.K.vet.Akao.Forhandl., 4:5-17
Anderson, G.W., 1953 A note on the seaweed resources of Zanzibar protectorate. Proc.Int.Seaweed Symp., 1:102-3
Burman, N.L., 1768 Flora Indica: cui accedit series zoophytorum indicorum, prodromus florae capensis. Leiden Cornelis Haak, 242 p.
Collins, F.S., and A.B. Hervey, 1917 The algae of Bermuda. Proc.Am. Acad.Arts Sci., 53:195 p.
Dawes, C.J., 1981Marine botany. New York, John Wiley and Sons, 628 p.
Dawes, C.J., S.A. Earle, and P.C. Croley, 1967 The offshore benthic flora of the southwest coast of Florida. Bull.Mar. Sci., 12:211-31
Dawes, C.J., A.C. Mathieson, and D.P. Cheney, 1974 Ecological studies of floridian Eucheuma (Rhodophyta, Gigartinales). 1. Seasonal growth and reproduction. Bull.Mar.Sci., 24: 235-73
Dawes, C.J., et al., 1974 Ecological studies of floridian Eucheuma (Rhodophyta, Gigartinales). 3. Seasonal variation of carrageenan, total carbohydrate, protein, and lipid. Bull.Mar.Sci., 24:286-99
Dixon, P.S., 1962 Taxonomic and nomenclatural notes on the Florideae. 3. Bot.Not., (115):245-60
Deveau, L.E., and J.R. Castle, 1979 The industrial development of farmed marine algae: the case-history of Eucheuma in the Philippines and USA. In Advances in aquaculture, edited by T.V.R. Pillay and W.A. Dill. Farnham, Surrey, Fishing News Books Ltd., for FAO, pp. 410-5
Doty, M.S., 1971 Physical factors in the production of tropical benthic marine algae. In Fertility of the sea, edited by J.D. Costlow. New York, Gordon and Breach Science Publishers, Vol.1:99-121
Doty, M.S., 1972 Algal standing crop data from Philippine Eucheuma communities. Hawaii Bot.Sci.Pap.Univ.Hawaii (27): 648 p.
Doty, M.S, 1973 Eucheuma farming for Carrageenan. Sea Grant Advis. Rep.Univ.Hawaii, (UNIHI-SEAGRANT-AR-73-02):21 p.
Doty, M.S, 1973 Farming the red seaweed, Eucheuma. for carrageenans. Micronesica, 9:59-73
Doty, M.S, 1978 Eucheuma - current marine agronomy. In The marine plant biomass of the Pacific northwest coast, edited by R. Krauss. Corvallis, Oregon State University Press, pp. 204-14
Doty, M.S., 1979 Status of marine agronomy, with special reference to the tropics. Proc. Int. Seaweed Symp., 9:35-58
Doty, M.S., 1982 The diversified farming of coral reefs. University of Hawaii Harold L. Lyon Arboretum Lecture Number Eleven. Honolulu, Hawaii, University of Hawaii Press, 29 p.
Doty, M.S., 1982a Worldwide status of marine agronomy. In Proceedings of Republic of China - United States Cooperative Science Seminar on cultivation and utilization of economic algae, edited by R.T. Tsuda and Y.M. Chiang. Mongilao, Guam, University of Guam Marine Laboratory, pp. 17-22
Doty, M.S., 1985 Eucheuma alvarezii, sp- nov. (Gigartinales, Rhodophyta) from Malaysia. In Taxonomy of economic seaweeds with reference to some Pacific and Caribbean species, edited by J.A. Abbott and J. Norris. La Jolla, California, California Sea Grant College Program (T-.CSGCP-011):37-45
Doty, M.S., Estimating Gracilaria and Eucheuma productivity on line farms. Monographias Biologicas Marinas, 37 p. (in press)
Doty, M.S., and V.B. Alvarez, 1973 Seaweed farms: a new approach for U.S. industry. Annu.Conf.Proc.Mar.Technol.Soc., 9: 701-8
Doty, M.S., and V.B. Alvarez 1975, Status, problems, advances and economics of Eucheuma farms. Mar. Technol. Soc.J., 9(4):30-5
Doty, M.S., and V.B. Alvarez, 1981 Eucheuma farm productivity. Proc.Int.Seaweed Symp., 8:688-91
Doty, M.S., and J.N. Norris, 1985 Eucheuma species (Solieriaceae, Rhodophyta) that are major sources of carrageenan. In Taxonomy of economic seaweeds with reference to some Pacific and Caribbean species, edited by I.A. Abbot and J. Norris. La Jolla, California, California Sea Grant College Program (T-CSGCP-011):47-61
Esoer, E.J.C., 1797-1808 Icones fucorum. 357 p.
Gabrielson, P. W., 1983 Vegetative and reproductive morphology of Eucheuma isiforme (Solieriaceae, Gigartinales, Rodophyta). J. Phycol., 19 (2): 4 5 - 5 2
Gabrielson, P.W., and G.T. Kraft, 1984 The marine algae of Lord Howe Island (N.S.W.): the family Solieriaceae (Gigartinales, Rhodophyta). Brunonia, 1984:217-25
Glenn, E.P., and M.S. Doty, 1981 Photosynthesis and respiration of the tropical red seaweeds. Eucheuma striatum (Tambalang and Elkhorn varieties) and E. denticulatum. Aquat.Bot., 10:353-64.
Glicksman, M., 1969 Gum technology in the food industry. New York, Academic Press
Glicksman, M, 1982 The hydrocolloids industry in the '80s - problems and opportunities. Proc.Food Nutr.Sci., 6:299-321
Gmelin, S.G., 1768 Historia fucorum. St. Petersburg, Academia Scientiarum, 239 p.
Greer, C.W., and W. Yaphe, 1984 Characterization of hybrid (beta-kappa-gamma) carrageenan from Eucheuma gelatinae J. Agardh (Rhodophyta, Solieriaceae) using carrageenases, infrared and 13C-nuclear magnetic resonance spectroscopy. Bot.Mar., 27:473-8
Guisley, K.B., N.F. Stanley, and P.A. Whitehouse, 1980 Carrageenan. In Handbook of water soluble gums and resins. Edited by R-L. Davidson. New York, McGraw-Hill
Hollenbeck, L.S., 1983 Economic assessment of the Philippine Eucheuma seaweed industry with implications for future policy. Masters Thesis in University of Hawaii Library, Honolulu, Hawaii, 116 p.
Kraft, G.T., 1969 The red algal genus Eucheuma in the Philippines. Master's Thesis deposited in the University of Hawaii Library, 358 p.
Kraft, G.T, 1972 Preliminary studies of Philippine Eucheuma species (Rhodophyta). Part 1. Taxonomy and ecology of Eucheuma arnoldii. Pac.Sci., 26(3):318-35
Kylin, H., 1956 Die Gattungen der Rhodophyceen. Lund, K. Gleerups, 673 p.
Laite, P., and M. Riconermoso, 1981 Revolutionary impact of Eucheuma cultivation in the South China Sea on the carrageenan industry. Proc.Int.Seaweed Symp., 10:596-600
Lim, J.R., and H. Porse, 1981 Breakthrough in the commercial culture of Eucheuma spinosum in northern Bonol, Philippines, Proc.Int.Seaweed Symp., 10:601-6
Lim, J.R., et al., 1982 Farming the ocean (The Genu story). Publ. Hist.Conserv.Soc., Manila, 36:143 p.
Linnaeus, C., 1771 Mantissa plantarum altera. Impensis direct. Stockholm, Laurent. Salvii
List, J., 1966 Smithsonian meteorological taoles. Smithson.Misc. Collect, (1 1 4): 5 2 7 p.
Liu, S.-J., and P. Znuang, 1984 The commercial cultivation of Eucheuma in China, Hydrobiologia, 116/117:243-5
Mathieson, A.C., and C.J. Dawes. 1974 Ecological studies of floridian Eucheuma (Rhodophyta, Gigartinales). 2. Bull.Mar. Sci., 24:247-85: 274 - 85
Moss, J.R., and M.S. Doty, Hawaii as a site for production and processing of seaweed materials. Honolulu, Aquaculture Development Program of the Hawaii State Department of Land and Natural Resources, 150 p. (in press)
Mshigeni, K.E., 1982 Seaweed resources in Tanzania: a survey of potential sources for industrial phycocolloids and for other uses. In Marine algae in pharmaceutical science, 2, Berlin, edited by H.A. Hoppe and T. Levring. Walter de Gruyter and Co., pp. 174
Mshigeni, K.E, 1984 The red algal genus Eucheuma (Gigartinales Solieraceae) in East Africa: and an underexploited resource. Hydrobiologia, 116/117:347-50
Neish, A.C., and P.F. Shacklock, 1971 On the propagation of strain T-4 of Irish moss. Tech.Rep.Ser.Atl.Reg.Lab.Natl.Res. Counc.Can., (14):25 p.
Parker, M.S., 1974 The culture of the red algal genus Eucheuma in the Philippines. Aquaculture, 3:425-39
Ramage, C.S., EL Niño. Sci.Am., 1986 254:77-83
Russell, D.J., 1983 Ecology of the imported red seaweed Eucheuma striatum Schmitz on Coconut Island, Oahu, Hawaii. Pac.Sci., 37(2):87-107
Santos, G.A., and M.S. Doty, 1978 Carrageenans from tetrasporic and cystocarpic Eucheuma species. Aquat.Bot., 4:143-9
Tham, A.K., 1969 A review of research work carried out by the Fisheries Biology Unit of the University of Singapore during 1965 and 1966. In Proceedings of the Second Symposium on scientific and technological research in Malaysia and Singapore, 1967, pp. 136-40
Thomassin, B.A., 1976 Feeding behaviour of the felt-, sponge-, and coral-feeder sea stars, mainly Cylcita schmidelina. Helgol.Meeresunters., 28:51-75
Turner, D., 1808 Fuci, sive plantarum Fucorum generi a botanicis adscriptarium descriptiones et historia... Vol. 1. 64 p.
U.S. Navy Hydrographic Office, 1944 world atlas of sea surface temperature. Washington, D.C., U.S. Navy Hydrographic Office, Publ.U.S. Navy Hydrogr.Off., Wash., (225) 2nd ed.
Uyenco, F.R., L.S. Saniel and G.S. Jacino, 1981 The "ice-ice" problem in seaweed farming. Proc.Int.Seaweed Symp., 10:625-30
Vellin, J.L.C., and L., Hollenbeck, 1982 Report on Eucheuma seaweed farming in Nusa Tenggara Timur, Indonesia. Annex I: List of seaweed traders worldwide. Annex II: Manual on farming of Eucheuma spinosum. Jakarta, Government of the Republic of Indonesia
Weber-van Bosse, A., 1913 Marine algae. Rhodophyceae. Trans Linn. Soc.(Ser.2.Bot.), 8:105-42
Weber-van Bosse, A, 1928 Liste des algues du Siboga. 4. Pt. 3 Gigartinales Rhodymeniales. Siboga Exped., (59d):393-533
Whistler, R.L., and J.N. Bemiller, 1973 Industrial gums polysaccharides and their derivatives. New York. Academic Press, 337 p. 2nd ed.
![]()
![]()
![]()
{kind=link}
{kind=link}
{kind=link}
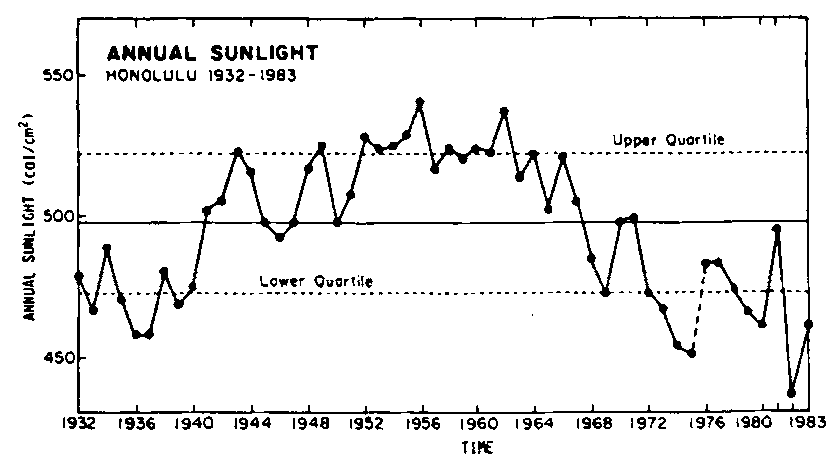{kind=link}
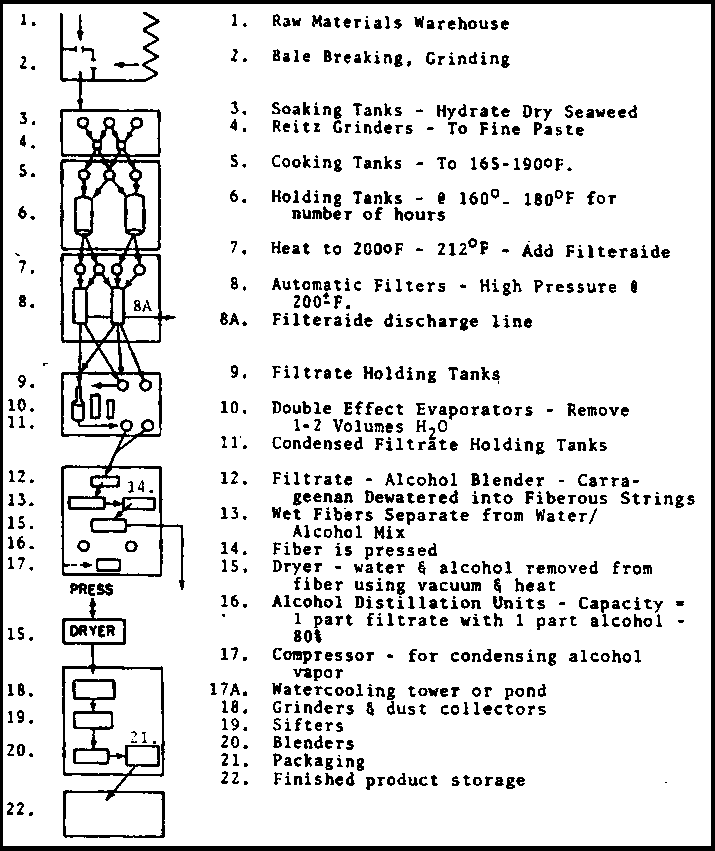{kind=link}