![]()
![]()
1. IDENTITY
2. DISTRIBUTION, ECOLOGY AND METABOLISM
3. LIFE HISTORY
4. POPULATION STRUCTURE AND MORTALITY
5. PRODUCTIVITY OF THE RESOURCE
6. METHOD OF HARVESTING AND HARVESTING CYCLE
7. EQUIPMENT USED FOR HARVESTING
8. PROTECTION AND MANAGEMENT OF THE RESOURCE
9. UTILIZATION
10. REFERENCES
By
Wheeler J. North
W.M. Keck Engineering Laboratories
California Institute of Technology
Pasadena; California 91125 USA
|
ABSTRACT The major North American source of alginate is Macrocystis from California. The area, standing crop and yield in California have varied greatly, e.g., from a maximum of 358,425 wet tons from 13,171 ha in 1918 to a minimum of 236 T from 13,171 ha in 1931 to recent tonnages of about 100,000 to, in 1984, 47,320 T from 4,765 hectares. To some extent variations in the southern sea level index and appearance of the El Nino current off the west coast of South America are related: a complex of grazer and other phenomena have been implicated.. A large amount of experimental physiological and population work is reviewed. |
The genus Macrocystis (Macro = large, Kystis = bladder) is presently recognized as containing three species1 M. pyrifera, (refers to the pear-shaped or pyriform bladders), M. angustifolia (narrow leaves), and M. integrifolia (complete leaves). There were formerly many more species recognized, based on frond morphology. Further study revealed that frond morphology could be profoundly affected by environmental factors. Later workers consequently chose holdfast morphology as a basis for species separation (Womersley, 1954). The holdfast of M. pyrifera is terete with an upright axis usually conical for plants a year or more old. Holdfasts of M. integrifolia are more or less prostrate with fronds developing from a creeping rhizome with a flattened axis. Holdfasts of M. angustifolia are intermediate between M. pyrifera and M. integrifolia. Neushul and coworkers have obtained viable crosses between all three species.
1Note added in proof, a fourth species, M. laevis, has just been proposed by C.H. Hay (Phycologia, Vol. 25, 1986, pp. 241-252.
Macrocystis belongs to the Order Laminariales, which includes those Phaeophyta growing by means of an intercalary meristem as well as a superficial meristoderm and has oogamous reproduction by microscopic dioecious gametophytes. Macrocystis above the holdfast is differentiated into a blade and stipe, with the blade being split forming several intercalary meristems during early sporophytic development. This places it in the Family Lessoniaceae.
Microscopic stages in the Macrocystis life cycle (i.e. gametophytes and embryonic sporophytes) resemble those of other members of the Lessoniaceae (see below) and they cannot be distinguished from one another. The morphologies of the macroscopic adult sporophytes differ widely in this family and constitute the basis for separation into genera and species. The typical Macrocystis adult sporophyte (Figure 1A) consists of numerous blade-bearing fronds that arise from a basal branching system of stipes. Lower portions of the basal stipe system also produce root-like haptera that grow downwards, wrapping around and forming attachment to irregularities in solid substrata. Uniquely the haptera of M. angustifolia can also attach to sedimentary bottoms (See Section 2. 3). The mass of haptera constitutes the holdfast.
New young fronds arise (Figure 1A) from longitudinal splits in meristematic blades located on the basal branches just above the holdfast (the basal meristems, Figure 1A). The outer blade of the two produced by the splitting becomes an apical frond initial while the inner portion remains as a basal meristem that will give rise to yet further new frond and basal meristem initials.
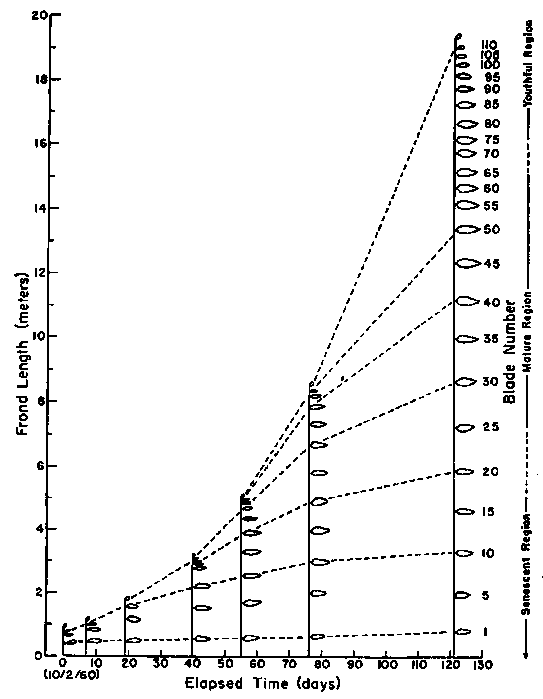
Splits appear near the base of the outer (apical) frond initial of a young frond, and developing longitudinally toward the blade edge, subdivide the original lamina into lanceolate strips each of which becomes one blade on the young frond. Elongation occurs in that part of the blade base where the splits arise, yielding the stipe of the young frond and causing separation of the newly forming blades. Further stipe growth and splitting produces an apical scimitar-shaped blade distinct from the remaining blades. This is the apical meristem which generates additional blades and stipes as the frond continues to develop. The lowermost 2 or 3 blades lack pneumatocysts (bladders) but thereafter all upper blades carry these pyriform flotation devices joining the laminae to the stipe. Continued elongation in the stipe plus production of new blades by the apical meristem results (Figure 1B) in elongation of the frond and increase in the numbers of blades. Eventually the apical meristem reaches the water surface and then is pushed horizontally in the canopy by continued growth throughout the stipe length. Finally, meristematic activity ceases in the apical blade and a terminal blade is formed. A fully grown frond may display (Figure 1B) 100 to 200 or more blades.
A mature frond thus ultimately consists (Figure 1B) of a basal senescent, a central mature and a terminal youthful region as sections of differing ages. The oldest blades occur near the base with a gradient extending upwards to the youngest blades at the apex. Senile deterioration often occurs among the lowest blades of a mature frond even while new youthful blades are developing in the upper regions. The lifespan of an entire frond is about six months but may be somewhat longer or shorter, depending on environmental conditions in the surrounding water. Senile, deteriorating fronds are replaced by young new fronds constantly arising from the basal meristems just above the holdfast. If an apical meristematic blade is severed from a growing frond, further blade production ceases. The remaining basal portion of this frond survives until it is overcome by senility near the end of its normal lifespan.
Harvesting a Macrocystis canopy removes all apical meristems lying above the cutting depth. Canopy regeneration is accomplished by growth from fronds whose apical meristems all lie beneath the cutting depth. The lowermost two to six laminae (called sporophylls [Figure 1A]) at the base of a frond develop reproductive sporangia. These sporangia are densely packed in certain areas, called sori, on the sporophyll blade surfaces. Each saclike sporangium at maturity releases numerous biflagellated zoospores which develop into gametophytes if they attach to a suitable substratum (see below). The chromosome numbers for a haploid M. integrifolia cell are 14 to 16 (Cole, 1968).
Adult Macrocystis sporophytes generally display increased numbers of fronds as they age. Very large plants in protected waters may bear several hundred fronds and may be many years old. In exposed locations, however, storms rip holdfasts loose from their attachments and such plants do not usually survive longer than 2 to 3 years. The largest plants under such circumstances may bear 40 to 60 fronds. Fronds growing from holdfasts in deep water do not produce substantially more blades than are found on comparable fronds growing in shallow water. Fronds compensate for greater water depth by increasing internodal distances (lengths of the stipe sections between adjacent pneumatocysts), not by increasing the total blade numbers.
Figure 1B. Time scale frond development illustrated from consecutive observations of one frond of an adult Macrocystis plant standing in a water depth of 20 m off La Jolla California. The senescent; mature, and youthful regions of the frond are shown but only some of the blades (from North, 1979a).
2.1 Geographical extent
2.2 Local vertical and horizontal distribution
2.3 Effects of ecological determinants
2.4 Nutrition and growth
The genus Macrocystis occurs naturally around every major land mass in the Southern Hemisphere except for Antarctica (i.e. South America, South Africa, Australia, New Zealand), and also along the eastern Pacific coast in the Northern Hemisphere. Presumably a cold water bridge that formed across the Gulf of Panama on the Pacific side of Latin America permitted transport of Macrocystis from South America northward to Mexico, United States and Canada. Macrocystis pyrifera was introduced to Asiatic waters in 1978 by scientists from the People's Republic of China (Liu et al., 1984). The species has also been successfully cultured in North Atlantic waters on at least three separate occasions. Thus it appears that Macrocystis will survive and grow in any of the temperate oceanic waters of our planet. It has not been able to colonize in either tropical or high arctic climates. M. pyrifera and M. integrifolia occur in both Hemispheres. M. angustifolia was described from Australia (Womersley, 1954). Neushul (1971) identified the Macrocystis growing near Santa Barbara, California, as M. angustifolia, based on Womersley's description.
Vertical distribution of Macrocystis is apparently affected more by local conditions than by preferences or a specific range of depths. For example, given year-round suitably low water temperatures, ample nutrients, and protection from wave action, both M. pyrifera and M. integrifolia can colonize low intertidal pools. On exposed coasts, however, the inner limit of large Macrocystis beds is usually at 7 to 10 m depths and is apparently controlled by wave action. The outer limit of Macrocystis beds in California coastal waters usually occurs at depths of 10 - 20 m. The depth range may increase to 25 - 30 m in very clear waters around offshore islands. Presumably the lower depth limit is governed by the bottom illumination required to sustain development of the microscopic gametophytes and small sporophytes. The deepest adult plant ever observed by the author was 40.2 m beneath the surface at Punta Banda, Mexico, an area of extremely clear water.
Local horizontal distributions are most frequently controlled by the availability of rocky substrata. Holdfasts of M. pyrifera or M. integrifolia require solid substrata for firm attachment. Macrocystis angustifolia does occur on sedimentary bottoms. (See Section below.)
The primary ecological factors affecting success and survival of Macrocystis populations in the United States and Mexico (and probably elsewhere in the world) are water motion, nutrient availability, water temperature, competition, grazing organisms, bottom illumination and sedimentation, more or less in that order of importance. We discuss nutrients and grazing below, so primary emphasis here will be on wave action, water temperature and illumination.
Importance of wave action as a disruptive factor is demonstrated by the distribution of Macrocystis in southern California, comparing the well-protected Santa Barbara coastline with the remaining exposed habitat. Harger (1983) reported that the coastline from Santa Barbara to Point Conception (including San Miguel Island) contained 65 percent of all kelp bed areas in southern California along only 18 percent of the entire shoreline. Variation of canopy coverage was smallest for the Santa Barbara region. These statistics suggest that some degree of protection against wave action is very beneficial for increasing area of coverage and stability of kelp beds.
Studies of plant mortality in exposed kelp beds clearly indicate that maximum life span of adult plants is short, usually around two to three years for most of the population (North, 1971a; Rosenthal et al., 1974; Dayton et al., 1984). Juvenile recruitment thus is necessary to assure continuation of the population. While wave action can be destructive, water motion is also necessary for nutrient renewal. Back and forth motion such as caused by waves creates a scrubbing action across blade surfaces enhancing nutrient exchange (W.N. Wheeler, 1982; Gerard, 1982a). Longer period motion resulting from tidal cycles, internal waves, and alongshore current replenishes dissolved nutrients within beds and transports food and waste products for the associated fauna. Resistance to flow reduces current velocities within the kelp beds and causes deviation of flow around kelp stands (Jackson and Winant, 1983).
Nutrient availability is negatively correlated with water temperature for values greater than 15.5°C (Zimmerman, 1983). Consequently, the many reported observations of summertime deterioration by Macrocystis attributed to high temperature, probably represent combined effects from elevated water temperatures and low nutrients (Jackson, 1977; Zimmerman and Kremer, 1984). North and Zimmerman (1984) showed that fertilizing an experimental group of kelp plants resulted in substantial canopy preservation during summertime compared to an unfertilized group of control plants. In natural beds, stratification in the water column during summer results in a warm, nutrient-poor environment above the thermocline and a cool, nutrient-enhanced milieu below. Consequently, kelp deterioration is greatest in the surface layers while basal portions may survive with little, if any, damage. When sea surface temperatures fall during autumn, basal portions may regenerate the canopies in a few weeks or months.
Summertime deterioration is typically accompanied by a number of symptoms or signs which may conveniently be grouped into a single term we shall call "summer syndrome". Most of these symptoms can be seen within kelp beds at other times of the year as well but less frequently. Prevalence may simply increase sharply during warm summers. Summer syndrome includes loss of coloration in kelp tissues, lesions in the blades, tissues may become brittle and break easily or they may become soft and slough away, coverage by encrusting animals may increase, bladders become perforated or may simply fill with liquid so that fronds lose buoyancy and sink, apical blades become scarce and roost fronds display terminal blades. At a physiological level, photosynthetic capacity declines (Gerard, 1984a). Some of these symptoms undoubtedly arise from nutrient insufficiency (i.e. loss of color). Others may be indirect effects of increased temperature (e.g. lesions and sloughing, caused by enhanced microbial activity). Clendenning (1971) reported that light-saturated photosynthesis was always highest over the range of 20° to 25°C among all kelp tissues he studied (except for kelp from central Baja California which peaked in the range 25° to 30°C) during short term exposures. Summer syndrome, however, usually develops at around 20°C. Appearance of summer syndrome apparently involves relationships between duration of a period of elevated temperature and the level to which temperature rises. Thus a temperature of 20° may be tolerated for 2 or 3 months while 23°C or more for a month will result in rapid deterioration.
The microscopic stages of Macrocystis may be more sensitive than adult plants to elevated water temperatures. Dean and Deysher (1983) reported that the upper limit for sporophyte production from gametophytes they outplanted to a natural kelp bed was 16.3°C.
Competition from other algal species takes several forms including shading of juvenile Macrocystis (i.e. competition for light), competition for space, substratum, and possibly competition for nutrients. There may also be poorly understood competitive mechanisms. For example, small Macrocystis and other lami-narian sporophytes appear to recruit poorly on substrata thickly covered by encrustations of coralline algae and animals when compared to non-encrusted surfaces. On the other hand, remains of old decaying Macrocystis holdfasts sometimes support numbers of young sporophytes while none occurs on the surrounding bottom. These observations suggest that chemical mechanisms may exist which affect suitability of a substratum for colonization by kelp spores. Barren rocky bottoms severely and constantly grazed by sea urchins are usually excellent substratum for colonization by Macrocystis and other brown algae after the urchins are eliminated; provided, of course, that fertile adult plants are close by enough to serve as spore sources. When territory dominated by Macrocystis is lost due to some catastrophic effect, other seaweed species may colonize the area and prevent Macrocystis from reestablishing. Persistence, resistance to invasion, and resilience or recoverability after loss of territory are discussed by Dayton and Tegner (1984) and by Dayton et al. (1984). Physical abrasion from nearby blades of tough species such as Laminaria and Agarum may damage small Macrocystis sporophytes, (R. McPeak, pers. comm.).
Significant grazing damage can be inflicted on Macrocystis plants by several herbivores including sea urchins, crustaceans, mollusks and fishes. Probably the most damaging grazers in California waters are the sea urchins, Strongylocentrotus franciscanus, S. purpuratus and Lytechinus anamesus. These echinoids can occur in enormous populations at concentrations well above 10/m². The animals migrate slowly across the sea floor, consuming all plant life except for crustose corallines, as well as many sessile invertebrates such as sponges, tunicates, and bryozoans. Under some conditions, urchin populations persist indefinitely at high concentrations, preventing vegetation from reestablishing and creating "urchin barrens" (Dean et al., 1984). Urchin barrens have been observed in many shallow benthic areas other than California. Disappearances or significant reductions of predators that control urchin numbers permit development of huge urchin populations. Urchin predators include lobster, fishes, sea otters, and asteroids (Tegner and Levin, 1983; Dayton et al., 1980; Estes and Palmisano, 1974). Human exploitation of urchins as food may also act as a significant control, (Wilson and North, 1983). Very severe storms may even remove urchins from kelp bed depths (Ebling et al., 1984). Techniques for artificially controlling urchin populations are described below.
Herbivorous fishes are not usually sources of significant damage to Macrocystis populations in dense healthy kelp forests because the amounts of plant tissue consumed are small in relation to total productivity. Fish grazing may assume importance when productivity declines as, for example, in a kelp bed under stressful conditions or where small plants colonize the sea floor after the adult Macrocystis plants were destroyed by some catastrophe. Two herbivorous fishes in California waters, Girella nigricans and Medialuna californiensis, preferentially feed on youthful kelp tissues such as the apical meristems and the blades of very young sporophytes (Harris et al., 1984). A third fish species, Oxyjulis californiensis adventitiously removes chunks of kelp tissue when feeding on encrusting bryozoans (Berstein, 1977), sometimes damaging blades significantly. For grazing of small sporophytes or apical meristems, the damage inflicted may be far out of proportion to actual amounts of tissue consumed. Effects from fish grazing may be especially significant when small young Macrocystis plants are first reestablished in an area where a catastrophe has eliminated all the adults.
Leighton (1971) studied food preferences of eleven invertebrate herbivores common in California kelp beds. All eleven showed preferences for Macrocystis tissues. He did not include herbivorous fishes in his studies but our field observations suggest that Macrocystis is strongly preferred by Girella and Medialuna. We frequently record instances where extensive damage from fish grazing occurs on blades of small Macrocystis plants but none is seen on other recruiting kelp species such as Eisenia, Egregia, Pterygophora, and Laminaria. Fish grazing undoubtedly enhances competition from other kelp species on such occasions. Grazing fishes can be a serious problem when we undertake to develop Macrocystis stands by transplantation techniques (see below).
Grazing by crustaceans and gastropods infrequently may cause significant damage to Macrocystis populations. Numbers of these herbivorous species are usually so small that their activities only result in trivial losses of kelp tissue. Very occasionally, however, dense concentrations of a species may appear so that grazing effects reach destructive proportions. We have observed significantly destructive grazing on Macrocystis by large numbers of kelp crabs (Pugettia), abalone (Haliotis), dove snails (Mitrella), and microcrustaceans (primarily Ampithoe) and Idothea).
Adequate sunlight is, of course, a requirement for all plants, but effects of cloudy days or turbid water are less for adult Macrocystis than for short-statured algae because of the extensive canopies which extend just beneath the surface where submarine illumination is maximal (cloudy days may affect kelp productivity but not survival of adult plants). The primary influence on Macrocystis populations by attenuation of submarine light is the effect on microscopic plants and recruitment of small plants. Dean and Deysher (1983) outplanted gametophytes to a natural kelp bed at San Onofre, California, and after six weeks recorded densities of resulting sporophytes on their artificial substrata. They found that no sporophytes were produced at irradiances below 0.4 E/m²/d. They also noted an interactive effect between water temperature and irradiance. As temperatures approached 16°C, about 1E/m²/d was required for moderate sporo-phyte production. These authors also measured daily irradiance on the bottom in clear areas within the San Onofre kelp bed for 3 1/2 years (July, 1978 to January, 1982). Favorable irradiances over 2-week intervals combined with suitable bottom temperatures occurred only eight times during the 3 1/2 year interval and each recruitment was traceable to a period of favorable light and temperature as defined by Dean and Deysher's measurements. These authors postulated that "windows" of environmental conditions favoring recruitment occurred only occasionally and were critical to persistence of this kelp bed. Photosynthesis by Macrocystis gametophytes and embryonic sporophytes became light saturated at 70 m E/m²/s (Fain and Murray, 1982). Compensation irradiance for embryonic sporophytes was 2.8 m E/m²/s. Luning and Neushul (1978) estimated that 20 m E/m²/s was adequate for saturation of vegetative growth in M. pyrifera and M. integrifolia gametophytes but 2 to 3 times this irradiance was necessary to induce gametogenesis.
Sedimentary deposits can affect M. pyrifera in several ways. Even very light sedimentary films may interfere with spore settling under laboratory conditions (Devinny and Volse, 1978). An approximately tenfold increase in sedimentary load caused burial of already settled spores and interfered with their survival. Presumably due to sediment scouring, water motion in the presence of fine sediment further reduced survival. Dean and Deysher (1983) noted a substantial influence from sedimentation on sporophyte production by their gametophyte outplants to the San Onofre kelp beds (in their regression analysis, sedimentation accounted for 11% of the variance observed, temperature 25%, light 10%, nitrogen 0%). The thin sedimentary deposits that may bury microscopic stages of Macrocystis are, of course, ineffective against larger plants. Major sedimentary shifts have been noted, however, damaging young and even adult sporophytes (Neu-shul, 1959; North, numerous observations, unpub.).
Partial burial of holdfasts does not appear to affect haptera adversely, e.g., the haptera of M. angustifolia actively growing downward into the sedimentary bottom. The haptera penetrate deeply into the sediments, and interstices within that part of the holdfast above the bottom become filled with sand or mud. This plant produces holdfasts that may be 3 to 5 m in diameter but less than one meter tall (i.e., low mounds). By trapping and penetrating the sediment, it creates a heavy mass that withstands displacement by wave action.
Burial of the primary or secondary stipes at the holdfast apex is not tolerated well. Stipe tissues rot in a few days if they are not freely exposed to surrounding water. Blades also are intolerant of even light sedimentary deposits and may disintegrate after a few weeks of coverage. This type of damage is common among Macrocystis plants growing in quiet waters of bays and inlets where scrubbing action by wave surge is lacking. The Santa Barbara region, however, is protected from long period waves by offshore islands and orientation of the mainland.
Ecological determinants of lesser importance affect Macrocystis from time to time. Aggregations of encrusting animals may appear on Macrocystis blades so densely as to cause loss of buoyancy and sinking of fronds. Encrustation of kelp by the Bryozoan, Membranipora, enhances blade loss by reducing flexibility and by encouraging fish grazing on the Bryozoan colonies (Dixon et al., 1981; Kirkwood, 1977). Compensating light intensity was about 50% higher for Membranipora encrusted than for clean Macrocystis blades (Wing and Clendenning, 1971). Isolated plants and those near the edges of kelp beds are more susceptible to colonization by Membranipora (Bernstein and Jung, 1979).
Salinity remains sufficiently constant along open coastlines so that distribution of Macrocystis populations is rarely, if ever, affected in California. Lowering of salinity in bays and estuaries may regulate Macrocystis distribution in these habitats, especially north of southern California where rainfall increases. According to Druehl (1979), beds of M. integrifolia are able to grow at Nootka, Vancouver Island, where the yearly salinity range was from about 23 ‰ to 30 ‰. Druehl indicated that high temperatures (i.e. 14° - 18°C) during periods of low salinity were associated with absence of Macrocystis and that the season of low salinity at Nootka coincided with a time of low temperatures (8° - 12°C). North (unpub.) grew gametophytes of M. pyrifera in serial dilutions of open ocean seawater and distilled water. Gametophytes did not survive at dilutions greater than 3 to 1 (i.e. just below 25 ‰ salinity if we assume the salinity of the undiluted seawater was 33 ‰. This conclusion was in rough agreement with Druehl's (1979) observations.
Kelp growth is probably rarely, if ever, limited by availability of micronutrients in surface seawater. Kuwabara and North (1980) demonstrated nine elemental requirements by Macrocystis including the macronutrients nitrogen and phosphorus as well as the micronutrients copper, zinc, cobalt, manganese, iron, molybdenum and iodine. Kuwabara (1982) found indications that sea-water from 300 m depths in southern California was somewhat deficient in manganese and cobalt for growth by Macrocystis gametophytes.
Nitrogen may at times be a limiting element in surface water and very occasionally phosphorus (Manley and North, 1984). Luxury uptake of nitrogen and phosphorus occurs in Macrocystis during times when these elements are plentiful and reserves are utilized when external supplies are inadequate (Gerard, 1982b; Manley and North, 1984). These authors indicated that values of critical tissue levels (where the element is neither in excess nor in short supply) for Macrocystis were 0.2% dry wt. for Phosphorus and 1.0% dry wt. for Nitrogen in blade tissues. P.A. Wheeler and North (1980) found that blade N-contents of juvenile sporophytes reached a maximal value of 3% dry weight when external nitrate concentration in a flowing water culturing system was at 13 m M. Gerard (1982b) transplanted large adult sporophytes with laminar tissues initially containing 2.2 to 3.5% dry wt. Nitrogen from a coastal kelp forest to an offshore island site where ambient external Nitrogen was well below m M (as indicated by continual temperature measurements). Growth rates stayed at relatively high levels for two weeks, then declined sharply. Ambient Nitrogen rose during the fourth week and growth rates recovered somewhat. The Nitrogen contents of laminae declined steadily during the initial 3 weeks of starvation, then remained constant or rose slightly thereafter. Holdfast Nitrogen remained constant throughout. Gerard (1982c) further proposed a mathematical model for nitrate uptake by whole plants. Zimmerman and Kremer (1984) demonstrated a relationship between estimated nitrate concentration and frond growth rate. Growth rates fell for nitrate concentrations below 1 m g-at/1.
Haines and P.A. Wheeler (1978) listed values of KS=13.1 m q-at N/1, Vmax=3.05 m g-at N/wet g/h for NO3 uptake by juvenile Macrocystis and KS= 5.3 m g-at N/1, Vmax=2.38 m g-at N/wet g/h for NH4 for concentrations below 22 m g-at/1. W.N. Wheeler (1978) reported maximum uptake rates by tissue discs cut from blades on an adult frond as 75 m m/cm²/h for NO3, 275 nM/cm²/h for NH4, and 3.8 nM/cm²/h for PO4 under saturating water motion. All workers reported that nitrate and ammonium were taken up simultaneously without any effects on uptake rates by presence of another form of nitrogen. Manley (1985) found values of KS= 3.51 m M and Vmax= 5.29 nM/cm²/h for PO4 uptake by blade tissue discs from adult sporophytes. Manley suggested that differences between his results and those of W.N. Wheeler might arise from differences in tissue Phosphorus levels. Gerard (1982c), using whole blades attached to adult plants, showed that NO3 uptake rates differed substantially for tissues from different parts of the plant and similar findings have been shown for PO4 uptake (Manley, 1985; Schmitz and Srivastava, 1979) and for NO3 uptake by M. integrifolia (W.N. Wheeler and Srivastava, 1984). Uptake of NO3 and PO4 was more rapid in light than in darkness or under shaded conditions (W.N. Wheeler, 1982; Gerard, 1982a; Manley, 1985). Culturing tissues in water high in NO3 or PO4 reduces uptake of these nutrients and Gerard (1982b) found an inverse relation between depth and NO3 uptake. W.N. Wheeler (1978, 1980a, 1982) found that water motion affected carbon and nutrient uptake rates with uptake saturating in the range of 3 to 5 cm/sec current velocity. Gerard (1982c) concluded that NO3 uptake regardless of concentration, became saturated at a water flow of 3 cm/sec. She found that water motion under field conditions exceeds this critical value even inside dense canopies during calm seas. W.N. Wheeler (1980a) reported that transport rate of inorganic carbon through the boundary layer on Macrocystis blades saturated at flow velocities of 4 cm/sec.
Uptake rates of the micronutrients I, Fe, Zn, Co, and Mn as a function of external concentration were presented by Manley and North (1981), using an artificial seawater. Manley (1984) showed that these elements as well as Mo and Ni were concentrated in sieve tube sap. He calculated that adult plants sometimes might not be able to fulfill their requirements for Mn and Co, resulting in growth limitation by availability of these elements. Manley (1981) showed that Fe uptake is light independent and energy dependent. Manley (1.c.) and Anderson (1984) both showed that iron is reduced from Fe3+ to Fe2+ during uptake.
Ability to translocate inorganic and organic ions and compounds is an important feature of nutrition in the Laminariales. Structural elements of the translocation system are maximally developed in the tree-like kelps such as Macrocystis. Translocation processes nourish portions of adult plants growing under low illumination near the bottom by photosynthate created in the upper well-lit parts of the water column. The translocation capability allows Macrocystis plants to grow in dense aggregates with over-lapping canopies which effectively shade out competitors on the bottom, yet supports rapid growth by young fronds, sporophylls, haptera and other tissues near bases of the plants. The sieve tubes occurring around the periphery of medullary tissue centrally within blades and stipes, are the conducting pathways supporting translocation. The sieve tubes are enclosed by cellular membranes. Osmotic pressure is believed to be the driving force causing movement of fluid through the sieve tubes (Schmitz, 1982). Synthesis of organics and rapid uptake of inorganics in the "source" region of a frond creates high concentrations of solutes there, creating a tendency for intake of water from outside and movement of water plus solutes through the sieve tubes towards more diluted solutions in the "sink" regions. Accommodation of the flow is accomplished by loss of water in the sink region. Flow can be bidirectional within a single mature frond (i.e. from the center toward both ends). Thus, both apical meristems in the upper parts of the water columns and newly developing fronds at the plant base receive photosynthate and nutrients from mature blades located in the central part of a long frond.
Lobban (1978a) showed that newly developing fronds of M. pyrifera act as sinks until they are approximately three meters long. Loss or disappearance of an apical meristem caused all translocation to occur toward the base. Translocation in M. integrifolia differed from that in M. pyrifera in terms of seasonal changes in direction of translocation and in distances from the apices where changes occur (Lobban, 1978b). Manley (1984) postulated that young short fronds probably depend on translocation of Mn, Co and Fe to meet growth requirements because their small total blade areas provided insufficient uptake capacities.
Translocation velocities of 25 to 45 cm/hr were measured using radioactive 32p compounds as tracers in M. integrifolia (Schmitz and Srivastava, 1979). These velocities were comparable to velocities shown for 14C compounds. Translocation velocities in M. pyrifera blades were only 6 to 22 cm/hr., possibly because of small diameter pores in sieve plates forming the interconnections between adjacent sieve tube cells (Buggeln et al., 1985).
Phosphorous is assimilated from the exterior in inorganic form but is primarily incorporated into organic compounds during translocation in the sieve tube sap. Similarly, organic forms of nitrogen are taken up very slowly or not at all by M. pyrifera (P.A. Wheeler, 1979), yet nitrogen in sieve tube sap occurs almost entirely as amino acids (Jackson, 1976). Jackson (1977) calculated that the flux of nitrogen due to translocation in Macrocystis stipes amounted to 0.2mM/hr, while that of carbon was 2mM/hr. The principal amino acid in M. pyrifera was alanine.
The initial product of photosynthesis in Macrocystis is mannitol (Vaughn, 1959) and the primary low molecular weight carbohydrate was mannitol in both M. pyrifera and M. integrifolia (Jackson, 1.c., Schmitz and Srivastava, 1979).
The basis of growth and productivity in Macrocystis, as in all plants, is photosynthesis. Photosynthesis in an adult Macrocystis sporophyte is affected by a number of factors, resulting in an extremely complex situation. Different tissue types (i.e. blades, stipes, sporophylls, haptera) display different photo-synthetic capacities (Arnold and Manley, 1985). Gradients in photosynthetic capacities exist from base to tip within blades and between blades along the frond length (Arnold and Manley, 1.c.; W.N. Wheeler, 1980b; Clendenning, 1963). The blades account for nearly all the photosynthetic activity of Macrocystis sporophytes in nature). The submarine light field that drives photosynthesis is extremely variable and complex in kelp beds as a consequence of canopy absorption, refraction by ripples and waves, and constant movement by blades resulting in flashing light patterns in the upper part of the water column (Gerard, 1984b). Vertical distribution of blade area is not uniform (Jackson et al., 1985) with 50% or more of the total area occurring near the surface in adult plants displaying full canopies. Very substantial seasonal changes in photosynthetic capacity may occur for both M. pyrifera (Clendenning 1971) and M. integrifolia (Smith et al., 1983) due to changes in water temperature, availability of micronutrients and general healthiness of the blades. Jackson (personal communication) is developing a computer-based mathematical model of kelp bed photosynthesis and productivity that takes account of the many factors affecting photosynthesis in Macrocyst is. The model is still in a preliminary state of development, but in its present state has yielded interesting predictions as to effects of planting density, changes in latitude, and seasonal changes in plant size (the seasonal changes in plant size predictions were confirmed by measurements conducted by Kirkwood, 1977, in a Macoystys bed at Palos Verdes, California).
Growth in Macrocystis, on a short term basis, consists of development by individual fronds and haptera and, over the longer term, increase in holdfast size and total numbers of fronds per plant. The short term growth parameters can be easily determined through in situ measurements of hapteral and frond lengths by divers. Long term results of growth can also be estimated in situ (measuring holdfast dimensions, counting numbers of fronds), or by removing plants from the water and weighing them (Neushul and Harger, 1983; the latter technique requires correction for added weights from extraneous factors such as entrapped water and sediment as well as encrusting organisms). Gerard (personal communication) developed a technique for estimating changes in frond biomass based on in situ measurements. First, a relationship is determined for frond length vs frond weight by sampling fronds from a target population. Then the frond size distribution pattern is determined by measuring overall lengths of a large number of randomly selected fronds from the population in situ. Concurrently, frond density is estimated in situ by counting all fronds within randomly positioned quadrats of known areas. Biomass per unit area is then estimated as the sum of the mean weights of the various size classes of fronds, adjusted for the frequency of occurrence of each size class in the total frond population, multiplied by the density of fronds per unit area. The determinations are repeated at a later date and the difference between the two sets of measurements provides an estimate of biomass change during the interval. The productivity of each frond size class can be assessed by measuring elongations of fronds in the population over a given time period and calculating the resulting weight increment from the length-weight relationship. Summing productivities of each frond size class (adjusted for frequency of each size class) yields productivity by the entire frond population.
Methods for assessing frond growth have been variously proposed (North, 1971b; Kain, 1982; Gerard, 1982b; Grua, 1964; Manley, 1984). Changes in mean frond growth rates are useful for assessing healthiness of Macrocystis populations. Frond growth rates, measured either as length or weight changes, are hot uniform with time, but change as the frond ages. It is thus necessary that measurements either be made on a sample of fronds all of similar ages or that corrections for the age factor be introduced if the sample includes fronds of various ages.
The Macrocystis life cycle (Figure 1A) consists of a dimorphic alternation of generations between a macroscopic diploid sporophyte and microscopic haploid gametophytes. Biflagellated zoospore production begins after the sporophytes are 6 to 12 months old. Spores are transported away from the parent plant primarily by natural water movements and secondarily by their own swimming activity. Those settling on suitable substrata produce germ tubes in a few hours. The spore content migrates down the germ tube in producing the first cell of the gametophyte (a few gametophytes apparently do grow directly from the spore without intervention of a germ tube). The initial gametophytic cell grows and divides into one or two cells (females) or several cells (males) in a week or two.
Male and female gametophytes can be differentiated after about a week in laboratory cultures by color and shape. Males are pale or bluish, the cells are small, and the overall shape is often filamentous. Females are yellow-green, gold, or brown with round or ovoid shapes. Gametogenesis may begin in 7 to 10 days. Antheridia on males liberate biflagellated sperm. The entire gametophyte in single celled females may form the deeply pig-mented ovum, or only one cell may become an ovum in multicellular females. All cells, however, may eventually develop into ova.
The ovum is extruded from the oogonium and becomes fertilized. Extrusion may or may not rupture attachment of the ovum to the oogonium. If detached, the ovum or zygote is capable of settling elsewhere and forming a new attachment. Rapid cell division in the zygote produces the embryonic sporophyte. Within a few weeks, growth yields tiny blades just visible to the naked eye.
The single blade stage in Macrocystis closely resembles that of other Laminariales. Positive identification as Macrocystis can be made when traces of the first or primary split occurs immediately at the junction of blade and stipe which eventually creates two separate blades or basal meristems (See Figure 1A). Further splitting and appearance of frond initials leads to development of the adult morphology in the young plant.
Although spore production is enormous (Neushul, 1959, estimated there were 10,000 sporangia/cm² in Macrocystis sporo-phylis), probably only a small fraction ever settle on suitable substrata and vastly smaller numbers survive through the microscopic phases of the life cycle (Anderson and North, 1966; Deysher, 1984).
Young sporophytes normally appear at only distances of a few meters to ca 100 m from attached plants. Drifting adult sporophytes, however, may disperse spores in areas lacking attached Macrocystis, possibly providing opportunities for establishing the species.
4.1 Age, weight or size composition
4.2 Sporophyte - gametophyte sex composition
4.3 Mortality, morbidity
Natural Macrocystis populations vary widely in their population structures. Sometimes nearly all plants will be of similar sizes and ages while in other cases plants of varying ages and sizes occur intermixed. The former type of age-size structure sometimes arises after a major catastrophe such as an exceptional storm destroys almost all Macrocystis throughout a large area. Normally, the bottom in such an area would be well seeded with microscopic spores and, thus, gametophytes. Removal of most adult plants would increase bottom illumination dramatically, stimulating garoetophytic growth and production of embryonic sporophytes. Assuming good survival, a population of adult sporophytes of uniform age would eventually develop. Once a canopy is formed, further recruitment of juvenile plants ceases and only the oldest plants with fronds in the canopy receive enough sunlight to survive. The shorter plants perish for lack of sunlight, leaving a population of uniform age and size. If a few of these survivors perish more or less randomly as time passes, canopies of the remaining plants will still prevent recruitment of juveniles. Random attrition and continued growth by the survivors produces a population of scattered old plants with large holdfasts, each supporting many fronds.
Populations of variously sized plants occur where forces of attrition are high so that mortalities occur frequently. Patches of open water can appear between adult plants, allowing enough sunlight into the water column to support areas of juvenile recruitment during periods of clear water. Thus, plants of all sizes and ages coexist (for harvesting effects, cf. Kimura, 1980).
One of the major agents of attrition in Macrocystis forests is dislodgement of plants by storm-generated waves. Storm-attrition is greater the shallower the water. Consequently, the offshore edges of kelp beds experience less storm attrition than inshore edges. We frequently observe uniformly large and widely spaced old plants along the offshore borders of Macrocystis beds, while inshore portions of the beds support mixed sizes of fairly young plants, often in dense aggregates.
A unit commonly employed for assessing size among adult Macrocystis plants is the number of fronds. The typical adult Macrocystis is composed of fronds of varying lengths ranging from initials a few cm long to fully mature fronds with 5 to 10 m or more of their length stretching horizontally in the canopy (the longest frond I ever measured was 44.8 m and probably 3 m was missing from the distal end). Although frond lengths on a given plant vary, frond size distribution per plant stays within narrower limits (North, 1968), particularly for populations growing at a given depth. The average weight of all fronds in a population thus tends to be similar, increasing slightly with depth. Thus the mean wet weight of all fronds from plants growing at depths of 10-20 m is slightly more than 1 kg, while for plants growing at 20-25 m depths, the mean wet weight is ca 1.5 kg (North, 1958). Total biomass in a population can be estimated by multiplying frond density (obtained by counting all fronds at the plant bases occurring within several randomly-located quadrats in the bed) by the mean wet weight of a frond (i.e. ca 1 kg). For natural kelp beds with coherent canopies, a range of biomasses of 3 to 22 kg/m² was reported by North (1971a).
Nothing is known about proportions of Macrocystis sporophytes to gametophytes in natural populations, or of the sex composition of gametophytes. The microscopic dimensions of gametophytes make them difficult to detect on natural substrates such as rock or hard organic material. Fates of gametophytes and sporophytes have been followed on outplanted artificial substrates (Dean and Deysher, 1983), and recruitment has been monitored on concrete blocks (Foster 1975a, 1975b). Outplanted Macrocystis gametophytes survived only 40 to 45 days off San Onofre, California (probably a fairly hostile environment). In laboratory cultures, approximately equal numbers of male vs female gametophytes develop from settled spores.
Rosenthal et al., (1974) monitored tagged adult Macrocystis in a bed off Del Mar, California for 5.7 years and reported that the primary causes of mortality were: storms, entanglement between drifting and attached plants, and kelp harvesting (holdfasts were presumably pulled loose during cutting). North (1971a) studied Macrocystis mortality by tagging plants and following their disappearances over a few months, computing half-lives by assuming that mortality remained fairly constant (a half-life is the time required for 50% of a tagged population to disappear). Half-lives ranged from two months for very exposed populations during winter, to several years for well-protected plants off Santa Barbara, California. Similar results were reported by Dayton et al. (1984).
Gametophyte mortalities are probably much greater. Deysher (1984) found that Macrocystis gametophytes outplanted on rope substrata, survived only 40 to 45 days off San Onofre (as judged by cessation of sporophyte production on the ropes). San Onofre is believed to be a somewhat hostile environment for gametophytes because of sedimentation (Devinney and Volse, 1978, showed that even light sedimentary deposits were very harmful towards Macrocystis gametophytes).
5.1 Productivity and harvesting yield
5.2 Principal factors affecting productivity
5.3 Genetic studies and breeding studies
5.4 Relative contributions from vegetative & sexual reproduction to the harvest
5.5 Environmental enhancement
Biomass production by plants depends on availability of sunlight and nutrients. Most of the photon flux through the water surface is presumably utilized when plant biomass is distributed so as to completely cover the surface, as in dense Macrocystis canopies. Nutrient availability is influenced not only by concentrations of nutrients in the surrounding waters, but also, by water motion and amounts of plant surfaces exposed for taking up nutrients. Ryther and coworkers have shown that approximately parabolic relationships exist between production and density of biomass when aquatic plants and seaweeds are grown under light limiting conditions (i.e. water motion and nutrients are not growth-limiting). That is, there is an optimum density of plant tissue at which biomass production and yield are maximal. The optimum tissue density is different for differing kinds of plants (e.g. 0.02 kg dry/m² for Lemna minor, 0.5 to 1 dry kg/m² for Eichornia crassipes, Ryther et al., 1979) and is affected somewhat by season (Ryther et al., 1977).
Neushul and coworkers attempted to determine the optimal biomass density for maximizing yield from Macrocystis angusti-folia by growing adult transplants at three biomass densities (initial values 1.7, 6.4 and 24.3 fronds/m², representing 0.0625 to 0.25 and 1 plant/m²) on test plots. Half of the plots were fertilized with ammonium sulfate, half were unfertilized. Harvested yields varied seasonally and were highest for the period May-July for all three planting densities (Neushul and Harger, 1983). Projected yields (i.e. assumes that no frond was lost and that all fronds produced were eventually harvested) from five harvests the yields averaged 5.6, 13.9 and 24.8 DAFMT/ha/yr (dry, ash-free metric tons per hectare per year; presumes that one DAFMT =1.786 dry ton = 14.286 wet ton) from the three densities respectively. Total yields from the fertilized plots were slightly higher than from the unfertilized areas for the two highest densities, but yields from the fertilized plots for all three densities were substantially higher for the July-September period, a time when ambient nutrients were low (Neushul and Harger, 1983). The study thus indicated that the optimal biomass density for Macrocystis lies above 24.3 fronds/m². Although I have measured a density of 30 fronds/m² in a natural Macrocystis bed, it seems likely that such very high densities cannot be sustained for long. During the course of this 15 month study, Neushul and Harger (1.c.) reported that plants in their high density plots decreased in size so that the overall frond density declined by a half during the study. North (1971a) found a range of 2 to 15/fronds/m² for large size samples from Macrocystis beds in California and Mexico. Most populations fall in the range of 3 to 6 fronds/m².
Gerard and North (1984) measured production and harvested yield on a group of 63 adult M. pyrifera held for 2 1/2 months in an enclosure with intermittent fertilizing (9-13 uM NO3 and 1 uM PO4). Deterioration in the basal portions of the plants occurred, probably from lack of water motion due to the enclosure. In spite of these difficulties, they estimated that 50% of the total productivity was recovered as harvested yield.
Considerable data are gathered by the California Department of Fish & Game as to fresh weights of Macrocystis harvested from beds within the State. In most cases, kelp bed areas involved in the harvesting were not distinguished. Neushul et al., (1982) conducted a special study in cooperation with a kelp harvesting concern in which both the weight of kelp and the area harvested were determined for a kelp bed off Goleta, California. They obtained a yield of 8.3 DAFMT/ha/yr. A similar one year study by North et. al., (1982) for a fertilized bed off Leucadia, California/gave a yield of 12.3 to 14.8 DAFMT/ha/yr. Barilotti (pers. comm.) reported that yields from the kelp bed off Pt. Loma, California, ranged from 1.2 to 4.9 DAFMT/ha/yr.
The total annual harvest of kelp (primarily Macrocystis) from California waters shows considerable year to year variation resulting from interactions of both environmental, economic, and other factors (Table 1). Data from Neushul (1981a) suggest that areas of canopies have varied substantially, probably accounting for some of the variations in total harvest. (Examples of canopy variation are shown in Figures 2 and 3 below). Macrocystis was harvested maximally from 1916 to 1918 and from at least 1955 onward because of favorable economics. Kelp was used as a source of potash during World War I, but the demand collapsed after the war terminated (Scofield, 1959). Interest in kelp for animal feed additives and chemicals led to renewed harvesting activity in the late 1920's. Demand for alginate products caused steady increases in harvests in the latter 1930's, the 1940's, and has sustained the industry ever since. High water temperatures, unusual storms and reduced availability of nutrients associated with major El Nino oceanographic events, were responsible for reduced harvests in certain years (1957, 1959, 1983).
Table 1. Estimated southern California kelp areas in hectares (modified from Neushul, 1981a) and annual Macrocystis harvest from California waters in wet metric tons (Pinkas, 1977 through 1976; subsequent figures courtesy of K. Wilson and E. Smith, California fish and Game). The area of kelp in hectares is underscored at the head of the respective list of paired years and tonnages.
|
Total area |
Total area |
Total area | |||
|
Year |
Tonnage |
Year |
Tonnage |
Year |
Tonnage |
|
13,171 |
1949 |
75,610 |
4,753 | ||
|
1911 |
|
1950 |
91,265 |
1967 |
119,290 |
|
1916 |
122,050 |
1951 |
104,109 |
1968 |
122,337 |
|
1917 |
358,314 |
1952 |
99,935 |
1969 |
119,058 |
|
1918 |
358,425 |
1953 |
114,894 |
1970 |
115,248 |
|
1919 |
15,125 |
1954 |
96,358 |
|
|
|
1920 |
23,101 |
|
|
5,971 | |
|
1931 |
236 |
7,110 |
1971 |
141,121 | |
|
1932 |
9,358 |
1955 |
112,548 |
1972 |
147,428 |
|
1933 |
19,614 |
1956 |
106,881 |
1973 |
138,872 |
|
1934 |
14,406 |
1957 |
84,961 |
1974 |
154,386 |
|
1935 |
27,762 |
1958 |
103,475 |
1975 |
155,671 |
|
1936 |
44,740 |
|
|
1976 |
143,672 |
|
1937 |
39,874 |
6,510 |
|
| |
|
1938 |
43,270 |
1959 |
81,283 |
6,202 | |
|
1939 |
51,470 |
1960 |
109, |
1977 |
143,918 |
|
1940 |
53,528 |
1961 |
117,259 |
1978 |
186,270 |
|
1941 |
50,546 |
1962 |
127,217 |
|
|
|
1942 |
56,153 |
|
|
4765 | |
|
1943 |
43,507 |
2,818 |
1979 |
188,464 | |
|
1944 |
48,108 |
1963 |
109,798 |
1980 |
162,693 |
|
1945 |
53,688 |
1964 |
115,443 |
1981 |
83,822 |
|
1946 |
82,616 |
1965 |
122,587 |
1982 |
95,326 |
|
1947 |
67,347 |
1966 |
108,376 |
1983 |
5,334 |
|
1948 |
71,342 |
|
|
1984 |
47,320 |
5.2.1 Physical factors
As noted above, wave exposure is reckoned as the primary environmental factor causing losses of kelp tissues and entire plants on exposed coasts (Rosenthal et al., 1974; Gerard 1976). High water temperatures and lack of nutrients lead to losses of canopy tissues during warm summers (Jackson, 1977; North, 1983). A combination of record storms, warm water and low nutrients associated with an El Nino event in 1983, destroyed more than 90 percent of the kelp canopies in southern California, leading to the poorest annual harvest yield in recent times (see Table 1). Factors affecting recruitment of juvenile plants have a delayed influence on the adult population. Increased sedimentation from construction projects and discharged waste materials may have temporary or long-lasting effects.
5.2.2 Biological factors
Grazing by sea urchins has led to declines and, in some cases, complete disappearances of major kelp areas in California and Baja California during the period 1940 to 1965 (Leighton et al., 1965). Probably declines in major urchin predators such as sea otters, lobster, and certain fishes eliminated some of the natural factors controlling size of urchin populations (Tegner and Levin, 1983; Leighton et al. 1.c.). Programs of artificial control of urchins were associated with reappearances of Macrocystis beds at Pt. Loma and Palos Verdes, California, in the 1960's and 1970's respectively (Figures 2 and 3).
Grazing by herbivorous fishes does not usually reach destructive proportions in large established Macrocystis beds. Fish grazing may cause significant difficulties, as noted above, when attempts are made to introduce transplants of adult or juvenile kelp into areas where no Macrocystis exists. Presence of the plants probably attracts herbivorous fishes to the transplant sites and a large amount of grazing is focused on their rather small biomass.
Competitive algae can be a serious impediment to either natural recruitment of juvenile Macrocystis or to reestablishment of this species in an area by transplanting or "seeding" with microscopic stages. Removal of competitors from 1 to 3 m of bottom provides an opportunity for a small transplant to grow to a size where it can overcome competitive effects.
Several kelp diseases have been described (North, 1979b; Scotten, 1971). Most are associated with summertime deterioration of canopies during periods of elevated water temperature and low nutrients. Some localized rotting of stipes near plant bases on Macrocystis holdfasts has been observed in areas of organic enrichment by sewage (North, 1979b).
5.2.3 Possibilities of control measures
Several techniques have been utilized in southern California for controlling overgrazing by sea urchins. Urchins have been destroyed by quicklime, crushed with hammers, and sucked up by dredging machinery (Wilson & McPeak, 1983; Wilson & North, 1983; Leighton et al., 1965). Attempts to control herbivorous fishes by gill-netting, spearing and dispersals of poisons were only partially successful.
As noted above, removal by cutting or plucking of competitive algae from the immediate vicinity of a young Macrocystis (either transplanted or naturally attached) will assist survival until the young plant attains a height of 2 to 3m and begins to eliminate surrounding competitors by shading them. There are no known methods for controlling diseases of Macrocystis plants. Since most of the diseases occur during summer when surface waters are warm and depleted of nutrients, fertilizing might be expected to improve plant healthiness. Research is needed, however, because excessive fertilizing might benefit the disease organisms.
Neushul and coworkers have undertaken breeding studies involving crosses between the three species of Macrocystis, between Macrocystis and closely related kelps such as Pelagophycus and development of clones derived from highly productive adult Macrocystis raised on their test farm at Ellwood, California (Sanbonsuga and Neushul, 1978a, 1978b; Neushul, 1981b; Neushul and Harger, 1985). Work is still in progress and it is not yet established whether enhanced productivity will be possible. Stock cultures of gametophytes of the various species and strains are maintained by this research group. One possibility for enhancing Macrocystis productivity would involve development of a strain which would have tolerance for elevated water temperatures and could be cultivated at low latitudes where wintertime reduction in available sunlight is not as pronounced as in California. Clendenning (1971) reported that Macrocystis from Turtle Bay, Baja California (near the southern limit of the Macrocystis range in North America) exhibited a photosynthetic capacity per unit area that was about 50% higher than the capacity found for plants from California. The temperature optimum for photosynthesis from Turtle Bay kelp was between 25°C and 30°C, about 5°C higher than for kelp from southern California.
Another interesting possibility for improving harvest yield might be utilization of a Macrocystis strain occurring only in Paterson/inlet, New Zealand (Gerard and Kirkman, 1984). This plant can generate several apical meristems on a single frond through production of side branchings along the frond length. Use of this characteristic in a harvested crop might substantially reduce the period required for canopy regeneration after cutting.
Both sexual and vegetative reproduction are important to maintaining the Macrocystis crops available for harvest. Effects of sexual reproduction, however, are indirect and serve only to replace adult plants lost to storm or other adversities. Vegetative growth by young fronds with intact apical blades is entirely responsible for regenerating kelp canopies following harvesting.
Macrocystis populations in southern California are sometimes adversely affected by nutrient starvation. As noted above, depletion of nutrients in surface waters above the thermocline is probably a major factor causing canopy deterioration during summer. Nutrient depletion probably also occurs in association with major El Nino events (a complex sequence of oceanographic events beginning near the equator, spreading to higher latitudes, sometimes lasting from a few months to 2-3 years; Dayton and Tegner, 1984; Gerard, 1984a). Fertilizing Macrocystis canopies prior to deterioration might serve to preserve for harvesting much organic material that might otherwise be lost. Neushul and Harger (1983) observed greater yield during summer from fertilized than from unfertilized plots on their test farm. Fertilizing a kelp bed by crop dusting techniques using helicopters was described by North et al., (1982).
Fertilizing might also be helpful if a Macrocystis crop growing under low-nutrient conditions were to be utilized in preparation of animal feeds. Contents of N and C can be raised substantially in a few days by addition of nitrogen as nitrate or as ammonium to the medium (North, 1978). P-contents of kelp tissues also rise as external concentration of inorganic phosphate is raised (Manley et al., 1982b).
Sizes of Macrocystis beds can be increased by provision of artificial substrate for transplanting (Figure 4). All that is required is an anchoring mechanism. The adult plant does not need to be attached to the bottom (Wilson and North, 1983). Small plants can, however, easily be affixed to solid substrates if so desired (McPeak et al., 1973).
6.1 Annual operations
6.2 Manpower productivity
6.3 Alternate employment
Given appropriate environmental conditions favoring Macrocystis growth, canopies can be available throughout the year. Kelp beds occur in a variety of environments in southern California, so at least some canopy is available somewhere for harvesting at all times of the year. Winter storms and warm waters during summer can create seasonal scarcities of plant material available for harvesting.
Operational and economic statistics from the California kelp harvesting industry are closely guarded and not available to the public. It is believed, however, that manpower requirement for operation of even large harvesting vessels is modest, amounting to four to six individuals. Manpower for unloading and pro cessing the cut kelp is unknown and undoubtedly depends to some extent on intended uses of the kelp and the degree of automation available in the processing factory.
Two economic analyses of kelp harvesting operations and costs were prepared for marine biomass studies undertaken by the U.S. Naval Ocean Systems Center (NOSC) and the Gas Research Institute (GRI). The NOSC study envisioned a Macrocystis farm of about 40,000 ha with an annual productivity of 946 wet metric tons per ha (Integrated Sciences, Inc., 1976). The personnel estimate specified 220 individuals required for harvesting the kelp and 403 shore-based personnel for processing the raw material into the ultimate product (methane gas). These figures indicate that 172,000 t/yr would be produced per individual involved in the harvesting. If we assume that the annual California kelp harvest of about 180,000 t (Table 1) can be obtained by three vessels or 15 individuals, a figure of 12,000 t/man-yr is calculated, considerably below the productivity estimate from NOSC. The discrepancy arises partially from the circumstance that productivity of the wild crop in California at about 35 t/ha/yr, is much lower than that assumed in the NOSC study. The comparison is much closer if it is drawn on the basis of areas harvested, 182 ha/yr/individual for the NOSC study and 333 ha/yr/individual for the California kelp industry.
The GRI-sponsored study envisioned a Macrocystis farm of 16/300 ha yielding an estimated 674 wet t/ha/yr (R.M. Parsons Co., 1983). Their scheme depicted two individuals working 12 hr. shifts in operating the harvesting vessel. This is equivalent to six individuals per day working 8 hr. shifts, giving 1,830,000 t/man-yr. or 2,700 ha/man-yr. These higher figures assume a much greater degree of automation in the harvesting machinery than actually exists in the California kelp harvesting industry.
There is no need for alternate employment in the California kelp harvesting industry, as far as is known.
7.1 Wild resources
7.2 Macrocystis culturing
7.3 Sources of credit and insurance
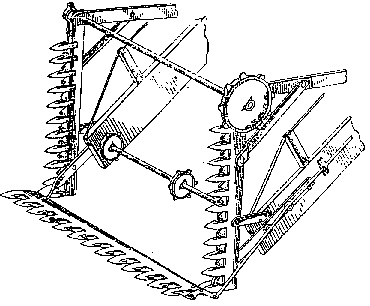
As stated above, detailed operating characteristics and economics of harvesting are not available to the public from the California kelp harvesting industry. The harvesting barges carry the cutting apparatus at the sterns of the vessels and proceed in reverse when operating in a kelp canopy. Cutting is accomplished by reciprocating blades attached to a horizontal bar that is lowered to a fixed depth of 1.2 m below the waterline. Cut ends of the kelp fronds are carried against a conveyor belt just behind the cutting bar. The cut kelp is carried up on the belt over the gunwale, and deposited in an open hold that extends for most of the barge's length. A mechanically driven steel claw or grapnel is used to distribute the cut kelp evenly throughout the hold. The bridge crew's quarters and other necessary spaces are on the bow, forward of the hold. Typical harvesting vessels may load several hundred wet tons of kelp per trip. Cameron (1915) described a harvesting vessel of his day in considerable detail (Figures 5 to 7). The essential features of a kelp harvester, as described above, were fully developed even then.
Harger (1979) studied harvesting activity in Goleta Bay for four years, seeking correlations between amount of kelp obtained and ten environmental variables which he monitored. The most important were surface irradiance, water temperature, swell height, and nitrate concentrations (surface & bottom). He developed a model wherein environmental conditions accounted for 46% of the variability in harvest. Harger found that the harvest rate could vary from about 13 to 110 wet t/hr/with a majority of values lying between 40 and 80 wet t/hr.
Gerard and North (1984) calculated that their simulated harvesting operations removed about 50% of the plant biomass. Macrocystis beds are generally harvested only when dense canopies justify such activity, otherwise a bed is left unharvested until an adequate canopy develops. A bed may thus be left uncut for several years. Productive beds, however, will usually support three harvests per year (C. Martin, personal communication). Brandt (1923) reported that some beds were harvested continuously for five or more months per year, but yields fell significantly following such intense usage.
Capital and maintenance costs of harvesting vessels, docking and unloading facilities, etc., for the California industry are not public information. The NOSC economic analysis budgeted $133,800,000 for start-up harvesting costs with annual operating expenses of $17,400,000 (Integrated Sciences, Inc., 1976). Ralph M. Parsons Co. (1983) listed the cost of a 12x27 m harvesting vessel as $1,677,000 plus $805,000 for a 12x45 m barge to receive the cut kelp. Their operating expenses were $4,830,200 which included all activities, not just harvesting. Thus the NOSC estimates seem extremely high, but the number of harvesting vessels involved was not specified. Also, they planned to operate a kelp farm 160 km offshore, which would increase operating costs considerably since the processing plant was onshore.
The manpower requirement, as stated above appears to be in the range of 200 to 300 ha of kelp bed that can be harvested per man-year using mechanical harvesters as they presently exist. Automation might be able to increase these values.
Presently no commercial-scale culturing of Macrocystis is conducted in California. North (1976, 1972) described experimental facilities for mass-culturing Macrocystis gametophytes and embryonic sporophytes for dispersal in kelp areas to enhance the recruitment processes. Wilson and North (1983) reviewed techniques for transplanting young and adult Macrocystis plants to initiate new populations in depleted areas. McPeak (1975) described a four-year transplantation effort that involved installation of about 28,000 young Macrocystis at the north end of the La Jolla kelp bed.
Figure 7. Perspective view showing details of the cutting apparatus located at the bow of the harvesting vessel of Figure 4 (from Cameron, 1915). Modern harvesters have the cutting and loading equipment at the vessel's stern and the ship moves in reverse while cutting kelp. The prow is V-shaped/providing for maximum efficiency in travelling to and from port.
Industrial loans from commercial banking sources presumably have been available and used by kelp harvesting concerns in California. The major remaining such organization at the present time is Kelco Company of San Diego, a division of Merck & Co., Inc. During periods of crop shortages in California waters, Kelco imports raw material from foreign sources. Dependence on imports, rather than insurance, appears to be the preferred method for sustaining operations in case of natural disasters.
Commercial harvesting of Macrocystis in the USA does not occur north of Monterey Bay in central California. Macrocystis is also harvested in northern Baja California, Mexico, and transported to the processing plant of Kelco.
The kelp harvesting concerns in California have all operated as private entrepreneurs, independent of linkages to governmental agencies. Interest in kelp resources led to formation of several kelp harvesting enterprises in the early 1900's (Scofield, 1959; P. Neushul in Neushul and Harger, 1985). Some of the organizations utilized harvesting techniques that may have damaged the resource and contributed unnecessarily to beach litter. Since all the kelp beds lie within three miles of the shoreline, their ownership resides in the State of California. The responsible State agency for management of living marine resources is the Department of Fish and Game with policy direction from the Fish and Game Commission. Concern about welfare of the kelp resource around 1916 led to a joint effort by the Commission, conserva-tionists, and the larger kelp harvesting concerns, to control harvesting methods and practices. Results were the first set of regulations covering kelp harvesting activities, enacted July 26, 1917. The regulations have been expanded and amended since then on several occasions.
Regulations now divide the California kelp beds into two categories: open beds (available for harvest by anyone) and closed beds (restricted for harvesting by a lessee only). A third of the beds are required to be held in the open category. Harvesters must purchase a license and pay a privilege fee of 10 cents for each wet ton harvested and a second assessment of $100 for each square mile of kelp bed harvested. Since 1950, leases are granted by negotiation (before, leases were awarded on the basis of sealed bids). Leases are granted for periods of fifteen years or less. Lease rights confer only harvesting privileges and have no control over fishing or navigational activities.
In 1956, the Fish and Game Commission decreed that depth of kelp cutting should be no greater than four feet (1.22 m) below the surface and all material cut must be fully utilized (i.e. no escapement of cut fronds is allowed. This latter is to prevent increasing beach litter as a result of harvesting). Violations may result in fines or suspensions of the license to harvest. Enforcement is accomplished by wardens operating from patrol vessels.
9.1 Chemical and nutritional content
9.2 Human Food
9.3 Animal fodder and feed additives
9.4 Manure and fertilizer
9.5 Industrial products and processes
Paine and Vadas (1969) reported the caloric content of M. pyrifera as 2.85 kcal/g dry wt, or 4.3 kcal/g dry minus ash. Similar values were obtained for a closely related kelp, Nereocystis leutkeana. Because of the high ash content, the above caloric values were among the lowest of seventeen seaweed species assayed by these workers. With ash corrections most species were very similar. Effects of various light intensities and nitrate concentrations on caloric value of cultured juvenile Macrocystis sporophytes were determined by Shivji (1985).
Extensive analyses of organic and salt composition of the Pacific coast kelps, including Macrocystis, were reported by Balch and by Turrentine from work in the early 1900's (cf. Cameron, 1912, 1915). The most recent general studies were conducted in connection with the Marine Biomass Program (Tables 2 and 3). As a result of these studies, it became known that concentrations of some of the organic and inorganic constituents vary substantially according to environmental conditions and the recent history of the tissue under analysis. Thus the ranges of mannitol contents of M. pyrifera fronds increased from 5 to 15% dry wt to 14 to 33% dry wt in a plant transferred for three weeks into a low-nutrient environment (Gerardy 1982b). Tissue contents of nitrogen and phosphorus in young M. pyrifera sporophytes reflected concentrations of the inorganic salts of these elements in the culturing media (P.A. Wheeler and North, 1980; Manley et al., 1982). Seasonal variations in dry weight, ash, N, P, S, Cu, Zn, Fe, ether solubles, algin, mannitol, laminarin, and fucoidin were studied for 2 years in M. integrifolia by Wort (1955).
Table 2. Gross composition of Macrocystis, combining data from Point Loma and San Clemente Island (Lindner et al. 1977) and from Monterey Bay, Santa Cruz, and Soquel Point (Hart et al., 1976). Standard deviations are shown for ash, solids, and alginic acid for all samples combined.
|
Substance and % Wet or Dry Wt. |
Percent Composition |
||
|
All Samples |
So. Calif. |
Cen. Calif. |
|
|
% WET WT |
|
|
|
|
Water |
88.2 |
|
|
|
Solids |
11.8 ± 1.0a |
12.2 |
11.3 |
|
% DRY (550°C) |
|
|
|
|
Ash |
38.96 ± 6.24b |
37.29 |
40.49 |
|
KCl |
29 |
|
|
|
NaCl |
7 |
|
|
|
Na2SO4 |
4 |
|
|
|
Other |
4 |
|
|
|
Volatiles |
66-53 |
|
|
|
Mannitol |
6-22 |
|
|
|
Laminarin |
|
1-2 |
|
|
Fucoidan |
|
0.5-2 |
|
|
Alginic Acid |
15.71 ± 2.56c |
18.6 |
11.4 |
|
Cellulosics |
3-8 |
|
|
|
Protein |
5-14 |
|
|
|
Fat |
0.5-2 |
|
|
a 42 samples
b44 samples
c35 samples
Table 3. Concentrations of selected elements in wet Macrocystis tissues (modified from North, 1980) and in seawater according to Brewer (1975). The ratio of these two sets of numbers was taken as the concentration factor for each element. The tissue and seawater values are in micromoles per given unit.
|
Element |
Concentration |
||
|
Tissue per gm wet wt. |
Seawater per liter |
Concentration Factor |
|
|
Phosphorus |
11.6 |
0.2 |
58,000 |
|
Beryllium |
0.025 |
0.00063 |
40,000 |
|
Nitrogen |
10.5 |
0.5 |
21,000 |
|
Antimony |
0.0125 |
0.002 |
6,000 |
|
Iron |
0.175 |
0.05 |
5,000 |
|
Cobalt |
0.00375 |
0.0008 |
4,700 |
|
Arsenic |
0.175 |
0.05 |
3,500 |
|
Manganese |
0.0125 |
0.0036 |
3,500 |
|
Aluminum |
0.120 |
0.074 |
1,600 |
|
Copper |
0.01 |
0.008 |
1,300 |
|
Iodine |
0.588 |
0.5 |
1,200 |
|
Zinc |
0.0688 |
0.076 |
900 |
|
Vanadium |
0.0375 |
0.05 |
750 |
|
Nickel |
0.00625 |
0.028 |
220 |
|
Molybdenm |
0.00125 |
0.01 |
130 |
|
Boron |
3.13 |
410 |
8 |
|
Lithium |
0.19 |
26 |
7 |
|
Strontium |
0.475 |
91 |
5 |
|
Silicon |
0.35 |
71 |
5 |
|
Potassium |
15 |
10,200 |
1.5 |
|
Uncertain |
|||
|
Selenium |
0.00125 |
0.0025 |
500 |
|
Chromium |
0.00125 |
0.0057 |
220 |
Bromine, Rubidium, and Fluorine are not concentrated.
Usage of Macrocystis tissues as human food is minor in North America. Macrocystis and Nereocystis slices are occasionally prepared as pickles by individuals, but there is no known commercial product of this kind. Dried powdered kelp (probably M. pyrifera) was formerly marketed by Philip R. Park, Inc. A fraction of the product entered specialty items in health food stores (P. Neushul in Neushul and Harger, 1985).
Dried kelp (probably Macrocystis pyrifera) was formerly produced by Philip R. Park Inc. and Kelp Organic Products Co. of Los Angeles. Most of the material was incorporated into animal feeds. Neither of these concerns exist at this time (P. Neushul, 1.c.). The value of this Macrocystis as fodder might be enhanced by fertilizing shortly before harvesting to increase nitrogen and phosphorus contents of the tissues via (cf. Mateus et al., 1976) its amino acid composition.
No present usage of Macrocystis as a manure exists in California. A large and successful effort was mounted during the first World War to secure kelp as a source of potash for manufacture of fertilizer and explosives (Cameron, 1915; Scofield, 1959; P. Neushul, 1.c.). Presently, all Macrocystis harvested in California and Baja California is processed for alginate production and the resource is fully utilized. An economic study of the feasibility of using Macrocystis for methane production proposed dispersal of sludge remaining from digestion of the kelp, into their kelp farm as a fertilizer (Ralph M. Parsons, 1983).
As noted above, alginates are the only products currently manufactured from harvested Macrocystis in California. Various compounds of the complex polysaccharides, alginic acid, serve as stabilizers, thickeners, emulsifiers, water-holders, and film-forming or suspending agents. Most of the usages are in foods, pharmaceuticals, beverages, dairy products, and in the manufacture of latex, textiles, as well as paper and cardboard, plus other applications (Kelco, 1961; P. Neushul, 1.c.). Two methods for alginate manufacture are the Le Gloahec-Herter and Green's cold processes, both described with flow charts by Chapman (1970). Salts are first leached from cut or slurried kelp in weak acid. The alginate is then solubilized or induced to form a gel by soaking the kelp in a solution of sodium carbonate or other weak alkali. Reduction of pH by dilution with water or addition of acid causes precipitation and the recovered material can be sold as crude alginate or refined further by repeated solubilization and precipitation (Tseng, 1945).
In the past, a number of other industrial products have been derived from Macrocystis in California, particularly during the period of intense utilization during World War I. These included potash and acetone for explosives manufacturing (potash was also employed in manufacture of matches), iodine, and char (P. Neushul, 1.c.). The early kelp industry was unable to compete with German sources of potash which once more became available to American industry after termination of the war in 1919. The California kelp industry collapsed for about a decade until interests in alginate production and the uses of kelp in animal feeds revived harvesting activities in 1927-28.
A possible future usage of Macrocystis is as alternative sources of energy in the form of gaseous or liquid fuels. The feasibility of liquid fuel production has already been demonstrated on a commercial scale by production of acetone in World War I by Hercules Powder Company (P. Neushul, 1.c.). Extensive investigations of the potential for methane production from kelp were recently completed (Tompkins and Bryce, 1984). This study, the Marine Biomass Program, also yielded considerable information relating to Macrocystis biology, and many of the resulting publications were cited above. We also referred to two of the economic analyses that were performed (Integrated Sciences, Inc., 1976; Ralph M. Parsons, 1983). Macrocystis tissue proved to be an excellent substrate for methane production by bacterial digestion processes. Yields of 4.5 SCF/1bVS (0.28 liters of combustible gas per gram of volatile solids) were reported by Chenoweth et al. (1978). Maximal methane yields were recovered from those kelp samples highest in mannitol.
In closing, we should mention that the Macrocystis forests in nature provide an extensive habitat for fishes and shellfish utilized by humans (North and Hubbs, 1968; Leighton, 1971). As such, Macrocystis and other large kelps are an important sustaining element in recreational and commercial fisheries, both of which are well-established activities in California.
Anderson, L.M., 1984 Iron reduction by juvenile Macrocystis pyrifera (L.) C. Agardh. Hydrobiologia,116/117:493-97
Anderson, E.K., and W.J. North, 1966 In situ studies of spore production and dispersal in the giant kelp, Macrocystis. Proc.Int.Seaweed Symp., 5:73-86
Arnold, K.E. and S.L. Manley, 1985 Carbon allocation in Macrocystis pyrifera (Phaeophyta): intrinsic variability in photosynthesis and respiration. J. Phycol., 21:154-67
Bernstein, B., 1977 Selective pressures and coevolution in a kelp canopy community in southern California. Ph.D. Thesis. University of California, San Diego, 135 p.
Bernstein, B., and N. Jung, 1979 Selective pressures and coevolution in a kelp canopy community in southern California. Ecol.Monogr., 49:335-55
Brandt, R.P., 1923 Potash from kelp: early development and growth of the giant kelp, Macrocystis pyrifera. Bull.U.S. Dept-Agric., (1191):40 p.
Brewer, G., 1975 Minor elements in seawater. In Chemical oceanography, edited by J.P. Riley and G. Skirrow. London, Academic Press, Vol.1:415-96 2nd ed.
Buggeln, R.G., D.S. Fensom, and C.J. Emerson, 1985 Translocation of 14C-photoassimilates in the blade of Macrocystis pyrifera (Phaeophyceae). J. Phycol., 21:35-40
Cameron, F.K., 1912 A preliminary report on the fertilizer resources of the United States (with special reference to seaweeds). Washington, D.C., U.S. Senate Doc. (100): 40-290
Cameron, F.K, 1915 Potash from kelp. I. Pacific kelp as a source of potassium salts. Rep.U.S.Dep.Agric., (100):9-32
Chapman, V.J., 1970 Seaweeds and their uses. London, Methuen and Co. Ltd., 304 p. 2nd ed.
Chenoweth, D.P., L. Klass, and S. Ghosh, 1978 Biomethanation of giant brown kelp Macrocystis pyrifera. In Energy from biomass and wastes. Chicago Institute of Gas Technology, pp. 229-51
Clendenning, K.A., 1963 Photosynthesis and growth in Macrocystis pyrifera. Proc.Int.Seaweed Symp., 4:55-65
Clendenning, K.A, 1971 Photosynthesis and general development in Macrocystis. In The biology of Giant Kelp beds (Macrocystis) in California, edited by W.J. North. Lehre, Germany, J. Cramer, pp. 169-90
Cole, K., 1968 Gametophytic development and fertilization in Macrocystis integrifolia. Can.J.Bot., 46:777-82
Dayton, P.K., B.D. Keller, and D.A. Ven Tresca, 1980 Studies of a nearshore community inhabited by sea otters. Report to U.S. Marine Mammal Commission. Washington D.C., Rep.No.(MMC-78/14):91 p. Available from National Technical Information Service, Springfield, Virginia, PB81-109860)
Dayton, P.K., and M.J. Tegner, 1984 Catastrophic storms and patch stability in a southern California kelp community. Science, Mash., 224:283-5
Dayton, P.K., et al., 1984 Patch dynamics and stability of some California kelp communities. Eco L. Monogr., 54:253-89
Dean, T.A., and L.E. Deysher, 1983 The effects of suspended solids and thermal discharges on kelp. In The effects of waste disposal on kelp communities edited by W. Bascom, Long Beach, Southern California Coastal Water Research Project, pp. 114-35
Dean, T.A., S.C. Schroeter, and J. Dixon, 1984 Effects of grazing by two species of sea urchins (Strongylocentrotus franciscanus and Lytechinus anamesus) on recruitment and survival of two species of kelp (Macrocystis pyrifera and Pterygophora californica). Mar.Biol., 78:301-13
Devinny, J.S., and L.A. Volse, 1978 Effects of sediments on the development of Macrocystis pyrifera gametophytes. Mar.Biol., 48:343-8
Deysher, L.E., 1984 Recruitment processes in benthic marine algae. Ph.D. Thesis, University of California, San Diego, 324 p.
Dixon, J., S.C. Schroeter, and J. Kastendiek, 1981 Effects of the encrusting bryozoan, Membranipora membranacea, on the loss of blades and fronds by the giant kelp, Macrocystis pyrifera (Laminariales). J.Phycol., 17:341-5
Druehl, L.D., 1979 The distribution of Macrocystis integrifolia in British Columbia as related to environmental parameters. Can.J.Bot., 56:69-59
Ebeling, A.W., D.R. Lauer, and R.J. Rowley, 1984 Severe storm disturbances and reversal of community structure in a southern California kelp forest. Mar.Biol., 75:1-8
Estes, J.A., and J.F. Palmisano, 1974 Sea otters: their role in structuring nearshore communities. Science, Wash., 185:1058-60
Fain, S.R., and S.N. Murray, 1982 Effects of Light and temperature on net photosynthesis and dark respiration of gametophytes and embryonic sporophytes of Macrocystis pyrifera. J.Phycol., 18:92-8
Foster, M.S., 1975 Algal succession in a Macrocystis pyrifera forest. Mar.Biol., 32:313-29
Foster, M.S, 1975a Regulation of algal community development in a Macrocystis pyrifera forest. Mar.Biol., 32:331-42
Gerard, V.A., 1976 Some aspects of material, dynamics and energy flow in a kelp forest in Monterey Bay, California. Ph.D. Thesis, University of California Santa Cruz, 173 p.
Gerard, V.A, 1982 In situ rates of nitrate uptake by giant kelp, Macrocystis pyrifera (L.) C. Agardh: tissue differences, environmental effects, and predictions of nitrogen-limited growth. J.Exp.Mar.BioL.Ecol., 62:211-24
Gerard, V.A, 1982a Growth and utilization of internal nitrogen reserves by the giant kelp Macrocystis pyrifera in a Low-nitrogen environment. Mar. Biol., 66:27-35
Gerard, V.A., 1982b In situ water motion and nutrient uptake by the giant kelp Macrocystis pyrifera. Mar.Biol., 69:51-4
Gerard, V.A, 1984 Physiological effects of EL Nino on giant kelp in southern California. Mar.Biol. Lett., 5:317-22
Gerard, V.A, 1984a The Light environment in a giant kelp forest: influence of Macrocystis pyrifera on spatial and temporal variability. Mar.Biol., 84:189-95
Gerard, V.A., and H. Kirkman, 1984 Ecological observations on a branched, Loose-Lying form of Macrpcystis pyrifera (L.) C. Agardh in New Zealand. Bot.Mar., 27:105-9
Gerard, V.A., and J. North, 1984 Measuring growth, production, and yield of the giant kelp, Macrocystis pyrifera. Hydrobiologia, 116/117:321-4
Grua, P., 1964 Premières donnèes sur le biomasses de herbier a Macrocystis pyrifera de la baie du Morbihan (Archi-pel Kerguelen). Terre Vie, 2:215-20
Haines, K.C., and P.A. wheeler, 1978 Ammonium and nitrate uptake by the marine macrophytes Hypnea musciformis (Rhodophyta) and Macrocystis pyrifera (Phaeophyta). J.Phycol., 14: 319-24
Harger, B.W.W., 1979 Coastal oceanography and hard substrate ecology in a California kelp bed. Ph.D. Thesis, University of California, Santa Barbara, 437 p.
Harger, B. W., 1983 A historical overview of kelp in southern California. In The effects of waste disposal on kelp communities, edited by W. Bascom. Long Beach Southern California Coastal water Research Project, pp. 70-83
Harris, G., et al., 1984 Community recovery after storm damage: a case of facilitation in primary succession. Science, Wash., 224:1336-8
Hart, M.R., et al., 1976 Ocean food and energy farm kelp pretreatment and separation processes. Berkeley, California, Western Regional Research Center, U.S. Department of Agriculture Report for Agreement No. 12-14-5001-6402:125 p.
Integrated Sciences Corp., 1976 Systems analysis of the ocean food and energy farm (DFEF). Vol. 1, overall economic analysis of the base line design. Santa Monica, California, Integrated Sciences Corporation, 69 p.
Jackson, G.A., 1976 Nutrients and productivity of the giant kelp, Macrocystis pyrifera in the nearshore. Ph.D. Thesis, California Institute of Technology, Pasadena, 134 p.
Jackson, G.A, 1977 Nutrients and production of giant kelp, Macrocystis pyrifera, off southern California. Limnol.Oceanogr., 22:979-95
Jackson, G.A., D.E. James, and W.J. 1985 North submitted, Morphological relationships among fronds of giant kelp, Macrocystis pyrifera, off La Jolla, California, Mar.Ecol.(Prog. Ser.), 26(3):261-70
Jackson, G.A., and C.D. Winant, 1983 Effect of a kelp forest on coastal currents. Cont.Shelf Res., 2:75-80
Kain, J.M., 1982 Morphology and growth of the giant kelp Macrocystis pyrifera in New Zealand and California. Mar.Biol., 67:143-57
Kelco Company, 1961Algin. San Diego, Kelco Co., 12 p.
Kimura, S., 1980 The effects of harvesting Macrocystis pyrifera on understory algae in Carmel Bay, California. M.S. Thesis, California State University, Fresno, 108 p.
Kirkwood, P.D., 1977 Seasonal patterns in the growth of the giant kelp, Macrocystis pyrifera. Ph.D. Thesis, California Institute of Technology, Pasadena, 139 p.
Kuwabara, J.S., 1982 Micronutrients and kelp cultures: evidence for cobalt and manganese deficiency in southern California deep seawater. Science, wash., 216:1219-21
Kuwabara, J.S., and W.J. North, 1980 Culturing microscopic stages of Macrocystis pyrifera (Phaeophyta) in Aquil, a chemically defined medium. J.Phycol., 16:546-9
Leighton, D.L., 1971 Grazing activities of benthic invertebrates in kelp beds. In The biology of giant kelp becs (Macrocystis) in California, edited by W.J. North. Lehre, Germany, J. Cramer, pp. 421-53
Leighton, D.L., G. Jones and J. North, 1966 Ecological relationships between giant kelp and sea urchins in southern California. Proc.Int.Seaweed Symp., 5:141-53
Lindner, E., C.A. Dooley, and R.H. Wade, 1977 Chemical variation of chemical constituents in Macrocystis pyrifera. Final report. San Diego, U.S. Naval Ocean Systems Center, 39 p.
Liu, Tianjing, et al., 1984 Studies on the artifical cultivation and propagation of giant kelp (Macrocystis pyrifera). Hydrobiologia, 116/117:259-62
Lobban, C.S., 1978 Translocation of 14C in Macrocystis pyrifera (giant kelp). Plant Physiol., 61:585-9
Lobban, C.S, 1978 Translocation of 14C in Macrocystis integrifolia (Phaeophyceae). J.Phycol., 14:178-82
Luning, K., and M. Neushul, 1978 Light and temperature demands for growth and reproduction of Laminarian gametophytes in southern and central California. Mar. Biol., 45:297-309
Manley, S.L., 1981 Iron uptake and translocation by Macrocystis pyrifera. Plant Physiol., 68:914-8
Manley, S.L, 1984 Micronutrient uptake and translocation by Macrocystis pyrifera (Phaeophyta). J.Phycol., 20:199-201
Manley, S.L, Phosphate uptake by blades of Macrocystis pyrifera (Phaeophyta). Sot.Mar. (in press)
Manley, S.L., and W.J. North, 1981 micronutrient uptake by juvenile Macrocystis pyrifera (L.) C. Agardh. Proc.Int.Seaweed Symp., 10:467-72
Manley, S.L., and W.J. North, 1984 Phosphorus and the growth of juvenile Macrocystis pyrif ra (Phaeophyta) sporophytes. J. Phycol., 20:389-93
Manley, S.L. et al., 1981 Environmental considerations involved in 1982 farming Macrocystis. In Proceedings of the International Gas Research Conference, Rockville, Maryland, Government Institutions Inc., pp. 693-8
Mateus, H., J.M. Regenstein, and R.C. Baker, 1976 The amino acid composition of the marine brown alga Macrocystis py r i fe ra from Baja California. Mexico. Bot. Mar., 19:155-9
McPeak, H., 1975 Observations and transplantation studies at Point Loma and La Jolla. Annu.Rep.KeIp Habitat Improv. Proj.Calif.Inst.Technol. Pasadena, (1974/5):80-91
McPeak, R.H., H.C. Fastenau, and D. Bishop, 1973 observations and transplantation studies at La Jolla and Point Loma. Annu.Rep.Kelp Habitat Improv.Proj.Calif.Inst.Technol., Pasadena, (1972/3): 57-73
Neushul, M., 1959 Studies on the growth and reproduction of the giant kelp, Macrocystis. Ph.D. Thesis, University of California, Los Angeles, 134 p.
Neushul, M, 1971 The species of Macrocystis with particular reference to those of North and south America. In The biology of Giant Kelp beds (Macrocystis) in California, edited by W.J. North. Lehre, Germany, J. Cramer, pp. 211-22
Neushul, M., 1981 Historical review of kelp beds in the southern California Bight. Goleta, California, Neushul Mariculture Inc., 74 p.
Neushul, M, 1981 The domestication and cultivation of Californian amacroalgae. Proc. Int..Seaweed Symp., 10:71-96
Neushul, M. and B.W.W. Harger, 1983 Kelp biomass coastal test farm program. Annual report for 1982. Goleta, California, Neushul Mariculture Inc., 25 p. + 75 p. Appendices.
Neushul, M. and B.W.W. Harger, 1985 Kelp biomass production yield, genetics, and planting technology. Annual report Jan. 1983-Aug. 1984. Goleta, California, Neushul Mariculture Inc., 85 p. + 39 p. Appendices
Neushul, M., B.W.W. Harger, and J.W. Woessner, 1982 Laboratory and nearshore field studies of the giant California kelp as an energy crop plant. In Proceedings of the 1981 International Gas Research Conference, Rockville, Maryland, Government Institutions Inc., pp. 699-708
North, W.J., 1958 Experimental ecology. Kelp Investigations Program, Quarterly progress report, July 1 - Sept. 30. La Jolla, University of California, Institute of Marine Research, pp. 2-8
North, W.J, 1968 Effects of canopy cutting on kelp growth: comparison of experimentation with theory. In Utilization of kelp-bed resources in Southern California, edited by W.J. North and C.L. Hubbs. Fish Bull.Calif.Dep.Fish Game, (139):223-54
North, W.J, 1971 Growth of individual fronds of the mature giant kelp. Macrocystis. In The biology of Giant kelp beds (Macrocystis) in California, edited by W.J. North. Lehre, Germany, J. Cramer, pp. 123-68
North, W.J, 1972 Mass-cultured Macrocystis as a means of increasing kelp stands in nature. Proc.Int. Seaweed Symp., 7:394-99
North, W.J,1976 Aquacultural techniques for creating and restoring beds of giant kelp, Macrocystis spp. J.Fish.Res.Board Can, 33(4)Pt2:1015- 23-
North, W.J, 1978 Progress in studies of oceanic production of biomass. In Proceedings of the Second Annual Symposium on Fuels from biomass, edited by W.W. Shuster. Washington, D.C., U.S. Department of Energy.(CONF-7806107-P2):99-1040. Available from National Technical Information Service, Springfield, Virginia.
North, W.J, 1979 Adverse factors affecting giant kelp and associated seaweeds. Experientia, 35:445-7
North, W.J., 1980 Trace metals in giant kelp. Macrocystis. Am.J.Bot., 67:1097-101
North, W.J, 1983 Separating the effects of temperature and nutrients. In The effects of waste disposal on kelp communities, edited by W. Bascom. Long Beach, Southern California Coastal Water Research Project, pp. 243-55
North, W.J., V.A. Gerard, and J.S. Kuwabara, 1982 Farming Macrocystis at coastal and oceanic sites. In Proceedings of the Conference on Synthetic and degradative processes in marine macrophytes, edited by L.M. Srivastava. New York, de Gruyter, pp. 247-62
North, W.J., and C.L. Hubbs, (eds). 1968 Utilization of kelp-bed resources in Southern California. Fish Bull.Calif. Dep.Fish Game, (139):264 p.
North, W.J., and R.C. Zimmerman, 1984 Influences of macronutrients and water temperatures on summertime survival of Macrocystis canopies. Hydrobiologia, 116/117:419-24
Paine, R.T., and R.L. Vadas, 1969 Calorific values of benthic marine algae and their postulated relationship to invertebrate food preferences. Mar.Biol., 4:79-86
Parsons Company, Ralph M., 1983 An economic and systems assessment of the concept of nearshore kelp farming for methane production. Final report. Chicago, Gas Research Institute (GRI-82/0067):86 p + 234 p Appendices
Pinkas, L., 1977 California marine fish landings for 1975. Fish.Bull. Calif.Dep.Fish.Game, (168):5 5 p.
Rosenthal, J., D.Clarke, and P.K. Dayton, 1974 Observations on the ecology and natural history of a stand of giant kelp, Macrocystis pyrifera (Linnaeus) Agardh, off Del Mar, California. Fish.Bull.NOAA/NMFS, 72:670-84
Ryther, J.H., et al., 1977 Cultivation of seaweeds as a biomass source for energy. In Proceedings Fuel from biomass Symposium. Urbana, University of Illinois, pp. 83-98
Ryther, J.H., et al., 1979 Biomass production by marine and freshwater plants. In Proceedings of the Third Annual Biomass Energy Systems Conference. Golden, Connecticut, SERI, (SERI/TP-33-285):13-23. Available from National Technical Information Service, Springfield, Virginia.
Schmitz, K., 1982 Translocation of organic compounds in Lam Laminariales. In Synthetic and degradative processes in marine macrophytes, edited by L.M. Srivastava, de Gruyter, pp. 16-179
Schmitz, K. and L.M. Srivastava, 1979 Long distance transport in Macrocystis integrifolia. 2. Tracer experiments with 14C and 32P. Plant Physiol., 63:1003-9
Scofield, W.L., 1959 History of kelp harvesting in California. Calif. Fish Game Q., 45:135-57
Scotten, H.L., 1971 Microbiological aspects of the kelp bed environment. In The biology of Giant Kelp beds (Macrocystis) in California, edited by W.J. North. Lehre, Germany, J. Cramer, pp. 315-8
Shivji, M., 1985 Interactive effects of Light and nitrogen on growth and chemical composition of juvenile Macrocystis pyrifera (L.) C.Ag. (Phaeophyta) sporphytes. J. E x p. Mar.Biol-Ecol., 89(1):81-96
Smith, R.G., N. Wheeler, and L.M. Srivastava, 1983 Seasonal photo-synthetic performance of Macrocystis integrifolia (Phaeophyceae). J.Phycol., 19:352-9
Tegner, M.J., and L.A. Levin, 1983 Spiny Lobsters and sea urchins: analysis of a predator-prey interaction. J.Exp.Mar. Biol.Ecol., 73:125-50
Tompkins, A.N., and A.J. Bryce, 1984 Marine biomass program, Final report. Chicago, Gas Research Institute, 166 p.
Tseng, Cheng K., 1945Colloids from kelp. Chem-Metall.Eng., 52:97-100
Vaughn, O.W., 1959 Carbohydrate metabolism in a marine brown alga Macrocystis pyrifera. Ph.D. Thesis, University of California, Berkeley, 65 p.
Wheeler, A., 1979 Uptake of methylamine (an ammonium analogue) by a marine macrophyte Macrocystis pyrifera (Phaeophyta). J.Phycol., 15:12-7
Wheeler, P.A., and W.J. North, 1980 Effect of nitrogen supply on nitrogen content and growth rate of juvenile Macrocystis pyrifera (Phaeophyta) sporophytes. J.Phycol., 16:577-82
Wheeler, W.N., 1978 Ecophysiological studies on the giant kelp, Macrocystis. Ph.D. Thesis, University of California, SantaBarbara, 179 p.
Wheeler, W.N, 1980 Effect of boundary Layer transport on the fixation of carbon by the giant kelp Macrocystis pyrifera. Mar.Biol., 56:103-10
Wheeler, W.N, 1980a Pigment content and photosynthetic rate of the fronds of Macrocystis pyrifera. Mar.Biol., 56:97-102
Wheeler, W.N, 1982 Nitrogen nutrition of Macrocystis. In Synthetic and degradative processes in marine macrophytes, edited by L.M. Srivastava. New York, de Gruyter, pp. 121-35
Wheeler, W.N., and L.M. Srivastava, 1984 Seasonal nitrate physiology of Macrocystis integrifolia Bory. J.Exp.Mar.Biol. Ecol., 76:35-50
Wilson, K.C., and R. H. McPeak, 1983 Kelp restoration. In The effects of waste disposal on. kelp communities, edited by W. Bascom. Long Beach, Southern California Coastal Water Research Project, pp. 199-216
Wilson, K.C., and W.J. North, 1983 A review of kelp bed management in southern California. J.WorId Maricult.Soc., 14:347-59
Wing, B.L., and K.A. Clendenning, 1971 Kelp surfaces and associated invertebrates. In The biology of Giant Kelp beds (Macrocystis) in California, edited by W.J. North. Lehre, Germany, J. Cramer, pp. 319-41
Womersley, H.B.S., 1954 The species of Macrocystis with special reference to those on southern Australian coasts. Univ.Calif.Publ.Bot., 27:109-32
Wort, D.J., 1955 The seasonal variation in chemical composition of Macrocystis integrifolia and Nereocystis leutkeana in British Columbia coastal waters. Can.J. Bot., 33, pp. 323-40
Zimmerman, R.C., 1983 Seasonal patterns in the productivity of a giant kelp (Macrocystis pyrifera) forest: the effect of nutrient availability. Ph.D. Thesis, University of Southern California Los Angeles, 182 p.
Zimmerman, R.C., and J.N. Kremer, 1984 Episodic nutirent supply to a kelp forest ecosystem in southern California. J.Mar. Res., 42:591-604
![]()
![]()
{kind=link}
{kind=link}
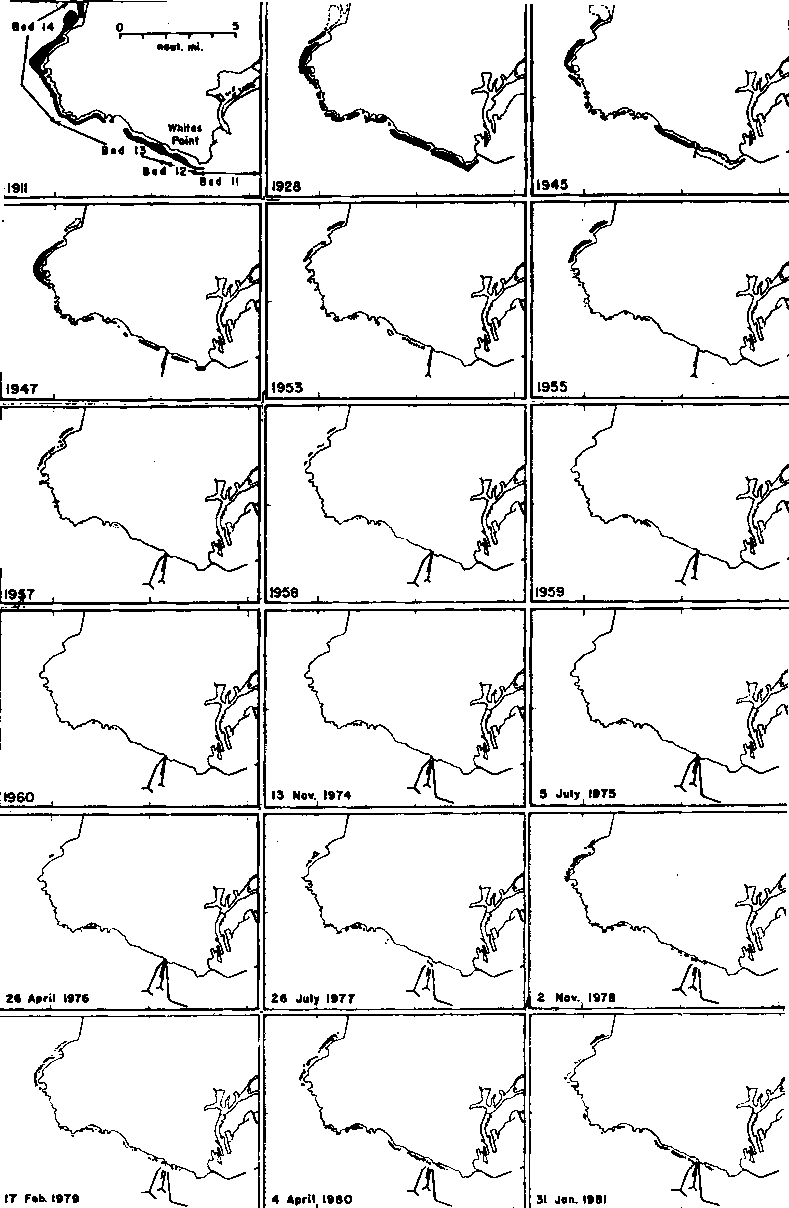{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}