![]()
![]()
![]()
J.C. ELGUE y G. FABIANO
Instituto Nacional de Pesca
Montevideo, Uruguay
R. DELFINO
Instituto Nacional de Investigación y Desarrollo Pesquero
Mar del Plata, Argentina
2.1 RESUMEN
Durante 1984 se realizaronn 46 estaciones de pesca con redes agalleras en el Embalse de Salto Grande (31 °S; 58 °W), ubicadas en 7 brazos: Itapebí, Gualeguaycito, Mandisovi, Arapey, Mocoretá, Yacui y Mandiyú, y una estación, Campamento, correspondiente al antiguo lecho del cauce del río Uruguay, cercana a la represa.
Se calcularon los índices de diversidad específica (H'), sobre la base de un inventario que comprendía la presencia de 49 taxones. El máximo de H' registrado correspondió a la estación Campamento en invierno, con 3.911 bits individuo; el mínimo fue registrado en el brazo Mandiyú, con 0,978 bits/individuo. El promedio para todo el período considerado es de 2.677 bits/individuo.
Se practicó un análisis en componentes principales con los valores de H' calculados, utilizándose éstos como descriptores, y se obtuvo una ordenación en la cual los dos primeros ejes explican el 96% de la varianza total con una buena discriminación de cada estación anual Se reafirmó el comportamiento particular de cada ambiente estudiado, lo cual concuerda con otros estudios realizados en el área.
ABSTRACT
During 1984. 46 stations utilizing gill nets were carried out in Salto Grande Reservoir (31 °S; 58° W). The stations were distributed along seven boundaries of the Uruguay river: Itapebí, Gualeguaycito, Mandisoví, Arapey, Mocoretá, Yacuí and Mandiyú. Another station was conducted at the old ditch of the Uruguay river, close to the dam, in the area known as Campamento.
The specific diversity index (H') was calculated for an inventory of 49 fish taxa. The maximum of H' was recorded in winter at Campamento, with the value of 3,911 bits/individual. The minimum was recorded at Mandiyú, with a value of 0.978 bits/ individual. The average for the period considered was 2,677 bits/individual.
An analysis of principal components was carried out with the H' calculated values. The two first axes explain about the 96% of the total variance, giving a good discrimination of each annual station. It is reaffirmed the particular behaviour of each studied environment, wich corresponds to other researches carried out in the above mentioned area.
2.2 INTRODUCCION
Se ha establecido que una característica de los ríos sudamericanos es poseer una fauna íctica extremadamente rica y diversificada, en comparación, por ejemplo, con los ríos europeos (Margalef, 1983). Por una parte el distinto origen y por otra un regímenes histórico más antiguo, están en la base de la riqueza específica antes indicada.
La construcción de un embalse es una fuente de perturbación para el ecosistema original, perturbación que es posible estudiar desde diversos puntos de vista. El que nos ocupa en esta oportunidad es el relacionado con las variaciones espaciales y temporales de ciertos recursos ícticos en el área del Embalse de Salto Grande durante el año 1984.
Desde la construcción del embalse en 1960, la Comisión Técnica Mixta de Salto Grande (CIMSG), ha emprendido una serie de relevamientos tendientes a mejorar el conocimiento sobre este ambiente. Dado que se trata de un ecosistema de funcionamiento forzado, el embalse está predispuesto a sufrir alternaciones importantes. La práctica eventual o sistemática de la pesca y la influencia de dichas perturbaciones, han llevado a la CIMSG a poner en práctica un programa de monitoreo de las poblaciones de peces del embalse
Trataremos de desarrollar ciertos puntos de vista que se relacionan con el concepto de diversidad específica, tomando datos que fueron recolectados con otro objetivo y dejando en claro que los registros que se poseen anteriores a la construcción del embalse, son muy pocos y fragmentarios
La diversidad especifica (H') podría definirse en forma general como una medida de la composición en especies de un ecosistema, explicada en términos del número de especies y de sus abundancias relativas (Legendre y Legendre, 1984). Dicho índice de diversidad es aplicable a inventarios de cualquier tipo de comunidades, cualesquiera sea la forma de distribución de los taxones participantes (Margalef, 1974). Pielou (1977) describe este índice en cuanto a la variable especie, como la varianza que se refiere a una variable métrica
Las medidas de diversidad obtenidas a partir de la teoría de la información propuesta por Shannon y Weaver (1963) son razonablemente independientes de los valores obtenidos en cada muestreo para las especies consideradas y sus respectivas abundancias (Vieira da Silva, 1979).
El rango de variación de H', cuando se usan logaritmos de base 2, queda acotado entre 0 y algo más de 5 bits; a modo de ejemplo, valdrá 0 cuando todos los individuos considerados pertenezcan a la misma especie, obteniéndose el valor máximo cuando cada especie presente esté representada por un solo individuo. Este parece ser un buen indicador del estado del ecosistema, y las variaciones registradas para un mismo lugar en dos tiempos diferentes nos puede dar una idea con valores comparables de los cambios ocurridos. Margalef (1974; 1980) presenta una excelente revisión del tema, ofreciéndose asimismo valores de diversidad obtenidos en distintos ecosistemas y su posible significación ecológica.
Por otra parte, se debe dejar constancia de que es posible criticar las conclusiones que aquí se exponen, a partir del sesgo que el arte empleado tiene. Un examen primario de los taxones presentes y de su numerosidad, nos obliga a realizar un comentario más: de las especies de las cuales se tiene conocimiento, presentes en el área, no todas aparecen, y cuando lo hacen, su numerosidad es relativamente baja. La elección de un arte pasivo parece ser una de las causas de ello, especialmente para aquellas especies que por sus características biológicas (hábitos alimentarios y reproductivos, de desplazamiento, etc.), no son posibles de ser capturadas. Provoca, asimismo, una pobre representación de individuos de tallas pequeñas próximas al límite inferior del rango de malla utilizado.
Evidentemente, no existe un arte de pesca ideal y la mejor solución sería utilizar artes pasivos y activos combinados, solución ésta que conlleva otros problemas. De cualquier forma, no se deben considerar los resultados aquí expuestos como definitivos, sino más bien como una aproximación más o menos gruesa, que fue posible efectuar dadas las posibilidades de trabajo y el objetivo inicial de monitorear las eventuales modificaciones de la fauna íctica, luego de la construcción de la represa.
2.3 MATERIALES Y METODO
Los inventarios utilizados en el presente trabajo fueron obtenidos mediante la pesca en estaciones fijas de muestreo, considerándose que el embalse es un ambiente heterogéneo y posible de ser estudiado por subambientes (Prenski y Baigún, 1984). La caracterización limnológica del embalse muestra que existen diferencias significativas entre el centro y los brazos, y de estos entre sí (Quirós, 1980; Quirós y Cuch, 1982).
Se establecen de esta manera cinco estaciones de muestreo en el centro de los brazos: Gualeguaycito, Itapebi, Mandisovi, Arapey y Mocoretá, y una en el cauce del antiguo río: Campamento. En las campañas correspondientes a otoño, invierno y primavera se agregan dos estaciones, correspondientes a los brazos del Yacuí y Mandiyú (Figura 1).
El muestreo correspondiente al verano se realizó entre el 1 de febrero y el 3 de marzo; el de otoño, del 16 de mayo al 1 de junio; el de invierno, del 14 al 30 de agosto; y el de primavera, entre el 30 de noviembre y el 15 de diciembre. La pesca de tipo pasivo se efectuó mediante la utilización de una batería de nueve redes agalleras, las que se ubicaron en forma perpendicular a la costa y con los tamaños de malla crecientes hacia el centro del brazo. El tamaño de malla para la batería queda comprendido entre 42 y 170 mm y un detalle de las características de esta batería de redes se describe en Chediak et al (1986). Asimismo se define la estrategia de muestreo abarcando dos jornadas sucesivas, recogiéndose las capturas en la primera mañana siguiente, para evitar el efecto de saturación en el arte utilizado.
El total de la captura de la batería de redes fue identificado y contado, contabilizándose en conjunto e identificándose 49 taxones, los cuales aparecen en la Tabla I. Se calcula el índice de diversidad específica (H') de acuerdo con el propuesto por Shannon-Weaver (1963), donde:
H' = - Σni= 1 Pi log2 P1
donde: P1 = 1, y P1 = N1/N
Una vez obtenidos los valores de H' para el conjunto de los datos, pareció oportuno explorar primariamente si los valores de diversidad específica obtenidos en cada estación de muestreo para las diferentes estaciones anuales consideradas pudieran tener un comportamiento coherente. En efecto, se ensaya un análisis en componentes principales considerados a partir de la matriz de correlación, utilizándose como fuente los datos de H' de aquellas estaciones que fueron muestreadas a lo largo del año: Campamento, Itapebi, Gualeguaycito, Mandisoví, Arapey y Mocoretá. Para tal fin se utiliza la rutina PCAR tomada de Orloci (1975) y desarrollada en un computador de mesa tipo PC.
2.4 RESULTADOS Y DISCUSION
Los valores de H' para cada estación de muestreo aparecen en la Tabla II. El valor máximo para el período examinado corresponde a la estación de Campamento, en el invierno, con 3.911 bits/individuo; el mínimo a la estación de Mandiyú, en otoño, con 0.978 bits/individuo. Estacionalmente, es durante el invierno cuando se registran los valores más altos en promedio, con una media de 3.128 bits/individuo; los mínimos corresponden al otoño, con una diversidad media de 2.256 bits/individuo.
En relación con los ambientes estudiados, los valores medios de diversidad en Campamento, Mocoretá, Mandisoví e Itapebí son parecidos: entre 2.841 y 2.994 bits/individuo. El brazo Mandiyú, sobre el margen uruguayo y con sólo tres muestreos, presenta el valor medio mínimo, con 1.786 bits/individuo. El invierno es la estación del año que presenta la menor variación en el índice; las mayores diferencias entre ambientes se registran durante otoño y primavera (Figura 2). La distribución especial de los valores de H' para cada estación anual se ilustran en las Figuras 3 a 6.
En el verano de 1984, la distribución de la diversidad parece seguir el sentido este-oeste del embalse; del lado argentino, los brazos Gualeguaycito y Mandisoví presentan valores mayores de 3,0 bits/individuo; del lado uruguayo, los brazos Itapebí y Arapey presentan valores de 2,0 bits/individuo; en tanto que el resto del cuerpo de agua presenta valores que no superan los 2,5 bits (Figura 3).
En el otoño la situación es algo diferente; en la cola del embalse se registran valores de menos de 1 bit, los que van aumentando hasta 2,5 bits a la altura del brazo Itapebí. La situación cambia en el brazo Mocoretá, con valores de más de 3,0 bits/individuo; en el brazo Gualeguaycito y en la estación Campamento los valores son más bajos (1,5 bits/individuo) (Figura 4).
Valores de 2,5 bits/individuo -1 bit hasta el brazo Arapey y más 3,0 bits para el resto del embalse, excepto en Campamento donde se registran valores de más de 3,5 bits/individuo-fueron característicos en el invierno de 1984 (Figura 5).
Finalmente, en la primavera la situación es parecida a la del invierno, con valores de 1,5 bits para las estaciones correspondientes a la cola del embalse y de 3,0 bits en la estación Campamento; sin embargo, existen dos excepciones: brazo Mocoretá y brazo Gualeguaycito, con algo más de 3,0 bits/individuo -1 bit en la primera y más de 3,5 en la segunda, ambas del lado argentino.
Esta situación es en general concordante con los datos que sobre el embalse presentan Prenski y Baigún (1984) y Quirós et al (1984), en el sentido de que los subambientes tienen una dinámica propia y que el comportamiento de la estación correspondiente al tramo final (Campamento), reflejaría aproximadamente el comportamiento del embalse. No obstante esto no siempre es así, especialmente en otoño, donde podría influir el manejo de regulación del nivel del embalse realizado en las proximidades.
Los resultados del análisis en componentes principales aparecen en la Tabla III. Los dos primeros ejes reúnen algo más del 96% de la varianza total y son los que se representan en la Figura 7. En el plano definido por los dos primeros ejes participan las cuatro estaciones anuales de manera aproximadamente equitativa. Para el primer eje, los valores de verano y otoño, por un lado, y los de invierno y primavera, por el otro, explican casi el 60% de la varianza total. En el segundo eje, los valores de primavera se asocian con los de otoño y los de invierno se asocian con los correspondientes al verano, dando cuenta de algo menos del 37% de la varianza total. Esto provoca que, en el conjunto, las cuatro estaciones anuales ocupen lugares prácticamente equidistantes, ocupando espacios bien diferenciados (Morrison, 1976).
Por estar la matriz constituida exclusivamente con valores de diversidad, la distribución de las épocas anuales viene dada por las diferencias entre los valores de H' obtenidos en cada estación de muestreo y para estación del año considerada.
2.5 CONCLUSIONES
La presencia de 49 taxones en el conjunto del embalse para el período estudiado apoya naturalmente la noción de un ecosistema muy diversificado. El valor máximo de H' de 3.911 bits/individuo en la estación Campamento, en invierno, y el mínimo de 0.978 bits/individuo en el brazo Mandiyú, en otoño, conforman los valores extremos del rango de diversidades encontradas en el embalse, el que se podría caracterizar por una media global de 2.667 bits/individuo.
Se constatan diferencias entre los ambientes estudiados, lo cual concuerda con estudios realizados anteriormente en el área: Macoretá en dos ocasiones, y Mandisoví y Gualeguaycito parecen ser los ambientes que aportan más diversidad, por ser los más diversificados y que poseen una dinámica atribuible probablemente a sus propias cuencas.
Las medidas de diversidad específica parecen reflejar bien los cambios estacionales y entre los ambientes, aunque lo restringido del período estudiado (un año), sugiere la conveniencia de extender este estudio a un período más largos. Igualmente parece conveniente comparar los valores de diversidad con otros parámetros ambientales y biológicos, así como ensayar el índice propuesto con otro tipo de unidades de uso frecente, tales como las CPUE o las de biomasa, lo que aportaría nueva información.
Hay un comportamiento propio de cada estación anual considerada, el que se pone de manifiesto a través del análisis en componentes principales. El sesgo producido por el arte de pesca utilizado puede considerarse como una restricción importante en la validez global de los resultados; de cualquier manera, el compromiso entre las posibilidades materiales (que restringen el uso de otros tipos de artes) y el límite que se impone en general a la obtención de inventarios exhaustivos, son situaciones comunes a la actividad del biológo.
Finalmente, parece ser conveniente disponer de series de datos que abarquen períodos más largos, así como la exploración de los brazos aguas arriba, con el fin de determinar los rangos de variación de cada brazo y consecuentemente, tratar de establecer más ajustadamente el aporte de cada uno de éstos a la dinámica del embalse.
2.6 BIBLIOGRAFIA
CHEJDIAK G. et al. 1986. Metodología de muestreo y estimación de abundancia relativa de peces en el embalse de Salto Grande. EN: Editores Vila, I. y E.Fagetti. Trabajos presentados al Taller Internacional sobre Ecología y Manejo de peces en lagos y embalses. Santiago, Chile, COPESCAL. Doc. Técnico. (4):237.
LEGENDRE, L. y LEGENDRE, P. 1984. Ecologie numerique. Masson et Cie. Vols. 260 + 350 p.
MARGALEF, R. 1974. Ecología. Barcelona, España, Ed. Omega. 944 p.
MARGALEF, R. 1980. La Biosfera, entre la termodinámica y el juego. Barcelona, España, Ed. Omega. 236 p.
MARGALEF, R. 1983. Limnología. Barcelona, España, Ed. Omega. 1010 p.
MORRISON, D.F. 1976. Multivariate statistical methods. 2nd. Ed. New York, U.S.A., McGraw Hill Book Co. 415 p.
PIELOU, E.C. 1977. Mathematical ecology. New York, U.S.A., J. Wiley & Sons. 385 p.
ORLOCI, L. 1975. Multivariate analysis in vegetation research. The Hague, Dr. W. Junk B.V. 276 p.
PRENSKI, L.B. y BAIGUN, C.R. 1984. Resultados de la prospección pesquera en el embalse de Salto Grande (febrero 1980–febrero 1981). B. Aires, Argentina, Inst. Nac. Invest. Des. Pesq. 440 p.
QUIROS, R. 1980. Evaluación del rendimiento pesquero potencial del embalse de Salto Grande. B. Aires, Argentina, Inst. Nac. Invest. Des. Pesq. (395):18 p.
QUIROS, R. y CUCH, S. 1982. Características limnológicas del embalse de Salto Grande. I. Cambios estacionales de ciertos parámetros físico químicos. Ecología. 7:195–224.
QUIROS, R., PRENSKI, L.B. y BAIGUN, C.R. 1984. Relación entre resultados de ensayos de captura y factores ambientales en el embalse de Salto Grande. B. Aires, Argentina, Inst. Nac. Invest. Des. Pesq. (425):21.
SHANNON, C.E. and WEAVER, W. 1963. The mathematical theory of communication. Urbana, Illinois, U.S.A., Univ. Illinois Press. 154 p.
VIEIRA DA SILVA, J. 1979. Introduction a la theorie ecologique. Paris, France, Masson et Cie. Coll. d'Ecol. 14. 112 p.
TABLA I. Código y nombre común de las especies estudiadas en el presente trabajo.
| cód. | nombre común | taxones presentes |
| Familia ENGRAULIDAE | ||
| 201 | anchoa | Lycongraulis olidus |
| Familia TETRAGONOPTERIDAE | ||
| 401 | mojarra | Holoshesthes pequira |
| 402 | mojarra | Astyanax (P.) abramis |
| 403 | mojarra | Astyanax (A.) fasciatus fasciatus |
| 404 | dientudo transparente | Asiphonicthys stenopterus |
| 405 | dientudo jorobado | Cyrtocharax squamosus |
| 406 | dorado | Salminus maxillosus |
| 407 | chafalote | Raphiodon vulpinus |
| 408 | dientudo paraguayo | Acestrorhyncus altus |
| 409 | dientudo común | Oligosarcus hepsetus |
| 410 | virolito | Apareidon affinis |
| 412 | sabalito plateado | Curimatorbis platanus |
| 413 | sabalito (1) chico | Pseudocurimata gilberti |
| 414 | sabalito (2) rayado | Pseudocurimata nitens |
| 415 | sabalito (3) | Curimatinae sp. |
| 416 | sábalo | Prochilodus platensis |
| 417 | boga (1) lisa o lagunera | Schizodon platae |
| 418 | boga (2) común | Leporinus obtusidens |
| 419 | boga (3) juncalera | Schizodon nasutus |
| 422 | mojarra (4) | sin determinar |
| 423 | sabalito (4) | sin determinar |
| 424 | mojarra (5) | Astianax sp. |
| Familia ERYTHRINIDAE | ||
| 501 | tararira | Hoplias malabaricus malabaricus |
| Familia SERRASALMIDAE | ||
| 601 | piraña (1) | Serrasalmus spilopleura |
| 602 | piraña (2) | Serrasalmus nattereri |
| Familia AGENEIOSIDAE | ||
| 701 | manduvi | Ageneiosus valenciennesi |
| Familia AUCHENIPTERIDAE | ||
| 801 | buzo | Auchenipterus nuchalis |
| 802 | torito | Parauchenipterus galeatus |
| Familia DORADIDAE | ||
| 901 | armado común | Pterodoras granulosus |
| 902 | armado marieta | Rhinodoras d'orbigny |
| Familia PIMELODIDAE | ||
| 1001 | bagre porteño | Parapimelodus valenciennesi |
| 1003 | cucharón | Sorubim lima |
| 1004 | bagre trompudo | Iheringichtys westermanni |
| 1005 | bagre amarillo | Pimelodus clarias |
| 1006 | bagre sapo o negro | Rhamdia sapo |
| 1007 | pati | Luciopimelodus pati |
| 1008 | surubi | Pseudoplatystoma coruscans |
| Familia LORICARIDAE | ||
| 1201 | vieja de agua (1) | Plecostomus sp. |
| 1202 | vieja de agua (2) | Rhinelepis sp. |
| 1203 | vieja de agua (3) | Loricaria anus |
| 1204 | vieja de agua (4) | Loricaria sp. |
| Familia SCIANIDAE | ||
| 1301 | corvina de rio | Pachyurus bonariensis |
| Familia CICHLIDAE | ||
| 1401 | chanchita | Geophagus sp. |
| 1402 | cabeza amarga | Crenicichla sp. |
| Familia ACHIRIDAE | ||
| 1501 | lenguado | Achirus jenynsi |
| Familia GYMNOTOIDEI | ||
| 1601 | banderita | Eigenmannia virescens |
| 1602 | morena (1) | Gymnotus carapo |
| 1603 | morena (2) | Gymnotoidei sp. |
| Familia SYNBRANCHIDAE | ||
| 1801 | anguila criolla | Symbranchus marmoratus |
TABLA II. Diversidad especifica (H') para cada una de las estaciones de muestreo realizadas en 1984. H' expresada en bits.individuo-1.
| estación de muestreo | verano | otoño | invierno | primavera | media anual |
| CAMPAMENTO | 2.991 | 1.923 | 3.911 | 3.151 | 2.994 |
| ITAPEBI | 2.276 | 2.864 | 3.079 | 3.146 | 2.841 |
| GUALEGUAYCITO | 2.318 | 1.843 | 3.382 | 3.737 | 2.820 |
| MANDISOVI | 3.557 | 2.765 | 3.402 | 2.101 | 2.956 |
| ARAPEY | 2.357 | 2.339 | 3.239 | 2.196 | 2.548 |
| MOCORETA | 2.896 | 3.032 | 2.836 | 3.177 | 2.985 |
| YACUI | --- | 2.034 | 2.504 | 1.566 | 2.125 |
| MANDIYU | --- | 0.978 | 2.667 | 1.713 | 1.786 |
| media estacional | 2.733 | 2.256 | 3.128 | 2.598 |
TABLA III. Resultados del Análisis en Componentes Principales utilizando valores de H'.
| vector propio | eigen value | % de varianza explicada | % de varianza acumul. |
| 1 | 1.242224 | 59.8096 | 59.8096 |
| 2 | 0.765138 | 36.8392 | 96.6488 |
| 3 | 6.96.10-2 | 3.3511 | 99.9999 |
| 4 | 3.28.10-7 | 1.57.10-6 | 99.9999 |
| 5 | 1.74.10-8 | 8.38.10-8 | 99.9999 |
| 6 | 1.89.10-9 | -9.11.10-7 | 100. |
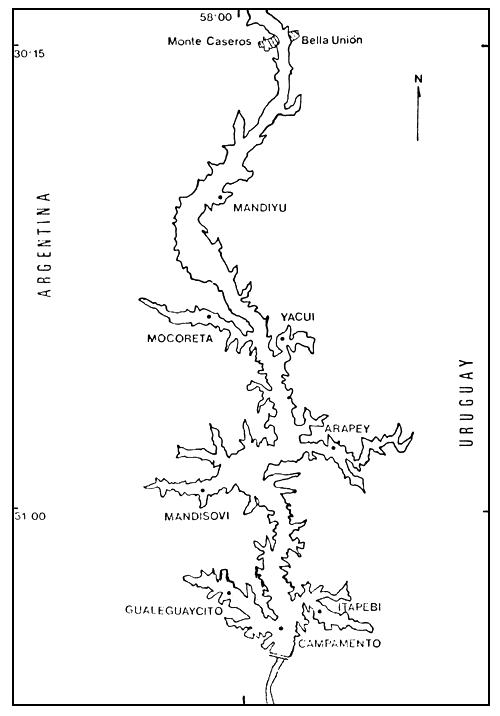
Figura 1. Ubicación de las estaciones de muestreo en el embalse de Salto Grande en el presente trabajo.
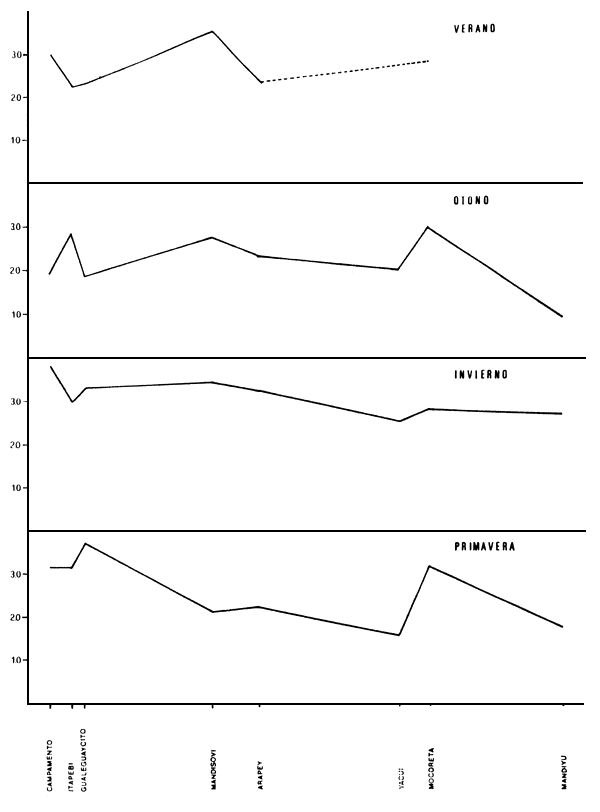
Figura 2. Variación de los valores de diversidad (H'), para cada estación anual durante el año 1984. Los valores de H' expresados en bits.individuo-1.
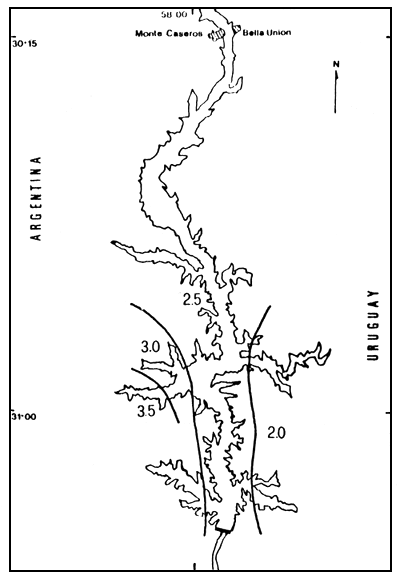
Figura 3. Distribución espacial de los valores de diversidad (H'), para el verano de 1984. H' expresado en bits.individuo-1.
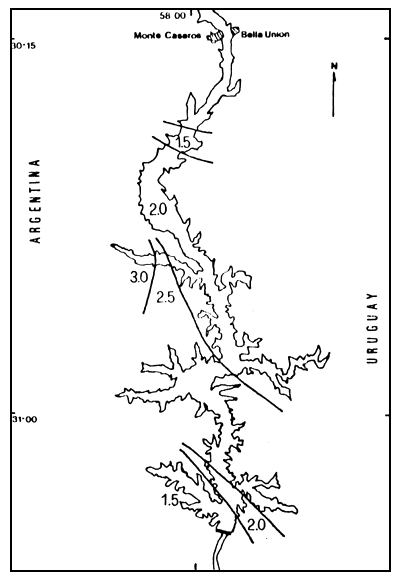
Figura 4. Distribución espacial de los valores de diversidad (H'), para el otoño de 1984. h" expresado en bits. individuo-1.
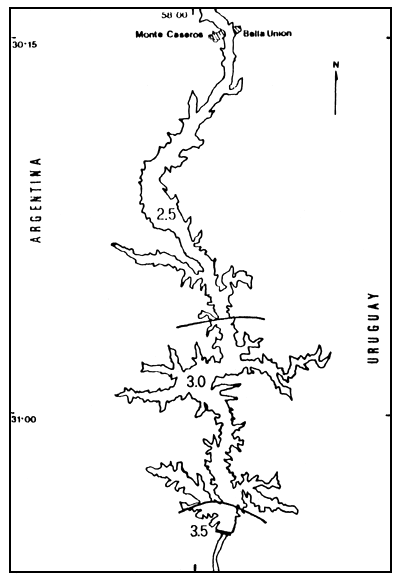
Figura 5. Distribución espacial de los valores de diversidad (H'), para el invierno de 1984. h' expresado en bits.individuo-1.
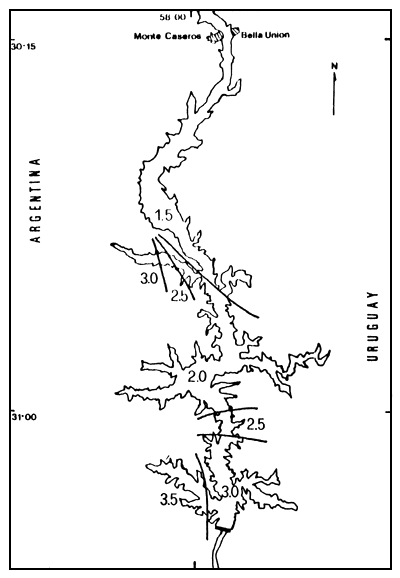
Figura 6. Distribución espacial de los valores de diversidad (H'), para la primavera de 1984. H' expresado en bits.individuo-1.
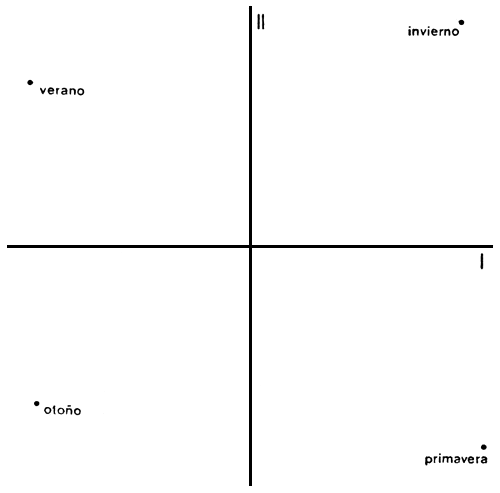
Figura 7. Distribución de las estaciones anuales en función de los dos primeros ejes resultantes del análisis en componentes principales.
![]()
![]()
![]()