![]()
![]()
![]()
A.M. Lafarga y C.A. Guerrero
Departamento de Aguas Continentales
Instituto Nacional de Investigación y Desarrollo Pesquero
Buenos Aires, Argentina
4.1 RESUMEN
Se realizó un estudio de crecimiento de distintas poblaciones de trucha arco iris, Oncorhynchus mykiss, en un conjunto de cinco lagos y lagunas de la Patagonia Argentina. La edad fue leída en escamas provenientes de campañas realizadas entre los meses de mayo de 1983 y marzo de 1985.
El estudio revela que existe relación entre la tasa de crecimiento, el estado trófico del ambiente y las densidades poblacionales. Así, en ambientes con gran disponibilidad de alimento y altas densidades poblacionales, los individuos crecen rápidamente, pero no alcanzan tallas elevadas. La presencia de aterínidos afectaría negativamente el crecimiento de la especie Asimismo, se observó que existe una relación inversa entre la tasa de crecimiento y el tiempo de permanencia en los ríos donde nacieron.
ABSTRACT
The growth of different populations of rainbow trout (Oncorhynchus mykiss) from five Patagonian lakes and ponds (Argentina) was studied. Age has been determined on scales obtained from samples performed since May 1983 to March 1985.
The results show a relation between the individual growth rate, the trophic environmental characteristics and the population densities Thus in environments with high food availability and high population densities, the individuals tend to grow faster but with lower lengths. The presence of atherinids seems to affect negatively the growth of this species. Also, a negative relation was found between the growth rate and the time spent by individuals at the nursery streams.
4.2 INTRODUCCION
Se define crecimiento como el incremento en peso o tamaño de un sistema orgánico, resultado del balance entre los procesos catabólicos y anabólicos, distinguiéndolo del proceso de diferenciación, el cual implica un incremento en la organización (Bartalanffy, 1938).
El estudio revela diferencias en las capacidades de asimilación de los peces, no sólo entre especies, sino también a través de las distintas etapas del desarrollo o entre distintos ambientes (Weartherley, 1976). Muchos autores han tratado este tema, encontrando relación entre el crecimiento y la temperatura (Baldwin, 1956), la altitud y latitud (Hazel et al., 1983) el estado trófico de los ambientes (Campbell, 1971, 1979; Ryan y Harvey, 1977) la interacción entre especies (Larkin y Smith, 1954; Crossman, 1959a, 1959b; Burrough y Kennedy, 1978) y la densidad poblacional (Mottley, 1941; Havey, 1980). Asimismo, se ha comparado, en distintos ambientes, el efecto que tienen dichas variables sobre el crecimiento (Leggett y Power, 1969; Pivnicka, 1983).
El método más frecuentemente usado en la asignación de edad en peces es el de estudio y determinación de marcas de crecimiento en estructuras óseas (vértebras, espinas, opérculos, escamas, otolitos), las cuales se forman durante períodos alternantes de crecimiento rápido y lento, y reflejan tanto influencias ambientales como internas. Otros métodos utilizados son el análisis de la distribución de las frecuencias de tallas en la población, métodos de marcado y recaptura y métodos de observación directa en cautiverio (Bagenal, 1978).
El objetivo del presente trabajo es estudiar el crecimiento de la trucha arco iris (Oncorhynchus mykiss en cinco ambientes de la Patagonia e intentar relacionarlo con los factores abióticos y bióticos que lo afectan. Si bien con anterioridad se han realizado en el país estudios en dicha especie en relación con la biología general (Baiz y Candia, 1978; Guijo et al., 1980), taxonomía (Cordiviola de Yuan, 1974), reproducción (Baix, 1979), interacciones tróficas (Baix et al., 1982, 1983) y crecimiento en cautividad (Baix, 1973), desafortunadamente no se han emprendido estudios sobre el crecimiento y los factores que lo afectan (Ringuelet et al., 1981, 1982), por lo que se consideró importante en esta oportunidad desarrollarlo, dentro de un plan de relevamientos de distintos ambientes.
4.3 MATERIALES Y METODOS
Entre mayo de 1983 y marzo de 1985 se desarrolló un convenio entre el Instituto Nacional de investigación y Desarrollo Pesquero y la Provincia del Chubut, para estudiar el potencial pesquero en ambientes lénticos de dicha Provincia. Durante el mismo se realizaron muestreos limnológicos y pesqueros en los lagos Musters y Rosario, y en las lagunas Esquel, Brechman y Zeta, realizándose campañas trimestrales, en coincidencia con las estaciones del año.
El lago Musters se halla en la planicie patagónica y pertenece a la cuenca del río Senguer, el cual constituye su afluente; mientras que el lago Rosario, ubicado en la región precordillerana, pertenece a la cuenca del río Futaleufú y cuenta con el río Nantifal como afluente. Las lagunas Esquel, Brechman y Zeta se encuentran también en la región precordillerana, pero mientras que las lagunas Esquel y Brechman poseen arroyos temporarios que las alimentan, la laguna Zeta carece de arroyos de importancia (Figura 1). En la Tabla 1 se presentan algunas características de los ambientes considerados.
Durante las campañas se recolectó material de escamas de las especies presentes, ocupando parte de ellas para realizar el presente trabajo. Las escamas de O mykiss se limpiaron en el laboratorio, bajo lupa, con detergente enzimático de uso doméstico, seleccionando cuatro a seis escamas por ejemplar, entre aquéllas que no presentaban signos de regeneración. Posteriormente estas escamas se montaron en seco entre dos portaobjetos, midiendo de dos a cuatro escamas por ejemplar en un proyector óptico de perfiles, con un aumento de 20 X y con una precisión de 1 mm de proyección.
En cada escama se midió el radio total (R) y la distancia del foco a cada marco (r). Ambas mediciones se realizaron en el campo anterior de la escama, sobre el eje que une el foco con el margen anterior más extremo, promediando luego las lecturas entre escamas del mismo ejemplar. Para la asignación de marcas, se siguió el criterio de considerar la marca como una banda de circuli apretados y poco robustos, que no se extiende más allá de los campos antero-laterales de la escama, cortando en su recorrido a los circuli espaciados (Greeley, 1933; Brown y Holton, 1953; Hunt y Jones, 1972a).
Las marcas fueron consideradas anuales, estimándose mediados de primavera como la época de marcación (Baix y Candia, 1978; Wetzlar, 1979; Guerrero y Lafarga, 1986). Se diferenciaron en las escamas períodos de crecimiento en río o arroyo, y períodos de crecimiento en lagos, sobre la base del mayor espaciamiento que muestran los circuli en el último caso durante la estación de crecimiento (Grelley, 1933).
Una vez adjudicada la edad a cada marca, se retrocalculó la talla individual a cada edad. Para ello se estudió, en cada ambiente, la relación entre el radio total de la escama (R) y el largo total de los individuos (L). En todos los ambientes se encontró que el mejor ajuste era una relación lineal:
L = a + b . R
Las constantes fueron calculadas por el método de regresión lineal funcional (Ricker, 1973). Posteriormente se realizó el retrocálculo de tallas para cada marca de las escamas de los individuos de cada ambiente, utilizando la siguiente ecuación (Fraser, 1916, Lee, 1920):
ln = a + (L - a) . (rn/R)
donde: L : largo total del individuo del que se tomó la escama
R : radio total de la escama
ln : largo total del individuo cuando formó la marca n
rn : radio de la marca n
Con los datos de tallas retrocalculadas a cada edad y para cada individuo, se ajustó para el lago Musters y la laguna Esquel una curva de crecimiento de von Bertalanfly, por el método de Allen, (1966):
lt = L∞ . [1 - e -K (t - to)]
donde: lt: largo a la edad t
L∞ : largo asintótico de la curva de crecimiento
K : medida de la velocidad a la cual la curva de crecimiento se
aproxima al largo asintótico
to : edad (hipotética) a la cual comienza el crecimiento.
Para el resto de los ambientes fue imposible realizar el ajuste a tal modelo, pues no lo permite el número de clases de edad encontradas. A fin de analizar las posibles relaciones entre el crecimiento y las variables físicas, químicas y biológicas de los ambientes estudiados, se realizó un análisis de correlación lineal entre dichas variables y el crecimiento (expresado éste por medio de las tallas retrocalculadas a las edades I y II). Para disminuir la correlación entre el valor medio y la varianza de las variables, se realizó una transformación logarítmica previa en las variables.
Por último, se estudió la relación largo-peso en los distintos ambientes, utilizando la ecuación:
W = a . Lb
donde: W : peso total
L : largo total
a y b: son constantes
En cada caso las relaciones fueron determinadas a través de su transformación logarítmica previa, según el método de regresión lineal funcional (Ricker, 1973).
4.4 RESULTADOS
En las escamas de O mykiss de las lagunas Esquel y Brecham, la zona central consiste en pocos circuli apretados y es seguida por una zona de gran crecimiento. Se asocia la primera con crecimiento en arroyo, y la zona adyacente de circuli muy espaciados, con crecimiento en la laguna. Es decir, que estos individuos permanecerían menos de un año en el arroyo y luego pasarían a la laguna.
Por el contrario, en las escamas de la mayoría de los individuos del lago Rosario se encontró una zona central de circuli apretados, rodeadaa por una banda de circuli muy apretados (la marca anual). Esta zona es seguida por una zona de gran crecimiento, indicada por circuli muy espaciados. En este caso, se adjudica la marca (I) como de río, por lo cual los individuos permanecerían durante un año en el. Luego de esto pasarían al lago, tal cual lo indica la zona de gran crecimiento posterior a esta marca. En el lago Musters se encontrarían individuos que permanecen menos de un año en el río, e individuos que estarían hasta un año en él.
La laguna Zeta carece de afluentes o efluentes de importancia, lo cual se manifiesta en las escamas, por ejemplo en la ausencia de una zona central marcada. La Tabla 2 muestra el resultado para cada ambiente del ajuste de la relación entre el largo total de los individuos y el radio total de las escamas. Estas relaciones están graficadas en las Figuras 2 a 6.
Las lagunas Zeta y Brecham presentan tallas retrocalculadas mayores que el resto de los ambientes. Para el primer año de edad, la laguna Esquel presenta una talla retrocalculada mayor que la del lago Musters. Pero ya al segundo año de edad, los individuos del lago Musters que permanecen menos de un año en el río, presentan una talla mayor que la respectiva de la laguna Esquel.
Los individuos del lago Rosario y aquéllos del lago Musters que permanecen hasta un año en el río, presentan las menores tallas retrocalculadas encontradas (Tabla 3, Figura 7). La curva de crecimiento de O mykiss de la laguna Esquel posee un largo asintótico (L∞) menor y una tasa de crecimiento (K) mayor que los respectivos valores del lago Musters (Tabla 4). Esto determina que dichas curvas se crucen aproximadamente al año y medio de edad (Figura 7).
Estadísticamente, los coeficientes de correlación entre los parámetros físicos, químicos y biológicos y las tallas retrocalculadas a las edades I y II son significativos, con P < 0,05, para los niveles totales de nutrientes (Tabla 5). Se puede observar que existe una relación positiva entre los niveles de nutrientes y las tallas retrocalculadas a dichas edades. Esta relación se cumple sobre todo con el nitrógeno total y es mayor para el primer año de edad. Cabe señalar que se obtienen resultados semejantes si para la relación L versus R se utiliza una única recta elaborada con los valores de L y R de todos los ambientes.
En la Tabla 6 se detallan las relaciones largo-peso para los peces de cada ambiente. Para la misma longitud, los individuos de la laguna Zeta alcanzan un peso mucho mayor que en el resto de los ambientes (Figura 8).
4.5 DISCUSION Y CONCLUSIONES
Los valores de tallas retrocalculadas encontradas en el presente trabajo para Esquel, Musters y Rosario, estarían dentro de los intervalos señalados por Wetzlar (1979) para O. mykiss en siete lagos chilenos. Los valores para Brecham y Zeta superan a los de este último autor y estarían dentro de los valores más altos encontrados para O. mykiss según Carlander (1969), quien realizó una revisión de datos sobre dicha especie hasta ese momento.
Se puede considerar las características climáticas, morfométricas y edáficas de los lagos como determinantes de su producción pesquera (Quiros et al., 1986). Teniendo en cuenta esto, la laguna Esquel sería un ambiente relativamente más productivo que el lago Musters (Tabla 1). En dicha laguna, el crecimiento de O. mykiss coincide con lo que se esperaría en un ambiente que ofrece mayor disponibilidad de alimento, encontrando así individuos con un alto ritmo de crecimiento inicial (Weatherley, 1972). Contrariamente, en el lago Musters, el ritmo de crecimiento inicial es menor. Por otro lado, debido al efecto que ejercerían las mayores densidades de O. mykiss y de otras especies presentes, se alcanzaría en el primer ambiente una talla asintótica menor que en el lago Musters.
Son conocidos los efectos negativos de la densidad poblacional (Wetzlar, 1979) y de la competencia interespecífica (Weatherley, 1976) sobre la talla asintótica. El mismo patrón de crecimiento, mayores crecimientos iniciales y menores largos asintóticos para los lagos más productivos con poblaciones autosostenidas, fue hallado para lagos del sur de Chile (Wetzlar, 1979). Quedan por explicar diferencias en el crecimiento de O. mykiss entre ambos conjuntos de lagos, con mayores crecimientos iniciales en los lagos chilenos. Las mayores tasas de crecimiento (K) de los lagos chilenos (Wetzlar, 1979) posiblemente estén ligadas a las mayores temperaturas de éstos en relación a los lagos del lago argentino, con características climáticas continentales (Quirós y Drago, 1985).
Para el primer año de edad, la talla media retrocalculada para la laguna Esquel es mayor que la del lago Musters, pero para los individuos del lago Musters que permanecen menos de un año en el río este proceso se revierte a partir del segundo año de edad (Tabla 3). Esta es la causa de que la relación entre las tallas medias retrocalculadas y los niveles de nutrientes sea mayor para el primer año de edad.
Las altas tallas encontradas para esta especie en las lagunas Brecham y Zeta podrían deberse a la gran disponibilidad de alimento y a las bajas densidades poblacionales presentes. Si bien la composición en especies en los distintos ambientes no es uniforme, se observa una relación negativa entre la densidad de aterínidos y las tallas de la trucha arco iris (Tabla 5). La competencia interespecífica ejercería un efecto negativo sobre el crecimiento.
Una interacción de este tipo entre O. mykiss y los aterínidos Basilichthys microlepidotus y Odontesthes bonariensis fue señalada por Baiz et al. (1982, 1983) para una laguna de la meseta patagónica. Una relación negativa entre abundancia relativa de salmónidos y de aterínidos, a escala amplia, luego de eliminar el efecto climático, fue señalada por Quirós (1987b) para lagos y embalses de Argentina. Este hecho, sumado al efecto producido por una baja disponibilidad de alimento y una baja densidad poblacional, podrían ser la causa de las bajas tallas encontradas para la trucha arcoiris en el lago Rosario.
Los resultados para el lago Musters muestran que existe una relación entre el crecimiento y el tiempo de permanencia de las crías en el río donde nacieron. Se observa así una tasa de crecimiento mayor para los individuos que abandonaron antes el río, comparada con la de aquéllos que permanecieron en él durante un año. La diferencia entre estos dos grupos disminuye con la edad, es decir, los individuos de edades mayores tienden a tener tallas semejantes, independientemente de su tiempo de permanencia en el río.
Estos resultados son coincidentes con los señalados por Kwain (1971) para esta especie en el lago Superior, y con los señalados por Kennedy (1976), en Irlanda, para Salmo trutta. La ampliación de la presente base de datos con la inclusión de un mayor número de ambientes, permitiría contar con mayores elementos para predecir el crecimiento de O. mykiss en función de determinados factores abióticos y bióticos.
4.6 AGRADECIMIENTOS
El presente trabajo fue financiado por el Instituto Nacional de Investigación y Desarrollo Pesquero, la Provincia del Chubut y por PID/CONICET 3-017400/85 otorgado a R. Quirós. Se agradece expresamente la colaboración de la Dirección de Intereses Marítimos y Pesca Continental de la Provincia del Chubut.
4.7 BIBLIOGRAFIA
ALLEN, K.R. 1966. A method of fitting growth curves of the von Bertalanffy type to observed data. IN: J. Fish. Res. Board Canada. 23(2):163–179.
BAGENAL, T. 1978. Methods for assessment of fish production in freshwaters. Blackwell Scientific Publications Ltd. IBP Handbook No 3. 365 p.
BAIZ, M. de L. 1973. Crecimiento en cautividad de Salmo gairdneri Richardson, 1936, y sus posibilidades comerciales. Servicio Nacional de Pesca, Inst. Nac. de Inv. Des. Pesq. 11.
BAIZ, M. de L. 1979. Observaciones sobre la reproducción de Salmo gairdneri de lago Gutiérrez, Parque Nacional Nahuel Huapi. EN: Reunión Argentina de Ecología, 7. Mendoza, Argentina, 22–27 de abril.
BAIZ, M. de L. y CANDIA, C.R. 1978. Características estructurales de la población de trucha arco iris (Salmo gairdneri-Salmonidae) de laguna Ñe-Luán, Maquinchao, Río Negro; datos Preliminares. EN: Ecología. 3:31–42.
BAIZ, M. de L., BELLO, M.T. y BUNGE, M.M. 1982. Respuesta de Salmo gairdneri a la introducción del pejerrey en la laguna Ñe-Luán, Maquinchao (R.N.). EN: Reunión Argentina de Ecología, 10. Mar del Plata, Argentina, 18–23 de abril.
BAIZ, M. de L., BELLO, M.T. y BUNGE, M.M. 1983. Interacción trófica entre trucha y pejerrey de laguna Ñe-Luán (R.N.). EN: Reunión Argentina de Ecología, 11. Villa Giardino, Córdoba, Argentina, 17–23 de abril.
BALDWIN, N.S. 1956. Food consumption and growth of brook trout at different temperatures. Trans. Am. Fish. Soc. 86:323–328.
BERTALANFFLY, L. von. 1938. A quantitative theory of organic growth inquiries on growth laws. II. Human Biology. 10(2):181–213.
BROWN, C.J.D. and HOLTON, G.D. 1953. Time of annulus formation on the scales of brook and rainbow trout. Trans. Am. Microsc. Soc. 72(1):47–48.
BURROUGH, R.J. and KENNEDY, C.R. 1978. Observations on the brown trout (Salmo trutta) and perch (Perca fluviatilis) of Malham Tarn, North Yorkshire. Fld. Stud. 4:631–643.
CAMPBELL, R.N. 1971. The growth of brown trout Salmo trutta L. IN: Northern Scottish lochs with special reference to the improvement of fisheries. IN: J. Fish. Biol. 3:1–28.
CAMPBELL, R.N. 1979. Ferox trout, Salmo trutta L., and charr, Salvelinus alpinus (L.), in Scottish lochs. IN: J. Fish. Biol. 14:1–29.
CARLANDER, K.D. 1969. Handbook of freshwater fishery biology. Vol. 1. U.S.A., Iowa State University Press. 752 p.
CORDIVIOLA DE YUAN, E. 1974. Sobre la ubicación taxonómica de la “Trucha Arcoiris” del lago Cardiel, Prov. de Santa Cruz (Pisces, Salmonidae). Physis. B33(87):247–248.
CROSSMAN, E.J. 1959a. Distribution and movements of a predator, the rainbow trout and its prey, the redside shiner, in Paul lake, British Columbia. IN: J.Fish. Res. Bd. Canada. 16(3):247–267.
CROSSMAN, E.J. 1959b. A predator prey interaction in freshwater fish. IN: J. Fish. Res. Board Canada. 16(3):269–281.
FRASER, C.McL. 1916. Growth of the spring salmon. Seattle, U.S.A., Trans. Pacif. Fish. Soc. (1915). pp. 29–39.
GREELEY, J.R. 1933. The growth rate of rainbow trout from some Michigan waters. Trans. Am. Fish. Soc. 63:361–379.
GUERRERO, C.A. y LAFARGA, A. 1986. Edad de la trucha arco iris (Salmo gairdneri) de la laguna Zeta (Prov. de Chubut). Argentina, Inst. Nac. de Investigación y Desarrollo Pesquero. Serie Informes Técnicos del Departamento de Aguas Continentales No 4. 15 p.
GUIJO, A., COMPAGNUCCI, L. y HERRERA, C. 1980. Estudio sobre la biología de la trucha arcoiris (Salmo gairdneri) del lago Cardiel (Provincia de Santa Cruz). EN: Reunión Argentina de Ecología, 8. Santa Fé, Argentina, 11–17 de mayo.
HAVEY, K.A. 1980. Stocking rate and the growth and yield of landlocked atlantic salmon at Long Fond, Maine. Trans. Am. Fish. Soc. 109:502–510.
HAZEL, P.P., LAMOUREUX, J., MAGNIN, E. et NAULT, R.. 1983. Croissance de six especes de poissons vivant pres de leur limite de repartition en latitude et en altitude sur le territoire de la Baie James, Cybium. 7(4):57–69.
HILE, R. 1970. Body-scale relation and calculation of growth in fishes. Trans. Am. Fish. Soc. 99(3):468–474.
HUNT, P.C. and JONES, J.W. 1972a. Trout in Llyn Alaw, Anglesey, North Wales. I. Population structure and angling returns. IN: J. Fish. Biol. 4:395–408.
HUNT, P.C. and JONES, J.W. 1972b. Trout in Llyn Alaw, Anglesey, North Wales. II. Growth. J. Fish. Biol. 4:409–424.
KENNEDY, G.J.A. 1976. Age, growth and feeding studies on Lough Neagh trout Salmo trutta (L). Fish. Res. Lab., Coleraine. IN: Fish. Res. Leaflet. 10:14.
KWAIN, W.H. 1971. Life history of rainbow trout (Salmo gairdneri) in Batchawana Bay, Eastern Lake Superior. IN: J. Fish. Res. Board Canada. 8(5):771–775.
LAFARGA, A.M. y GUERRERO, C.A. 1987. Crecimiento de Salmo gairdneri y Percichthys trucha en dos ambientes lénticos de la Patagonia. EN: Reunión Argentina de Ecología, 13. Bahía Blanca, 5–9 de abril.
LARKIN, P.A. and SMITH, S.B. 1954. Some effects of introduction of the redside shiner on the Kamloops trout in Paul lake, British Columbia. Trans. Am. Fish. Soc. 83:161–175.
LEE, R.M. 1920. A review of the methods of age and growth determination by means of scales. Fishery Invest. Lond. Ser. II,4,2. 32 p.
LEGGETT, W.C. and POWER, G. 1969. Differences between two populations of landlocked Atlantic salmon (Salmo salar) in Newfoundland. IN: J. Fish. Res. Board Canada 26:1585–1596.
LOPEZ, H.L., MENNI, R.C. y RINGUELET, R.A. 1981. Bibliografía de los peces de agua dulce de Argentina y Uruguay; 1967–1981. EN: Biología Acuática № 1. La Plata, Argentina, Inst. de Limnología ILPLA. 100 p.
LOPEZ, H.L., MENNI, R.C. y RINGUELET, R.A. 1982. Bibliografía de los peces de agua dulce de Argentina y Uruguay; Suplemento 1982. EN: Biología Acuática N№ 3. La Plata, Argentina, Inst. de Limnología ILPLA. 26 p.
MOTTLEY, C. McC. 1941. The effect of increasing the stock in a lake on the size and condition of rainbow trout. Trans. Am. Fish. Soc. 70:414–420.
PIVNICKA, K. 1983. Growth capacity of some fish species in different environmental conditions. Vest. Cs. Spolec. Zool. 47:272–287.
QUIROS, R. 1987a. Relationships between air temperature, depth, nutrients and chlorophyll in 103 Argentinian lakes. Verh. Internat. Verein. Limnol. 23:1013–1023. In press.
QUIROS, R. 1987b. Factores que afectan la distribución de salmónidos en Argentina. EN: Curso Taller Lat. Enfoques integrados para el estudio de lagos y reservorios, como base para su gestión. Santiago, Chile, 28 septiembre–10 octubre 1987.
QUIROS, R. y SCAPINI DE ORFILA, M. 1984. Características limnológicas de los ambientes. EN: Editor Quirós R. Evaluación del potencial pesquero en seis lagos de la Provincia del Chubut. Informe de Avance N№ 1, Primera Parte. Departamento de Aguas Continentales, Inst. Nac. de Investig. y Desarrollo Pesquero. Mimeografiado.
QUIROS, R. y DRAGO, E. 1985. Relaciones entre variables físicas, morfo métricas y climáticas en lagos patagónicos. EN: Rev. Asoc. Cs. Nat. Litoral. 16:181–199.
QUIROS, R. y BAIGUN, C. 1986. Prospección pesquera en 33 lagos y embalses patagónicos (Argentina). EN: Editoras Vila I. y Fagetti, E. Trabajos presentados al Taller Internacional sobre Ecología y manejo de peces en lagos y embalses. Santiago, Chile, 5–10 noviembre 1984. COPESCAL. Documento Técnico. (4):159–179.
QUIROS, R., CUCH, S. y BAIGUN, C. 1986. Relación entre abundancia de peces y ciertas propiedades físicas, químicas y biológicas, en lagos y embalses patagónicos (Argentina). EN: Editoras Vila, I. y Fagetti, F. Trabajos presentados al Taller Internacional sobre Ecología y manejo de peces en lagos y embalses. Santiago, Chile, 5–10 noviembre 1984. COPESCAL. Documento Técnico. (4):180–202.
RICKER, W.E. 1973. Linear regressions in fishery research. IN: J. Fish. Res. Board Canada. 30:409–434.
RICKER, W.E. 1975. Computation and interpretation of biological statistics of fish populations. IN: J. Fish. Res. Board Canada. pp. 191–382.
RINGUELET, R.A., ARAMBURU, R.H. y ALONSO DE ARAMBURU, A. 1967. Los peces argentinos de agua dulce. Provincia de Buenos Aires, Comisión de Investigación Científica. 602 p.
RYAN, P.M. and HARVEY, H.H. 1977. Growth of rock bass, Ambloplites rupestris, in relation to the morphoedaphic index as an indicator of an environmental stress. IN: J. Fish. Res. Board Canada. 34:2079–2088.
SCOTT W.B. and CROSSMAN, E.J. 1973. Fishwater fishes of Canada. IN: J. Fish. Res. Board Canada. pp. 184:966.
WEATHERLEY, A.H. 1972. Growth and ecology of fish populations. London, England, Academic Press Inc. 294 p.
WEATHERLEY, A.H. 1976. Factors affecting maximization of fish growth. IN: J. Fish. Res. Board Canada. 33:1046–1058.
WETZLAR, H.J. 1979. Beiträge zur Biologie und Bewirtschaftung von Forellen (Salmon gairdnerii und S. trutta) in Chile. Freiburg/Br. 264 p. Dissertation zur Erlangung des Doktorgrades vorgelegt der Fakultät fur Biologie der Albert-Ludwigs-Universität.
TABLA 1: Parámetros físicos, químicos y biológicos de los
ambientes considerados.
(A = area;
Z = profundidad media;
ALT = altura;
TEMP = temperatura media anual del aire;
LS = lectura del disco de Secchi;
TP = fósforo total;
TON = nitrógeno orgánico total;
CHLeq = clorofila equivalente;
K20 = conductividad a 20°C;
CPUET = captura total por unidad de esfuerzo;
CPUEAIR = captura de trucha arco iris por unidad de esfuerzo;
CPUEPER = captura de percíctidos por unidad de esfuerzo;
CPUEATE = captura de aterínidos por unidad de esfuerzo;
CPUEGAL = captura de galáxidos por unidad de esfuerzo;
CPUEFON = captura de trucha fontinalis por unidad de esfuerzo.
(Datos tomados de Quirós y Scapini de Orfila, 1984, Quirós y
Drago, 1985, Quirós, 1987a).
| ESQUEL | MUSTERS | ROSARIO | BRECHAM | ZETA | |
| A (Km2) | 2,8 | 414 | 14,5 | 0,3 | 0,8 |
| Z (m) | 2,0 | 20,0 | 24,9 | 3,3 | 5,9 |
| ALT (m) | 650 | 275 | 650 | 480 | 850 |
| TEMP (°C) | 8 | 10,9 | 8 | 8 | 8 |
| LS (m) | 3,1 | 1,1 | 7,0 | 1,5 | 3,6 |
| TP (mg P/m3) | 53 | 28 | 17 | 43 | 74 |
| TON (mg N/m3) | 652 | 429 | 294 | 626 | 808 |
| CHLeq (mg/m3) | 3,7 | 5,6 | 1,55 | 9,4 | 3,1 |
| PH | 7,8 | 8,1 | 7,9 | 7,8 | 8,1 |
| K20 (μs/cm) | 186 | 418 | 118 | 105 | 229 |
| IME (K20/Z) | 93,0 | 20,9 | 4,7 | 31,8 | 38,8 |
| CPUET (Kg) | 200,5 | 53 | 31,5 | 46,0 | 26,4 |
| CPUEAIR (Kg) | 40,1 | 6,9 | 7,6 | 45,5 | 26,4 |
| CPUEPER (Kg) | 160,4 | 44,5 | -- | -- | -- |
| CPUEATE (Kg) | -- | 1,6 | 23,6 | -- | -- |
| CPUEGAL (Kg) | -- | -- | 0,3 | -- | -- |
| CPUEFON (Kg) | -- | -- | -- | 0,5 | -- |
TABLA 2: Relación largo total de los individuos versus radio total de la escama, para los distintos ambientes.
| MUSTERS | L = 12,23 + 8,64 R | r = 0,943 | N = 67 |
| ROSARIO | L = 45,69 + 8,22 R | r = 0,899 | N = 41 |
| ESQUEL | L = 57,56 + 7,49 R | r = 0,895 | N = 150 |
| BRECHAM | L = 52,48 + 7,71 R | r = 0,918 | N = 66 |
| ZETA | L = 118,06 + 7,37 R | r = 0,908 | N = 61 |
TABLA 3: Tallas medias retrocalculadas (en mm) para cada edad y ambiente.
| AÑO | ESQUEL | MUSTERS (O arroyo) | MUSTERS (1 arroyo) | ROSARIO | BRECHAM | ZETA | |
| I | X | 199,41 | 196,4 | 155,05 | 151,58 | 232,43 | 290,42 |
| DS | 27,22 | 27,54 | 22,48 | 19,74 | 32,03 | 24,03 | |
| N | 25 | 20 | 21 | 20 | 18 | 47 | |
| II | X | 304 | 310,40 | 220,82 | 245,37 | 345,57 | 445,11 |
| DS | 31,92 | 46,86 | 19,02 | 26,74 | 40,33 | 34,01 | |
| N | 61 | 5 | 11 | 4 | 4 | 28 | |
| III | X | 376,54 | 406,50 | 294,75 | 352,57 | - | 666,76 |
| DS | 42,77 | - | 27,38 | 44,79 | - | 64,67 | |
| N | 28 | 2 | 4 | 9 | - | 4 | |
| IV | X | 434,27 | 485 | 403 | - | - | - |
| DS | 24,08 | - | - | - | - | - | |
| N | 11 | 2 | 2 | - | - | - | |
| V | X | 493,5 | 558 | 486 | - | - | - |
| DS | - | - | - | - | - | - | |
| N | 2 | 2 | 1 | - | - | - | |
| VI | X | - | 565 | 567 | - | - | - |
| DS | - | - | - | - | - | - | |
| N | - | 1 | 1 | - | - | - | |
| VII | X | - | 634 | 647 | - | - | - |
| DS | - | - | - | - | - | - | |
| N | - | 1 | 1 | - | - | - | |
| N | TOTAL | 127 | 33 | 41 | 33 | 22 | 79 |
TABLA 4: Resultados de los ajustes a las curvas de crecimiento de von Bertalanffy para el lago Musters (para individuos que permanecen menos de 1 año en el río) y para la laguna Esquel.
| Parámetros de la curva de crecimiento | ESQUEL | MUSTERS | |
| L ∞ (mm) | 607,279 | 783,720 | |
| DS (mm) | 71,212 | 141,825 | |
| K | 0,288 | 0,223 | |
| DS | 0,073 | 0,086 | |
| to (anos) | -0,388 | -0,287 | |
| DS (anos) | 0,199 | 0,269 | |
TABLA 5: Matríz de correlación entre las tallas retrocalculadas (para las edades I y II) y distintas variables físicas, químicas y biológicas de los ambientes estudiados. Para el lago Musters no se consideran los individuos que permanecen más de un año en el río.
| I | II | |
| (n = 5 ) | ( n = 5 ) | |
| CPUET | -0,183 | -0,246 |
| CPUEAIR | 0,602 | 0,522 |
| CPUEPER | -0,209 | -0,224 |
| CPUEATE | -0,796 | -0,735 |
| A | -0,549 | -0,492 |
| Z | -0,521 | -0,439 |
| ALT | 0,267 | 0,273 |
| TEMP | -0,147 | -0,109 |
| LS | -0,319 | -0,285 |
| TP | 0,902 * | 0,876 * |
| TON | 0,912 * | 0,876 * |
| CHLeq | 0,457 | 0,391 |
| PH | 0,326 | 0,417 |
| K20 | 0,195 | 0,251 |
| IME | 0,275 | 0,218 |
TABLA 6: Relaciones largo total versus peso total para cada ambiente.
| ESQUEL | W = 5,73. 10-6. L 3,12 |
| MUSTERS | W = 3,69. 10-6 . L 3,14 |
| ROSARIO | W = 5,04.10-6 . L 3,12 |
| BRECHAM | W = 2,46.10-5 . L 2,85 |
| ZETA | W = 7,26.10-6 .L 3,13 |
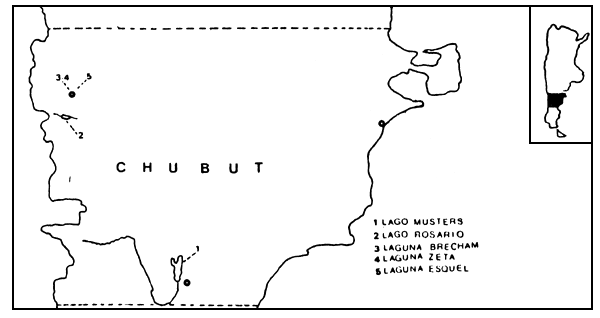
FIGURA 1: Ubicación geográfica de los ambientes considerados.
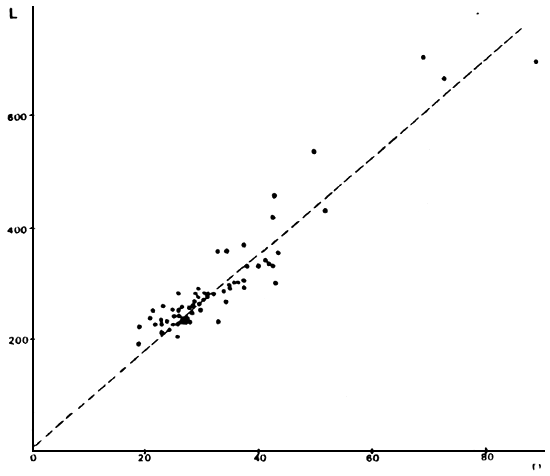
FIGURA 2: Gráfico de los valores observados y recta teórica calculada para la relación largo total (en mm) versus radio total de la escama (en mm) para los individuos del lago Musters.

FIGURA 3: Gráfico de los valores observados y recta teórica calculada para la relación largo total (en mm) versus radio total de la escama (en mm) para los individuos del lago Rosario.
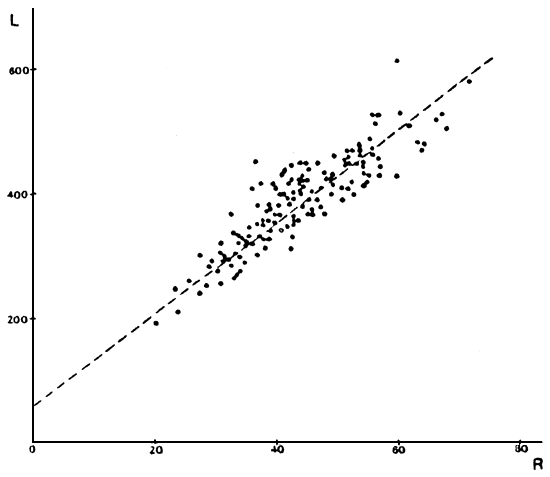
FIGURA 4: Gráfico de los valores observados y recta teórica calculada para la relación largo total (en mm) versus radio total de la escama (en mm) para los individuos de la laguna Esquel.
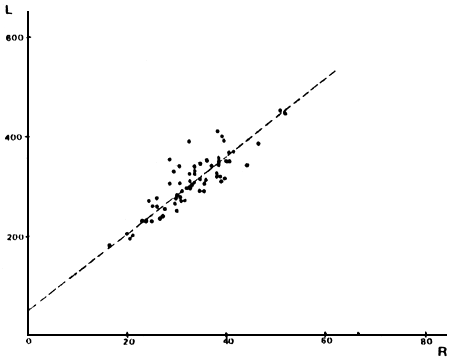
FIGURA 5: Gráfico de los valores observados y recta teórica calculada para la relación largo total (en mm) versus radio total de la escama (en mm) para los individuos de la laguna Brecham.
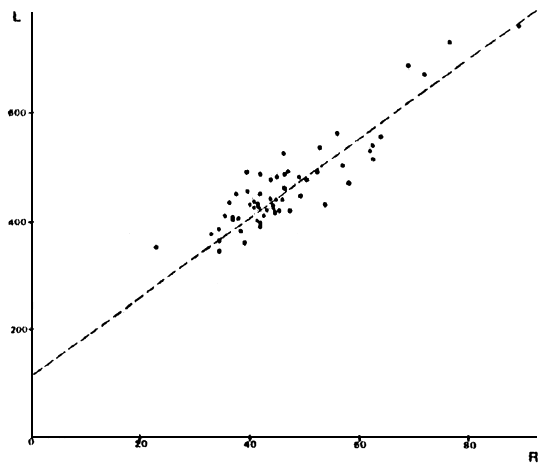
FIGURA 6: Gráfico de los valores observados y recta teórica calculada para la relación largo total (en mm) versus radio total de la escama (en mm) para los individuos de la laguna Zeta.
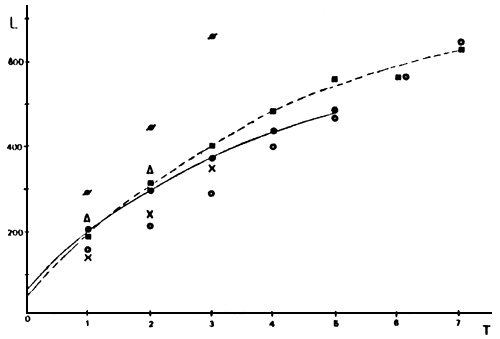
FIGURA 7: Tallas medias retrocalculadas (en mm), para cada edad y ambiente, y curvas de crecimiento de von Bertalanffy para el lago Musters (trazo continuo) y para la laguna Esquel (trazo punteado) [ Musters O ano en arroyo (□), Musters 1 ano en arroyo (○), Rosario ( X ), Esquel (•), Brecham (∆), Zeta (ø) ].
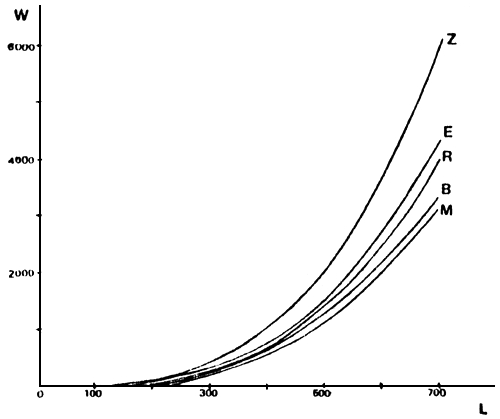
FIGURA 8: Relaciones largo total (en mm) versus peso total (en g) para cada ambiente. (Musters (M), Rosario (R), Esquel (E), Brecham (B), Zeta (Z)).
![]()
![]()
![]()