![]()
![]()
![]()
4.1. Vue d'ensemble des effets du CO2 sur les processus de croissance des plantes
4.2. Réponses spécifiques des cultures a une hausse du CO2
4.3. Résume de comptes rendus d'ensemble
4.4. La modélisation des cultures: Prédictions pour le futur
4.5. Résume et conclusions
Remerciements
Références
L. HARTWELL ALLEN, JR.
US Department of Agriculture, Agricultural Research Service, University of
Florida, Gainesville, Florida, USA
Jeff T. BAKER et Ken J. BOOTE
Département d'Agronomie, Université de Floride, Gainesville, Floride, USA
La hausse de la concentration en dioxyde de carbone (CO2) atmosphérique de l'ordre de 280 (m mol/mol avant la révolution industrielle à environ 360 m mol/mol actuellement est bien documentée (par ex., Baker et Enoch, 1983; Keeling et al., 1995). Le consensus de beaucoup d'études des effets d'une hausse du CO2 sur les plantes est que l'effet fertilisant du CO2 est réel (voir Kimball, 1983; Acock et Allen, 1985; Cure et Acock, 1986; Allen, 1990; Rozema et al., 1993; Allen, 1994; Allen et Amthor, 1995). Cependant, l'effet fertilisant du CO2 peut ne pas se manifester dans des conditions où un autre facteur de croissance est très limitant, tel qu'une basse température (Long, 1991). De plus, les plantes poussant dans certaines conditions où des limitations du volume des racines (Arp, 1991), de lumière, ou d'autres facteurs restreignent la croissance, n'ont pas montré une réponse soutenue à la hausse de CO2 (Kramer, 1981).
Les principaux objectifs de ce chapitre sont d'évaluer les effets directs de la hausse du CO2 atmosphérique et les effets indirects des changements potentiels de climat sur la croissance et le rendement des cultures. L'approche sera de (1) fournir une vue générale des effets du CO2 sur les processus de croissance des plantes; (2) analyser quelques données expérimentales spécifiques sur les réponses des plantes de culture à une hausse du CO2 et aux facteurs du changement de climat; (3) résumer les comptes rendus récents sur les réponses des plantes à la hausse du CO2; et (4) discuter quelques évaluations par modélisation des cultures de la hausse du CO2 et des facteurs du changement de climat sur la productivité agricole basées sur les prédictions des modèles de changement global du climat. En outre, on suggérera certaines adaptations pour améliorer la productivité des cultures dans un monde à teneur en CO2 plus élevée.
La plus grande partie de la discussion suivante à propos des effets du CO2 sur les plantes concerne les espèces au parcours photosynthétique C3 et pas nécessairement les espèces C4. D'autres facteurs aériens environnementaux non biotiques qui affectent la croissance et le développement des plantes sont la lumière et la température. En général, les régimes de photosynthèse des plantes croissent linéairement avec la lumière dans les gammes d'intensité lumineuse relativement basses et ensuite les régimes ralentissent jusqu'à atteindre un maximum asymptotique. A cause de la quantité et de l'ombrage dés nombreuses feuilles, le couvert végétal de la plupart des cultures n'atteint pas la saturation lumineuse en plein soleil; c'est dire qu'elles seraient capables de répondre à des niveaux de lumière bien supérieurs au plein rayonnement solaire. De même, les vitesses de photosynthèse des cultures répondent aux teneurs croissantes en CO2 puis ralentissent à des concentrations plus élevées (autour de 700 m mol/mol ou plus en fonction des espèces et autres facteurs). Cependant, la photosynthèse foliaire croît d'habitude avec la température jusqu'à une certaine valeur maximum et puis décroît. De plus, la température affecte non seulement la photosynthèse mais aussi la respiration, la croissance, les phases de développement et les processus de reproduction.
Une teneur élevée en CO2 peut avoir des effets sur la phénologie des cultures quoique les stades de développement sont principalement gouvernés par la température, le temps et la photopériode. Si les dates de plantation devaient être changées à cause de l'effet de serre, alors le calendrier phénologique des plantes pourrait être affecté. Par exemple, des températures plus élevées pourraient réduire les rendements en raccourcissant la durée de la période de remplissage des grains ou des modifications de la photopériode raccourciraient ou allongeraient le stade végétatif.
L'effet fertilisant du CO2 commence avec une fixation photosynthétique prononcée du CO2. Les hydrates de carbone non structurels tendent à s'accumuler dans les feuilles et autres organes de la plante sous forme d'amidon, d'hydrates de carbones solubles ou de polyfructose en fonction de l'espèce. Dans certains cas, il peut y avoir une inhibition en retour de la photosynthèse liée à l'accumulation d'hydrates de carbones non structurés. Une accumulation carbohydratée accrue, surtout dans les feuilles, peut être le signe de l'inadaptation de plantes cultivées sous enrichissement de CO2 à profiter complètement de celui-ci. Ceci peut être dû au fait que les plantes enrichies en CO2 n'ont pas une capacité de croissance adéquate (organes-puits) ou de chargement du phloème et de translocation des hydrates de carbone solubles. L'amélioration de l'utilisation de photoassimilats devrait être un but de conception des cultivars du futur (Hall et Allen, 1993).
Dans le processus de croissance, les photoassimilats sont destinés aux pousses végétatives, au système de racines ou aux organes reproductifs. Dans certains cas, une plus grande partie du photoassimilat de plantes enrichies en CO2 est allouée aux racines qu'aux pousses. Au-dessus du sol, plus de photoassimilats vont d'habitude aux branches et aux structures de soutien qu'aux feuilles. Ce phénomène peut ne pas être une réponse inhérente à une hausse de CO2 mais un sous-produit de la taille plus grande des plantes vivant en atmosphère enrichie en CO2, surtout parmi les espèces qui produisent des bouts de branche le long des branches principales (Allen et al., 1991).
La croissance de la biomasse reproductive et celle de la biomasse végétative sont d'habitude accrues par une hausse du CO2. Cependant, l'indice de récolte, ou le rapport entre le rendement en grains et le rendement en biomasse au-dessus du sol, est typiquement plus bas dans des conditions de CO2 élevé (Allen, 1991; Baker et al., 1989). Ceci peut aussi être une preuve de l'incapacité d'utiliser complètement une quantité plus abondante de photoassimilats.
Dans de nombreux cas, à la fois la quantité et l'activité de carboxylation de l'enzyme ribulose 1,5-biphosphate carboxylase-oxygenase (rubisco) diminuent dans les feuilles des plantes cultivées sous une teneur élevée en CO2. Ce phénomène d'acclimatation peut occasionner une dérégulation de la photosynthèse; cependant, ce n'est pas universel. Par exemple, il y a peu d'évidence de cette dérégulation chez le soja (Glycine max L.Merr.), une légumineuse C3. En fait, la capacité de photosynthèse par unité de surface foliaire du soja croît avec l'enrichissement en CO2. Souvent, les feuilles développent une couche supplémentaire de cellules du mésophylle. De plus, un hydrate de carbone plus structuré peut être produit dans les feuilles et dans les tiges des plantes enrichies en CO2.
On peut savoir pourquoi certaines plantes dérégulent et d'autres ne répondent pas à une hausse de CO2 en examinant les capacités du soja. Cette plante a (a) une fixation symbiotique de N2; (b) la capacité de former des couches supplémentaires de cellules palissades dans le tissu foliaire; (c) la capacité de dériver beaucoup de photoassimilats en amidon relativement inerte plutôt qu'en sucres solubles pendant la photosynthèse; (d) un "puits feuille-tige" assez fort au cours du développement végétatif; et (e) un fort "puits grain" pendant le développement reproductif. Les plantes qui n'ont pas ces capacités, soit de façon inhérente ou du fait de croître dans des environnements limitants, sont plus sujettes à un certain degré de dérégulation de la photosynthèse (Allen, 1994).
Le rapport carbone-azote des feuilles croît d'habitude avec une hausse du CO2. Les plantes peuvent s'acclimater à une teneur élevée en CO2 en exigeant moins de rubisco et d'appareil de photosynthèse, ce qui conduirait à des teneurs en azote plus faibles. La modification du rapport C/N est due en gros, à la fois, aux accroissements en hydrates de carbone structurels et non structurels et aux diminutions en protéines. Cependant, la teneur en azote du grain est peu affectée (Allen et al., 1988).
La vitesse spécifique de respiration peut être réduite aussi bien par une exposition à court terme à une hausse de CO2 qu'à une croissance à long terme sous une teneur élevée en CO2 (Amthor, 1995). Cependant, l'effet à long terme peut être similaire lorsque les vitesses de respiration sont calculées par unité d'azote.
Dans les scénarios du changement de climat, on s'attend à une augmentation des températures suite à la hausse du CO2 et des autres gaz à effet de serre. Des interactions entre le dioxyde de carbone et la température ont été observées dans la croissance végétative (c'est-à-dire que l'effet fertilisant du CO2 est plus grand quand il fait plus chaud que plus froid). Les hausses de température dans un monde enrichi en CO2 augmenteraient la productivité globale de biomasse des cultures végétatives (prairies et fourrages) à la fois par l'allongement de la saison de croissance en régions tempérées et par l'interaction CO2 x température dans la stimulation de la croissance végétative. Cependant, les interactions CO2 x température semblent très faibles ou négligeables dans les processus de reproduction (semence et grain) bien qu'il puisse y avoir plus de fleurs initiales formées sur un plus grand nombre de branches et de tiges stimulées par un enrichissement en CO2 (Baker et Allen, 1993a).
Des modifications dans les précipitations peuvent avoir lieu avec d'autres effets du changement de climat. En général, les prédictions des modèles culturaux montrent qu'une hausse de CO2 devrait accroître la productivité des plantes C3 mais celles associées à une hausse des températures seraient nuisibles. Il n'est pas surprenant que les modifications dans l'allure des précipitations (moins de pluie pendant la période de croissance) puissent être encore plus préjudiciables à la production des cultures que les changements de température.
Avec une hausse du CO2 la conductance stomatique de la plupart des espèces diminuera, ce qui peut réduire la transpiration par unité de surface foliaire. Cependant, l'indice foliaire de certaines cultures peut aussi croître. La réduction typique de 40% de la conductance stomatique induite par un doublement du CO2 n'a résulté, en général, qu'en une réduction de 10% (ou moins) de l'utilisation de l'eau par la culture dans des conditions expérimentales de laboratoire ou de terrain. Les modifications actuelles de l'évapotranspiration seront guidées par le bilan énergétique de la culture atténuée par la conductance stomatique, l'indice foliaire, la structure de la culture et tout changement de facteurs météorologiques.
L'efficience de l'utilisation de l'eau (WUE: rapport entre la fixation du CO2 et l'évapotranspiration) augmentera avec une hausse du CO2 Cet accroissement est causé plus par une photosynthèse accrue que par une réduction de la perte d'eau à travers des stomates partiellement fermés. Donc, il peut y avoir plus de biomasse produite par unité d'eau utilisée même si une culture exigeait encore presque autant d'eau du semis à la récolte. Cependant, si les températures croissent, la WUE accrue causée par l'effet fertilisant du CO2 pourrait diminuer ou s'annuler à moins que les dates de plantation ne soient déplacées à des saisons plus favorables.
Plusieurs évaluations des impacts du changement de climat sur la productivité des cultures ont été publiées. Le progrès réalisé consiste en une intégration des impacts sur des pays particuliers et des interactions économiques et sociales. Pour la plupart, ces évaluations prévoient des climats plus favorables à l'agriculture dans les latitudes nord et des climats moins favorables dans les zones tropicales et subtropicales. Cependant, les prédictions des modèles culturaux dépendent des résultats des scénarios des Modèles de Circulation Générale (GCMs) appliqués à l'effet de serre. La chaîne de dépendance des évaluations est la suivante: Scénarios de Changement de Climat ® Prédiction du Modèle Cultural et Systèmes de Production Agricole (avec ou sans stratégies de réponse d'atténuation et d'adaptation disponibles) ® Scénarios Nationaux sur l'Economie et le Bien-être des Agriculteurs, le Commerce Agricole et les Consommateurs ® Scénarios Pays par Pays et d'Interaction Globale de l'Echange Mondial (alimentation et toutes autres matières premières), de la Dynamique des populations et du Bien-être Economique ainsi que des Impacts sur les Systèmes Sociaux.
Aussi longtemps que le monde continue à consommer des combustibles fossiles, les concentrations en CO2 continueront de monter. Les autres gaz à effet de serre, tels que le méthane, les oxydes d'azote, les chlorofluorocarbones et leurs substituts, ainsi que l'ozone troposphérique monteront sans doute aussi. L'effet fertilisant du CO2 sur les plantes va croître et les changements de climat peuvent avoir lieu à cause de l'augmentation combinée de tous les gaz à effet de serre. L'agriculture globale s'adaptera aux changements graduels régionaux mais des changements soudains risquent d'être plus graves. Des actions d'adaptation et/ou d'atténuation pourraient inclure ce qui suit:
1. La sélection de plantes qui peuvent mieux utiliser les hydrates de carbone produits quand elles poussent à une teneur élevée en CO2.2. La sélection de plantes qui produisent moins de matière structurale et plus de capacité reproductive sous enrichissement en CO2. (Ceci s'applique aux plantes de culture de grains et pas nécessairement à celles produisant une biomasse végétative)
3. Recherche de gènes adaptés à des températures de jour et de nuit plus élevées et les incorporer dans les cultivars de cultures de production désirables pour améliorer la floraison et les semences.
4. Changer les dates de plantation et les autres pratiques culturales pour optimiser les rendements sous les nouvelles conditions climatiques, et sélectionner des cultivars qui sont adaptés à ces nouvelles pratiques.
5. Passer à des espèces qui ont une production plus stable sous des températures élevées ou la sécheresse.
6. Déterminer si des rapports C/N plus favorables peuvent être atteints dans des cultivars adaptés à une hausse du CO2.
7. Si nécessaire et si possible, développer des systèmes d'irrigation pour les cultures.
Cette section est consacrée aux réponses de deux cultures C3 à une hausse du CO2: le soja, une légumineuse fixatrice d'azote symbiotique représentative des impulsions; et le riz (Oryza sativa L.), une culture de céréale alimentaire globalement importante. Les résultats de sept cycles culturaux de soja et de riz cultivés en chambres éclairées par le soleil à Gainesville, Floride aux USA seront présentés (Baker et Allen, 1993a, b). Les chambres disposent de grands volumes de vrai sol pour l'enracinnement. Certaines études comprennent les réponses à des concentrations en CO2 inférieures ou supérieures à l'atmosphère (par ex., Allen et al., 1991; Baker et Allen, 1993a, b, 1994). (Les expériences sur le riz sont désignées chronologiquement par RICE I, II, III, IV, V, VI et VII.)
4.2.1. LE SOJA
Les flux photosynthétiques
Les flux photosynthétiques des feuilles et du couvert végétal total ont toujours été accrus par une hausse du CO2. Jones et al. (1984) ont signalé des flux photosynthétiques maximum du couvert à midi de 60 et 90 m mol CO2/m²/s, 70 jours après semis (JAS), pour du soja cultivé à des concentrations en m mol/mol correspondantes de 330 et 800 m mol/mol/mol. L'indice foliaire (LAI) était respectivement de 6.9 et 9.0. Les flux photosynthétiques maximum du couvert à midi étaient dans d'autres études de 40 et 75 m mol/mol CO2/m²/s pour le soja cultivé à 320 et 640 m mol/mol CO2 (Jones et al., 1985c) et, de 40 et 80 m mol/mol CO2/m²/s pour le soja cultivé à 330 et 800 m mol/mol CO2 (Jones et al., 1985a, b).
Valle et al. (1985) ont trouvé que les flux d'assimilation photosynthétique maximum du CO2 à midi pour les feuilles de soja variaient de 30 à 50 et de 15 à 25 m mol CO2/m²/s pour des plantes cultivées respectivement à 660 et 330 m mol/mol CO2. Allen et al. (1990) rapportent qu'à tous les niveaux de lumière, les flux photosynthétiques des feuilles croissent linéairement avec la concentration en CO2 dans la gamme de 330 à 800 m mol/mol.
Valle et al. (1985) ont utilisé une hyperbole rectangulaire du type Michaelis-Menten pour résumer les réponses photosynthétiques de feuilles de soja en fonction de la concentration en CO2. Les plantes avaient été cultivées à 330 et 660 m mol/mol de CO2 et, ensuite, exposées à une large gamme de CO2 pendant une courte période.
|
Y = (Ymax x [C])/([C]+ Km)+ Yi |
(4.1) |
où Y est le flux photosynthétique en m mol CO2/m²/s; [C] est la concentration en m mol/mol; Yi est l'intercept sur l'axe y à [C] égal zéro, le régime de respiration apparent en m mol CO2/m²/s; Ymax est la réponse limite de (Y- Yi) à très haute [C], le flux photosynthétique asymptotique en m mol CO2/m²/s; Km est la valeur de [C] où (Y- Yi) = Ymax/2, la constante apparente de Michaelis-Menten en m mol/mol; et G c est la [C] calculée à l'intercept Y égale zéro, le point de compensation de CO2 en m mol/mol (ne se trouve pas dans l'équation). Les paramètres moyens pour les réponses à 330 et 660 m mol/mol sont données au Tableau 4.1. Il n'y a pas eu de dérégulation évidente de la photosynthèse foliaire du soja en réponse à une teneur élevée en CO2 en fait, la capacité photosynthétique a augmenté. Le rendement en feuilles a augmenté de 0,05 à 0,09 pour le soja exposé respectivement à 330 et 660 m mol/mol de CO2 (Valle et al., 1985).
Campbell et al. (1988) ont montré que les flux photosynthétiques foliaires du soja étaient plus élevés pour les plantes cultivées à 660 qu'à 330 m mol/mol de CO2 lorsque la mesure était faite à des concentrations intercellulaires en CO2 communes. En outre, Campbell et al. (1988) ont mesuré l'activité et la quantité de la rubisco dans les feuilles de soja cultivé à des concentrations en CO2 de 160, 220, 280, 330, 660 et 990 m mol/mol. Ils ont trouvé que l'activité de la rubisco était constante à 1,0 m mol CO2/min/mg de protéine soluble dans toute cette gamme de traitement au CO2 La protéine foliaire soluble était proche d'une constante, 2,4 g/m2 avec 55% de rubisco. Le poids spécifique foliaire a crû dans la gamme de concentration en CO2 de 160 à 990 m mol/mol de sorte que l'activité de la rubisco sur base du poids foliaire sec décrût.
Campbell et al. (1990) ont montré que la capacité photosynthétique de couverts de soja cultivé à 330 et 660 m mol/mol de CO2 était, pour de courtes expositions, semblable à celle des concentrations inférieures à 500 m mol/mol mais, au-dessus de cette concentration, les couverts cultivés à 660 m mol/mol avaient une capacité photosynthétique légèrement supérieure. Donc, le soja ne perd pas sa capacité photosynthétique comme le font certaines autres espèces de plantes (voir Allen, 1994). Cependant, dans d'autres études, le flux photosynthétique et l'activité de la rubisco diminuent tous deux pendant l'enrichissement à long terme en CO2 (Thorne et Koller, 1974; Delucia et al., 1985).
La respiration
Les flux de respiration du couvert complet avant l'aurore 40 à 60 jours après semis étaient 2-3 et 4-5 (m mol/m²/s pour du soja cultivé respectivement à des niveaux de CO2 de 320 et 640 m mol/mol et à une température jour/nuit constante de 25°C (Jones et al, 1985c). Après avoir interchangé deux chambres de traitement 52 jours après semis, les flux photosynthétiques et de respiration se sont rapidement ajustés aux nouvelles conditions d'exposition au CO2. Donc, les flux de respiration avant l'aube étaient fort liés aux flux précédents de fixation du CO2.
Tableau 4.1. Flux photosynthétique asymptotique maximal moyen (Y, max) en rapport avec le paramètre (Yi) ou l'intercept de l'axe y, la constante apparente de Michaelis-Menten pour le CO2 (Km), et le point de compensation du CO2 (Fg) pour les feuilles qui se sont développées sous deux niveaux de traitement en CO2 et soumises à différents niveaux de concentration en CO2 pendant de courtes expositions (Valle et al., 1985)
|
Traitement de croissance |
Ymax |
Km |
Yi |
G c |
|
au CO2 |
m mol/m²/s |
m mol/mol |
m mol/m²/s |
m mol/mol |
|
330 |
51,8 |
359 |
-7,8 |
63 |
|
660 |
126,6 |
1133 |
-4,6 |
42 |
Les moyennes de Ymax et Yi sont signitivement différentes, p = 0.05, par le t-test.
La répartition
La croissance des plantes à une teneur élevée en CO2 résulte de modifications de la répartition dans le temps des photoassimilats vers les divers organes de la plante (Tableau 4.2). Chez le soja, une hausse du CO2 provoque en général une plus grande répartition du carbone (matière sèche) vers les structures de soutien (tiges, pétioles et racines) que vers les feuilles pendant les stades végétatifs de la croissance (Allen et al., 1991). Durant les stades reproductifs, il y a une tendance à une répartition relativement faible à la croissance reproductive (gousses) par les plantes sous atmosphère riche en CO2.
Les régimes de croissance
Pendant la phase linéaire de la croissance végétative et après que la couverture totale du sol fut réalisée, les régimes de croissance des plantes exposées à une gamme de concentrations en CO2 ont varié de 5,0 à 20,7g/m2/j pour des expositions de 160 à 990 (m mol/mol (Allen et al., 1991). Le poids sec total variait de 12,88 à 39,12 g/plante et le poids final de graines allait de 5,77 à 17,85 g/plante pour des traitements en CO2 de 160 à 660 (m mol/mol.
Tableau 4.2. Parties constituantes de la plante de soja exprimées en % du total de la matière sèche cultivées dans des conditions de concentrations du CO2 inférieures ou supérieures à la concentration à l'air libre en 1984. (Allen et al; 1991).
|
Constituants |
Concentration en CO2, m ml/mol |
|||||
|
(%) |
160 |
220 |
280 |
330 |
660 |
990 |
|
13 JAS1 (V2 Phase) |
||||||
|
Racine |
11,3 |
9,8 |
9,3 |
12,4 |
9,8 |
10,5 |
|
Cotylédon |
19,5 |
15,6 |
13,9 |
13,8 |
11,2 |
9,4 |
|
Tige |
22,8 |
25,0 |
25,8 |
25,5 |
26,3 |
26,7 |
|
Feuille |
46,1 |
49,5 |
50,7 |
48,2 |
52,5 |
53,2 |
|
34 JAS (V8 Phase) |
||||||
|
Racine |
6,4 |
7,1 |
6,9 |
7,6 |
8,9 |
7,9 |
|
Tige |
23,4 |
24,4 |
27,6 |
25,9 |
28,2 |
29,3 |
|
Pétiole |
12,2 |
13,1 |
14,6 |
14,4 |
15,7 |
15,7 |
|
Feuille |
57,6 |
55,3 |
50,8 |
52,2 |
47,2 |
46,9 |
|
66 JAS (R5 Phase) |
||||||
|
Tige |
16,5 |
19,6 |
25,0 |
21,7 |
25,2 |
26,8 |
|
Pétiole |
9,9 |
11,3 |
12,9 |
12,7 |
14,1 |
13,6 |
|
Feuille |
35,3 |
35,3 |
35,4 |
32,2 |
31,5 |
29,8 |
|
Gousse |
38,2 |
33,6 |
26,5 |
33,3 |
28,9 |
29,6 |
|
94 JAS (R7 Phase) |
||||||
|
Tige |
10,2 |
12,8 |
14,9 |
14,3 |
16,3 |
19,5 |
|
Pétiole |
5,5 |
6,2 |
6,0 |
6,7 |
7,0 |
6,5 |
|
Feuille |
17,4 |
16,3 |
14,9 |
14,4 |
11,9 |
10,9 |
|
Gousse |
66,7 |
64,7 |
64,1 |
64,6 |
64,7 |
64,0 |
¹Jours Après le Semis.
Les hydrates de carbone
Le soja accumule des hydrates de carbone non structurels, surtout l'amidon, sous enrichissement en CO2. Allen et al. (1988) ont fait pousser du soja avec des traitements en CO2 de 330, 450, 600 et 800 m mol/mol. La sucrose, les sucres réducteurs et les sucres solubles totaux des feuilles sont restés à peu près constants pendant la journée mais l'amidon a augmenté constamment à un rythme d'environ 6 g/kg de matière sèche/heure. La moyenne totale des sucres solubles a crû de 24 à 36 g/kg de poids sec et l'amidon de 85 à 204 g/kg de poids sec sous toute la gamme de 330 à 800 m mo/mol. Une teneur élevée en hydrates de carbone non structurels dans les plantes de soja enrichies en CO2 fut confirmée par Baker et al. (1989) et Allen et al. (1995). Les concentrations ont aussi varié pendant toute la vie des plantes.
L'azote
La teneur en azote a diminué de 50 à 37 g/kg de poids sec dans la gamme de 330 à 800 m mol/mol. Quand la teneur en azote fut ajustée pour annuler l'effet des hydrates de carbone non structurels totaux, les modifications relatives furent plus petites, 55 à 48g N/kg de matière sèche dans la gamme de 330 à 800 m mol/mol (Allen et al., 1988).
Le rendement
Le rendement en graines de soja a toujours crû suite à une hausse de CO2 Allen et al. (1987) ont résumé les réponses en rendements photosynthétiques, en biomasse et en graines de plusieurs expériences avec le modèle de l'hyperbole rectangulaire de l'équation (4.1) en utilisant des données normalisées par rapport aux réponses obtenues à 330 m mol/mol. Les valeurs des paramètres.. Km, Ymax et Yi, des flux photosynthétiques relatifs furent de 279 m mol/mol, 3,08 et - 0,68 respectivement pour des rendements relatifs en biomasse de 182 m mol/mol, 3,02 et - 0,91 et pour des rendements relatifs en graines de 141 m mol/mol, 2,55 et - 0,76. Ce modèle fut utilisé pour prévoir les rendements dans plusieurs gammes de concentrations accrues en CO2 atmosphérique (Tableau 4.3). Pour un doublement du CO2 ce modèle prédit une augmentation de 32,2% du rendement en graines de soja et de 42,7% en biomasse. Le rapport de ces deux nombres, 1,322/1,427=0,926, donne la fraction attendue de l'indice de récolte pour un doublement du CO2 en comparaison avec le CO2 ambiant.
Le dioxyde de carbone et la température
L'effet fertilisant du CO2 semble être stimulé à des températures élevées, du moins jusqu'à un certain point. Idso et al. (1987) et Kimball et al. (1993) ont montré que le facteur de modification de croissance (ou le rapport de modification de croissance de la biomasse) dû à un enrichissement de 300 m mol/mol était de 0,08 par °C (moyenne de température journalière) dans la gamme de 12 à 34 °C. Cependant, pour le soja, Allen (1991) a calculé un rapport de modification de croissance de la biomasse de -0,031 par °C pour le rendement de la biomasse des graines et de -0,026 par °C pour l'accumulation de biomasse totale. Des températures élevées tendent à raccourcir la période de remplissage des graines de cette culture.
Tableau 4.3. Accroissements en % des flux photosynthétiques du soja à midi, rendement en biomasse et en grain estimés à travers diverses gammes de concentration de dioxyde de carbone [CO2] associés à des points de repère dans le temps. (Allen et al., 1987)
|
Période de temps (années) |
[CO²]-Midi |
Photosynthèse de la biomasse |
Rendement en grains la |
Rendement de la biomasse |
|
|
Initiale |
Finale |
|
|||
|
(m mol/mol) |
(% d'accroissement au-dessus du niveau initial [CO2]) |
||||
|
AG-1700¹ |
200 |
270 |
38 |
33 |
24 |
|
1700-1973 |
270 |
330 |
19 |
16 |
12 |
|
1973-2073?2 |
330 |
660 |
50 |
41 |
31 |
¹ AG, L'Age Glaciaire soit il y a environ 13 000 à 30 000 ans. Durant le dernier Age Glaciaire et depuis la fin de la fonte glaciaire jusqu'aux périodes de la révolution pré-pionnière et pré-industrielle, les concentrations en CO2 dans l'atmosphère qui ont prévalu étaient de 200 et 270 m mol/mol.
2 Le premier choc pétrolier s'est produit en 1973 quand la concentration en CO2 était de 330 m mol/mol. Cette valeur est utilisée comme référence dans beaucoup d'études de doublement du CO2. Il est prévu que la concentration en CO2 va doubler tôt ou tard au 21ème siècle
Le rendement en graines du soja tend à décroître légèrement avec la température dans la gamme jour/nuit de 26/19 à 36/29 °C (Tableau 4.4). Le nombre de graines par plante croît légèrement avec une augmentation, à la fois, du CO2 et de la température. La masse par graine décroît nettement avec une hausse de température. Malgré que l'enrichissement en CO2 aboutit à un accroissement du rendement en graines et en biomasse au-dessus du sol, l'indice de récolte décrût à la fois avec le CO2 et la température (Baker et al., 1989). Les données du Tableau 4.4 n'indique aucune tendance à l'augmentation du facteur de modification de croissance avec la température ni pour le rendement en graines ni pour l'accumulation de biomasse.
Des expériences ultérieures avec le soja renforcent ces données quoiqu'on a trouvé que les rendements en graines chutaient très fort à des températures jour/nuit de 40/30 °C et au-dessus, et que les rendements en biomasse se maintenaient jusquà 44/34 °C avant de chuter rapidement (Deyun Pan et al., non publié).
La sensitivité des données de Idso et al. (1987) et Kimball et al. (1993) sur le facteur de modification de croissance peuvent avoir été affectés par d'autres facteurs à Phoenix, Arizona. Apparemment, les données de croissance vs température furent obtenues sur toute l'année et peuvent avoir été influencées par des facteurs tels que le rayonnement solaire total, la photopériode, les stades de développement ou d'autres conditions dues au changement de saisons. Bien sûr, ces autres facteurs environnementaux font tous partie du complexe de réponses des plantes aux conditions climatiques. Cependant, un meilleur test des effets de la température seule serait un enrichissement en CO2 pendant toute la saison ou le cycle de vie des plantes dans des conditions naturelles mais avec des écarts de températures consistants (plus froides et plus chaudes).
Tableau 4.4. Rendement en grains, composantes de rendement, total de la biomasse au-dessus du sol et indice de récolte du soja cultivé en 1987 à deux niveaux de concentrations du CO2. et à trois températures (Baker et al., 1989)
|
Concentration en CO2. ((m mol/mol) |
Jour/Nuit température (°C) |
Rendement en grains (g/plante) |
Grains/plante (no./plante) |
Poids des grains (mg/graine) |
Biomasse au-dessus du sol (g/plante) |
Indice de récolte |
|
330 |
26/19 |
9,0 |
44,7 |
202 |
17,1 |
0,53 |
|
330 |
36/29 |
10,1 |
58,9 |
172 |
22,2 |
0,45 |
|
660 |
26/19 |
13,1 |
58,8 |
223 |
26,6 |
0,49 |
|
660 |
31/24 |
12,5 |
63,2 |
198 |
27,6 |
0,45 |
|
660 |
36/29 |
11,6 |
70,1 |
165 |
26,5 |
0,44 |
|
F-Valeurs | ||||||
|
Concentration en CO2 |
12,3** |
11,4** |
2,5* |
ND |
ND | |
|
Température |
0,0 NS |
8,4** |
106,2** |
ND |
ND | |
|
CO2 x Température |
2,0 NS |
0,1 NS |
11,2** |
ND |
ND | |
*,** Significatif à des niveaux de probabilités de 0,05 et 0,01 respectivement.
NS = Non significatif; ND = Non disponible
L'évapotranspiration et l'efficience de l'utilisation de l'eau (WUE)
L'effet direct de l'accroissement des températures dans la gamme de 28 à 35°C semble être l'élévation du flux de transpiration de 4 à 5% par °C, basé sur des études expérimentales et de modélisation (Allen, 1991). Ceci est en bon accord avec la hausse de la pression de vapeur à saturation de l'ordre de 6% par °C. Allen et al. (1985) montrèrent que la hausse de la WUE du soja provoquée par une photosynthèse accrue due à une teneur élevée en CO2 était beaucoup plus grande que celle provoquée par une diminution de la transpiration. La part de l'accroissement en WUE attribuable à la hausse de la photosynthèse et à la baisse de la transpiration était respectivement de 0,8 et 0,2 (sur base des données de Jones et al., 1985b).
4.2.2. LE RIZ
La photosynthèse des feuilles et du couvert
Les flux photosynthétiques foliaires du riz cv.IR72, mesurés (à la température ambiante) en pleine lumière en 1992 (RICE VI) sur des plantes cultivées à trois régimes de températures jour/nuit, 32/23, 35/26 et 38/29°C, étaient en moyenne de 18,8 et 30,4 mmol CO2/m²/s respectivement pour des traitements de 330 et 660 m mo/mol CO2 (Allen et al., 1995). Cette augmentation causée par une hausse du CO2 est d'environ 60%. La courbe A/Ci obtenue pour les deux traitements aux températures de croissance de 35/26 °C est illustrée à la Figure 4.1. Il n'y a essentiellement pas de différence entre les flux d'assimilation, A, dans les deux traitements en CO2 à chaque niveau spécifique de CO2 intercellulaire, Ci. Des couples similaires de courbes (non présentées) furent obtenus pour les autres traitements de température. Chaque couple de courbes de A/Ci foliaires étaient semblables au sein de chaque régime de températures. Les courbes de A/Ci foliaires n'indiquent donc pas de modification de la capacité photosynthétique. L'extrapolation des courbes de réponse aux valeurs nulles de A donnent un intercept (point de compensation en CO2 foliaire) d'environ 60 m mol/mol CO2.
Après la fermeture du couvert végétal, le flux photosynthétique net du couvert du riz (Pn) vs les réponses de densité de flux de photons photosynthétiques (PPFD) était linéaire et n'approcha pas la saturation lumineuse dans aucune expérience, probablement à cause de l'orientation des feuilles érigées du cv. IR30 et de la forte densité de population végétale. Le Pn du couvert vs PPFD à 60 jours après plantation (JAP) pour les six traitements en CO2 de l'expérience RICE II (Baker et Allen, 1993b; Allen et al., 1995) donnait des valeurs d'environ 34, 50, 60, 80 et 90 mmol CO2/m²/s à fort éclairage (1600 mmol photons/m²/s) dans les traitements de 160, 250, 330, 500, 660 et 900 m mol/mol.
Les réponses linéaires de Pn du couvert au PPFD du jour 60 de l'expérience RICE IV furent similaires dans tous les traitements de températures (25/18, 28/21, 31/24, 34/27 et 37/30 °C) à 660 m mol/mol CO2; les réponses au traitement de 330 m mol/mol CO2 furent à peu près 25% moindres. D'autres études du Pn du couvert végétal n'ont montré que des petites différences sur de grandes gammes de températures; par ex., le coton (Baker et al., 1972) et le soja (Jones et al., 1985a). Il peut y avoir deux raisons pour le manque de réponse photosynthétique d'un couvert clairsemé à la température de l'air dans la gamme de 25 à 37°C. Le refroidissement évaporatif peut abaisser la température foliaire en-dessous de la température de l'air au fur et à mesure de l'augmentation de la température de l'air et du déficit de pression de vapeur (Allen, 1990; Pickering et al., 1995). La réponse cumulée des flux photosynthétiques foliaires à la température à toutes les expositions de lumière peut élargir la réponse à la température des flux photosynthétiques du couvert entier (Pickering et al., 1995).
Baker et Allen (1993b) ont introduit les données de jours entiers de Pn du couvert végétal dans une hyperbole du type de l'équation (4.1) 61 jours après semis dans l'expérience RICE II. Les paramètres étaient respectivement 70,8 m mol/mol, 3,96 et -2,21 pour Km, Ymax, et Yi, avec une réponse relative de Pn plafonnant à 1,75 à [C] infinie. Le pourcentage d'augmentation calculé en réponse à Pn de jours entiers du couvert à 660 vs 330 m mol/mol était de 36%.
Acclimatation photosynthétique au CO2 au niveau du couvert végétal
Pour tester la capacité photosynthétique d'acclimatation du couvert végétal, Baker et al. (1990b) ont utilisé du riz cultivé à 160, 250, 330, 500, 660 et 900 m mol/mol CO2. Pendant des périodes d'un demi-jour (les matins) des jours 62, 63 et 64 après plantation, des concentrations communes de CO2 de 160, 330 ou 660 m mol/mol furent imposées dans chaque chambre à ces jours respectifs. En comparant ces expositions à court terme au CO2, Baker et al. (1990b, 1996b) et Baker et Allen (1993a) ont montré que le flux photosynthétique net à court terme du couvert diminue avec la hausse du traitement en CO2 à long terme (Figure 4.2). Les effets relatifs de l'exposition à court terme au CO2 étaient plus grands pour la concentration en CO2 à court terme la plus basse. Par exemple, lorsque comparé à une exposition commune à court terme de CO2 de 160 m mol/mol, le flux photosynthétique du couvert de la chambre soumise au traitement de croissance à long terme de 900 m mo/mol CO2 n'était que d'un tiers de celui du traitement de croissance à long terme de 160 m mol/mol CO2 (Figure 4.2). Cependant, une bonne partie de l'effet de cette acclimatation apparente peut être attribuée aux flux respiratoires plus grands des plantes cultivées à une teneur élevée en CO2 (Boote et al., 1994).
Le pourcentage en protéine rubisco permit de vérifier l'acclimatation foliaire dans une large gamme de concentrations en CO2 (Baker et Allen, 1994; Allen et al., 1995) comme montré au Tableau 4.5 au jour 34 après semis pour le soja (Campbell et al., 1988) et au jour 75 après plantation pour le riz (Rowland-Bamford et al., 1991). Pour le riz, l'activité rubisco exprimée sur base de la surface foliaire diminua de 66% dans la gamme de 160 à 900 m mol/mol des traitements en CO2 à long terme (Rowland-Bamford et al., 1991). Une cause majeure de ce déclin de l'activité rubisco fut une chute de 32% de la quantité de protéine rubisco par rapport aux autres protéines solubles (Rowland-Bamford et al., 1991).
Quoique Rowland-Bamford et al. (1991) signalent des diminutions de la quantité et de l'activité du rubisco avec la hausse en CO2 dans les plants de riz cultivés dans la gamme de 160 à 900 m mol/mol, les courbes de réponse foliaire A/Ci n'indiquent pas une chute de la capacité photosynthétique d'acclimatation dans la gamme plus restreinte de 330 à 660 m mol/mol CO2. Une possibilité serait que le rubisco n'est pas aussi limitant qu'attendu pour la photosynthèse sous enrichissement en CO2. On a besoin de plus de travaux sous traitements de CO2 pour explorer les effets d'interaction de la capacité puits, de l'alimentation en azote et d'autres processus internes de fixation du CO2. sur le comportement photosynthétique et le rendement des cultures.
Tableau 4.5. Pour le soja, la protéine foliaire soluble est exprimée sur base de la surface de la feuille et le pourcentage en protéine rubisco exprimé sur base de protéine foliaire soluble d'une plante de soja âgée de 34 jours cultivée sous une large gamme de concentrations en CO2 (Campbell et al., 1988) et (Baker and Allen, 1994). Pour le riz, la teneur en azote foliaire est exprimée sur base de la surface foliaire et le pourcentage en protéine rubisco défini sur base de protéine foliaire soluble d'une plante de riz âgée de 75 jours cultivée sous une large gamme de concentrations en CO2 (Rowland-Bamford et al., 1991; Baker and Allen, 1994).
|
Soja |
Riz | ||||
|
Concentration en CO2 pendant la croissance (m mol/mol) |
Protéine foliaire soluble (g/m2) |
Protéine rubisco (%) |
Concentration en CO2 pendant la croissance (m mol/mol) |
Protéine azotée foliaire (m mol/m2) |
Protéine rubisco (%) |
|
160 |
2,5 |
56 |
160 |
95 |
62 |
|
220 |
3,2 |
54 |
250 |
90 |
59 |
|
280 |
2,6 |
- |
330 |
81 |
54 |
|
330 |
2,3 |
57 |
500 |
62 |
49 |
|
660 |
2,3 |
54 |
660 |
78 |
43 |
|
990 |
2,3 |
55 |
900 |
64 |
42 |
Les stades de développement
Chez le riz deux régimes différents d'apparition des feuilles furent aussi trouvées dans les phases végétative et reproductive de croissance (Baker et al., 1990, 1996; Baker et Allen, 1994) comme observées précédemment (Yoshida, 1977; Vergara, 1980). Le régime d'apparition foliaire (feuilles par jour) est à peu près deux fois plus grand dans la phase végétative que dans la phase reproductive.
Dans l'expérience RICE II de plantation 'tardive' (23 juin 1987), le nombre de feuilles sur les branches principales à l'initiation du panicule et le nombre final de feuilles sur les branches principales décroissent pour les traitements en CO2 de 160 à 500 (m mol/mol et restent similaires de 500 à 900 (m mol/mol. L'initiation du panicule et le stade de la gaine ont lieu environ 12 jours plus tôt avec une ambiance plus concentrée en CO2 dans le traitement à 160 m mol/mol (Baker et al, 1990a; Baker et Allen, 1993b). Donc, le rythme du développement de la plante fut clairement accéléré par une hausse du CO2 jusqu'à environ 500 (m mol/mol.
Les données sur l'effet de la température furent réunis dans les traitements des essais RICE 1 et RICE V allant de 25/18/21 à 40/33/37 °C (températures jour/nuit de l'eau du paddy). La température de l'air affecte fortement la vitesse d'apparition foliaire (Figure 4.3-A), le rythme du développement et la durée totale de croissance alors que les effets étaient comparativement petits dans la gamme limitée de 330 à 660 m mol/mol (Baker et Allen, 1993b; Baker et al., 1992a, b, 1995). On n'a pas observé de différences consistantes dans l'intervalle de foliation (jours par apparition de feuille) entre 330 à 660 m mol/mol tandis que cet intervalle a augmenté avec la hausse de température dans la gamme 25/18/21 à 40/33/37 °C, surtout dans la phase reproductive (Figure 4.3-B). Dans les essais RICE III, IV et V, l'anthèse arrive de 0 à 6 jours plus tôt dans les essais de 660 vs 330 (m mol/mol (Baker et al., 1995). De plus, le nombre de jours jusqu'à la floraison fut réduit d'à peu près 10 jours dans la gamme 25/18/21 à 40/33/37 °C de l'essai RICE IV.
Croissance et rendement
Le rendement et les données composant le rendement des essais RICE 1 et RICE II pour une gamme de concentrations en CO2 de 160 à 900 (m mol/mol sont présentées au Tableau 4.6. Une hyperbole rectangulaire (équation (4.1)) fut ajustée aux données de rendement en grain. Des valeurs de Km= 284 m mol/mol, Ymax = 2,24 et Yi=-0,13 furent obtenues pour un rendement relatif en grain dont le plafond asymptotique (Ymax+ Yi) = 2,11 à une [C]
Dans une gamme relativement grande de températures allant de 25/18/21 à 37/30/34°C, Baker et al. (19920) trouvèrent un large optimum de températures pour la production de biomasse dans les gammes de températures moyennes. Les plantes cultivées à 40/33/37°C étaient près de la limite supérieure de survie. La stérilité des épillets de riz à haute température est induite presqu'exclusivement le jour de l'anthèse (Satake et Yoshida, 1978) quand des températures atteignant plus de 35 °C pendant plus d'une heure induisent un haut pourcentage de stérilité (Yoshida, 1981). Dans les traitements à 40/33/37°C, les plantes dans les chambres à 330 m mol/mol mourrurent pendant l'élongation internodale tandis que celles à 660 m mol/mol produisirent des petits panicules mal formés qui étaient stériles (Baker et al., 1992a). Donc, une teneur élevée en CO2 permet d'accroître légèrement le maximum de température auquel les plantes de riz peuvent survivre.
Aux deux niveaux de CO2, le rendement en grain fut le plus haut dans le traitement 28/21/25°C suivi par une chute à zéro dans le traitement 40/33/37°C (Tableau 4.7). L'enrichissement en CO2 de 330 à 660 m mol/mol accrût le rendement par augmentation du nombre de panicules par plant tandis que le nombre de grains pleins par épi et la masse individuelle des grains étaient moins influencés. Les effets de la température sur le rendement et ses composantes furent hautement significatifs. Le nombre de panicules par plant augmenta tandis que le nombre de grains pleins par épi chuta fortement avec la hausse de la température. La masse des grains individuels fut stable aux températures modérées mais a eu tendance à diminuer au-delà de 34/27/31°C. La biomasse finale au-dessus du sol et l'indice de récolte augmentèrent avec l'enrichissement en CO2 tandis que l'indice de récolte chuta fortement avec la hausse de la température. Notamment, il n'y a pas eu d'effet significatif d'interaction CO2 x température sur le rendement, ses composantes ou la biomasse finale au-dessus du sol (Tableau 4.7).
A chaque concentration en CO2, des équations polynomiales de régression furent ajustées aux données de rendement en grain du riz (Y) vs température (X) du Tableau 4.7. Une polynomiale du 3ème degré, Y = -239,226 + 25,557* X - 0,848* X2 + 0,00896* X3 (R2 = 0,91, Sy,x = 1,36), donna le meilleur ajustement pour les traitements de 660 m mol/mol CO2 (Baker et Allen, 1993a). Cependant, une polynomiale du 2ème degré, Y = -1,196 + 1,042* X - 0,0278* X2 (R2 =0,91, Sy,x = 1,36), s'ajuste mieux aux données des traitements de 330 (m mol/mol. A partir d'un nombre d'essais repris dans les tableaux et figures de l'article de Baker et Allen (1993a), l'effet de la température sur le rendement en grains et la biomasse finale est illustré respectivement aux Figures 4.4 et 4.5. Alors que les futures augmentations en CO2 atmosphérique devraient favoriser les rendements du riz, des effets fortement négatifs sont vraisemblables si les températures montent aussi. Les figures montrent que la productivité végétative se maintient à plus hautes températures que la croissance reproductive.
Les flux de respiration nocturne du couvert
Les flux de respiration des plantes peuvent décroître à cause d'expositions à court et long termes à des concentrations élevées en CO2 (Bunce, 1990; Amthor, 1991); cependant, Baker et al. (1992c) trouvèrent que les flux de respiration nocturne du couvert (Rd, mmol CO2/m2 de sol/s) croissent pour du riz exposé pendant le jour à des teneurs en CO2 allant de 160 à 900 m mol/mol. Similaires aux flux photosynthétiques, les Rd augmentèrent avec des expositions au CO2 allant de 160 à 500 m mol/mol mais plafonnèrent quelque peu dans la gamme de 500 à 900 m mol/mol. Les Rd des traitements en CO2 ambiant et superambiant ont atteint un large maximum 30 à 50 jours après plantation tandis que le maximum des traitements subambiants avaient lieu plus tard aux environs de 50 à 70 JAP. Les valeurs maximum de Rd furent à peu près de 6, 8 et 9 mmol/m²s pour les traitements en CO2 de 160, 250 et 330 (m mol/mol et d'environ 11 à 12 m mol/mol/m²s pour les trois traitements superambiants en CO2.
Le flux spécifique de respiration (Rdw, m mol/mol CO2/s/kg de matière séche totale au-dessus du sol) diminua exponentiellement avec les JAP pour toutes les expositions au CO2 et fut plus élevé dans les traitements subambiants (160 et 250 m mol/mol) que dans ceux ambiants (330 (m mol/mol) et superambiants (550, 660 et 900 m mol/mol). A chaque niveau d'exposition au CO2, les allures des Rdw en fonction des JAP furent très similaires à celles de la concentration en azote des tissus végétaux avec JAP (Baker et al., 1992c). De plus, Rdw était linéairement lié à la concentration totale en azote des tissus végétaux au-dessus du sol (mg de N/g de poids sec) dans toute la gamme d'expositions au CO2 et les six dates d'échantillonnage (Rdw = -13,0 + 0,952* (N); R2 =0,91, P = 0,001). Baker et al. (1992c) concluèrent que les traitements de CO2 affectent Rdw principalement en altérant la teneur en protéine du tissu végétal. Une autre explication est qu'une teneur élevée en CO2 accroît la quantité d'hydrates de carbone structurels et non structurels dans les tissus de la plante en sorte qu'une plus grande partie de la matière sèche est bloquée dans les matériaux non protéiniques. Cette étude n'a pas montré d'acclimatation de la respiration à l'enrichissement à long terme en CO2 (acclimatation indirecte) du riz qui ne pourrait pas être expliquée par la concentration en azote des tissus de la plante.
Tableau 4.6. Rendement en grains, composantes du rendement, biomasse totale des parties aériennes et indices de récolte du riz dans les essais menés en 1987 à des niveaux de concentration en CO2 supérieurs ou inférieurs à la concentration de CO2 à l'air libre (Baker et al., 1988, Baker et Allen 1993b, 1994, Baker et al. 1995, 1996).
|
CO2 |
Température |
Rendement en grains |
Panicule/plante |
Grains pleins |
Masse des grains |
Biomasse |
Indice de récolte |
Essai RICE No. |
|
m mol/mol |
°C |
Mg/ha |
no./plante |
no./panicule |
mg/grain |
g/plante |
|
|
|
160 |
31/31/27 |
3,4c2 |
3,6c |
24,8a |
17,0a |
4,0c |
0,36a |
I&II |
|
250 |
31/31/27 |
4,1c |
4,8bc |
20,8a |
18,2a |
5,1bc |
0,34a |
I&II |
|
330 |
31/31/27 |
4,8bc |
5,7ab |
21,0a |
17,6a |
6,3ab |
0,34a |
I&II |
|
500 |
31/31/27 |
6,83 |
7,3 |
23,0 |
18,1 |
9,8 |
0,30 |
II uniquement |
|
660 |
31/31/27 |
6,6ab |
6,5ab |
25,0a |
17,8a |
8,4a |
0,35a4 |
I&II |
|
900 |
31/31/27 |
7,3a |
7,4a4 |
24,8a |
17,8a |
8,2a |
0,39a |
I&II |
¹ Température diurne de l'air/Température nocturne de l'air/Température de l'eau de rizière.
2 Les valeurs suivies d'une même lettre dans une même colonne ne sont pas différentes de manière significative après le Test de Gamme Multiple de Duncan (p = 0,05).
³Les plantes de l'essai RICE 1 pour un traitement en CO2 de 500 m mol/mol ont subi un test de toxicité hydro-sulfide dans la zone racinaire ce qui a empêché le remplissage des grains et réduit le rendement en grains aux niveaux inférieurs à ceux mesures dans des conditions de traitements en CO2 inférieures à la concentration de l'atmosphère. Ces données n'ont pas été utilisées dans cette analyse. Les valeurs de traitements de la ligne 500 m mol/mol de CO2 correspondent uniquement aux résultats de l'essai RICE II.
4 Indique une différence significative entre les dates de plantation telle que déterminée par le t-test à un niveau de confiance de 0, 05.
Tableau 4.7. Rendement en grain, composantes du rendement, biomasse finale de la partie aérienne et indice de récolte de cinq essais différents menés sur le riz. D'après Baker et Allen (1993a) et Baker et al. (1996)
|
CO2 |
Température |
Température2 moyenne de l'air |
Rendement en grain |
Panicule/plante |
grains pleins |
Masse de grains |
Biomasse |
Indice de récolte |
NO. Essai |
|
(m mol/mol |
°C |
°C |
Mg/ha |
no./plante |
no./panicule |
mg/graine |
g/plante |
|
|
|
330 |
28/21/25 |
24,2 |
7,9 |
5,1 |
34,5 |
17,4 |
7,3 |
0,47 |
RICE III |
|
28/21/25 |
25,1 |
6,6 |
3,9 |
39,6 |
17,5 |
6,5 |
0,44 |
RICE IV |
|
|
28/21/25 |
25,1 |
8,0 |
4,0 |
47,5 |
18,5 |
8,1 |
0,43 |
RICE V |
|
|
31/31/27 |
31,0 |
5,2 |
5,9 |
23,0 |
17,1 |
5,5 |
0,42 |
RICE I |
|
|
31/31/27 |
31,0 |
4,3 |
5,4 |
19,0 |
17,9 |
7,2 |
0,26 |
RICE II |
|
|
34/27/31 |
30,2 |
4,2 |
7,7 |
15,2 |
16,2 |
5,6 |
0,43 |
RICE III |
|
|
40/33/37 |
36,2 |
0,0 |
-,- |
-,- |
-,- |
-,- |
-,- |
RICE III |
|
|
660 |
25/18/21 |
22,1 |
8,4 |
4,4 |
46,0 |
18,2 |
7,8 |
0,47 |
RICE IV |
|
28/21/25 |
24,2 |
8,4 |
5,0 |
37,7 |
18,0 |
7,9 |
0,46 |
RICE III |
|
|
28/21/25 |
25,1 |
10,4 |
4,2 |
58,3 |
18,3 |
8,9 |
0,50 |
RICE IV |
|
|
28/21/25 |
25,1 |
10,1 |
4,4 |
54,2 |
19,0 |
9,3 |
0,47 |
RICE V |
|
|
31/31/27 |
31,0 |
6,8 |
6,9 |
25,1 |
17,2 |
7,5 |
0,40 |
RICE I |
|
|
31/31/27 |
31,0 |
6,4 |
6,0 |
24,8 |
18,4 |
9,3 |
0,29 |
RICE 11 |
|
|
4/27/31 |
30,2 |
4,8 |
6,5 |
18,5 |
16,7 |
6,3 |
0,32 |
RICE III |
|
|
34/27/31 |
31,1 |
3,4 |
7,5 |
12,9 |
16,3 |
8,1 |
0,18 |
RICE IV |
|
|
37/30/34 |
34,1 |
1,0 |
8,0 |
3,0 |
14,2 |
7,1 |
0,06 |
RICE IV |
|
|
40/33/37 |
36,2 |
0,0 |
-,- |
-,- |
-,- |
-,- |
-,- |
RICE III |
|
|
F-valeurs |
|||||||||
|
CO2 |
4,2* |
4,3* |
0,4 NS |
0,1 NS |
13,1** |
4,6* |
|
||
|
Température |
51,8** |
22,6** |
51,1** |
27,0** |
3,2* |
32,8** |
|
||
|
CO2 x température |
1,9 NS |
1,7 NS |
1.3 NS |
0,3 NS |
0,2 NS |
1,9 NS |
|
||
¹Température diurne de l'air/température nocturne de l'air/température de l'eau de rizière.
2 La température moyenne de l'air est la moyenne entre la température du jour et celle de la nuit ajustée pour une thermopériode
** * Significatif à un niveau de probabilité de 0,01 et 0,05 respectivement. NS = Non Significatif
Figure 4.4. Le rendement en grains du riz en fonction de la moyenne pondérée de la température jour/nuit de l'air pour les plantes cultivées jusqu'à maturité à des concentrations en CO2 de 330 et 660 (m mol/mol dans cinq essais différents

Figure 4.5. L'ajustement polynomial du second degré du rendement final en biomasse de riz en fonction de la température moyenne de l'air pondérée jour/nuit pour des plantes cultivées à des teneurs en CO2 de 330 et 660 m mol/mol jusqu'à maturité dans cinq essais différents.

L'évapotranspiration et l'efficience de l'utilisation de l'eau
A la fois, la transpiration de la plante et l'évaporation directe de la surface inondée contribuent à l'utilisation de l'eau (évapotranspiration, ET) par le riz cultivé en chambres SPAR. Les tendances diurnes de ET suivirent le rayonnement solaire (Baker et al., 1990b; Allen et al., 1995). Après la fermeture du couvert, les flux maximum de ET et les pertes diurnes totales d'eau furent environ 35 et 30% plus grands respectivement, pour le riz cultivé à 160 (m mol/mol que pour celui à 900 (m mol/mol (Tableau 4.8). Les flux de ET furent similaires quand toutes les chambres étaient exposées pendant un demi jour à la même concentration en CO2 (données non présentées, Baker et al., 1990b), ce qui démontre l'effet du niveau d'exposition du CO2 sur le contrôle stomatique de la transpiration.
La température a aussi un grand effet sur les flux diurnes de ET (Baker et Allen, 1993a; Allen et al., 1995) et l'utilisation diurne de l'eau (Tableau 4.8) lorsque le déficit de pression de vapeur de l'air intervient principalement. Le rayonnement solaire a aussi un grand effet, probablement, à la fois, par les entrées d'énergie dans le couvert végétal et par l'ouverture stomatique en réponse à la lumière (directement ou indirectement). Les flux maximum de ET à midi furent environ 75 et 35% plus élevés chez le riz cultivé à 40/33/37°C et 34/27/31°C respectivement, comparé à celui à 28/21/25°C (Baker et Allen, 1993a; Allen et al., 1995).
L'efficience de l'utilisation de l'eau (WUE): les effets du CO2
Les totaux diurnes d'assimilation du CO2 l'ET et les chiffres de WUE vs traitement en CO2 sont montrés au Tableau 4.8. La conductance stomatique décroît avec l'augmentation de la concentration en CO2 qui peut causer une réduction de la transpiration et des feuilles ainsi que de tout le couvert. Cependant, l'enrichissement en CO2 peut aussi accroître la surface foliaire du couvert pour la transpiration, ce qui déséquilibre un peu l'économie en eau (Jones et al., 1985B; Allen et al., 1985). L'indice foliaire de l'essai RICE II variait de 7, 6 à 10, 8 dans les traitements en CO2 de 160 à 900 m mol/mol. Les chiffres du WUE ont augmenté avec le CO2 (Tableau 4.8) à cause de la chute de ET dans toute la gamme de CO2 et de la hausse de Pn avec le CO2 jusqu'à 500 (m mol/mol.
L'efficience de l'utilisation de l'eau (WUE): effets de la température
A une température de l'air de 28°C, l'enrichissement en CO2 de 330 à 660 m mol/mol a résulté en un gain de 30% dans l'assimilation diurne totale du CO2 et en une perte de 22% en ET avec une augmentation concommittante de WUE de 1, 42 à 2, 37 mmol (CO2)/mmol (H2O), soit un gain de 67% (Tableau 4.8). L'ET de jour a plus que doublé de 25 à 37°C. La WUE de jour a chuté de 28 à 37°C à cause de la forte augmentation de l'ET et de Pn relativement stable dans cette gamme de températures. Si WUE était basée sur le rendement en grain, elle aurait chuté considérablement avec la hausse de température non seulement à cause de la hausse en ET mais aussi de la forte diminution de la production de grains.
Tableau 4.8. Comparaison des réponses diurnes totales du couvert végétal du riz soumis pendant toute la saison à diverses concentrations en CO2 (RICE II, 23 Août 1987, 61 jours après plantation) et à différents régimes de températures (RICE IV, 10 Septembre 1989, 58 jours après plantation). Les traitements à température de l'air du jour/température de l'air de nuit/température de l'eau de rizière/température du point de rosée sont donnés dans la deuxième colonne. D'après Baker et al. (1990b) et Baker et Allen (1993a, b, 1994).
|
CO2 |
Température1 |
Flux diurne total de photons |
Quantité diurne de CO2 prélevé |
Perte diurne totale de H2O |
Efficience2 à l'utilisation de l'eau |
|
m mol/mol |
°C |
mol (Photons)/m2 |
mol (CO2)/m2 |
mol (H2O)/m2 |
m mol/mol |
|
Effets du dioxyde de carbone: RICE II, 23 Août 1987, 61 JAP | |||||
|
160 |
31/31/27: 18 |
39,4 |
0,74 |
608,5 |
1,22 |
|
250 |
31/31/27: 18 |
|
1,13 |
643,4 |
1,76 |
|
330 |
31/31/27: 18 |
|
1,34 |
611,0 |
2,19 |
|
500 |
31/31/27: 18 |
|
1,80 |
553,6 |
3,25 |
|
660 |
31/31/27: 18 |
|
1,79 |
536,2 |
3,34 |
|
900 |
31/31/27: 18 |
|
1,83 |
469,5 |
3,90 |
|
Effets de la température: RICE IV, 10 Septembre 1989, 58 JAP | |||||
|
330 |
28/21/25: 12, 0 |
26,4 |
0,87 |
612,7 |
1,42 |
|
660 |
25/18/21: 10,5 |
|
0,83 |
359,3 |
2,30 |
|
660 |
28/21/25: 12,0 |
|
1,13 |
475,8 |
2,37 |
|
660 |
31/24/28: 13,5 |
|
0,83 |
491,3 |
1,69 |
|
660 |
34/27/31: 15,0 |
|
0,89 |
736,4 |
1,20 |
|
660 |
37/30/34: 16,5 |
|
0,97 |
909,4 |
1,06 |
¹ Température de l'air le jour/Température de l'air la nuit/température de l'eau de rizière: température du point de rosée
2 mmol (CO2)/mol (H2O). JAP: Jour Après Plantation
La hausse en CO2 atmosphérique joue probablement en faveur de la production de riz par l'augmentation de la photosynthèse, de la croissance et du rendement en grain tout en réduisant l'utilisation de l'eau qu'en augmentant la WUE. Dans les régions chaudes du monde, cependant, le réchauffement global futur possible peut résulter en des pertes de rendements substantielles à cause de la sensibilité de la floraison et de l'épiaison aux températures élevées et les pénuries en eau possibles qui peuvent résulter de l'augmentation de l'évapotranspiration.
Les comptes rendus et revues de plusieurs symposiums récents laissent peu de doute sur le fait que les plantes de cultures peuvent bien répondre à une hausse du CO2 (Rozema et al., 1993; Woodwell et Mackensie, 1995; Wittwer, 1995). Poorter (1993) a rassemblé l'information sur 156 espèces de plantes et a trouvé qu'un doublement du CO2 produirait un gain moyen de croissance de 37%. La distribution des rapports de poids des plantes enrichies en CO2 et de celles de contrôle est illustrée à la Figure 4.6. La compilation de Poorter montre un gain respectif de 41 et de 22% pour les plantes C3 et C4. Comme groupe, les plantes de cultures herbacées C3 répondent plus que les espèces herbacées sauvages (58 vs 35%). De plus, les espèces sauvages à croissance rapide répondent plus fortement que celles à croissance lente (54 vs 23%).
Poorter (1993) a mis deux restrictions à sa synthèse qui peuvent avoir conduit à des réponses plus larges qu'attendues à la hausse du CO2. D'abord, les plantes poussant en compétition ne sont pas comprises. Deuxièmement, seuls les stades végétatifs des plantes furent comparés puisque les données furent sélectionnées avant la floraison.
Plusieurs études ont montré que les réponses du stade végétatif peuvent être plus grandes que celles du stade reproductif (rendement en grain). Par conséquent, les données compilées par Poorter peuvent donner l'impression d'une plus grande réponse que si l'observation avait été faite sur tout le cycle de vie. Deuxièmement, les cultures au champ poussent d'habitude en populations denses où elles luttent pour l'espace et la lumière. Dans des conditions plus réelles du champ, les plantes de culture répondent vraisemblablement comme une communauté que comme des plantes individuelles, la lumière (rayonnement solaire) devenant ainsi un facteur limitant de croissance. Dans ces conditions, une hausse du CO2 ne peut favoriser l'expansion latérale ni une meilleure prise de lumière. Quoique les réponses réelles au champ peuvent être moindres, l'effet fertilisant du CO2 est clairement bien établi.
L'effet fertilisant du CO2 des espèces forestières est aussi devenu bien établi. Wulischleger et al. (1994) ont estimé le facteur de croissance biotique d'espèces forestières dans 58 études d'exposition controlée comprenant 398 observations. Leur distribution de fréquence de la réponse relative de croissance d'arbres soumis à une teneur en CO2 élevée vs CO2 ambiante est donnée à la Figure 4.7. Ils trouvèrent aussi un rapport moyen de réponse de 1,32. Des 398 observations, 51 montrèrent une réponse relative de croissance inférieure à 1, et 31 des réponses supérieures à 2. Cependant, dans des conditions de compétition, la réponse des arbres peut être bien moindre (Bazzaz et al., 1995).
Figure 4.6. Distribution du facteur de modification de croissance de la biomasse (rapport pondéral) pour les plantes enrichies en CO2 (600 à 700) m mol/mol en comparaison avec des traitements de contrôle (300 à 360 (m mol/mol). L'histogramme a été construit à partir des moyennes de tous les rapports pondéraux de 156 espèces sélectionnées dans littérature. Source: Poorter (1993).

Kimball et al. (1993) ont discuté les données de Idso et al. (1987) qui relatèrent le facteur de modification de croissance (ou le facteur de modification de croissance de la biomasse) causé par une hausse de 300 (m mol/mol de concentration en CO2 au-dessus de celle ambiante. Leurs données sont reproduites à la Figure 4.8. Dans les conditions de Phoenix, Arizona, le facteur de modification de croissance dans une gamme de température moyenne de 12 à 34°C fut assez linéaire. Donc, la réponse végétative au CO2 devrait être favorisée à des températures accrues. Comme discuté précédemment, le facteur de modification de croissance peut ne pas être d'application aux réponses de croissance reproductives (rendement en grain) des cultures (riz).
Figure 4.7. Distribution de fréquence du facteur de modification de croissance de la biomasse transformé en logarithme (log10 du rapport des masses totales sèches après exposition à des teneurs en CO2 élevées/ambiantes) de 73 espèces d'arbres. Source: Wullschleger et al.(1994)

4.4.1. MODELISATION DES REPONSES DES CULTURES AUX CHANGEMENTS EN CO2 ET DE CLIMAT
Peart et al. (1989) et Curry et al. (1990a, b, 1995) ont prédit des réponses de croissance et de rendement du soja et du maïs à des scénarios de changement de climat sous un doublement du CO2 dans le sud-est des USA. Leurs simulations utilisèrent 30 ans de données météorologiques de base (1951-1980) sur 19 sites dans 11 états. Les changements de climat prédits par le modèle (Hansen et al., 1988) de l'Institut Goddard d'études spatiales (GISS) et par le modèle (Manabe et Wetherald, 1987) du Laboratoire Géophysique de Dynamique des Fluides (GFDL) furent utilisés pour changer mois par mois les températures, précipitation et rayonnement solaire des 30 années de données de chaque site. Ces données de base modifiées fournirent les scénarios de changement de climat GISS et GFDL (Smith et Tirpak, 1989).
Figure 4.8. Facteur de modification de croissance de la biomasse (facteur de modification de croissance ou augmentation relative dans la croissance de la biomasse) résultant d'un accroissement de teneur en CO2 de 300 m mol/mol au-dessus de la teneur ambiante (presque doublée) en fonction de la température moyenne journalière de l'air pour les plantes indiquées dans la légende. Source: Kimbal et al. (1993)

A chaque site, les principaux types de sol, dates de plantation et cultivars furent utilisés dans les simulations.
Les données de précipitation et de température de base et des scénarions GISS et GFDL utilisées par Peart et al. (1989) furent condensées pour deux localisations représentatives; Columbia en Caroline du Sud et Memphis au Tennessee (Tableau 4.9). Les réponses du rendement des cultures au changement de climat furent simulées dans quatre conditions: avec ou sans effet fertilisant direct du CO2 et pour des cultures pluviales ou irriguées de façon optimum. L'équation de Penman-Monteith, qui contient un terme pour la conductance du couvert végétal, fut utilisée pour calculer les effets d'une hausse du CO2 sur la transpiration du couvert. Les simulations ont été faites pour un doublement du CO2 avec un facteur de stimulation photosynthétique de 1, 35 pour le soja (une plante C3) et 1, 10 pour le maïs (une plante C4). Il n'y eut pas de simulations avec les seuls effets fertilisants du CO2 (sans effets du changement de climat).
Les rendements en grain simulés du soja furent moyennés sur 30 ans et 19 sites (Tableau 4.10). Pour les cultures pluviales avec les effets du changement de climat seulement, les rendements simulés du soja avec le scénario GDFL furent réduits de 71% comparés au climat de base alors que les rendements moyens n'étaient réduits que de 23% avec le scénario GISS.
Les rendements du scénario GFDL furent sévèrement influencés à cause des réductions de pluie prédites par ce modèle (Tableau 4.9).
Tableau 4.9. Résumé de la moyenne des données météorologiques sur une période de base de 30 ans (1951-1980) (BASE) de deux sites avec les scénarios GISS et GFDL. Résumé de Peart et al. (1989)
|
|
BASE |
GISS |
GFDL |
||||||
|
Préc. (mm) |
Tmax (°C) |
Tmin (°C) |
Préc. (mm) |
Tmax (°C) |
Tmin (°C) |
Préc. (mm) |
Tmax (°C) |
Tmin (°C) |
|
|
Columbia, SC |
|||||||||
|
AVR-SEP; Juil |
684 |
33,3 |
21,2 |
817. |
35,2 |
23,1 |
471 |
38,2 |
26,1 |
|
OCT-MAR; Jan |
561 |
13,5 |
0,7 |
602 |
15,8 |
3,0 |
571 |
15,6 |
2,8 |
|
TOTAL; Moy. |
1245 |
24,1 |
10,7 |
1419 |
26,6 |
13,2 |
1042 |
27,3 |
13,9 |
|
Memphis. TN |
|||||||||
|
AVR-SEP; Juil |
656 |
33,1 |
22,6 |
748 |
35,4 |
24,9 |
549 |
36,0 |
25,5 |
|
OCT-MAR; Jan |
654 |
9,0 |
-0,6 |
565 |
12,0 |
2,4 |
758 |
11,3 |
1,7 |
|
TOTAL; Moy. |
1310 |
22,0 |
11,1 |
1313 |
25,4 |
14,5 |
1307 |
24,8 |
13,9 |
Dans les conditions optimales d'irrigation, les rendements moyens avec les deux scénarios GDFL et GISS furent réduits d'environ 18 à 19%. Cependant, malgré des températures plus hautes, les rendements en irrigué avec les scénarios GDFL et GISS furent environ 25% plus grands que ceux avec le climat de base sans irrigation.
Quand les effets fertilisants du CO2 étaient inclus avec les effets du changement de climat (Tableau 4.10), les rendements moyens simulés du scénario du climat GISS crûrent de 11 % sous conditions de pluie alors que les rendements avec le scénario du climat GFDL étaient encore plus bas (-52%). Avec une irrigation optimale et les effets fertilisants du CO2, les rendements des deux scénarios GISS et GFDL ont crû environ 13 à 14%.
Les rendements du maïs furent simulés par des effets fertilisants d'un doublement du CO2 à quatre sites de station météorologique seulement (Charlotte en Caroline du nord; Macon en Géorgie; Memphis au Tennessee; Meridian au Mississippi). Pour les seuls effets de changement du climat, les rendements prédits en maïs chutèrent seulement de 6% dans le scénario GISS mais de 73% dans le scénario GFDL (Tableau 4.11). Quoique l'irrigation accroît les rendements prédits des cultures, les scénarios GISS et GFDL donnèrent des chutes de rendement respectives de 18 et 27% par rapport au scénario de base avec irrigation. La réduction de rendement du scénario GFDL par rapport à GISS était de 10%, attribuable à des températures légèrement plus hautes. L'ajout des effets fertilisants du CO2 aux scénarios de changement du climat a eu peu d'effet sur les rendements prédits du maïs parce que c'est une plante C4.
Tableau 4.10. Simulations du rendement de soja (SOYGRO) pour un doublement du CO2 dans le Sud-Est des Etats Unis.
|
BASE |
Modèle GISS |
Modèle GFDL |
Modèle | ||
|
Rendement (kg/ha) |
Rendement (kg/ha) |
Diff.% |
Rendement (kg/ha) |
Diff% |
Diff% |
|
Effets du Changement de Climat uniquement, pluvial | |||||
|
2497 |
1929 |
-23 |
733 |
-71 |
-62 |
|
Effets du Changement de Climat uniquement, irriguée | |||||
|
3837 |
3158 |
-18 |
3092 |
-19 |
-2 |
|
Fertilisation par le CO2 plus Effets du Changement de Climat, pluvial | |||||
|
2497 |
2780 |
+ 11 |
1206 |
-52 |
-57 |
|
Fertilisation par le CO2 plus Effets du Changement de Climat, irriguée | |||||
|
3837 |
4350 |
+ 13 |
4393 |
+ 14 |
+ 1 |
Source: Peart et al. (1989).
Dans les Grands Lacs et la région du Corn Belt, les simulations par Ritchie et al. (1989) montrèrent qu'une plus haute température serait le facteur de changement du climat qui aurait le plus grand effet sur les rendements prédits du soja et du maïs, surtout par la réduction de leurs cycles de vie. Pour la partie méridionale de cette région, les réductions de rendement furent plus grandes avec le scénario GFDL. Les rendements prédits croissent dans les stations plus nordiques parce que les températures et la durée de la saison de croissance deviennent plus favorables. Les besoins moyens en eau d'irrigation dans cette région ont crû environ 90% dans les scénarios GISS et GFDL.
Rosenzweig (1989) a modélisé les rendements du maïs et du blé dans les Grandes Plaines avec les scénarios GISS et GFDL. Les rendements diminuèrent dans les Grandes Plaines du Sud à cause de températures plus élevées qui réduisent le cycle de vie des cultures. Là où la pluie prédite diminuait, les besoins d'irrigation augmentaient. Dans une étude de modélisation séparée, Allen et Gichuki (1989) prédirent une hausse de 15% du besoin en irrigation dans cette région, avec des besoins plus grands pour l'alfalfa parce que sa saison de croissance était allongée, et des besoins plus bas pour le maïs et le blé d'hiver parce que leurs saisons de croissance étaient diminuées. L'effet fertilisant du CO2 compensait les effets adverses du changement de climat en certains des endroits étudiés par Ritchie et al. (1989) et par Rosenzweig (1989).
Tableau 4.11. Simulations du rendement de maïs pour doublement du CO2 (CERES-Maize) à Charlotte NC, Macon GA, Meridian MS et Memphis TN
|
NORMALE |
Modèle GISS |
Modèle GFDL |
Modèle | ||
|
Rendement (kg/ha) |
Rendement (kg/ha) |
Diff.% |
Rendement (kg/ha) |
Diff %. |
Diff%. |
|
Effets du Changement de Climat uniquement, pluvial | |||||
|
8468 |
7926 |
-6 |
2289 |
-73 |
-71 |
|
Effets du Changement de Climat uniquement, irrigué | |||||
|
13899 |
11455 |
-18 |
10257 |
-26 |
-10 |
|
Fertilisation par le CO2 plus les Effets du Changement de Climat, pluvial | |||||
|
8577 |
8136 |
-5 |
2224 |
-74 |
-73 |
|
Fertilisation par le CO2 plus les Effets du Changement de Climat, irrigué | |||||
|
14052 |
11545 |
-18 |
10363 |
-26 |
-10 |
Source: Peart et al. (1989).
Dudek (1989) prédit des changements de productivité de plusieurs cultures de légumes, de fruits et de noix en Californie dans les scénarios GISS et GFDL avec un doublement du CO2. Sans les effets fertilisants du CO2, les changements moyens de rendement dans tout l'état seraient de -8 à -34% pour le scénario GISS et de -6 à -31% pour le GFDL, en fonction de la culture. Avec les effets fertilisants du CO2, ces changements de rendement variaient d'environ + 17 à - 12% pour le scénario GISS et de + 21 à -8% pour le GFDL.
Dudek (1989) a utilisé le California Agriculture and Resource Model (CARM) pour prédire aussi l'impact des changements de productivité sur l'économie et le marché. En général, la production chutait dans les scénarios du changement de climat sans effet fertilisant du CO2; cependant, les prix des matières premières augmentaient. Dans les scénarios avec effets fertilisants du CO2 et du changement de climat, les impacts sur les prix étaient bien moindres.
Les effets de la température sur la phénologie des plantes de culture jouent un rôle critique. Il y a un besoin crucial pour une recherche plus détaillée sur les réponses des plantes à la température et aux interactions température x CO2 comme données d'entrée aux modèles de cultures. Il faut aussi plus d'études de modélisation sur les dates de plantation (ou de semis) comme stratégie d'adaptation. Des cultivars doivent être conçus pour des conditions climatiques futures (Hall et Allen, 1993). Les facteurs qu'il faudrait considérer sont: l'extension de la période de remplissage des grains et peut-être la diminution de la durée de la période végétative (ce qui améliorerait aussi l'indice de récolte); une floraison et une épiaison à plus hautes températures; des interactions adaptatives des photo- et thermopériodes; une sélection pour une acclimatation photosynthètique positive là où une acclimatation photosynthétique négative a été observée; et une capacité d'utiliser plus efficacement les photoassimilats (stockage des hydrates de carbone). Ces facteurs doivent être intégrés dans la physiologie de la plante entière sous conditions réelles.
4.4.2. ADAPTATIONS ET EXIGENCES DE L'EVAPOTRANSPIRATION
Les réponses simulées du rendement des cultures aux changements climatiques fournies par Peart et al. (1989) et Curry et al. (1990a, b, 1995) révèlent deux points principaux: (a) l'impact très néfaste des scénarios de pluies insuffisantes couplés à ceux d'une montée des températures sur la production des cultures, et (b) l'importance bénéfique des effets fertilisants du CO2 face aux températures élevées. Cependant, le changement de climat correspondant à un doublement effectif du CO2 peut avoir lieu à des concentrations en CO2 moindres que celles utilisées en simulation si des traces d'autres gaz actifs sur le rayonnement jouent un grand rôle dans l'effet de serre. Dans ce cas, les effets directs du CO2 seraient un peu inférieurs à ceux donnés en exemples aux Tableaux 4.10 et 4.11 pour un changement de climat équivalent. Toutes les simulations supposent que les changements climatiques auraient lieu en même temps qu'un accroissement des concentrations en CO2 et autres gaz à effet de serre. Si le réchauffement global suit la hausse du CO2 atmosphérique, alors il y aura vraisemblablement des effets fertilisants bénéfiques du CO2 avant que l'impact total du changement de climat ne se révèle. Cependant, Broecker (1987) et d'autres préviennent que les changements de climat n'ont pas toujours été graduels au cours des périodes interglaciaires du Pléistocène. Il y a des preuves d'oscillations climatiques relativement rapides dans l'hémisphère nord pendant l'interglaciaire précédente, 110.000 à 140.000 ans d'ici, dans les enregistrements du Greenland Ice-core Project (GRIP) (Anklin et al., 1993). Ces oscillations ont provoqué des périodes froides aussi dures que la période glaciaire précédente.
La diminution des rendements en soja rapportée par Peart et al. (1989) et Curry et al. (1990a, b, 1995) était causée en grande partie par le raccourcissement de la période de remplissage des grains à des températures plus hautes. Des modifications de pratiques culturales telles que les dates de semis ou la sélection d'autres cultivars, pourrait empêcher un peu les réductions potentielles de rendement. Dans l'avenir, les reproducteurs de plantes peuvent devoir combiner une tolérance à la température et des réponses à la photopériode dans de nouveaux germoplasmes. Dans les situations où les hydrates de carbone non structurels s'accumulent en réponse à l'effet fertilisant du CO2, de nouveaux germoplasmes devraient être développés pour faire meilleur usage de la source de photoassimilats.
L'irrigation ne sera probablement pas une panacée pour le changement de climat. D'après les simulations de Peart et al. (1989) et Curry et al. (1990a, b), les besoins prédits en irrigation du soja dans le sud-est des USA augmenteraient respectivement de 33 et 134% avec les scénarios GISS et GFDL. Cependant, dans le scénario GFDL, les ressources en eau deviendraient rares et ne seraient pas facilement disponibles pour les cultures. Certaines régions des USA devraient s'adapter à irriguer moins de terres. La hausse des températures et la baisse des précipitations aux USA prédites par le modèle GFDL auraient un impact global fort négatif sur la productivité agricole quoique des producteurs dans des régions favorables pourraient bénéficier de prix plus élevés dus à la rareté (Adams et al., 1990).
4.4.3. EVALUATION DES IMPACTS INTERNATIONAUX DE LA HAUSSE DU CO2 ET DU CHANGEMENT DE CLIMAT
Plusieurs évaluations ont été menées sur les impacts de la hausse du CO2 et des changements climatiques globaux sur les tendances de production des cultures et les réponses économiques dans des zones nationales ou régionales (Adams et al., 1990; Crosson, 1993; Parry et al., 1988; Smit et Tirpark, 1989) et dans divers pays du monde (Rosenzweig et Iglesias, 1994; Rosenzweig et al., 1995). De plus, un progrès a été réalisé dans la prédiction des impacts du changement de climat en utilisant des modèles d'échange mondial (Rosenzweig et Parry, 1993, 1994).
Les études conduites par Rosenzweig et ses collègues ont été faites avec des modèles de cultures existants et des scénarios de changement de climat de trois GCMs (GISS, GFDL et le modèle du United Kingdom Meteorological Office (UKMO), Wilson et Mitchell, 1987) pour prédire les modifications de rendement des cultures de plusieurs pays du monde. Les scénarios du doublement du CO2 (changements de température, précipitation et évaporation) étaient basés sur le changement de climat potentiel attendu des hausses de tous les gaz à effet de serre. Ces changements de climat arriveraient bien avant que la concentration en CO2 n'ait en fait doublé. Par conséquent, les teneurs en CO2 pour obtenir des effets de fertilisation furent estimés être de 555 m mol/mol au lieu de 660 m mol/mol. D'où, les rapports photosynthétiques de l'effet fertilisant du CO2 du soja, du blé, du riz et du maïs furent pour ces quatre cultures respectives de 1,21, 1,17, 1,17 et 1,06.
L'impact des scénarios de changement du climat fut plus sévère aux latitudes tropicales qu'aux latitudes moyennes ou élevées. Par exemple, les modifications de production moyenne prédite par les scénarios des trois GCMs au Brésil, aux Indes, en Chine et au Canada furent, sans effet du CO2, de -47, -43, -11 et -20% respectivement, et de -28, -13,+ 8 et + 16% avec les effets fertilisants du CO2 (calculés à partir des données de Rosenzweig et Parry, 1993; Rozenzweig et Iglesias, 1994),
Les simulations de rendements du soja à travers les USA faisaient partie de cette étude internationale (Curry et al., 1995). Ils trouvèrent que les rendements d'ensemble étaient de 2,42, 2,80, 2,37 et 1,31 Mg/ha pour les scénarios respectifs BASE, GISS, GFDL et UKMO. Les températures étaient de l'ordre de 4°C plus élevées que BASE pour les scénarios GISS et GFDL mais de 5,2°C pour le modèle UKMO.
Des perfectionnements et améliorations continueront à se faire dans la méthodologie de prédiction mais ces évaluations donnent le meilleur aperçu actuellement disponible des effets fertilisants du CO2 et du changement de climat sur la productivité globale des cultures.
Une teneur élevée en CO2 accroît la taille et le poids sec de la plupart des plants C3 et de leurs composantes. Les photoassimilats sont relativement plus répartis dans les composants structuraux (les tiges et pétioles) pendant le développement végétatif pour soutenir l'appareil capteur de lumière (les feuilles). L'indice de récolte a tendance à diminuer avec l'augmentation de la concentration en CO2 et de la température. Une sélection des plantes qui pourraient répartir plus de photoassimilats à la croissance reproductive devrait être un but de recherche future. Au fur et à mesure qu'on apprend plus au sujet des effets des changements anticipés du climat sur les cultures, plus d'efforts devraient être orientés vers l'exploration des adaptations biologiques et des systèmes de gestion pour réduire ces impacts sur l'agriculture et l'humanité. Il reste à voir si les climats régionaux deviendront plus secs ou plus humides suite au réchauffement global.
Ce travail fut mené en coopération avec l'Université de Floride à Gainesville et supporté en partie par le US Department of Energy dans le cadre des Accords Interagences DE-AI02-93ER61720, DE-AI05-88ER60001 ainsi que par la US EPA dans le cadre de l'Accord Interagences DW12934099 avec l'Agricultural Research Service du US Department of Agriculture. Florida Agricultural Experiment Station Journal Series No. R-00000.
Acock, B. and Allen, L.H. Jr. 1985. Crop responses to elevated carbon dioxide concentration. In: Direct Effects of Increasing Carbon Dioxide on Vegetation. DOE/ER-0238. B.R. Strain and J.D. Cure (eds.). US Dept. of Energy, Carbon Dioxide Res. Div., Washington DC. pp. 53-97.
Adams, R.M., Rosenzweig, C., Peart, R.M., Ritchie, J.T., McCarl, B.A., Glyer, J.D., Curry, R.B., Jones, J.W., Boote, K.J. and Allen, L.H. Jr. 1990. Global climate change and US agriculture. Nature 345: 219-224.
Allen, L.H. Jr. 1990. Plant responses to rising carbon dioxide and potential inter-actions with air pollutants. J. Environ. Qual. 19: 15-34.
Allen, L.H. Jr. 1991. Effects of increasing carbon dioxide levels and climate change on plant growth, evapotranspiration, and water resources. In: Proceedings of a Colloquium on Managing Water Resources in the West Under Conditions of Climatic Uncertainty. 14-16 Nov. 1990, Scottsdale, AZ. National Research Council, National Academy Press, Washington DC. pp. 101-147.
Allen, L.H. Jr. 1994. Carbon dioxide increase: Direct impacts on crops and indirect effects mediated through anticipated climatic changes. In: Physiology and Determination of Crop Yield. K.J. Boote, J.M. Bennett, T.R. Sinclair and G.M. Paulsen (eds.). American Society of Agronomy, Crop Science Society of America, and Soil Science Society of America, Madison, Wisconsin, pp. 425-49.
Allen, L.H. Jr. and Amthor, J.S. 1995. Plant physiological responses to elevated CO;, temperature, air pollution, and UV-B radiation. In: Biotic Feedbacks in the Global Climatic System: Will the Warming Increase the Warming? G.M. Woodwell and F.T. Mackenzie (eds.). Oxford University Press, New York. pp. 51-84.
Allen, R.G. and Gichuki, F.N. 1989. Effects of projected CO2-induced climatic changes on irrigation water requirements in the Great Plains States (Texas, Oklahoma, Kansas, and Nebraska). In: The Potential Effects of Global Climate Change on the United States. Appendix C, Agriculture, Vol. 1. EPA-230-05-89-053. J.B. Smith and D.A. Tirpak (eds.). US Environmental Protection Agency, Washington DC. pp. 6-1 to 6-42.
Allen, L.H. Jr., Jones, P. and Jones, J.W. 1985. Rising atmospheric CO2 and evapotranspiration. In: Advances in Evapotranspiration. ASAE Pub. 14-85. American Society of Agricultural Engineers, St. Joseph, Michigan, pp. 13-27.
Allen, L.H. Jr., Boote, K.J., Jones, J.W., Jones, P.H., Valle, R.R., Acock, B., Rogers, H.H. and Dahlman, R.C. 1987. Response of vegetation to rising carbon dioxide: Photosynthesis, biomass, and seed yield of soybean. Global BiogeochemicalCycles 1: 1-14.
Allen, L.H. Jr., Vu, J.C.V., Valle, R.R., Boote, K.J. and Jones, P.H. 1988. Nonstructural carbohydrates and nitrogen of soybean grown under carbon dioxide enrichment. Crop Sci. 28: 84-94.
Allen, L.H. Jr., Valle, R.R., Mishoe, J.W., Jones, J.W. and Jones, P.H. 1990. Soybean leaf gas exchange responses to CO2 enrichment. Soil Crop Sci. Soc. Fla. Proc. 49: 192-198.
Allen, L.H. Jr., Bisbal, B.C., Boote, K.J. and Jones, P.H. 1991. Soybean dry matter allocation under subambient and superambient levels of carbon dioxide. Agron. J. 83: 875-883.
Allen, L.H. Jr., Baker, J.T., Albrecht, S.L., Boote, K.J., Pan, D. and Vu, J.C.V. 1995. Carbon dioxide and temperature effects on rice. In: Climate Change and Rice. S. Peng, K.T. Ingram, H.-U. Neue, and L.H. Ziska (eds.). Springer-Verlag, Berlin, Heidelberg, pp. 256-277.
Amthor, J.S. 1991. Respiration in a future, higher-CO2 world. Plant Cell Environ. 14: 13-20.
Amthor, J.S. 1995. Plant respiration responses to elevated partial CO2 pressures. In: Advances in Carbon Dioxide Effects Research. L.H. Allen, Jr., M.B. Kirkham, C. Whitman and D.M. Olzyk (eds.). Special Pub., American Society of Agronomy, Madison, Wisconsin.
Anklin, M., and thirty-nine other Greenland Ice-core Project (GRIP) members. 1993. Climate instability during the last interglacial period recorded in the GRIP ice core. Nature 364: 203-207.
Arp, W.J. 1991. Effects of source-sink relations on photosynthetic acclimation to elevated CO2. Plant Cell Environ. 14: 869-875.
Baker, J.T. and Allen, L.H. Jr. 1993a. Contrasting crop species responses to CO2 and temperature: Rice, soybean, and citrus. Vegetatio 104/105: 239-260. Also: pp. 239-260. In: CO2 and Biosphere. (Advances in Vegetation Science 14). J. Rozema, H. Lambers, S.C. van de Geijn and M.L. Cambridge (eds.). Kluwer Academic Publishers, Dordrecht.
Baker, J.T. and Allen, L.H. Jr. 1993b. Effects of CO2 and temperature on rice: A summary of five growing seasons. J. Agric. Meteorol. (Japan) 48: 575-582.
Baker, J.T. and Allen, L.H. Jr. 1994. Assessment of the impact of rising carbon dioxide and other potential climate changes on vegetation. Environ. Pollut. 83: 223-235.
Baker, D.N. and Enoch, H.Z. 1983. Plant growth and development. In: CO2 and Plants. E.R. Lemon (ed.). Westview Press, Boulder, Colorado, pp. 107-130.
Baker, D.N., Hesketh, J.D. and Duncan, W.G. 1972. Simulation of growth and yield in cotton: I. Gross photosynthesis, respiration, and growth. Crop Sci. 12: 431-435.
Baker, J.T., Allen, L.H. Jr., Boote, K.J., Rowland-Bamford, A.J., Jones, J.W., Jones, P.H., Bowes, G. and Albrecht, S.L. 1988. Response of vegetation to carbon dioxide, Ser. No. 043. Response of rice to subambient and superambient carbon dioxide concentrations, 1986-1987. Report of the US Dept. of Agriculture, Agricultural Research Service (in cooperation with the University of Florida) for the US Dept. of Energy, Carbon Dioxide Research Division, Office of Energy Research, Washington DC.
Baker, J.T., Allen, L.H. Jr., Boote, K.J., Jones, P. and Jones, J.W. 1989. Response of soybean to air temperature and carbon dioxide concentration. Crop Sci. 29: 98-105.
Baker, J.T., Allen, L.H. Jr., Boote, K.J., Jones, J.W. and Jones, P. 1990a. Developmental responses of rice to photoperiod and carbon dioxide concentration. Agric. For. Meteorol. 50: 201-210.
Baker, J.T., Allen, L.H. Jr., Boote, K.J., Jones, P. and Jones, J.W. 1990b. Rice photosynthesis and evapotranspiration in subambient, ambient, and superambient carbon dioxide concentrations. Agron. J. 82: 834-840.
Baker, J.T., Allen, L.H. Jr. and Boote, K.J. 1992a. Response of rice to CO2 and temperature. Agric. For. Meteorol. 60: 153-166.
Baker, J.T., Allen, L.H. Jr. and Boote, K.J. 1992b. Temperature effects on rice at elevated CO2 concentration. J. Exp. Bot. 43: 959-964.
Baker, J.T., Laugel, F., Boote, K.J. and Allen, L.H. Jr. 1992c. Effects of daytime carbon dioxide concentration on dark respiration of rice. Plant Cell Environ. 15: 231-239.
Baker, J.T., Allen, L.H. Jr. and Boote, K.J. 1995. Potential climate change effects on rice: Carbon dioxide and temperature. In: Climate Change and Agriculture: Analysis of Potential International Impacts. C. Rosenzweig, L.A. Harper, S.E. Hollinger, J.W. Jones and L.H. Allen, Jr. (eds.). ASA Special Pub. No. 59, American Society of Agronomy, Madison, Wisconsin, pp. 31-47.
Baker, J.T., Allen, L.H. Jr., Boote, K.J. and Pickering, N.B. 1996. Assessment of rice responses to global climate change: CO2 and temperature. In: Terrestrial Ecosystem Response to Elevated CO2. G.W. Koch and H.A. Mooney (eds.). Physiological Ecology Series, Academic Press, San Diego, pp. 265-282.
Bazzaz, F.A., Jasienski, M., Thomas, S.C. and Wayne, P. 1995. Microevolutionary responses in experimental populations of plants in CO2-enriched environments. Parallel results from two model systems. Proc. of the National Academy of Sciences S.92: 8161-8165.
Boote, K.J., Pickering, N.B., Baker, J.T. and Allen, L.H. Jr. 1994. Modelling leaf and canopy photosynthesis of rice in response to carbon dioxide. International Rice Research Notes 19: 47-48.
Broecker, W.S. 1987. Unpleasant surprises in the greenhouse? Nature 328: 123-126.
Bunce, J.A. 1990. Short- and long-term inhibition of respiratory carbon dioxide efflux by elevated carbon dioxide. Ann. Bot. 65: 637-642.
Campbell, W.J., Allen, L.H. Jr. and Bowes, G. 1988. Effects of CO2 concentration on rubisco activity, amount, and photosynthesis in soybean leaves. Plant Physiol. 88: 1310-1316.
Campbell, W.J., Allen, L.H. Jr. and Bowes, G. 1990. Response of soybean canopy photosynthesis to CO2, light, and temperature. J. Exp. Bot. 41: 427-433.
Crosson, P. 1993. Impacts of climate change on the agriculture and economy of the Missouri, Iowa, Nebraska, and Kansas (MINK) region. In: Agricultural Dimensions of Global Climate Change. H.M. Kaiser and T.E. Drennen (eds.). St. Lucie Press, Delray Beach, Florida, pp. 117-135.
Cure, J.D. and Acock, B. 1986. Crop responses to carbon dioxide doubling: A literature survey. Agric. For. Meteorol. 38: 127-145.
Curry, R.B., Part, R.M., Jones, J.W., Boote, K.J. and Allen, L.H. Jr. 1990a. Simulation as a tool for analyzing crop response to climate change. Trans. ASAE 33: 981-990.
Curry, R.B., Peart, R.M., Jones, J.W., Boote, K.J. and Allen, L.H. Jr. 1990b. Response of crop yield to predicted changes in climate and atmospheric CO2 using simulation. Trans. ASAE 33: 1383-1390.
Curry. R.B., Jones, J.W., Boote, K.J., Peart, R.M., Allen, L.H. Jr. and Pickering, N.B. 1995. Response of soybean to predicted climate change in the USA. 1995. In: Climate Change and Agriculture: Analysis of Potential International Impacts. C. Rosenzweig, L.A. Harper, S.E. Hollinger, J.W. Jones, and L.H. Allen, Jr. (eds.). ASA Special Pub. No. 59, American Society of Agronomy, Madison, Wisconsin, pp. 163-182.
Delucia, E.H., Sasek, T.W. and Strain, B.R. 1985. Photosynthesis inhibition after long-term exposure to elevated levels of atmospheric carbon dioxide. Photosyn. Res. 7: 175-184.
Dudek, D.J. 1989. Climate change impacts upon agriculture and resources: A case study of California. In: The Potential Effects of Global Climate Change on the United States. Appendix C, Agriculture, Vol. 1. EPA-230-05-89-053. J.B. Smith and D.A. Tirpak (eds.). US Environmental Protection Agency, Washington DC. pp. 5-1 to 5-38.
Hall, A.E. and Allen, L.H. Jr. 1993. Designing cultivars for the climatic conditions of the next century. In: International Crop Science 1. D.R. Buxton, R. Shibles, R.A. Forsberg, B.L. Blad, K.H. Asay, G.M. Paulsen and R.F. Wilson (eds.). Crop Science Society of America, Madison, Wisconsin, pp. 291-297.
Hansen, J., Fung, I., Lacis, A., Lebedeff, S., Rind, D., Ruedy, R., Russell, G. and Stone, P. 1988. Global climate changes as forecast by the GISS 3-D model. J. Geophys. Res. 98: 9341-9364.
Idso, S.B., Kimball, B.A., Anderson, M.G. and Mauney, J.R. 1987. Effects of atmospheric CO2 enrichment on plant growth: The interactive role of air temperature. Agric. Ecosystems Environ. 20: 1-10.
Jones, P., Allen, L.H. Jr., Jones, J.W. Boote, K.J. and Campbell, W.J. 1984. Soybean canopy growth, photosynthesis, and transpiration responses to whole-season carbon dioxide enrichment. Agron. J. 76: 633-637.
Jones, P., Allen, L.H. Jr. and Jones, J.W. 1985a. Responses of soybean canopy photosynthesis and transpiration to whole-day temperature changes in different CO2 environments. Agron. J. 77: 242-249.
Jones, P., Allen, L.H. Jr., Jones, J.W. and Valle, R.R. 1985b. Photosynthesis and transpiration responses of soybean canopies to short- and long-term CO2 treatments. Agron. J. 77: 119-126.
Jones, P., Jones, J.W. and Allen, L.H. Jr. 1985c. Carbon dioxide effects on photosynthesis and transpiration during vegetative growth in soybeans. Soil Crop Sci. Soc. Fla. Proc. 44: 129-134.
Keeling, C.D., Whorf, T.P., Wahlen, M. and van der Plicht, J. 1995. Interannual extremes in the rate of rise of atmospheric carbon dioxide since 1980. Nature 375: 660-670.
Kimball, B.A. 1983. Carbon dioxide and agricultural yield: An assemblage and analysis of 430 prior observations. Agron. J. 75: 779-788.
Kimball, B.A., Mauney, J.R., Nakayama, F.S. and Idso, S.B. 1993. Effects of increasing atmospheric CO2 on vegetation. Vegetatio 104/105: 65-75. Also: pp. 65-75. In: CO2 and Biosphere. (Advances in Vegetation Science 14). J. Rozema, H. Lambers, S.C. van de Geijn and M.L. Cambridge (eds.). Kluwer Academic Publishers, Dordrecht.
Kramer, P.J. 1981. Carbon dioxide concentration, photosynthesis, and dry matter production. Bioscience 31: 29-33.
Long, S.P. 1991. Modification of the response of photosynthetic productivity to rising temperature by atmospheric CO2 concentrations: Has its importance been underestimated? Plant Cell Environ. 14: 729739.
Manabe, S. and Wetherald, R.T. 1987. Large scale changes of soil wetness induced by an increase in atmospheric carbon dioxide. J. Atmos.Sci. 44: 1211-1235.
Parry, M.L., Carter, T.R. and Konijn, N.T. (eds.). 1988. The Impact of Climatic Variations on Agriculture. Vols. 1 and 2. Kluwer Academic Publishers, Dordrecht.
Peart, R.M., Jones, J.W., Curry, R.B., Boote, K.J. and Allen, L.H. Jr. 1989. Impact of climate change on crop yield in the Southeastern U.S.A.: A simulation study. In: The Potential Effects of Global Climate Change on the United States. Appendix C, Agriculture, Vol. 1, EPA-230-05-89-053. J.B. Smith and D.A. Tirpak (eds.). US Environmental Protection Agency, Washington DC. pp. 2-1 to 2-54.
Pickering, N.B., Jones, J.W. and Boote, K.J. 1995. Adapting SOYGRO V5.42 for prediction under climate change conditions. In: Climate Change and Agriculture: Analysis of Potential International Impacts. C. Rosenzweig, L.A. Harper, S.E. Hollinger, J.W. Jones and L.H. Allen, Jr. (eds.). ASA Special Pub. No. 59, American Society of Agronomy, Madison, Wisconsin, pp. 77-98.
Poorter, H. 1993. Interspecific variation in the response of plants to an elevated ambient CO2 concentration. Vegetatio 104/105: 77/97. Also: pp. 77-97. In: CO2 and Biosphere. (Advances in Vegetation Science 14). J. Rozema, H. Lambers, S.C. van de Geijn and M.L. Cambridge (eds.). Kluwer Academic Publishers, Dordrecht.
Ritchie, J.T., Baer, B.D. and Chou, T.Y. 1989. Effect of global climate change on agriculture: Great Lakes Region. In: The Potential Effects of Global Climate Change on the United States. Appendix C, Agriculture, Vol. 1. EPA-230-05-89-053. J.B. Smith and D.A. Tirpak (eds.). US Environmental Protection Agency, Washington DC. pp. 1-1 to 1-42.
Rosenzweig, C. 1989. Potential effects of climate change on agricultural production in the Great Plains: A simulation study. In: The Potential Effects of Global Climate Change on the United States. Appendix C, Agriculture, Vol. 1. EPA-230-05-89-053. J.B. Smith and D.A. Tirpak (eds.). US Environmental Protection Agency, Washington DC. pp. 3-1 to 3-43.
Rosenzweig, C. and Iglesias, A. (eds.). 1994. Implications of Climate Change for International Agriculture: Crop Modelling Study. EPA 230-B-94-003. US Environmental Protection Agency, Washington DC.
Rosenzweig, C. and Parry, M.L. 1993. Potential impact of climate change on world food supply: A summary of a recent international study. In: Agricultural Dimensions of Global Climate Change. H.M. Kaiser and T.E. Drennen (eds.). St. Lucie Press, Delray Beach, Florida, pp. 87-116.
Rosenzweig, C. and Parry, M.L. 1994. Potential impact of climate change on world food supply. Nature 367: 133-138.
Rosenzweig, C., Harper, L.A., Hollinger, S.E., Jones, J.W. and Allen, L.H. Jr. (eds.). 1995. Climate Change and Agriculture: Analysis of Potential International Impacts. ASA Special Pub. No. 59, American Society of Agronomy, Madison, Wisconsin.
Rowland-Bamford, A.J., Allen, L.H. Jr., Baker, J.T. and Bowes, G. 1991. Acclimation of rice to changing atmospheric carbon dioxide concentration. Plant Cell Environ. 14: 577-583.
Rozema, J., Lambers, H., van de Geijn, S.C. and Cambridge, M.L. (eds.). 1993. CO2 and Biosphere. (Advances in Vegetation Science 14). Kluwer Academic Publishers, Dordrecht.
Satake, T. and Yoshida, S. 1978. High temperature induced sterility in indica rices at flowering. Japan. J. Crop Sci. 47: 6-17.
Smith, J.B. and Tirpak, D.A. (eds.). 1989. The Potential Effects of Global Climate Change on the United States. EPA-230-05-89-050. US Environmental Protection Agency, Washington DC.
Thorne, J.M. and Koller, R.M. 1974. Influence of assimilate demand on photosynthesis, diffusive resistance, translocation and carbohydrate levels of soybean leaves. Plant Physiol. 54: 201-207.
Valle, R., Mishoe, J.W., Campbell, W.J., Jones, J.W. and Allen, L.H. Jr. 1985. Photosynthetic responses of 'Bragg' soybean leaves adapted to different CO2 environments. Crop Sci. 25: 333-339.
Vergara, B.S. 1980. Rice plant growth and development. In: Rice Production and Utilization. B.S. Luh (ed.). Avi Publishing, Westport, CT. pp.75-86.
Wilson, C.A. and Mitchell, J.F.B. 1987. A doubled CO2 climate sensitivity experiment with a global model including a simple ocean. J. Geophys. Res. 92: 13315-13343.
Wittwer, S.H. 1995. Food, Climate, and Carbon Dioxide: The Global Environment and World Food Production. CRC Press/Lewis Publishers, Boca Raton, Florida.
Woodwell, G.M. and Mackenzie, F.T. (eds.). 1995. Biotic Feedbacks in the Global Climatic System: Will the Warming Increase the Warming? Oxford University Press, New York.
Wullschleger, S.D., Post, W.M. and King, A.W. 1994. On the potential for a CO2 fertilization effect in forests: Estimates of the Biotic Growth Factor based on 58 controlled-exposure studies. In: Biotic Feedbacks in the Global Climatic System: Will the Warming Increase the Warming? G.M. Woodwell and F.T. Mackenzie (eds.). Oxford University Press, New York. pp. 85-107.
Yoshida, S. 1977. Rice. In: Ecophysiology of Tropical Crops. P.T. Alvim and T.T. Kozlowski (eds.). Academic Press, London, pp. 57-87. Yoshida, S. 1981. Fundamentals of Rice Crop
![]()
![]()
![]()
{kind=link}
![Figure 4.2. Comparaison du flux photosynthétique net (Pn) vs. le traitement d'acclimatation à long terme [CO2] d'un couvert de riz soumis en 1987 à des traitements de concentration en CO2 subambiant (160, 250), ambiant (330), et superambiant (500, 660, et 900 m mol CO2/mol air). Les estimations de Pn ont été obtenues durant une très courte période de changement de [CO2] au cours d'une étude où la concentration en CO2 était maintenu pendant la matinée aux six niveaux de concentrations à long terme. Les changements eurent lieu aux jours 62, 63 et 641 respectivement pour des concentrations de 160, 330 et 660 m mol/mol pour tous les six traitements à long terme, respectivement aux jours 62, 63 et 64 après plantation. Le Pn a été estimé à partir d'une équation de régression linéaire de Pn vs la densité de flux de photons photosynthétiques (PPFD) avec PPFD réglé à 1 500 (m mol/photons/m²s. Les barres verticales représentent les intervalles de confiance à 95%. Adapté de Baker et al. (1990b, 1996b) et Allen et al., (1995)](w5183f0n.gif){kind=link}
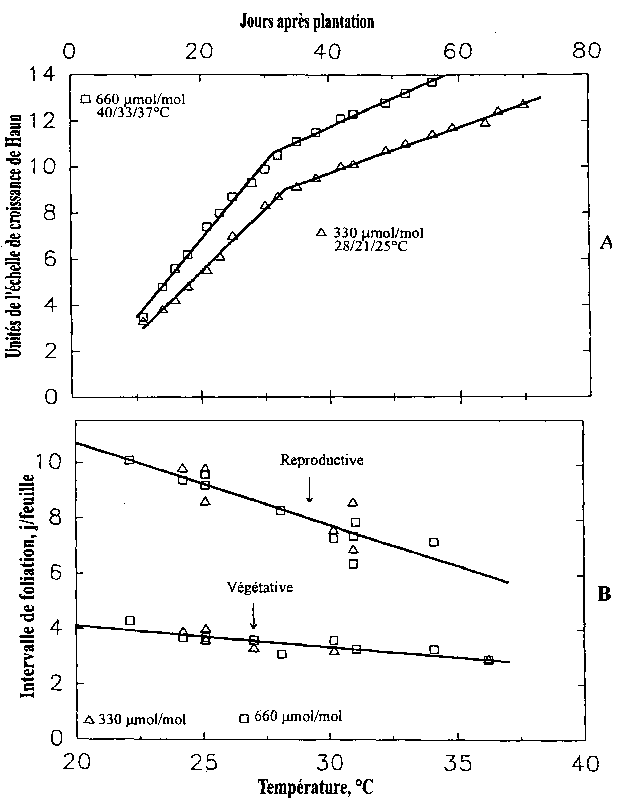{kind=link}