![]()
![]()
![]()
Il est important de comprendre que les bouleversements des ressources halieutiques ne sont pas le fait de quelques stocks particuliers dans le monde mais qu'il se sont produits et continuent de se produire dans de nombreuses régions avec une fréquence qui témoigne de l'existence d'un probléme majeur qui doit être pris en considération (Fig. 1). Bien que les captures ne soient pas de bons indices d'abondance, on admet généralement que ces grandes fluctuations reflétent essentiellement des variations d'abondance. Il faut noter l'accroissement rapide des captures et leur chute abrupte aprés quelques années d'exploitation intense, surtout pour les deux dernières décennies. La fig. 2 montre que ces variations peuvent prendre l'aspect d'oscillations régulières à grande période (proche des cycles solaires par exemple). La fig. 3 montre que les oscillations observées pour des stocks situés en divers endroits du monde peuvent être synchrones, suggérant un rôle important des facteurs climatiques à grande échelle.
Dans l'Atlantique oriental, des variations trés importantes ont également été observées. Les captures et la biomasse des sardines se sont accrues dans la zone située au nord du Cap Blanc de maniére exceptionnelle (Belvéze et Erzini 1983) alors que les fonds de pêche s'étendaient de 850 km environ vers le sud (voir fig. 1 et Garcia, 1982). Dans la même région et plus particulièrement au nord du Cap Bojador, les stocks inexploités de bécasse de mer (macroranphosus Scolopax) subissaient d'importantes fluctuations de biomasse (ISPM 1983) offrant vers la moitié des années 70 un potentiel récemment créé de 400 000 tonnes pratiquement disparu sans exploitation au début des années 80. Les sardinelles et les maquereaux ont simultanément fortement diminué au large des côtes du Sahara pendant que la distribution géographique des sardinelles régressait vers le sud et qu'un effondrement du stock de sardinelles et une disparition des maquereaux étaient observés dans l'ouest du golfe de Guinée, de la Côte d'Ivoire au Togo (Fig. 4). Au même moment, les balistes, une espèce peu abondante traditionnellement ont développé des biomasses estimées à plusieurs centaines de milliers de tonnes, dans le golfe de Guinée et dans le secteur Sherbro-Cap Vert, envahissant même saisonnièrement les côtes nord du Sénégal jusqu'à la Mauritanie (Caverivière 1982).
Au large de la Namibie, les oscillations semblent liées à l'effondrement des stocks de sardine et d'anchois et à l'accroissement progressif de certains stocks de chinchards (fig. 1).
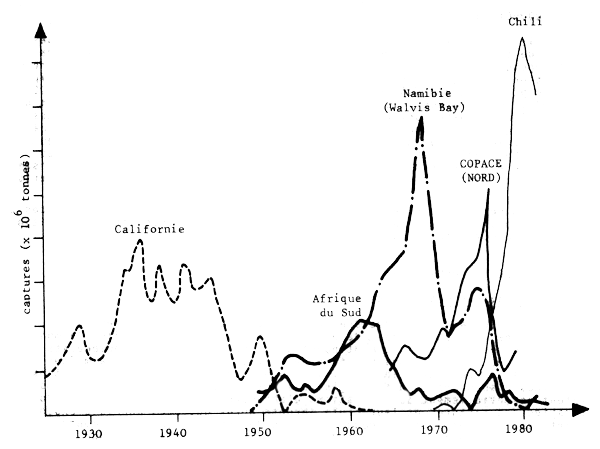
Fig. 1 - Variations des débarquements de sardine dans diverses régions d'updwelling (Parrish, 1983)
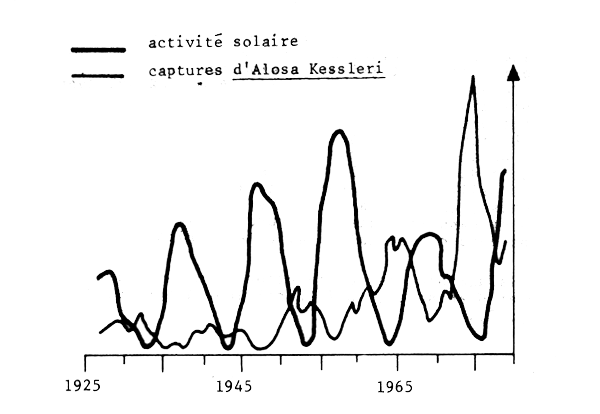
Fig. 2 - Variations des captures de l'Alosa Kessleri dans la Mer Noire (Ivanov, 1984)
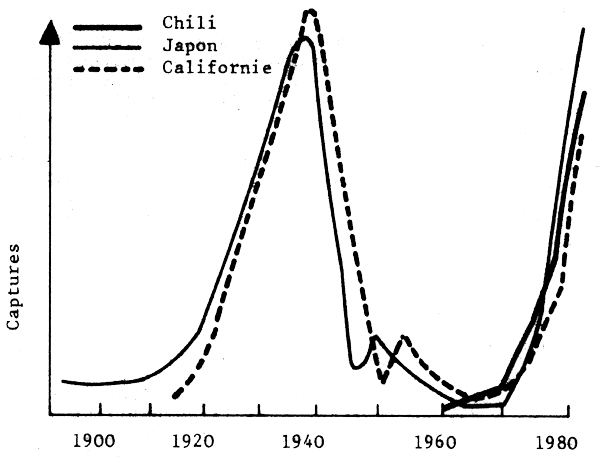
Fig. 3 - Variations simultanées des débarquements de sardine (réf.).
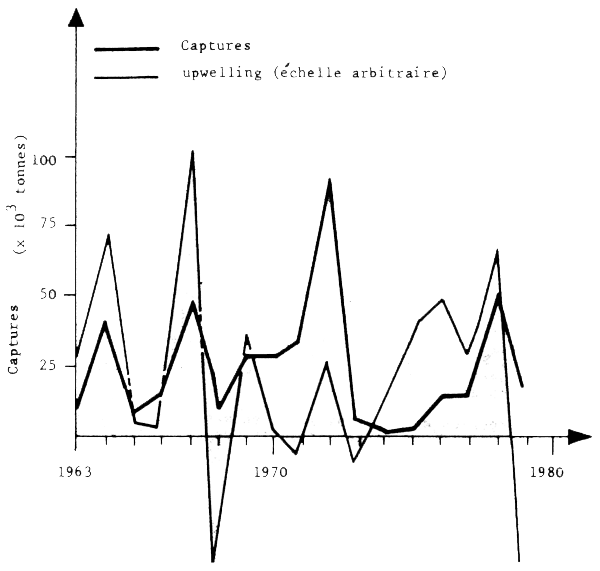
Fig.4 - Upwelling et captures de S. aurita dans le Golfe de Guinée (données CRO Abidjan, FAO/COPACE, 1980).
En Californie, Afrique du sud et Pérou, on a montré que les variations d'abondance étaient associées à des variations de très nombreux paramétres biologiques tels que la croissance, l'âge de première maturité sexuelle, les facteurs de condition, la fécondité, la mortalité naturelle, la distribution géographique, etc., indications de “mutations” profondes de la population en réponse à un stress trés important. Ces changements marqués dans le temps semblent n'être qu'un autre aspect de la variabilité, importance déjà démontrée au niveau géographique car les variations de paramétres biologiques sont également très importantes d'une région à l'autre. En outre, il est maintenant bien établi que le comportement grégaire de ces espéces provoque des changements importants de la capturabilité (qui augmente quand la biomasse diminue).
Il a été à maintes reprises suggéré que la biomasse produite par un écosystéme est plus constante que celle des éléments spécifiques qui la composent ou, en d'autres termes, qu'une espéce dont le stock s'est effondré sera remplacée par une autre. Le cas de la sardine et de l'anchois de Californie est même devenu un cas classique utilisé pour étayer la théorie de la compétition interspécifique. Cependant, plus récemment, McCall (1983) conteste l'hypothése du “remplacement” d'espéces au sens strict (impliquant des phénoménes de compétition interspécifiques) dans le cas de la Californie et considére qu'il peut s'agir d'avantage de changements importants de la biomasse et de la distribution géographique des ressources, sous l'action de l'environnement.
Les exemples de remplacements apparents sont trés nombreux (Fig. 5 à 9) dans la plupart des régions du monde et l'Atlantique centre-est a également été le siége de “remplacements” trés importants (Garcia, 1982).
Entre le Cap Blanc et le Cap Barbas, les remplacements observés chez les petits pélagiques sont dus à des changements de la distribution géographique et de la disponibilité (Belvéze, 1983). Il n'en reste pas moins vrai que ces déplacements, dans le cas de la sardine ainsi que du baliste se sont accompagnés d'accroissements considérables de la biomasse de ces espéces, au détriment semblet-il des autres espéces (Caveriviére 1982, Gulland et Garcia 1984). Il est important de noter que ces remplacements, qui n'apparaissent pas toujours comme immédiats ou automatiques, peuvent être favorables tels que le remplacement des sparidés par des céphalopodes dans la zone COPACE nord ou défavorables tels que le remplacement d'espèces traditionnellement commercialisées par des balistes dans tout le golfe de Guinée.
Des travaux importants ont été entrepris pour décrire les variations des ressources et tenter d'en comprendre les mécanismes, en Californie (McCall, 1983), au Pérou (Jordan, 1983), au Japon (Hayasi, Tanaka, 1983), etc., ainsi qu'au Maroc (Belvèze et Erzini, 1983), au Sénégal (Fréon, 1983) en Côte-d'Ivoire et au Ghana (FRU/ORSTOM, 1975; FAO/COPACE, 1980; Binet, 1982; Gulland et Garcia, 1983). La Consultation d'experts organisée par la FAO en avril 1983 s'est largement étendue sur le sujet (Sharp et Csirke, 1983).
Si les divergence de vue subsistent encore en ce qui concerne les mécanismes détaillés, il est maintenant largement admis que ces variations sont liées essentiellement à des fluctuations climatiques dont les conséquences sont aggravées par une exploitation trop intensive.
Les variations climatiques se traduisent vraisemblablement par des variations du taux de survie des recrues et donc de leur abondance. Les variations, qui se traduisent par des courtes séries de bons ou mauvais recrutements sont accompagnées de variations de la maturité sexuelle, de la croissance, de la fécondité, de la mortalité naturelle, etc. La relation stock-recrutement est également affectée et l'on pourrait dire que les variations d'abondance sont liées à des changements de la capacité biologique de l'environnement vis-à-vis de la phase larvaire.
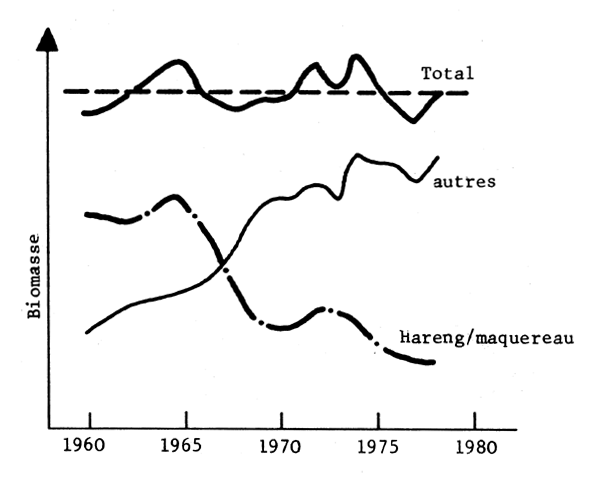
Fig. 5 - Mer du Nord
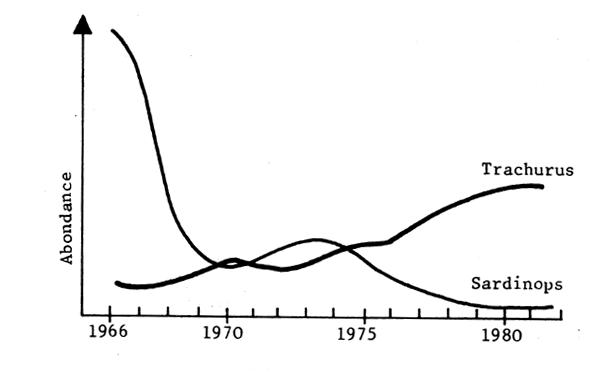
Fig .6 - Namibie
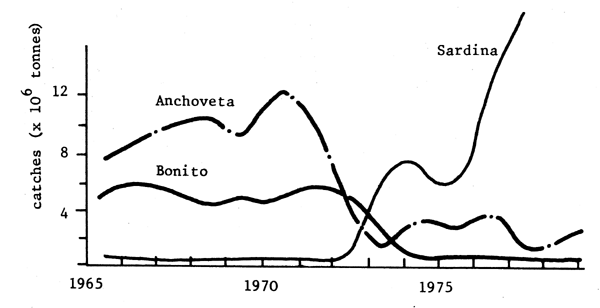
Fig .7 - Pérou
Fig. 5, 6, 7 - Expemples de “remplacements” d'espèces en Mer du Nord (Ursin, 1982), Namibie (Crawford, Shelton et Hutchings, 1983) et Pérou (Jordan, 1983).
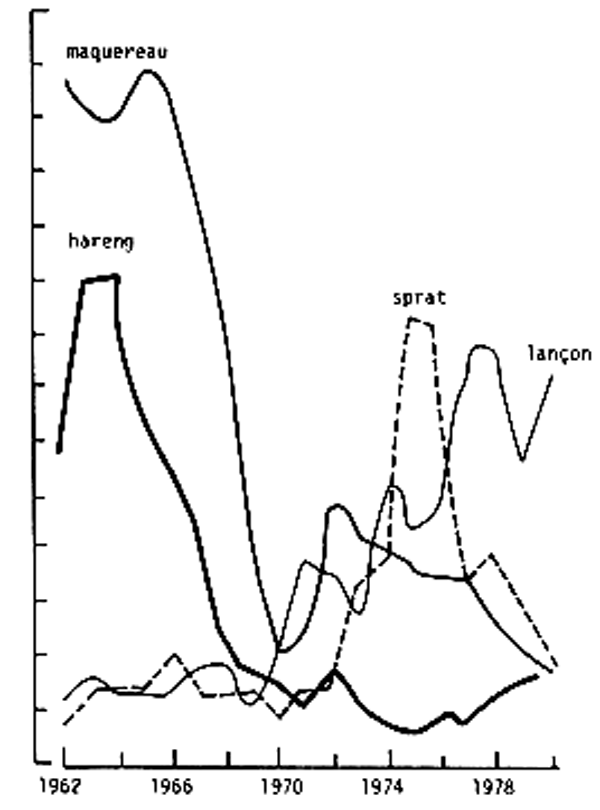
Fig. 8 - Variation de la biomasse de ponte du hareng et du maquereau et débarquements de sprat et de lançon en Mer du Nord (Jones, 1983)
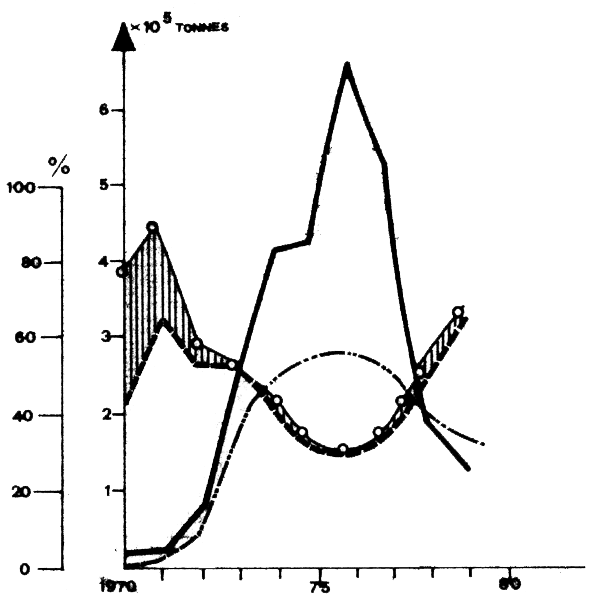
Fig. 9 - Vartiations des captures de sardine (—) et des proportions de sardine (—), chinchards et maquereaux (—) chinchards, maquereaux et sardinelles(0—0). Source Bull.Stat.COPACE no 3, 1981.
Il n'est pas impossible que la compétition interspécifique, les relations prédateur-proie, ou les effets secondaires de la pêche (sur une autre espéce que celle dont on considére les oscillations) aient également une influence, mais celle-ci n'a jamais pu être démontrée de maniére satisfaisante alors que les effects de l'environment sont souvent flagrants.
Des informations collectées dans les sédiments, sur l'abondance des stocks de sardines et d'anchois dans le passé ont montré que des effondrements biologiques avaient eu lieu, pratiquement en l'absence de toute exploitation au cours des deux derniers siécles (Soutar et Isaacs, 1974).
Il est cependant admis que les fluctuations sont aggravées par la pêche qui accroit également considérablement le risque d'effondrement.
L'analyse de cette variabilité est intéressante car elle permet de connaitre le type de variabilité auquel on doit s'attendre au niveau de la ressource. La fig. 10 démontre clairement que l'un des phénoménes marquant le climat de l'Afrique de l'Ouest est l'existence d'un cycle trés net de sécheresses sahéliennes. Ces sécheresses sont liées aux mouvements des fronts atmosphériques et ont pour conséquence directe des variations importantes des apports fluviaux et donc de la salinité et de la productivité primaire dans les zones côtiéres (Binet, 1982).
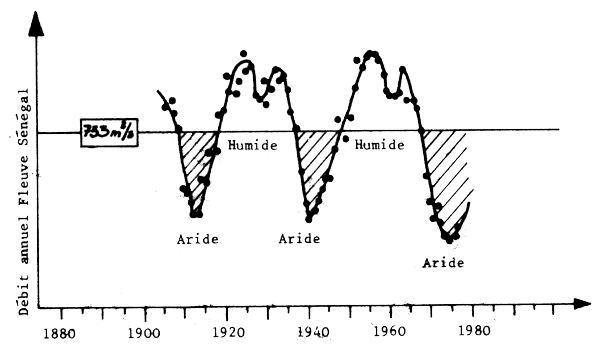
Fig. 10 - Variations climatiques à long terme en Afrique de l'Ouest (Faure et Gac, 1981).
Elles sont accompagnées de changements importants dans la vitesse et la direction des vents dominants et en particulier des vents d'upwellings (fig. 4, llA et llB) lesquels se traduisent également par des variations importantes de la productivité primaire. Les fig. llA et llB suggérent en outre que les variations hydroclimatiques observées le long de la côte et de l'Atlantique sont peut être corrélées. Les variations apparaissant similaires ou opposées suivant les cas compte tenu de la briéveté des séries chronologiques. Cette hypothése doit cependant être considérée avec la plus grande prudence. La fig. llB indique enfin qu'au-delà des oscillations à moyenne fréquence existent également des tendances à long terme à l'affaiblissement de l'upwelling (Sénégal) ou à son renforcement (Maroc).
Les travaux de Belvéze et Erzini (1983) au Maroc et de Fréon (1983) au Sénégal indiquent que les variations d'abondance des stocks sont liées à l'environnement (fig. 12 et 13). Fréon a même proposé un modéle de production pour la sardinelle du Sénégal intégrant les fluctuations de l'environnement.
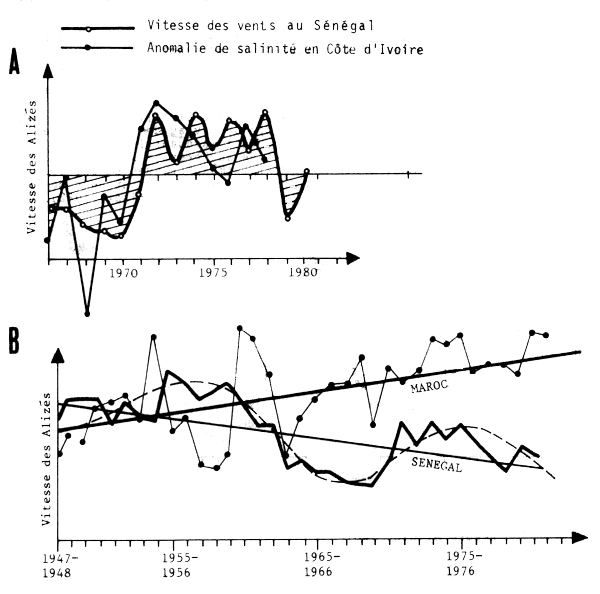
Fig. 11 - Variations hydroclimatiques en Afrique de l'Ouest : A - Anomalie de salinité à Abidjan (FAO/CECAF, 1981) et vitesse des alizés au Sénégal (Fréon, 1983)
B - Indice d'upwelling au Maroc (zone B), (Belvèze, 1983 et vitesse des vents au Sénégal (Fréon, 1983)
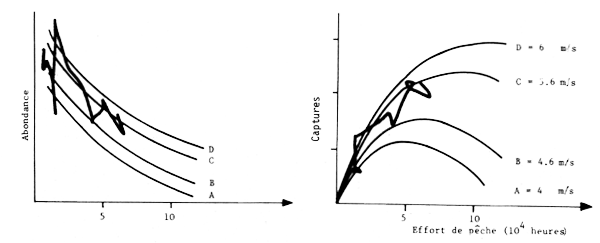
Fig. 12 - Modèle de production modifié par l'action du vent (et donc par l'upwelling pour les sardinelles du Sénégal) (Dréon, 1983)
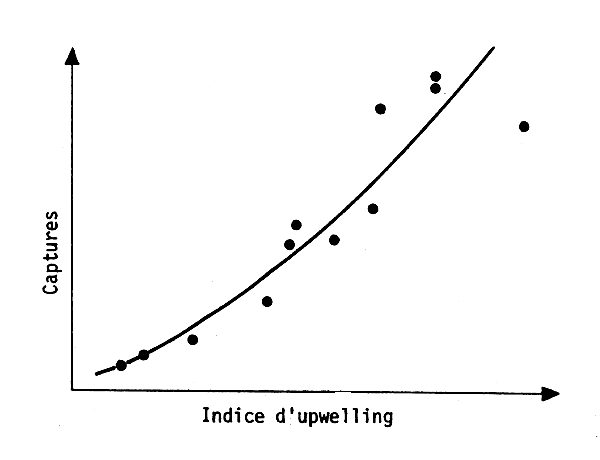
Fig. 13 - Captures de sardines et upwelling au Maroc (Belvèze et Erzini, 1983)
![]()
![]()
![]()