Chapter 12 : Dietary fat and immune response
Arachidonic acid (AA) metabolites
Polyunsaturated fatty acids (PUFA)
Conclusion
Interest in the contribution of lipids to immune function began with the recognition of their modifying effect on the reticuloendothelial system (Di Luzio, 1972). An additional stimulus for this interest was the suggestion that an association may exist between certain types of cancer and the quality and quantity of dietary lipids (Carroll and Khor, 1975; Tannenbaum and Silverstone, 1953). Also, dietary fat has been shown to affect the severity of autoimmune diseases as well as the length of allograft acceptance (Merlin and Hunt, 1976; Ring et al., 1974). Furthermore, the in vitro addition of fatty acids to lymphocyte cultures changed their mitogenic response (Merlin and Hughs, 1975).
Cooperation between different cells of the immune system via membrane-associated events and through different protein and lipid mediators is essential in mounting a successful immune system response. Dietary lipids can affect the immune system by influencing substrate availability in the formation of cyclooxygenase and lipooxygenase products. These products, in turn, act as lipid mediators in the control of the immune system (Rola-Plaszczynski, 1985 and Goodwin, Messner and Peake, 1974). Furthermore, the cells of the immune system are highly dependent on cell membrane function for operations such as the secretion of lymphokines and antibodies, antigen reception, lymphocyte transformation, and contact lysis. The importance of lipids in the maintenance of membrane integrity (Stubbs and Smith, 1984) indicates that they are potentially critical nutrients in the regulation of the immune function.
Arachidonic acid (AA) metabolites
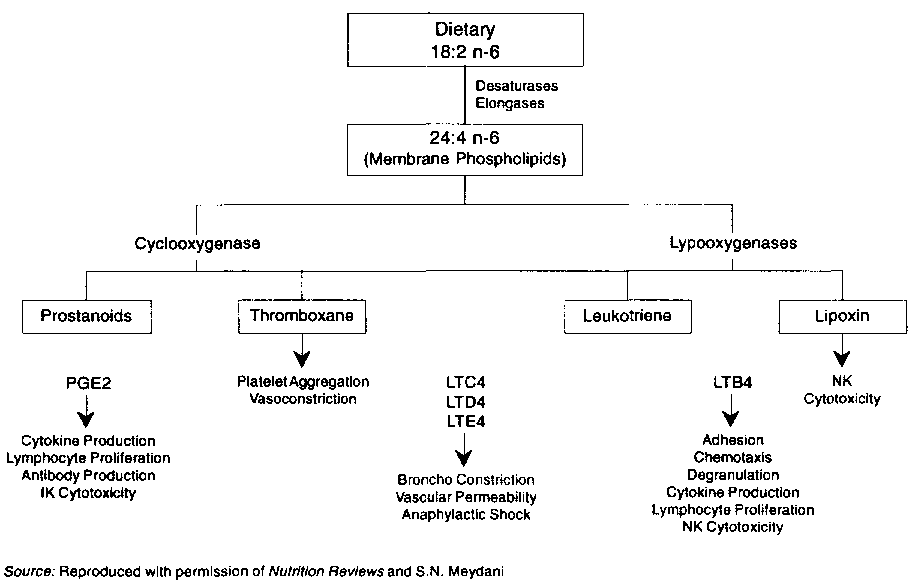
Arachidonic acid (AA) metabolites play a role in control of the immune system. Arachidonic acid (AA) metabolites, prostaglandins (PG), hydroxyeicosatetraenoic acid (METE) and leukotrienes (LT), are produced by human peripheral blood mononuclear cells (PBMC) and by mouse splenocytes in response to stimulation by mitogens or antigens. They also inhibit subsequent T cell proliferation. Similarly, inhibition of PG synthesis in vitro enhances T cell proliferation. Cellular and humoral immune responses operate under negative control by PG. In vitro, PGE2 inhibits T cell proliferation (Goodwin, Messner and Peake, 1974; Webb, Rogers and Nowowiejiski, 1980; Rola-Plaszczynski and Lemaire, 1985; Metzger, Hoffeld and Oppenheim, 1980; Fisher and Bostic-Bruton, 1982; Muscoplat, Rakich and Thoen, 1978), lymphokine production (Gordon, Bray, and Morley, 1976), the generation of cytotoxic cells (Plaut, 1979), and natural killer cell (NK) activity (Roder and Klein, 1979).
In addition to PGE2, lipooxygenase products, that is, LT and METE, have been shown to inhibit lymphocyte proliferation in mouse splenocytes and human peripheral lymphocytes (Goodman and Weigle, 1980). This effect may be mediated by decreased T helper and increased T suppressor/cytoxic cell proliferation (Payan, Missirian-Bastian and Goetzl, 1984; Gualde et al., 1984). In some but not all studies, LTB4 has been shown to increase interleukin (IL)-1 as well as IL-2 production and lymphocyte proliferation (Rola-Plaszczynski and Lemaire, 1985). Lipoxins (Lx) which are products of 15-lipoxygenase of AA have been shown to inhibit human NK activity when tested against K562 target cells (Ramstedt et al., 1985). On the other hand, LTB4 was shown to augment human NK activity (Rola-Plaszczynski, Gagnon and Sirois, 1983). The effect of AA metabolites on cell-mediated immunity is summarized in Figure 12.1.
Polyunsaturated fatty acids (PUFA)
Several investigators have evaluated the effect of dietary fat on the immune response (Johnson and Marshall, 1984; Erickson, 1986). Review of the literature presents a confusing picture mainly because the effect of PUFA on the immune response varies depending on a number of factors such as: concentration of dietary fat; duration of supplementation; genetic variation; existence of infectious or autoimmune and inflammatory diseases; age of the animal; status of other nutrients such as vitamin E, where the requirement is affected by the degree of saturation of fatty acids; presence or absence of essential fatty acid deficiency in the control diet; particular immunological test used in the experiment; and, series of PUFA tested that is, n-6 or n-3 PUFA.
FIGURE 12.1 : Role of eicosanoids in regulation of immunological and inflammatory processes
Plant-denved (n-6) PUFA. The effect of n-6 PUFA of plant origin on the immune response was recently reviewed (Meydani et al., 1991 b; Dupont et al., 1990). In the majority of animal studies, essential fatty acids (EFA) were necessary for normal immune function. Essential fatty acid deficiency impairs b and T cell-mediated responses. These impairments are normalized by the inclusion of essential fatty acids in the diet. High-fat diets (about 45 en %), regardless of the degree of saturation, suppress in vivo and in vitro indices of the immune system. With few exceptions, when animals which were not deficient in fatty acids were fed PUFA of plant origin (high percentage of n-6 FA) they had similar responses to those fed the same level of saturated fatty acids from animals or plant origin. This is especially evident at moderate levels of dietary fat (around 22 en %). At higher levels of dietary fat (around 45 en %) or when mice were challenged with infectious agents or chemical carcinogens, PUFA-fed mice tended to have lower responses to some, but not all, of the immunological tests than those mice fed the same level of saturated fatty acids. These conclusions cannot be readily extrapolated to humans because the immune systems of different species and strains within species respond differently to dietary fat alterations. Well-controlled human trials are needed to determine the effects of dietary fat levels and saturation on human immune responses.
The few studies of the effect of dietary fat on immune response in humans have been epidemiological, retrospective studies. Double-blind prospective studies of multiple sclerosis (MS), a possibly autoimmune demyelinating disease with lipid changes (Bornstein, 1973), suggests a deficiency of PUFA (Bernsohn and Stephanides, 1967; Mertin and Meade, 1977). One study showed that MS patients have less than normal linoleic acid in their lymphocyte lipids and that MS patients supplemented with safflower oil for two years had less frequent and less severe relapses than those supplemented with olive oil (Miller et al., 1973). Another study showed that lymphocytes from MS patients had lower mitogenic responses than control subjects and that supplementation with evening primrose seed oil (which contains 8 percent oleic, 75 percent linoleic, and 9 percent g -linoleic acid C18:3n-6) for 85 days improved the mitogenic response (Offner, Konat and Clausen, 1974).
Supplementation of MS patients and healthy subjects with 21 g/day safflower oil for 5 weeks increased agglutination with both measles antigen and concanavalin A (Con A), but supplementation with olive oil had no effect (Utermohlen et, al., 1981). PUFA supplementation, combined with conventional immunosuppressive therapy, facilitated the acceptance of kidney grafts in humans (Uldall, 1974).
In a cross-sectional study of lymphocyte mitogenesis to b and T cell mitogens and NK activity in 94 free-living subjects, there was no correlation between the degree of adipose tissue fatty acid unsaturation (presumably reflecting dietary consumption) or plasma lipoprotein and the immunological indices (Berry et al., 1987). The inter-individual variation and the influence of other dietary factors could mask a possible effect of fat.
A recent study showed that long term consumption of low fat diets with a moderate level of (n6) PUFA (6 percent) had no effect on delayed-type hypersensitivity skin response (DTH) which can in vivo measure T cell-mediated functions in healthy subjects. It did, however, significantly increase the proliferative response of lymphocytes to T cell mitogen Con A and ex vivo production of IL-1,B.
Plant-derived (n-3) PUFA. a -linolenic (C18:3n-3) can be converted to EPA (C20:5n-3) which can replace arachidonic acid in membrane phospholipids and be preferentially used by cyclooxygenase. This results in reduced production of PG of the 2 series and leukotrienes of the four series. Thus, it is proposed that consumption of oils containing C18:3n-3 might be associated with immunological changes. This proposition is supported by the many studies which have shown immunological modulation following marine oil consumption. There are very few immunologic studies of plant oils containing (n-3) PUFA. Plant oils containing (n-3) PUFA include perilla seed oil, which has the highest content of (n-3) PUFA (64 percent), followed by linseed oil (62 percent), canola oil (10 percent), and soybean oil (7 percent). In extrapolating the results obtained with (n-3) PUFA of marine oil to oils containing C18:3n-3, two questions should be kept in mind: first, what is the conversion rate of C18:3n-3 to longer-chain (n-3) PUFA (that is, C20:5n-3 and C22:6n-3); and second, is the percentage of (n-3) PUFA in the oil adequate to modify arachidonic acid metabolism and thereby immunological and inflammatory processes?
A recent review of the literature (Meydani et al., 1991 b) indicates that consumption of oils containing C18:3n-3 is associated with a modest decrease in eicosanoid production and has no or a slight stimulatory effect on immune response. Long term consumption of low fat diets containing 1.8 percent 18:3n-3 resulted in a significant decrease in plasma AA level associated with a significant increase in T cell mitogenic response to Con A and IL-1,6 production (Meydani et al., 1993) in humans. Other immunological indices were not affected.
Marine-developed (n-3) PUFA. Different aspects of inflammatory and cell-mediated immune response are under control by PGE2. PGE2 production is decreased following consumption of (n-3) PUFA. Several investigators have studied the effect of increased intake of these fatty acids on cytokine production and cell-mediated immune responses. Human studies have consistently demonstrated a decrease in production of pro-inflammatory cytokines when moderate to high levels of marine-derived (n-3) PUFA are taken orally (Meydani et al., 1993; Kremer et al., 1987; Meydani et al., 1991a; Endres et al., 1989). The decrease in production of pro-inflammatory cytokines and eicosanoids contributes to the beneficial effect of fish oil in reducing pathogenesis of inflammatory and atherosclerotic diseases.
Early epidemiological studies demonstrated low incidence of atherosclerotic, inflammatory and autoimmune diseases in Greenland Eskimos who consumed diets high in marine-derived (n-3) PUFA. A modest beneficial effect of fish oil has been demonstrated in human studies of arthritis (Kremer et al., 1990). Fish oil supplementation reduced the clinical symptoms of psoriasis, a skin inflammatory disease (Ziboh et al., 1986). A modest beneficial effect of fish oil was also demonstrated in ulcerative colitis (Lorenz et al., 1989).
Several studies have evaluated the effect of fish oil supplementation on cell-mediated immunity. Also, in vivo supplementation with fish oil has been shown to decrease T- and B-cell proliferation and DTH, an in vivo measure of cell-mediated immunity (Meydani et al., 1991; Kramer et al., 1991; Santoli and Zurier 1989; Virella et al., 1989).
The immunosuppressive effect of fish oil was also observed in subjects (more than 40 years old) consuming diets based on the National Cholesterol Education Panel (NCEP) recommendation (NCEP Step-2) enriched with fish-derived (n-3) PUFA (providing 0.54 percent energy or 1.23 g/day EPA and DHA) for 6 months (Meydani et al., 1993). This long-term consumption of the NCEP Step-2 diet enriched with fish caused a small but significant decrease in the percentage of helper T cells, whereas the percentage of suppressor T cells was increased. This change was accompanied by a significant reduction in DTH and mitogenic response to concanavalin A. A significant correlation between changes in DTH and plasma EPA levels was observed. As reported with fish oil supplementation, the production of cytokines, IL-1b , TNF-a , and IL-6 by PBMCs was also reduced (Ibid.). In contrast, long-term feeding of the NCEP Step-2 diet low in fish (0.13 percent energy or 0.27 g/day from EPA and DHA but with the same level of total PUFAs, that is, 10 percent) increased PBMC mitogenic response to the T-cell mitogen concanavalin A and increased IL-1b and TNF-a production but had no effect on the production of IL-6 or DTH. Although the NCEP Step-2 diet enriched with fish-derived (n-3) PUFA caused a significant decrease in the plasma ratio of tocopherol to PUFA the NCEP Step-2 diet low in fish did not cause a significant decrease in this ratio. The results of this study, as well as those observed in animal studies described above, indicate that the level of (n-3) PUFA and tocopherol status are important determinants of the nature of the effect on the immune response which was observed after consumption of these fatty acids.
The reduced production of pro-inflammatory cytokines and pro-aggregatory eicosanoids might be beneficial in cases where excess levels of these mediators are produced, for example, in the case of inflammatory diseases. However, in healthy subjects or those with compromised immune status, reduction in cytokines or eicosanoids production could compromise their normal biological functions which might result in an impairment of host defense or normal platelet aggregation. This can be better understood when the dual function of these compounds, that is, the physiological regulatory role as well as the contribution to the pathogenesis of diseases is considered.
Along these lines, fish oil supplementation has been shown to decrease IL-2 production, T cell mitogen-induced proliferation and the DTH in animal and human models. Increased cerebral hemorrhage has also been observed with fish oil consumption. Furthermore, fish oil feeding inhibited the ability of mouse accessory cells to present antigen to T helper cells (Fujikawa et al., 1992) and mice which were fed fish oil had a reduced capacity for cytotoxicity of mastocytoma cells upon stimulation with LPS or INF-g (Hubbard, Somers and Erickson, 1991).
It has been shown that long-term consumption of moderate levels of marine-derived (n-3) PUFA decreases percent T helper cells, mitogenic response to T cell mitogen (by 34 percent) and DTH (by 50 percent) (Meydani et al., 1993). The decrease in DTH is of particular interest as this test has been shown to be a predictor of morbidity and mortality. The clinical significance of this finding is supported by the observations that fish oil feeding decreased natural resistance of mice to infection with Salmonella typhimurium (Chang et al., 1992), and rats fed diets containing 9 percent menhaden oil had a shorter life span compared to those fed corn oil or beef tallow (Berdanier et al., 1992). Rabbits given a high fish or safflower supplement (5 g/kg.d) for 7 days after birth had decreased lung clearance of inspired Staphylococcus aureus by-50 percent compared with saline control or low doses of the oils (0.22 g/dg.d) (D'Ambola et al., 1991). Fish oil diets also augmented the nonhistamine-mediated bronchoconstrictor response in pulmonary anaphylaxis (Lee et al., 1985). Further studies are needed to determine the effect of fish oil supplementation on host defense in humans.
Without adequate antioxidant protection, the substitution of membrane fatty acids with highly oxidizable (n-3) PUFA of fish oil, that is, EPA and DHA, may potentiate the peroxidation of cellular membranes. Increased free radical activity has been demonstrated in laboratory animals fed diets high in (n-3) PUFA (Nalbone et al., 1989; Herbert and Wills 1987; Hu M-L et al., 1989). These compounds might contribute to the suppressive effect of fish oil on T cell-mediated functions. These risks which are associated with intake of (n-3) PUFA can be minimized by the intake of appropriate levels of an antioxidant nutrient such as vitamin E without compromising its beneficial effects.
Studies with animals, tissue cultures, as well as humans indicate that both the level and degree of saturation of dietary lipids influence inflammatory and immunological responses. The nature of the effect is dependent on the type of fatty acid, age, antioxidant and health status of the subject. Some of these immunological changes are associated with beneficial clinical effects, for example, reduction in pro-inflammatory cytokine by marine-derived (n-3) PUFA. Others might not be desirable, for example, decreased T cell-mediated function by marine-derived (n-3) PUFA in older people.
Thus, the effect of lipids on immune response should be considered before recommendations to change dietary lipids for other health benefits are made. Furthermore, recommendations to increase PUFA intake should be accompanied by appropriate recommendations for an increase in antioxidant intake, in particular vitamin E.
In general, for healthy individuals, recommendations to decrease dietary total fat intake, with the inclusion of moderate amounts of (n-6) and (n-3) PUFA with adequate antioxidant nutrients should provide for a competent immune response.
{kind=link}