![]()
![]()
![]()
4.1 Microbial consortia and biological aspects of methane fermentation
4.2 Molecular biology of methanogens
4.3 Developments in bioreactor technology
References
Methane fermentation is a versatile biotechnology capable of converting almost all types of polymeric materials to methane and carbon dioxide under anaerobic conditions. This is achieved as a result of the consecutive biochemical breakdown of polymers to methane and carbon dioxide in an environment in which a variety of microorganisms which include fermentative microbes (acidogens); hydrogen-producing, acetate-forming microbes (acetogens); and methane-producing microbes (methanogens) harmoniously grow and produce reduced end-products. Anaerobes play important roles in establishing a stable environment at various stages of methane fermentation.
Methane fermentation offers an effective means of pollution reduction, superior to that achieved via conventional aerobic processes. Although practiced for decades, interest in anaerobic fermentation has only recently focused on its use in the economic recovery of fuel gas from industrial and agricultural surpluses.
The biochemistry and microbiology of the anaerobic breakdown of polymeric materials to methane and the roles of the various microorganisms involved, are discussed here. Recent progress in the molecular biology of methanogens is reviewed, new digesters are described and improvements in the operation of various types of bioreactors are also discussed.
4.1.1 Hydrolysis and acidogenesis
4.1.2 Acetogenesis and dehydrogenation
4.1.3 Methanogenesis
Methane fermentation is the consequence of a series of metabolic interactions among various groups of microorganisms. A description of microorganisms involved in methane fermentation, based on an analysis of bacteria isolated from sewage sludge digesters and from the rumen of some animals, is summarized in Fig. 4-1. The first group of microorganisms secrete enzymes which hydrolyze polymeric materials to monomers such as glucose and amino acids, which are subsequently converted to higher volatile fatty acids, H2 and acetic acid (Fig. 4-1; stage 1). In the second stage, hydrogen-producing acetogenic bacteria convert the higher volatile fatty acids e.g., propionic and butyric acids, produced, to H2, CO2, and acetic acid. Finally, the third group, methanogenic bacteria convert H2, CO2, and acetate, to CH4 and CO2.
Polymeric materials such as lipids, proteins, and carbohydrates are primarily hydrolyzed by extracellular, hydrolases, excreted by microbes present in Stage 1 (Fig. 4-1). Hydrolytic enzymes, (lipases, proteases, cellulases, amylases, etc.) hydrolyze their respective polymers into smaller molecules, primarily monomeric units, which are then consumed by microbes. In methane fermentation of waste waters containing high concentrations of organic polymers, the hydrolytic activity relevant to each polymer is of paramount significance, in that polymer hydrolysis may become a rate-limiting step for the production of simpler bacterial substrates to be used in subsequent degradation steps.
Figure 4.1 - Stages of methane fermentation Source: McCarty, P.L., (1982)
Lipases convert lipids to long-chain fatty acids. A population density of 104 - 105 lipolytic bacteria per ml of digester fluid has been reported. Clostridia and the micrococci appear to be responsible for most of the extracellular lipase producers. The long-chain fatty acids produced are further degraded by p-oxidation to produce acetyl CoA.
Proteins are generally hydrolyzed to amino acids by proteases, secreted by Bacteroides, Butyrivibrio, Clostridium, Fusobacterium, Selenomonas, and Streptococcus. The amino acids produced are then degraded to fatty acids such as acetate, propionate, and butyrate, and to ammonia as found in Clostridium, Peptococcus, Selenomonas, Campylobacter, and Bacteroides.
Polysaccharides such as cellulose, starch, and pectin are hydrolyzed by cellulases, amylases, and pectinases. The majority of microbial cellulases are composed of three species: (a) endo-(3-l,4-glucanases; (b) exo-p-l,4-glucanases; (c) cellobiase or p-glucosidase. These three enzymes act synergistically on cellulose effectively hydrolyzing its crystal structure, to produce glucose. Microbial hydrolysis of raw starch to glucose requires amylolytic activity, which consist of 5 amylase species: (a) a-amylases that endocleave a±1-4 bonds; (b) p-amylases that exocleave a±1-4 bonds; (c) amyloglucosidases that exocleave a±l-4 and a±l-6 bonds; (d) debranching enzymes that act on a±l-6 bonds; (e) maltase that acts on maltose liberating glucose. Pectins are degraded by pectinases, including pectinesterases and depolymerases. Xylans are degraded with a²-endo-xylanase and a²-xylosidase to produce xylose.
Hexoses and pentoses are generally converted to C2 and C3 intermediates and to reduced electron carriers (e.g., NADH) via common pathways. Most anaerobic bacteria undergo hexose metabolism via the Emden-Meyerhof-Parnas pathway (EMP) which produces pyruvate as an intermediate along with NADH. The pyruvate and NADH thus generated, are transformed into fermentation endo-products such as lactate, propionate, acetate, and ethanol by other enzymatic activities which vary tremendously with microbial species.
Thus, in hydrolysis and acidogenesis (Fig. 4-1; Stage 1), sugars, amino acids, and fatty acids produced by microbial degradation of biopolymers are successively metabolised by fermentation endo-products such as lactate, propionate, acetate, and ethanol by other enzymatic activities which vary tremendously with microbial species.
Thus, in hydrolysis and acidogenesis (Fig. 4-1; Stage 1), sugars, ammo acids, and fatty acids produced by microbial degradation of biopolymers are successively metabolised by groups of bacteria and are primarily fermented to acetate, propionate, butyrate, lactate, ethanol, carbon dioxide, and hydrogen (2).
Although some acetate (20%) and H2 (4%) are directly produced by acidogenic fermentation of sugars, and amino acids, both products are primarily derived from the acetogenesis and dehydrogenation of higher volatile fatty acids (Fig. 4-1; Stage 2).
Obligate H2-producing acetogenic bacteria are capable of producing acetate and H2 from higher fatty acids. Only Syntrophobacter wolinii, a propionate decomposer (3) and Sytrophomonos wolfei, a butyrate decomposer (4) have thus far been isolated due to technical difficulties involved in the isolation of pure strains, since H2 produced, severely inhibits the growth of these strains. The use of co-culture techniques incorporating H2 consumers such as methanogens and sulfate-reducing bacteria may therefore facilitate elucidation of the biochemical breakdown of fatty acids.
Overall breakdown reactions for long-chain fatty acids are presented in Tables 4-1 and 4-2. H2 production by acetogens is generally energetically unfavorable due to high free energy requirements (a”Go, > 0; Table 4-1 and 4-2). However, with a combination of H2-consuming bacteria (Table 4-2, 4-3), co-culture systems provide favorable conditions for the decomposition of fatty acids to acetate and CH4 or H2S (a”Go, < 0). In addition to the decomposition of long-chain fatty acids, ethanol and lactate are also converted to acetate and H2 by an acetogen and Clostridium formicoaceticum, respectively.
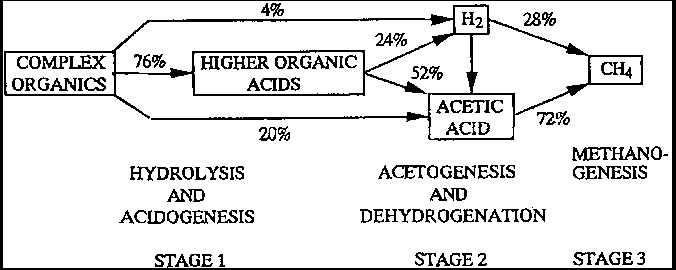
The effect of the partial pressure of H2 on the free energy associated with the conversion of ethanol, propionate, acetate, and H2/CO2 during methane fermentation is shown in Fig. 4-2. An extremely low partial pressure of H2 (10-5 atm) appears to be a significant factor in propionate degradation to CH4. Such a low partial pressure can be achieved in a co-culture with H2-consuming bacteria as previously described (Table 4-2,4-3).
Methanogens are physiologically united as methane producers in anaerobic digestion (Fig. 4-1; Stage 3). Although acetate and H2/CO2 are the main substrates available in the natural environment, formate, methanol, methylamines, and CO are also converted to CH4 (Table 4-3).
Table 4-1 Proposed Reactions Involved in Fatty Acid Catabolism by Syntrophomonas wolfei
|
Fatty Acids |
Reaction |
|
|
Even-numbered |
|
|
|
|
CH3CH2CH2COO- |
+ 2 H2O |
|
|
CH3CH2CH2CH2CH2COO- |
+ 4 H2O |
|
|
CH3CH2CH2CH2CH2CH2CH2COO- |
+ 6 H2O |
|
Odd-numbered |
|
|
|
|
CH3CH2CH2CH2COO- |
+1 H2O |
|
|
CH3CH2CH2CH2CH2CH2COO- |
+ 4 H2O |
|
Branched-chained |
|
|
|
|
CH3CHCH2CH2CH2COO- |
+ 2 H2O |
|
|
||
|
|
||
Source: McInerney et al. (1981)
Table 4-2 Free-Energy Changes for Reactions Involving Anaerobic Oxidation in Pure Cultures or in Co-Cultures with H2-Utilizing Methanogens or Desulfovibrio spp.
|
Equations |
a”G0,' (kJ/reaction) | |
|
1. Proton-reducing (H2-producing) acetogenic bacteria |
| |
|
|
A. CH3CH2CH2COO- + 2H2O |
+48.1 |
|
|
B. CH3CH2COO- + 3H2O |
+76.1 |
|
2. H2-using methanogens and desulfovibrios |
| |
|
|
C. 4H2 + HCO3- + H + |
-135.6 |
|
|
D. 4H2 + S042- + H+ |
-151.9 |
|
3. Co-culture of 1 and 2 |
| |
|
|
A + C 2 CH3CH2CH2COO- + HCO3- + H2O |
-39.4 |
|
|
A + D 2 CH3CH2CH2COO- + S042- |
-55.7 |
|
|
B + C 4 CH3CH2COO- + 12H2 |
-102.4 |
|
|
B + D 4 CH3CH2COO- + 3 S042" |
-151.3 |
Source: Boone, et al., (1980)
Table 4-3 Energy-Yielding Reactions of Methanogens
|
Reaction |
-G°, (kJ/mol substrate) | |
|
1. |
CO2 + 4 H2 ® CH4 + 2H2O |
-130.7 |
|
|
HCO3- + 4 H2 + H+ ® CH4 + 3 H2O |
-135.5 |
|
2. |
CH3COO-+ H+ ® H4 + CO2 |
-37.0 |
|
|
CH3COO-+ H2O ® CH4 + HCO3- |
-32.3 |
|
3. |
HCOO- + H+ ® 0.25 CH4 + 0.75 CO2 + 0.5 H2O |
-36.1 |
|
4. |
CO + 0.5 H2O ® 0.25 CH4 + 0.75 CO2 |
-52.7 |
|
5. |
CH3OH ® 0.75 CH4 + 0.25 CO2 + 0.5 H2O |
-79.9 |
|
6. |
CH3NH3+ + 0.5 H2O ® 0.75 CH4 + 0.25 CO2 + NH4+ |
-57.4 |
|
7. |
(CH3)2NH2+ + H2O ® 1.5 CH4 + 0.5 CO2 + NH4+ |
-112.2 |
|
8. |
(CH3)2NCH2CH3H+ + H2O ® 1.5 CH4 + 0.5 CO2 ++H3NCH2CH3 |
-105.0 |
|
9. |
(CH3)3NH+ 1.5H2O ® 2.25 CH4 + 0.75 CO2 + NH4+ |
-170.8 |
Source: Thauer, et al., (1977)
Since methanogens, as obligate anaerobes, require a redox potential of less than -300 mV for growth, their isolation and cultivation was somewhat elusive due to technical difficulties encountered in handling them under completely O2-free conditions. However, as a result of a greatly improved methanogen isolation techniques developed by Hungate (6), more than 40 strains of pure methanogens have now been isolated. Methanogens can be divided into two groups: H2/CO2- and acetate-consumers. Although some of the H2/CO2-consumers are capable of utilizing formate, acetate is consumed by a limited number of strains, such as Methanosarcina spp. and Methanothrix spp. (now, Methanosaeta), which are incapable of using formate. Since a large quantity of acetate is produced in the natural environment (Fig. 4-1), Methanosarcina and Methanothrix play an important role in completion of anaerobic digestion and in accumulating H2, which inhibits acetogens and methanogens. H2-consuming methanogens are also important in maintaining low levels of atmospheric H2.
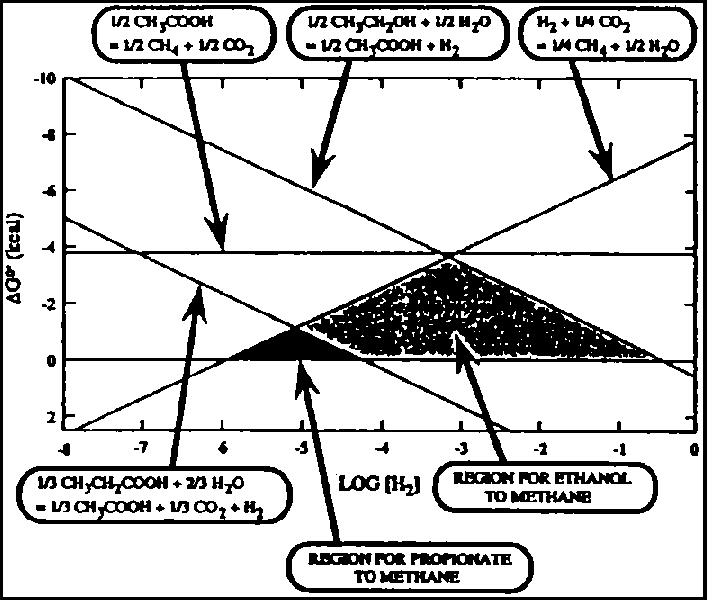
H2/CO2-consuming methanogens reduce CO2 as an electron acceptor via the formyl, methenyl, and methyl levels through association with unusual coenzymes, to finally produce CH4 (7) (Fig. 4-3). The overall acetoclastic reaction can be expressed as:
|
* |
o |
|
* |
o |
|
CH3COOH |
® |
CH4 + CO2 | ||
Since a small part of the CO2 is also formed from carbon derived from the methyl group, it is suspected that the reduced potential produced from the methyl group may reduce CO2 to CH4 (8).
4.2.1 Genetic markers
4.2.2 Molecular cloning of methanogenic genes
4.2.3 Genetic transformations
On the basis of homologous sequence analysis of 16S rRNAs, methanogens have been classified into one of the three primary kingdoms of living organisms: the Archaea (Archaebacteria). The Archaea also include major groups of organisms such as thermophiles and halophiles. Although Archaea possess a prokaryotic cell structure and organization, they share common feature with eukaryotes: homologous sequences in rRNA and tRNA, the presence of inn-ones in their genomes, similar RNA polymerase subunit organization, immunological homologies, and translation systems.
Recombinant DNA technology is one of the most powerful techniques for characterizing the biochemical and genetic regulation of methanogenesis. This necessitates the selection of genetic markers, an efficient genetic transformation system, and a vector system for genetic recombination as prerequisites.
Genetically marked strains are prerequisites for genetic studies: these strains can be employed to develop a genetic-exchange system in methanogens based on an efficient selection system. Since growth of M. thermoautotrophicum is inhibited by fluorouracil, analogue-resistant strains were isolated by spontaneous mutation. Other mutants resistant to DL-ethionine or 2-bromoethane sulfonate (coenzyme M analogue), in addition to autotrophic mutants, were obtained by mutagenic treatment. Several autotrophic strains were also obtained for the acetoclastic methanogen, M. voltae. These mutant strains are listed in Table 4-4.
Source: McCarty, (1982)
Although some methanogen genes such as amino acid and purine biosysnthetic genes, transcription and translation machinery genes, and structural protein genes, have been cloned, genes encoding enzymes involved in methanogenesis were chosen as "methane genes" here.
Figure 4.3 - C, Cycle for reduction of CO2 to CH4
Methyl CoM reductase (MR; Fig. 4-3) constitutes approximately 10% of the total protein in methanogenic cultures. The importance and abundance of MR inevitably focused initial attention on elucidating its structure and the mechanisms directing its synthesis and regulation. MR- encoding genes have been cloned and sequenced from Methanococcus vanielli, M. voltae, Methanosarcina barkeri, Methanobacterium thermoautotrophicum and M. fervidus.
Formylmethanofuran transferase (FTR) catalyzes the transfer of a formyl group from formylmethanofuran (MFR) to tetrahydromethanopterin (H4MPT) (Fig. 4-3, 4-2). The FTR-encoding gene from M. thermoautotrophicum has been cloned, sequenced, and functionally expressed in E. coli. Formate dehydrogenase (FDH) may sometimes account for 2 to 3% of the total soluble proteins in methanogenic cultures. The two genes encoding the a± and a² subunits of FDH have been cloned and sequenced from M formicicum. In addition, the genes encoding F420-reducing hydrogenase (Fig. 4-3), ferredoxin, and ATPase have also been cloned.
Table 4-4 Auxotrophic and Drug-Resistant Mutants Applicable To Gene Transfer Experiments
|
Strain |
Growth Requirement or Inhibitor |
Resistance Factor |
|
Methanococcus voltae PS
|
L-Histidine |
|
|
Purines |
||
|
Cyanocobalamin |
||
|
8-Aza-2,6-diaminopurine |
1,000 |
|
|
8-Azaguanine |
10,000 |
|
|
8-Azahypoxanthine |
>10,000 |
|
|
6-Mercaptopurine |
>500 |
|
|
6-Azauracil |
2,000 |
|
|
Pseudomonic acid |
40 |
|
|
1,2,4-Triazole-3-alanine |
800 |
|
|
Methanobacterium thermoautotrophicum Marburg
|
L-Leucine |
|
|
L-Tryptophan |
||
|
Adenosine |
||
|
Thiamine |
||
|
Formate |
||
|
Pseudomonic acid |
50 |
|
|
5-Fluorouracil |
>1,000 |
|
|
6-Mercaptopurine |
ND |
|
|
8-Aza-2,6-diaminopurine |
ND |
Source: Leisinger, et al., (1993)
A considerable amount of information relevant to natural DNA transformations of prokaryotic bacteria has been reported, and the natural competence of methanogens has been elucidated. M. thermoautotrophicum was transformed by DNA from fluorouacil resistant strains, resulting in the production of drug-resistant strains. In M voltae, auxotrophic mutants requiring histidine or purine were reverted with wild-type DNA, although the genetic transformation frequencies were very low. However, Gernhardt et al. (10) recently made a breakthrough with integration of a vector into M. voltae. Integration vector transformation techniques have been well exploited in yeasts, but not in methanogens. The hisA gene cloned from the methanogen was used as an integration site in homologous recombination. In methanococcus, a puromycin-resistant gene from Streptomyces alboniger was clearly shown to be expressed and stably maintained only under specific pressure conditions. Further characterization of the integration mode revealed that the integration vector was tandemly repeated in chromosomal genes of M maripaludis under intensive antibiotic pressure (11). Furthermore, genomic DNA from the recipient methanogen could directly transform E. coli to ampicillin resistance, indicating that integrated plasmid vectors can be used as recoverable shuttle vectors between methanogens and E. coli.
Electroporation has also been employed for transformation with wild-type DNA in several auxotrophic mutants (CoM, histidine, and purine) with 50- to 80-fold higher efficiency than natural transformation (12). Thus, within a period of only a few years, it has become possible to exchange genomic information in methanogens, or at least in methanococci, with several genetic markers, a recoverable integration vector, and improved transformation efficiency.
4.3.1 Upflow anaerobic sludge blanket (UASB)
4.3.2 Upflow anaerobic filter process (UAFP)
4.3.3 Anaerobic fluidized-bed reactor (AFBR)
4.3.4 Two-phase methane fermentation processes
Methane fermentation has been used since 1900 for treating excess sludge discharged from sewage-treatment plants. This technology has since been developed to treat waste waters, such as those derived from alcohol distillation, antibiotic production, and baker's yeast manufacture. However, the conventional system requires a long hydraulic retention time (HRT) due to factors such as low microbial concentration, and instability against environmental shocks.
Research and development efforts have been directed at retaining a high density of useful microorganisms, in order to achieve rapid and effective treatment, with the objective of improving the conventional system. To this end, considerable technological developments in microbial floe formation and in microbial adhesion onto carrier materials which retain cells in the reactor have been made. For the former purpose, the upflow anaerobic sludge blanket (UASB) (13) has proven useful, while for the latter, the upflow anaerobic filter process (UAFP) (14) and anaerobic fluidized-bed reactor (AFBR) (15) have been developed. In all of these newly developed processes, however, acidogenesis may occur more frequently than methanogenesis, leading to the accumulation of inhibitory products such as volatile fatty acids. Two-phase anaerobic digestion processes have been developed in order to resolve this problem (16).
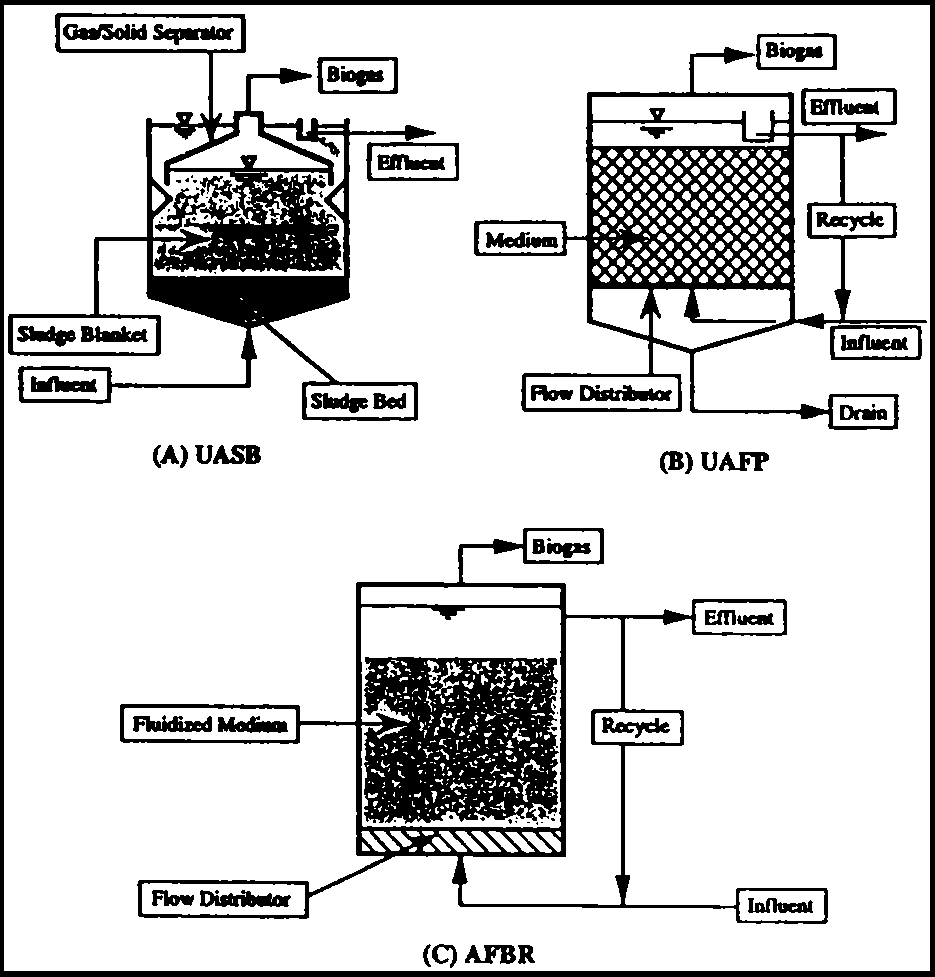
Successful construction of a UASB process capable of affording self-granulation (flocculation) of anaerobic microbes was first reported by Letting et al. (13). As shown in Fig. 4-4 (A), waste water entering from the bottom of the reactor passes through a sludge bed and sludge blanket where organic materials are anaerobically decomposed. Gas produced is then separated by a gas-solid separator and the clarified liquid is discharged over a weir, while the granular sludge naturally settles to the bottom. Granules range in size from 0.5-2.5 mm, and in concentration from 50-100 kg VSS/m3 at the bottom, to 5-40 kg VSS/m3 in the upper part of the reactor. Bench- and pilot plant-scale experiments indicate that it is possible to operate this system at a COD loading of 40 kg/m3/day at HRTs of 4-24 hours. Full-scale UASB reactors are now operational in Europe, the US and Japan, with more than 100 recently constructed plants in Japan. As shown in Table 4-5 (17), the COD concentration in the inlet waste water ranges from 2,000-20,000 mg/l at HRTs of 0.1-2 days (depending on the inlet COD concentration) in which the COD loads are 6-14 kg/m3/day, resulting in a COD reduction of more than 80%. UASB systems are primarily used in the treatment of waste water derived from the food processing industry. Studies on the application of this system to domestic sewage, however indicate that the UASB reactor can operate at a rather long HRT of 22 hours with a relatively low COD reduction of 60-80%.
Significant parameters in the UASB operation are floe diameter, microbial density, and the structure of the gas-solid separator which effectively retains the microbial granules within the reactor. The following criteria should be observed in order to achieve successful UASB operation: (a) selection of a suitable waste water capable of granule self-formation; (b) operation of the reactor without mechanical agitation; (c) start up at a relatively low COD load; (d) use of waste water containing Ca2+ and Ba2+ and (e) avoidance of bulking caused by filamentous microbial growth. Granule formation in a UASB system is influenced by the growth of rod-type Methanothrix spp. which produce spherical granules.
Table 4-5 Operational Methane-Generating Bioreactors and Their Current Applications
|
Reactor Type |
Location |
Waste Characteristics |
Design |
||||||
|
Type |
COD (mg/l) |
Flow (m3/day) |
HRT (day) |
COD load (kg/m3/day) |
COD removal (%) |
Medium |
|||
|
UASB
|
USA |
|
|
|
|
|
|
|
|
|
|
La Crosse, Wis. |
Brewery |
2,500 |
23,000 |
0.2 |
14.1 |
86 |
|
|
|
|
Caribou, Me. |
Starch |
22,000 |
910 |
2.0 |
11.0 |
85 |
|
|
|
|
Plover, Wis. |
Potato |
4,300 |
3,000 |
0.7 |
6.0 |
80 |
|
|
|
Europe |
|
|
|
|
|
|
|
||
|
|
Netherlands |
Sugar |
17,000 |
3,000 |
1.0 |
13.3 |
94 |
|
|
|
|
Netherlands |
Sugar beet |
3,000 |
6,960 |
0.2 |
14.5 |
85 |
|
|
|
|
Netherlands |
Starch |
7,700 |
1,750 |
0.1 |
8.0 |
85 |
|
|
|
|
Switzerland |
Potato |
2,260 |
2,210 |
0.3 |
8.3 |
80 |
|
|
|
|
Germany |
Sugar beet |
7,500 |
2,400 |
0.6 |
12,0 |
86 |
|
|
|
|
Netherlands |
Alcohol |
5,330 |
2,090 |
0.3 |
16.0 |
90 |
|
|
|
UAFP
|
Spokane Washington |
Starch gluten |
8,800 |
490 |
0.9 |
3.8 |
64 |
Graded rock |
|
|
Vernon, Texas |
Guar gum |
9,140 |
823 |
1.0 |
16.0 |
60 |
9 cm Pall rings |
||
|
San Juan Puerto Rico |
Rum distillery |
95,000 |
1,325 |
7.0-8.0 |
8.9 |
75 |
Synthetic ("Vinyl core") |
||
|
Bishop, Texas |
Chemical |
12,000 |
3,785 |
1.5 |
9.6 |
80 |
9 cm Pall rings |
||
|
Pampa, Texas |
Chemical |
14,400 |
3,785 |
1.5 |
10.4 |
90 |
9 cm Pall rings |
||
|
AFBR
|
Birmingham, Ala. |
Soft-drink bottling waste |
6,900 |
380 |
0.3 |
9.6 |
77 |
0.6 mm E.S. sand |
|
|
Midwest |
Soy-processing waste |
9,000 |
- |
<1.0 |
13.0 |
- |
0.4 mm E.S. sand |
||
Source: Bowker, 1983
UAFP systems were initially developed by Young and McCarty (14) using rocks and plastics for microbial fixation. These UAFP systems were applied to the treatment of domestic sewage and industrial waste waters containing relatively low levels of organic materials. As shown in Fig. 4-4 (B), the reactor contains a "medium", i.e. a microbial support. Granulated microorganisms exist not only in the spaces within the medium, but are also attached to its surface; hence, a high-density microbial population is retained within the reactor, creating a hybridization of microbial floe and adhesion. In order to avoid short-circuiting flow through the packed column, a distributor is fitted at the bottom to provide a homogeneous up-flow of waste water. At the top, treated waste water and the biogas produced are separated by a free board.
Data on full-scale UAFP systems (Table 4-5) show that alcohol distillery waste water can be treated at an HRT of 7.8 days with 74% COD removal. Application of this UAFP to domestic sewage treatment using Raschig rings (2.5 cm) as microbial supports, resulted in BOD removal of 50 to 60 % and suspended solids (SS) removal of 70 to 80%, at an HRT ranging from 5 to 33 hours.
Selection of a medium in which microbial adhesion is greatly influenced both by SS, and the chemical composition of the waste water, is extremely critical in UAFP systems. The effects of physical medium characteristics, such as size and shape, on COD removal have been investigated using modular corrugated blocks (porosity > 95%), pall rings, and perforated spheres. At a COD load of 2 kg/m/day, modular corrugated blocks exhibited superior behavior, removing 88 % of COD. A comparison of COD removal for cross- and tubular-flow systems, reveals that COD removal is 20 to 30 % greater in cross-flow systems.
In addition to plastic media, baked clay and a melted slug have also proven useful in laboratory experiments on methanogenesis from formate, acetate, and methanol. Pumice was used as a microbial supporter for methanogenesis from methanol-rich waste water of the evaporate condensate from a pulp mill (COD load: 12 kg/m3/day, COD removal: 96%).
A diagram of an anaerobic fluidized-bed process is shown in Fig. 4-4 (C). In such systems, the medium to which the microbes adhere is fluidized within the reactor, resulting in conversion of organic materials to CH4 and CO2. Anaerobic microbes grow on the surface of the medium, expanding the apparent volume of the medium; hence this reactor is also designated an "expanded bed reactor". Use of artificial sewage in an AFBR, resulted in COD removal exceeding 80 % at 20°C, and at a COD load of 2-4 kg/m3/day this system was tolerant of shock loading for step changes of temperature from 13 to 35°C and from 35 to 13°C. In the case of COD shock loading from 1.3 to 24 kg/m3/day, a steady state is established after 6 days. The AFBR thus seems to be capable of performing at relatively low temperatures with both low and high COD waste waters, without significant shock loading effects.
The AFBR has been progressively developed, as shown by the full-scale operation data in Table 4-5. Engineering improvements which can potentially minimize the mechanical power required for fluidization include reduction of the expanded volume, selection of a low density medium of high specific area; and avoidance of fragility. Media such as sand, quartzite, alumina, anthracite, granular activated carbon, or crystobalite with a particle size of approximately 0.5 mm are usually employed.
Novel bioreactors for methane fermentation such as the UASB, UAFP, and AFBR experience inherent problems when operated at high COD loads, due to the fact that the overall growth rate of acidogenic bacteria proceeds faster (10-fold) than that of methanogenic bacteria. When this occurs, inhibitory products such as volatile fatty acids and H2 accumulate in the reactor, slowing down the entire process. In order to overcome this, two-phase processes consisting of acidogenic and methanogenic fermentation's have been investigated (16).
In one full-scale two-phase system + the Anodek process (Belgium) + 70-97% COD removal and biogas production of 3-13 Kg/m2 day with a methane content of 65 to 80% was obtained when operated at COD loads of 20-60 kg/m3/day for acidogenic fermentation (1st phase) and 6-30 kg/n^/day for methanogenic fermentation (2nd phase). In another example, a two-phase system consisting of a complete stirred reactor for the first phase and a UASB for the second phase was constructed. When this system was applied in the treatment of alcohol distillery waste (COD =10,000 mg//) at HRTs of 16-72 hours in the first phase, and 14 hours in the second phase, 84% COD removal and 92% BOD removal were accomplished. A two-phase system consisting of a UAFP for the first phase and a horizontal AFP for the second phase has also been proposed, with which it should be possible to treat sewage waste water (COD 800 to 2,600 mg/l) at HRTs of 2-5.5 hours with a high methane content (~90%).
In addition, since SS in waste water greatly influences the performance of the UASB or UAFP, an acidogenic fermentation first phase in combination with a UASB or UAFP second phase is useful in reducing the SS which enter the second phase.
|
1. |
McCarty, P.L., In "Anaerobic Digestion 1981" Eds. Hughes, D.E. et.al., 3-22 (1982) Elsevier Biomedical Press, Amsterdam, New York, Oxford. |
|
|
|
|
2. |
Zeikus, J.G., Ann. Rev. Microbiol, 34, 423-464 (1980). |
|
|
|
|
3. |
Boone, D.R. et al., Appl. Environ. Microbiol., 40, 626-632 (1980). |
|
|
|
|
4. |
McInerney, M.J. et al., Appl. Environ. Microbiol., 41, 1029-1039 (1981). |
|
|
|
|
5. |
Thauer, R.K. et al., Bact. Rev., 41, 100-180 (1977). |
|
|
|
|
6. |
Hungate, R.E., Methods in Microbiol., 3B, 117-132 (1969). |
|
|
|
|
7. |
Wolfe, R.S., Trends in Biochem. Sci., 10, 396-399 (1985). |
|
|
|
|
8. |
Thauer, R.K. et.al., Ann. Rev. Microbiol., 43, 43-67 (1989). |
|
|
|
|
9. |
Leisinger, T. et.al., In "Genetics and Molecular Biology of Anaerobic Bacteria" Ed. Sebald, M., 1-12 (1993) Springer-Verlag, New York, Berlin, Heidelberg, London, Paris, Tokyo, Hong Kong, Barcelona, Budapest. |
|
|
|
|
10. |
Gernhardt, P. et.al., Mol. Gen. Genet., 221, 273-279 (1990). |
|
|
|
|
11. |
Sandbeck, K.A. et.al., Appl. Environ. Microbiol., 57, 2762-2763 (1991). |
|
|
|
|
12. |
Micheletti, P.A. et.al., J. Bacteriol, 173, 3414-3418 (1991). |
|
|
|
|
13. |
Lettinga, G. et.al., Biotech. Bioeng., 22, 699-734 (1980). |
|
|
|
|
14. |
Young, J.C. et.al., J. Wat. Poll. Contr. Fed., 41, 160-173 (1969). |
|
|
|
|
15. |
Jeris, J.S., Wat. Sci. Technol., 15, 167-187 (1983). |
|
|
|
|
16. |
Pohland, F.G. et.al., Environ. Lett., 1, 255-266 (1971). |
|
|
|
|
17. |
Bowker, R.P.G., Environ. Progress, 2, 235-242 (1983). |
Reviews recommended
|
1. |
Nagai, S. and Nishio, N., In "Handbook of Heat and Mass Transfer, vol. 3 Catalysis, Kinetics and Reactor Engineering" Ed. Cheremisinoff, N.P., 701-752 (1986) Gulf Publishing Corn., Houston, London, Paris, Tokyo. |
|
|
|
|
2. |
Kida, K. et al., In "Handbook of Heat and Mass Transfer, vol.3 Catalysis, Kinetics and Reactor Engineering" Ed. Cheremisinoff, N.P., 773-787 (1986) Gulf Publishing Corn., Houston, London, Paris, Tokyo. |
|
|
|
|
3. |
Palmer, J.R. and Reeve, J.N., In "Genetics and Molecular Biology of Anaerobic Bacteria" Ed. Sebald, M., 13-35 (1993) Springer-Verlag, New York, Berlin, Heidelberg, London, Paris, Tokyo, Hong Kong, Barcelona, Budapest. |
![]()
![]()
![]()
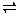 2 CH3COO-
+ 2H2 + H+
2 CH3COO-
+ 2H2 + H+
{kind=link}
{kind=link}
{kind=link}
{kind=link}